Sperm Morphology and Male Age in Black-Throated Blue Warblers, an Ecological Model System

1
Smithsonian Conservation Biology Institute, National Zoological Park, Washington, DC 20008, USA
2
Cornell Lab of Ornithology, Cornell University, Ithaca, NY 14850, USA
3
Premedical Education Department, Weill Cornell Medicine-Qatar, Doha 24144, Qatar
4
School of Biological Sciences, Washington State University, Pullman, WA 99164, USA
*
Author to whom correspondence should be addressed.
Animals 2020, 10(7), 1175; https://doi.org/10.3390/ani10071175
Submission received: 16 June 2020
/
Revised: 3 July 2020
/
Accepted: 6 July 2020
/
Published: 10 July 2020
(This article belongs to the Special Issue Reproductive Biotechnology in Wildlife)
Abstract
:Simple Summary
Sperm cell characteristics can impact the number of offspring a male sires, particularly when females copulate with more than one male in a reproductive cycle. Females copulate with multiple males in many songbird species, but for many of these species, little is known about sperm characteristics. In this paper, we examine the shape of sperm cells in black-throated blue warblers, a species where multiple copulations are common but where sperm morphology was previously unknown. Sperm shape did not correlate with plumage characteristics that may make males more successful at gaining copulation partners. However, older males tended to have longer sperm cells. Previous work shows that older males sire more offspring, and work in other species indicates that longer sperm cells are more effective at fertilizing eggs. Thus, sperm shape may help explain the higher success of older males in this species.
Abstract
Extra-pair paternity may drive selection on spermatozoa and ejaculate characteristics through sperm competition and cryptic female choice. Here, we examine sperm morphology in the black-throated blue warbler (Setophaga caerulescens), an ecological model species where extra-pair paternity is frequent and is linked with male age. We test whether sperm morphology relates to several aspects of male phenotype known or suspected to affect extra-pair paternity success. Sperm morphology did not correlate with the size of the white wing spot, a social status signal, nor with the volume of the cloacal protuberance. However, older males tended to have longer sperm cells. Although the sample size was limited, this pattern is intriguing, as longer cells may be advantageous in post-copulatory sexual selection and older males have larger testes and higher extra-pair paternity success in this species. Changes in sperm morphology with age are not observed in other birds, though they have been observed in insects and fishes. More research on sperm morphology is needed to clarify its role in extra-pair fertilizations in this well-studied species.
1. Introduction
Sperm cells evolve in response to diverse selection pressures [1]. Among these pressures are sperm competition and cryptic female choice [2,3], which can act when a female copulates with more than one male in a reproductive cycle, for instance in polyandrous mating systems or socially monogamous systems with extra-pair paternity (i.e., paternity by males other than the female’s social partner). Consistent with increased selection due to sperm competition and cryptic female choice, passerine bird species with higher levels of extra-pair paternity exhibit reduced among-male variation in sperm total length [4,5,6], longer spermatozoa [7,8], and faster change in sperm length over evolutionary time [9]. Sperm competition experiments with captive zebra finches (Taeniopygia guttata) show that males with longer and faster-swimming spermatozoa have higher fertilization success [10]. Despite the potential importance of sperm and ejaculate traits in extra-pair paternity, few studies have investigated sperm biology within passerine species.
Black-throated blue warblers (Setophaga carulescens) are socially monogamous, open-cup nesting songbirds with substantial extra-pair paternity (approximately 43% of offspring in 55% of broods), typically occurring among neighboring territories [11,12]. Males with lower food abundance on their territories are more likely to sire extra-pair paternity, and to sire offspring at a greater distance from their home territories, suggesting a possible role of foraging forays in extra-pair mating [11,13]. Older males are also more likely to sire extra-pair offspring [11]. Males and females differ substantially in plumage, with females being drab olive colored and males having a blue back, black face and flanks, and white underside [14]. Both sexes have a white wing patch; it is larger in males [14] and affects male-male interactions, particularly for young males [15]. Being sexually dimorphic, dorsal coloration and wing patch size may be hypothesized to affect extra-pair copulations, although one study found no relationship between wing patch and parentage [16]. Despite being the focus of intense study over many years [17], and particularly in the context of extra-pair paternity [11,12,13,18,19,20], sperm morphology was previously undescribed in this species.
Here, we examine the morphology of spermatozoa in black-throated blue warblers and assess its relationship to male traits known or hypothesized to affect extra-pair paternity: Male age, plumage, and cloacal protuberance volume (which may be used as a proxy for sperm production [4]). We focus on total sperm length, within-male variation in total sperm length, and the relative flagellum length, since sperm with longer flagella relative to head may swim more rapidly and thus have higher chances of fertilizing the eggs [7,21,22]; but see [8,23]. Because the direction and strength of correlation between male and ejaculate traits varies substantially in other taxa [24,25,26], no specific predictions were made. We show that older males tended to have longer sperm cells, an intriguing pattern given the importance of male age in extra-pair paternity in this species.
2. Materials and Methods
2.1. Animals and Sampling Procedures
Wild, free-living males were captured by mistnetting, from 22 May 2016 to 28 June 2016 at the Hubbard Brook Experimental Forest, Woodstock, New Hampshire, USA (43°56′ N, 71°45′ W). Birds were banded, bled, and measured for inclusion in on-going long-term studies. When weather and bird condition permitted slightly longer safe handling time, we collected a single semen sample per male by cloacal massage (n = 13 males). Semen samples were mixed with approximately 20 μL phosphate-buffered saline, and we transferred the sample into 5% formaldehyde immediately. We measured (1) the size of the white wing patch using digital calipers to the nearest 0.1 mm, placed around the longest extent of white visible while the wing was closed (n = 12; [15]), (2) the percentage of black feathers in the male’s backs, which was estimated while the bander held the bird in a modified bander’s grip to expose the back (n = 11 males [27]); (3) and the length and diameter of the cloacal protuberance (n = 10 males). Cloacal protuberance volume was calculated as length times radius squared times pi. Age was assigned as second-year (SY; first breeding season) if the alula had green edging or after second year (ASY) if the alula was pure blue [27]. Only four males had nests that survived until genetic sampling of offspring, so we did not attempt to examine genetic parentage. Male measurements are in Table S1.
2.2. Sperm Morphology Measurements
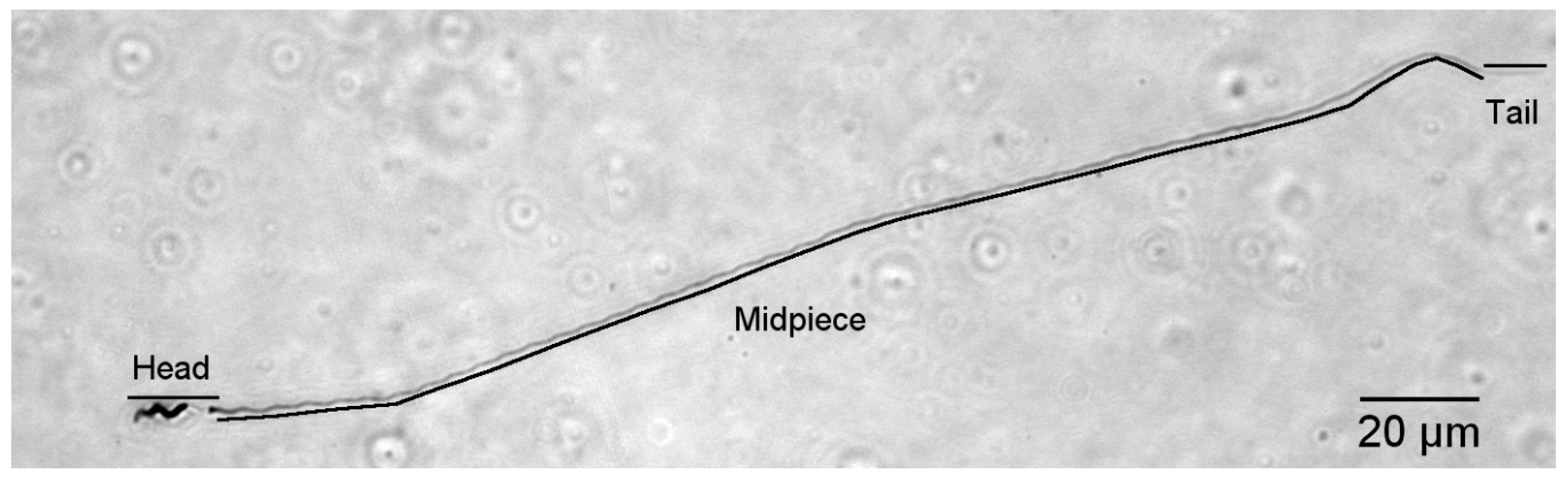
For each male, approximately 10 μL fixed semen sample was streaked onto a microscope slide, where it was allowed to air-dry before being immersed in Coomassie Blue stain for 1.5 min. Excess stain was rinsed off, a cover slip was applied using Permount, and slides were examined at 400× total magnification with an Olympus BX 41 microscope (Olympus, NewYork, NY, USA; the highest magnification allowing entire sperm cells in one field of view). Twenty morphologically normal sperm cells were photographed (Spot Imaging software, Sterling Heights, MI, USA), and images were analyzed using Image J software (National Institutes of Health, Bethesda, MD, USA) [28]. Cells were considered morphologically normal (e.g., Figure 1) if they exhibited a generally helical superstructure from the acrosome to the end of the midpiece and if there was no evidence of tail breakage. One SY male was excluded because of apparent damage to sperm cells, with the mitochondrial helix often appearing unwound. We measured the length of the head (including acrosome), midpiece, and tail (exposed flagellum; Figure 1), then calculated total sperm length as the sum of these components [8]. Raw data are in Table S2.
Flagellum length was the sum of midpiece and exposed flagellum. The coefficient of variation in sperm length within males (CVwm) was calculated as standard deviation in total length, divided by mean total length [29]. Because relatively few males were sampled, we applied a correction factor to avoid bias due to small sample sizes in calculating the coefficient of variation among males (CVam = (standard deviation/mean) × (1 + 1/(4n)) [4,29].
All measurements were taken by one person and were blind with respect to male phenotype. Measurements were repeatable (repeatability ± SE as assessed using package rptR [30] on four haphazardly chosen sperm cells from each of five males, with all 20 cells measured five times on different days; head 0.59 ± 0.10; midpiece 0.94 ± 0.02; tail 0.80 ± 0.06; total sperm length 0.99 ± 0.01; all p < 0.05 by likelihood ratio test; data in Table S3).
2.3. Statistical Analysis
To test whether males differed in sperm morphology, we performed ANOVA using data from individual sperm cells. To examine relationships among sperm component lengths, and between sperm measures and male phenotypes, we used the average length of each sperm component or measurement for each male. To test for correlations among different sperm components lengths, we performed linear regression among all pairs of sperm components (head, midpiece, and tail). To test whether sperm measurements (total sperm length, CVwm, and the relative length of the flagellum) related to sexual phenotypes (male age, the size of the wing spot, the percent of black feathers on the back, and the volume of the cloacal protuberance), we again used linear regression. For tests on relative flagellum length, we used a linear regression with flagellum length as the response variable, head length as a covariate, and one male phenotypic trait per model as the predictor of interest. For all other sperm measurements, we included only the male phenotypic trait as a predictor. Results were similar using data on individual sperm cells as the response variables, in mixed models controlling for male identity as a random effect (not shown; reporting only mean-level models was preferred for simplicity, since the single-cell approach is not available for CVwm). We further tested whether total sperm length changed with date of capture (e.g., [31] using linear regression on mean total sperm length).
To correct for multiple testing, we used false discovery rate correction, performing separate corrections for (1) comparisons of mean lengths across males; (2) assessing relationships among sperm component lengths; and (3) assessing relationships between sperm measurements and male phenotype. Uncorrected p-values are shown except where noted otherwise. All analyses were conducted in R version.3.6.1 (R Development Core Team 2019, Vienna, Austria [32]). Code used in analysis are in File S1.
2.4. Ethics Statement
This research was conducted with approval from the National Zoological Park Animal Care and Use Committee (14–20). The federal banding permit numbers is 23885.
3. Results
Total sperm length (mean ± SD) was 247.66 ± 5.11 μm and CVam was 2.1% (Table 1). Among-male variation was statistically significant for head, midpiece, and total sperm length, and it approached significance for tail (Table 1; p-values not qualitatively changed by FDR correction).
There were no significant correlations among sperm component lengths (head vs. midpiece, F1,10 = 1.23, p = 0.29; head vs. tail, F1,10 = 0.144, p = 0.71, midpiece vs. tail F1,10 = 2.52, p = 0.14). Sperm measurements did not correlate with capture date (total sperm length F1,10 = 2.99, p = 0.11; flagellum length controlling for head F1,9 = 2.81, p = 0.13; CVwm F1,10 = 0.17, p = 0.69). Older males tended to have longer sperm than young males (F1,10 = 11.74, uncorrected p = 0.006, corrected p = 0.08, estimated difference 8.3 ± 2.4 μm; Figure 2), with an accompanying trend for older males to have a longer flagellum relative to head length (F1,9 = 9.10, uncorrected p = 0.01, corrected p = 0.09). No other relationships between sperm traits and male phenotypes approached significance (Table 2).
4. Discussion
Older male black-throated blue warblers tended to have longer sperm, a pattern which has been observed in rove beetles (Aleochara bilineata [33]) and guppies (Poecilia reticulata [34]). Longer sperm are generally thought to have a competitive advantage over shorter sperm [10,35]. Increasing male age also correlates with greater total testicular volume and more asymmetrical testes in black-throated blue warblers [19], and with larger testes in other bird species ([36] and references therein). Larger testes are expected to allow older males to produce more sperm and therefore increase their chances of success in post-copulatory sexual selection [37]. Indeed, older males are more likely to sire extra-pair offspring in black-throated blue warblers [11] and in many other birds [38]. Recent experimental work in the house sparrow (Passer domesticus) suggests that the extra-pair success of older males may be due more to post-copulatory than pre-copulatory processes [39]. While the difference in total sperm length between age groups should be treated as preliminary due to our low sample size, it seems possible that age-dependent changes in reproductive traits such as sperm size and testes size and asymmetry play a role in differential siring success in this system.
In the other passerine birds that have been tested, sperm morphology does not differ between young and old males [40,41,42,43,44]. Furthermore, repeated samples across years of the same individuals show high repeatability in total sperm length, suggesting that between-season changes in sperm morphology within an individual are limited [23,31,42,45,46]. Small within-season changes in morphology, in contrast, appear to be relatively common, occurring in most of the species studied [31,42,43,46] (but see [45]). Older male black-throated blue warblers are more likely to obtain and defend territories with high quality nesting and foraging habitat [47], suggesting that, if they therefore have higher body condition (not assessed in this study), body condition could mediate the greater sperm length observed in older birds. Further study on black-throated blue warblers, with larger sample sizes, is warranted.
The degree of among-male variation in sperm total length is somewhat higher than expectations based on the extrapair paternity rate of the species (measured CVam 2.1; value of CVam associated with 43% extra-pair young, 1.43, using the calculator provided in [48]). Age-related changes in sperm total length may be partly responsible for this higher-than-expected between-male variation (though note that there is substantial unexplained variation in the relationship, such that our observed value is not extreme [4,48]). Average total sperm length for black-throated blue warblers is similar to other wood-warblers (average for black-throated blue warblers, 247.64 ± 5.11 μm; average for other wood-warblers in [4], 222.78 ± 41.08 μm; average for oscine passerines in [4], 147.82 ± 71.9 μm).
5. Conclusions
Extra-pair paternity is intricately linked with male age, ecology, and time-budget trade-offs in black-throated blue warblers [11,18]. Here we show that age may also affect sperm morphology. Further work is therefore needed to fully how these factors also interact with sperm and ejaculate traits in driving sexual selection in this ecological model system.
Supplementary Materials
The following are available online at https://www.mdpi.com/2076-2615/10/7/1175/s1, Table S1: Raw data on male characteristics, Table S2: Raw data on sperm morphology used in main analyses, Table S3: Data on sperm morphology used for assessing repeatability, File S1: R code used in analysis.
Author Contributions
Conceptualization, E.C.; sample collection, E.C. and N.K.; microscopy, E.C., T.R., P.C.; formal analysis, E.C.; resources, P.C.; writing—original draft preparation, E.C.; writing—review and editing, E.C., N.K., T.R., and P.C. All authors have read and agreed to the published version of the manuscript.
Funding
Long-term research on this population of birds has been supported by the US National Science Foundation.
Acknowledgments
We thank all of the field technicians that helped collect these data, and Sara Kaiser, Scott Sillett, Mike Webster, Nick Rodenhouse, and Dick Holmes for supporting the long-term study of black-throated blue warblers at Hubbard Brook. Special thanks to Danielle Aube and Thomas Aaron Hulsey for bird capture, measurements, and nest finding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lüpold, S.; Pitnick, S. Sperm form and function: What do we know about the role of sexual selection? Reproduction 2018, 155, R229–R243. [Google Scholar] [CrossRef] [PubMed]
- Parker, G. Sperm competition and its evolutionary consequences in the insects. Biol. Rev. 1970, 45, 525–567. [Google Scholar] [CrossRef]
- Eberhard, W. Female Control: Sexual Selection by Cryptic Female Choice; Princeton University Press: Princeton, NJ, USA, 1996. [Google Scholar]
- Lifjeld, J.T.; Laskemoen, T.; Kleven, O.; Albrecht, T.; Robertson, R.J. Sperm length variation as a predictor of extrapair paternity in passerine birds. PLoS ONE 2010, 5, e13456. [Google Scholar] [CrossRef] [Green Version]
- Calhim, S.; Immler, S.; Birkhead, T.R. Postcopulatory sexual selection is associated with reduced variation in sperm morphology. PLoS ONE 2007, 2, e413. [Google Scholar] [CrossRef]
- Kleven, O.; Laskemoen, T.; Fossøy, F.; Robertson, R.J.; Lifjeld, J.T. Intraspecific variation in sperm length is negatively related to sperm competition in passerine birds. Evolution 2008, 62, 494–499. [Google Scholar] [CrossRef] [PubMed]
- Lüpold, S.; Calhim, S.; Immler, S.; Birkhead, T.R. Sperm morphology and sperm velocity in passerine birds. Proc. R. Soc. B 2009, 276, 1175–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleven, O.; Fossøy, F.; Laskemoen, T.; Robertson, R.J.; Rudolfsen, G.; Lifjeld, J.T. Comparative evidence for the evolution of sperm swimming speed by sperm competition and female sperm storage duration in passerine birds. Evolution 2009, 63, 2466–2473. [Google Scholar] [CrossRef]
- Rowe, M.; Albrecht, T.; Cramer, E.R.A.; Johnsen, A.; Laskemoen, T.; Weir, J.T.; Lifjeld, J.T. Postcopulatory sexual selection is associated with accelerated evolution of sperm morphology. Evolution 2015, 69, 1044–1052. [Google Scholar] [CrossRef]
- Bennison, C.; Hemmings, N.; Slate, J.; Birkhead, T. Long sperm fertilize more eggs in a bird. Proc. R. Soc. B 2015, 282, 20141897. [Google Scholar] [CrossRef]
- Kaiser, S.A.; Risk, B.B.; Sillett, T.S.; Webster, M.S. Ecological and social factors constrain spatial and temporal opportunities for mating in a migratory songbird. Am. Nat. 2017, 189, 283–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webster, M.S.; Chuang-Dobbs, H.C.; Holmes, R.T. Microsatellite identification of extrapair sires in a socially monogamous warbler. Behav. Ecol. 2001, 12, 439–446. [Google Scholar] [CrossRef]
- Kaiser, S.A.; Sillett, T.S.; Risk, B.B.; Webster, M.S. Experimental food supplementation reveals habitat-dependent male reproductive investment in a migratory bird. Proc. R. Soc. B 2015, 282, 20142523. [Google Scholar] [CrossRef] [PubMed]
- Holmes, R.T.; Kaiser, S.A.; Rodenhouse, N.L.; Sillett, T.S.; Webster, M.S.; Pyle, P.; Patten, M.A. Black-throated Blue Warbler v. 3.0. In The Birds of North America Online; Cornell Lab of Ornithology: Ithaca, NY, USA, 2017. [Google Scholar]
- Cline, M.H.; Hatt, J.L.; Conroy, M.J.; Cooper, R.J. Experimental evidence for a phenotypic trait as an age-dependent intrasexual social signal between familiar individuals. Anim. Behav. 2016, 111, 319–327. [Google Scholar] [CrossRef]
- Cline, M. Communication, Reproduction, and Survival of a Long-Distance Migratory Sonbird, the Black-Throated Blue Warbler (Setophaga caerulescens); University of Georgia: Athens, GA, USA, 2015. [Google Scholar]
- Holmes, R.T. Avian population and community processes in forest ecosystems: Long-term research in the Hubbard Brook Experimental Forest. For. Ecol. Manag. 2011, 262, 20–32. [Google Scholar] [CrossRef]
- Chuang-Dobbs, H.C.; Webster, M.S.; Holmes, R.T. The effectiveness of mate guarding by male black-throated blue warblers. Behav. Ecol. 2001, 12, 541–546. [Google Scholar] [CrossRef]
- Graves, G.R.; Auk, S.T.; Apr, N. Testicular volume and asymmetry are age-dependent in black-throated blue warblers (Dendroica caerulescens). Auk 2004, 121, 473–485. [Google Scholar] [CrossRef]
- Cramer, E.R.A.; Kaiser, S.A.; Webster, M.S.; Sillett, T.S.; Ryder, T.B. Characterizing selection in black-throated blue warblers using a sexual network approach. J. Evol. Biol. 2017, 30, 2177–2188. [Google Scholar] [CrossRef] [Green Version]
- Humphries, S.; Evans, J.P.; Simmons, L.W. Sperm competition: Linking form to function. BMC Evol. Biol. 2008, 8, 319. [Google Scholar] [CrossRef] [Green Version]
- Helfenstein, F.; Podevin, M.; Richner, H. Sperm morphology, swimming velocity, and longevity in the house sparrow Passer domesticus. Behav. Ecol. Sociobiol. 2010, 64, 557–565. [Google Scholar] [CrossRef] [Green Version]
- Cramer, E.R.A.; Laskemoen, T.; Stensrud, E.; Rowe, M.; Haas, F.; Lifjeld, J.T.; Sætre, G.-P.; Johnsen, A. Morphology-function relationships and repeatability in the sperm of Passer sparrows. J. Morphol. 2015, 276, 370–377. [Google Scholar] [CrossRef]
- Evans, J.P.; Garcia-Gonzalez, F. The total opportunity for sexual selection and the integration of pre- and post-mating episodes of sexual selection in a complex world. J. Evol. Biol. 2016, 29, 2338–2361. [Google Scholar] [CrossRef]
- Lüpold, S.; Tomkins, J.L.; Simmons, L.W.; Fitzpatrick, J.L. Female monopolization mediates the relationship between pre- and postcopulatory sexual traits. Nat. Commun. 2014, 5, 3184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mautz, B.S.; Møller, A.P.; Jennions, M.D. Do male secondary sexual characters signal ejaculate quality? A meta-analysis. Biol. Rev. Camb. Philos. Soc. 2013, 88, 669–682. [Google Scholar] [CrossRef] [PubMed]
- Pyle, P. Identification Guide to North American Birds. Part I. Columbidae to Ploceidae; Slate Creek Press: Bolinas, CA, USA, 1997. [Google Scholar]
- Schneider, C.; Rasband, W.; Eliceiri, K. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Sokal, R.; Rohlf, F. Biometry: The Principles and Practice of Statistics in Biological Research; WH Freeman and Co.: San Francisco, CA, USA, 1995. [Google Scholar]
- Stoffel, M.A.; Nakagawa, S.; Schielzeth, H. rptR: Repeatability estimation and variance decomposition by generalized linear mixed-effects models. Methods Ecol. Evol. 2017, 8, 1639–1644. [Google Scholar] [CrossRef] [Green Version]
- Cramer, E.R.A.; Laskemoen, T.; Kleven, O.; Lifjeld, J.T. Sperm length variation in House Wrens Troglodytes aedon. J. Ornithol. 2013, 154, 129–138. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2019. [Google Scholar]
- Green, K. Age-related variation in mean sperm length, in the rove beetle Aleochara bilineata. J. Insect Physiol. 2003, 49, 993–998. [Google Scholar] [CrossRef]
- Gasparini, C.; Marino, I.A.M.; Boschetto, C.; Pilastro, A. Effect of male age on sperm traits and sperm competition success in the guppy (Poecilia reticulata). J. Evol. Biol. 2010, 23, 124–135. [Google Scholar] [CrossRef]
- Lüpold, S.; Manier, M.K.; Berben, K.S.; Smith, K.J.; Daley, B.D.; Buckley, S.H.; Belote, J.M.; Pitnick, S. How multivariate ejaculate traits determine competitive fertilization success in Drosophila melanogaster. Curr. Biol. 2012, 22, 1667–1672. [Google Scholar] [CrossRef] [Green Version]
- Laskemoen, T.; Fossøy, F.; Rudolfsen, G.; Lifjeld, J.T. Age-related variation in primary sexual characters in a passerine with male age-related fertilization success, the bluethroat Luscinia svecica. J. Avian Biol. 2008, 39, 322–328. [Google Scholar] [CrossRef] [Green Version]
- Calhim, S.; Montgomerie, R. Testis asymmetry in birds: The influences of sexual and natural selection. J. Avian Biol. 2015, 46, 175–185. [Google Scholar] [CrossRef]
- Hsu, Y.-H.; Schroeder, J.; Winney, I.; Burke, T.; Nakagawa, S. Are extra-pair males different from cuckolded males? A case study and a meta-analytic examination. Mol. Ecol. 2015, 24, 1558–1571. [Google Scholar] [CrossRef] [PubMed]
- Girndt, A.; Chng, C.W.T.; Burke, T.; Schroeder, J. Male age is associated with extra-pair paternity, but not with extra-pair mating behaviour. Sci. Rep. 2018, 8, 8378. [Google Scholar] [CrossRef] [PubMed]
- Laskemoen, T.; Kleven, O.; Fossøy, F.; Robertson, R.J.; Rudolfsen, G.; Lifjeld, J.T. Sperm quantity and quality effects on fertilization success in a highly promiscuous passerine, the tree swallow Tachycineta bicolor. Behav. Ecol. Sociobiol. 2010, 64, 1473–1483. [Google Scholar] [CrossRef]
- Sætre, C.L.C.; Johnsen, A.; Stensrud, E.; Cramer, E.R.A. Sperm morphology, sperm motility and paternity success in the bluethroat (Luscinia svecica). PLoS ONE 2018, 13, e0192644. [Google Scholar] [CrossRef] [Green Version]
- Edme, A.; Zobač, P.; Korsten, P.; Albrecht, T.; Schmoll, T.; Krist, M. Moderate heritability and low evolvability of sperm morphology in a species with high risk of sperm competition, the collared flycatcher Ficedula albicollis. J. Evol. Biol. 2019, 32, 205–217. [Google Scholar] [CrossRef]
- Schmoll, T.; Kleven, O.; Rusche, M. Individual phenotypic plasticity explains seasonal variation in sperm morphology in a passerine bird. Evol. Ecol. Res. 2018, 19, 561–574. [Google Scholar]
- Girndt, A.; Cockburn, G.; Sánchez-Tójar, A.; Hertel, M.; Burke, T.; Schroeder, J. Male age and its association with reproductive traits in captive and wild house sparrows. J. Evol. Biol. 2019, 32, 1432–1443. [Google Scholar] [CrossRef] [Green Version]
- Laskemoen, T.; Kleven, O.; Johannessen, L.E.; Fossøy, F.; Robertson, R.J.; Lifjeld, J.T. Repeatability of sperm size and motility within and between seasons in the Barn Swallow (Hirundo rustica). J. Ornithol. 2013, 154, 955–963. [Google Scholar] [CrossRef]
- Lüpold, S.; Birkhead, T.R.; Westneat, D.F. Seasonal variation in ejaculate traits of male red-winged blackbirds (Agelaius phoeniceus). Behav. Ecol. Sociobiol. 2012, 66, 1607–1617. [Google Scholar] [CrossRef]
- Holmes, R.T.; Marra, P.P.; Sherry, T.W. Habitat-specific demography of breeding black-throated blue warblers (Dendroica caerulescens): Implications for population dynamics. J. Anim. Ecol. 1996, 65, 183–195. [Google Scholar] [CrossRef] [Green Version]
- Lifjeld, J.T.; Gohli, J.; Albrecht, T.; Garcia-Del-Rey, E.; Johannessen, L.E.; Kleven, O.; Marki, P.Z.; Omotoriogun, T.C.; Rowe, M.; Johnsen, A. Evolution of female promiscuity in Passerides songbirds. BMC Evol. Biol. 2019, 19, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
A single, Coomassie-blue stained spermatozoan from a black-throated blue warbler (400× total magnification). The sections measured as head, midpiece and tail (exposed flagellum) are labelled. The unstained portion of the head fluoresced strongly with a DAPI stain in pilot work.
Figure 1.
A single, Coomassie-blue stained spermatozoan from a black-throated blue warbler (400× total magnification). The sections measured as head, midpiece and tail (exposed flagellum) are labelled. The unstained portion of the head fluoresced strongly with a DAPI stain in pilot work.

Figure 2.
Sperm length from males in different age categories. Older males (in their second or later breeding season, ASY) are in dark grey, and young males in their first-breeding season (SY) are in light grey. Males are in order by total length within an age category. Boxplots show median, interquartile range (IQR), and greatest data extent within 1.5 × IQR, with more extreme data points indicated by dots. n = 20 cells per male.
Figure 2.
Sperm length from males in different age categories. Older males (in their second or later breeding season, ASY) are in dark grey, and young males in their first-breeding season (SY) are in light grey. Males are in order by total length within an age category. Boxplots show median, interquartile range (IQR), and greatest data extent within 1.5 × IQR, with more extreme data points indicated by dots. n = 20 cells per male.


{kind=link}
{kind=link}
Table 1.
Sperm component length comparisons for 12 black-throated blue warblers.
| Description | Head | Midpiece | Tail | Total Sperm Length (TSL) | CVam 1 |
|---|---|---|---|---|---|
| Mean (±SD, μm) | 14.31 ± 0.44 | 224.64 ± 5.40 | 8.71 ± 1.30 | 247.66 ± 5.11 | 2.1 |
| ANOVA comparison among males | F11,228 = 6.38, p < 0.001 | F11,228 = 19.41, p < 0.001 | F11,228 = 1.75, p = 0.06 | F11,228 = 48.42, p < 0.001 | n/a |
1 CVam is the adjusted coefficient of variation of mean sperm length among males. No statistical comparison was performed on this measure.
Table 2.
Statistical results of the relationship between sperm and ejaculate measurements compared to male traits possibly linked to extra-pair paternity. Uncorrected p-values are shown; after correction, no tests had p < 0.05.
Table 2.
Statistical results of the relationship between sperm and ejaculate measurements compared to male traits possibly linked to extra-pair paternity. Uncorrected p-values are shown; after correction, no tests had p < 0.05.
| Predictor | Response Variable | ||
|---|---|---|---|
| Total Sperm Length | Flagellum | CVwm 1 | |
| Male age | F1,10 = 11.74, p = 0.006 | F1,9 = 9.10, p = 0.01 | F1,10 = 0.31, p = 0.59 |
| Size of white wing patch | F1,10 = 0.11, p = 0.75 | F1,9 = 0.01, p = 0.92 | F1,10 = 1.19, p = 0.30 |
| Percent black dorsal feathers | F1,9 = 1.54, p = 0.25 | F1,8 = 1.63, p = 0.24 | F1,9 = 0.10, p = 0.76 |
| Volume of cloacal protuberance | F1,8 = 0.45, p = 0.52 | F1,7 = 0.19, p = 0.68 | F1,8 = 0.30, p = 0.60 |
1 Results were similar when we instead analyzed CVwm using standard deviation in total sperm length as the response variable, with mean total sperm length as a covariate.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cramer, E.; Krauss, N.; Rowlison, T.; Comizzoli, P. Sperm Morphology and Male Age in Black-Throated Blue Warblers, an Ecological Model System. Animals 2020, 10, 1175. https://doi.org/10.3390/ani10071175
AMA Style
Cramer E, Krauss N, Rowlison T, Comizzoli P. Sperm Morphology and Male Age in Black-Throated Blue Warblers, an Ecological Model System. Animals. 2020; 10(7):1175. https://doi.org/10.3390/ani10071175
Chicago/Turabian StyleCramer, Emily, Nicole Krauss, Tricia Rowlison, and Pierre Comizzoli. 2020. "Sperm Morphology and Male Age in Black-Throated Blue Warblers, an Ecological Model System" Animals 10, no. 7: 1175. https://doi.org/10.3390/ani10071175
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.