Optogenetic Stimulation of Human Neural Networks Using Fast Ferroelectric Spatial Light Modulator—Based Holographic Illumination

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
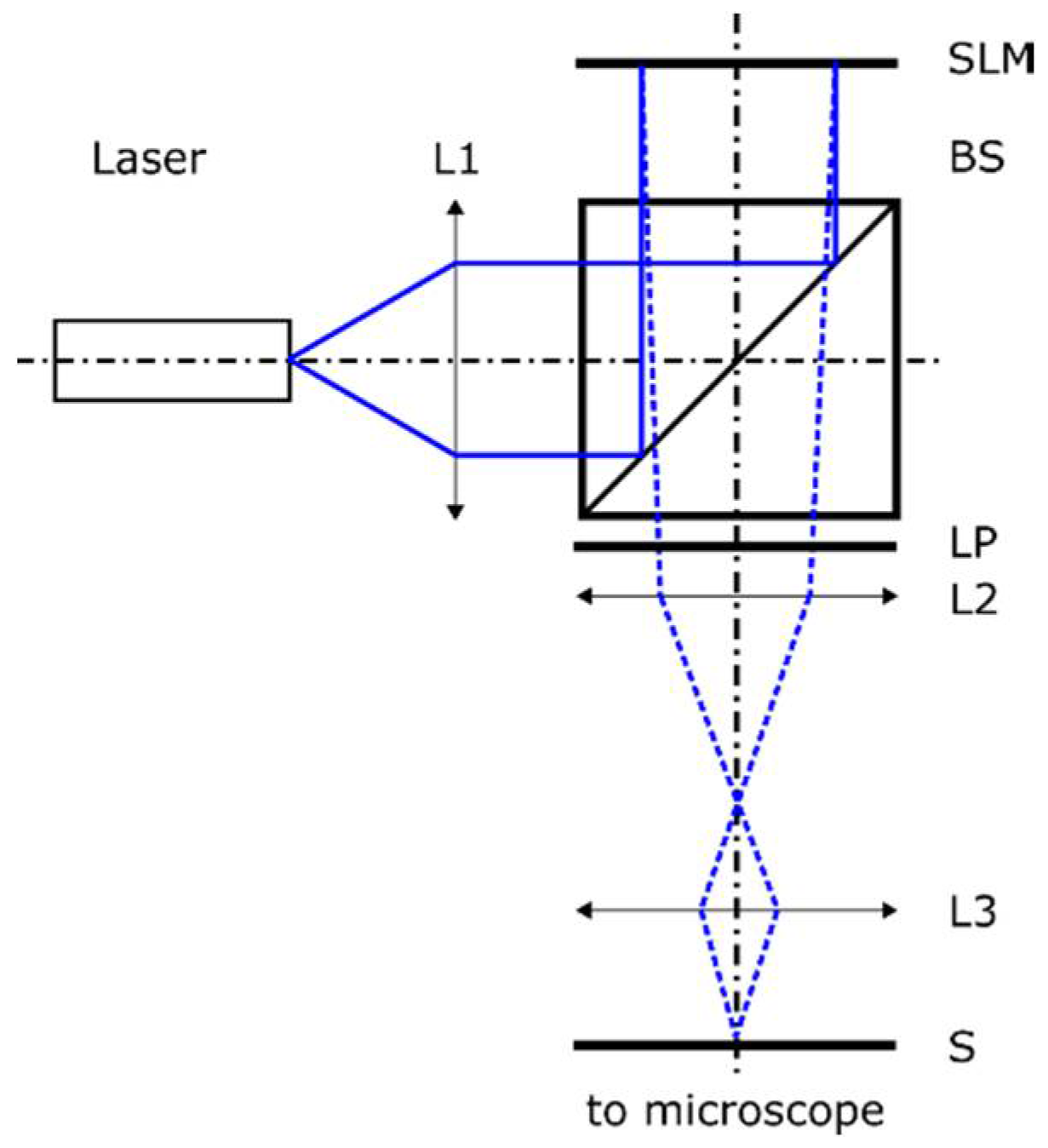
2. Holographic Single-Cell Illumination Setup
2.1. Modulator
2.2. Computer-Generated Holograms
2.3. Computer-Generated Hologram Calculation and Binarization
2.4. Sample
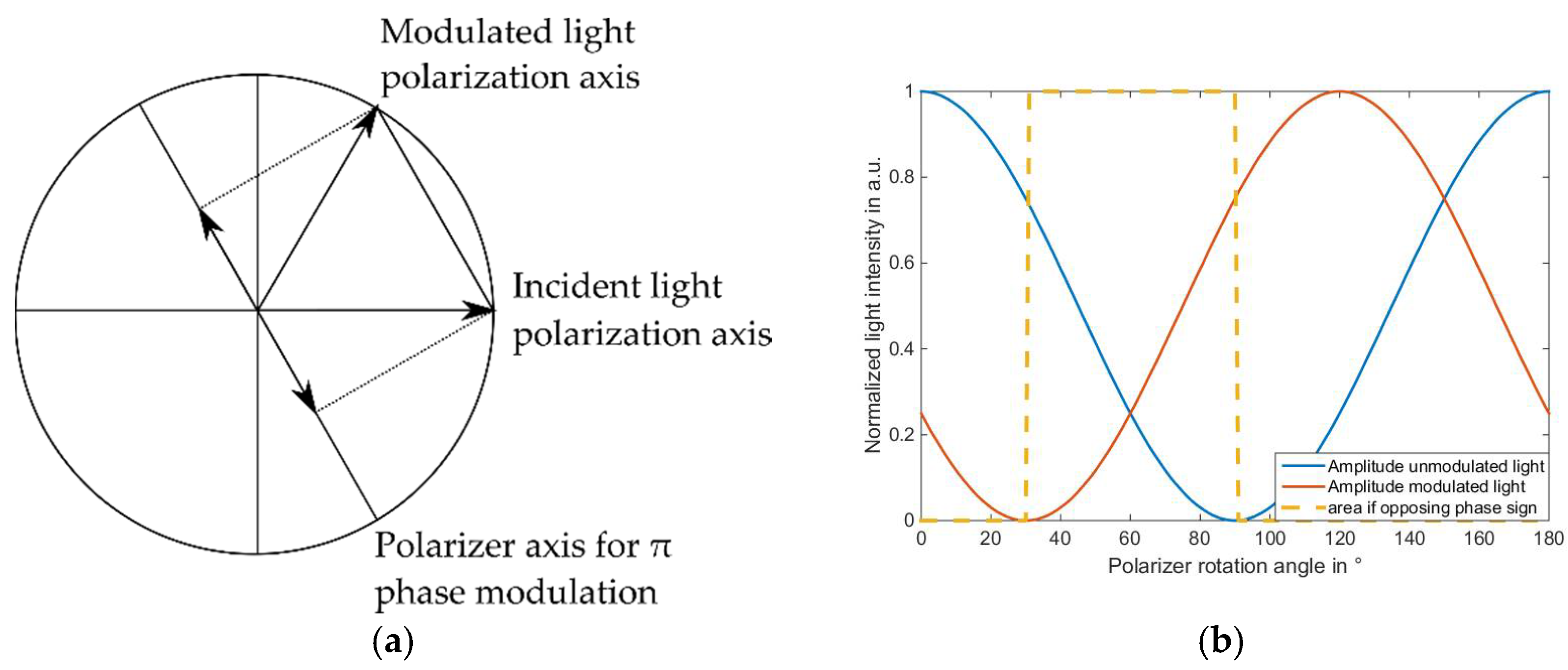
2.5. Calibration of the Illumination System
3. Results
3.1. Setup Specifications
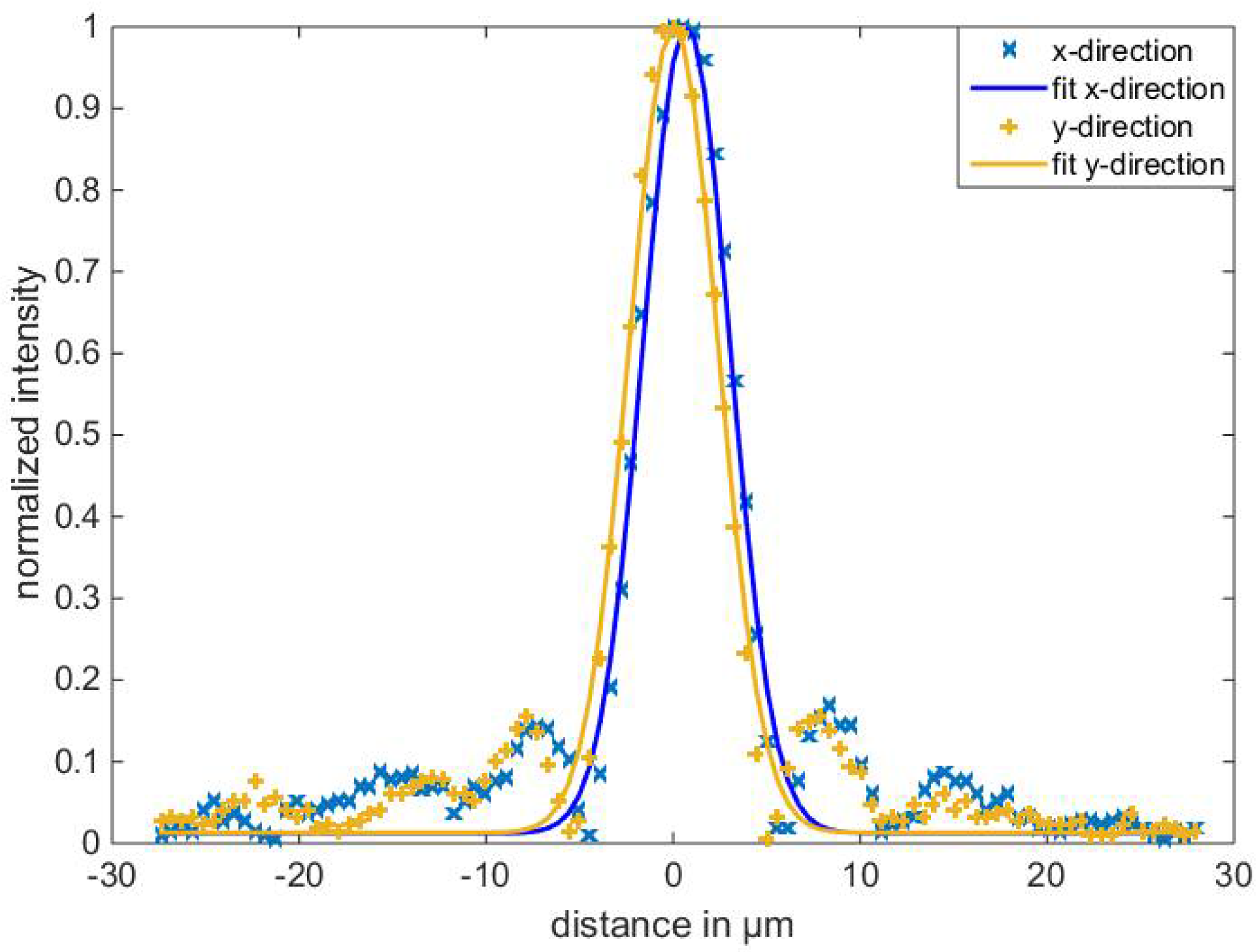
3.1.1. Minimal Focus Diameter
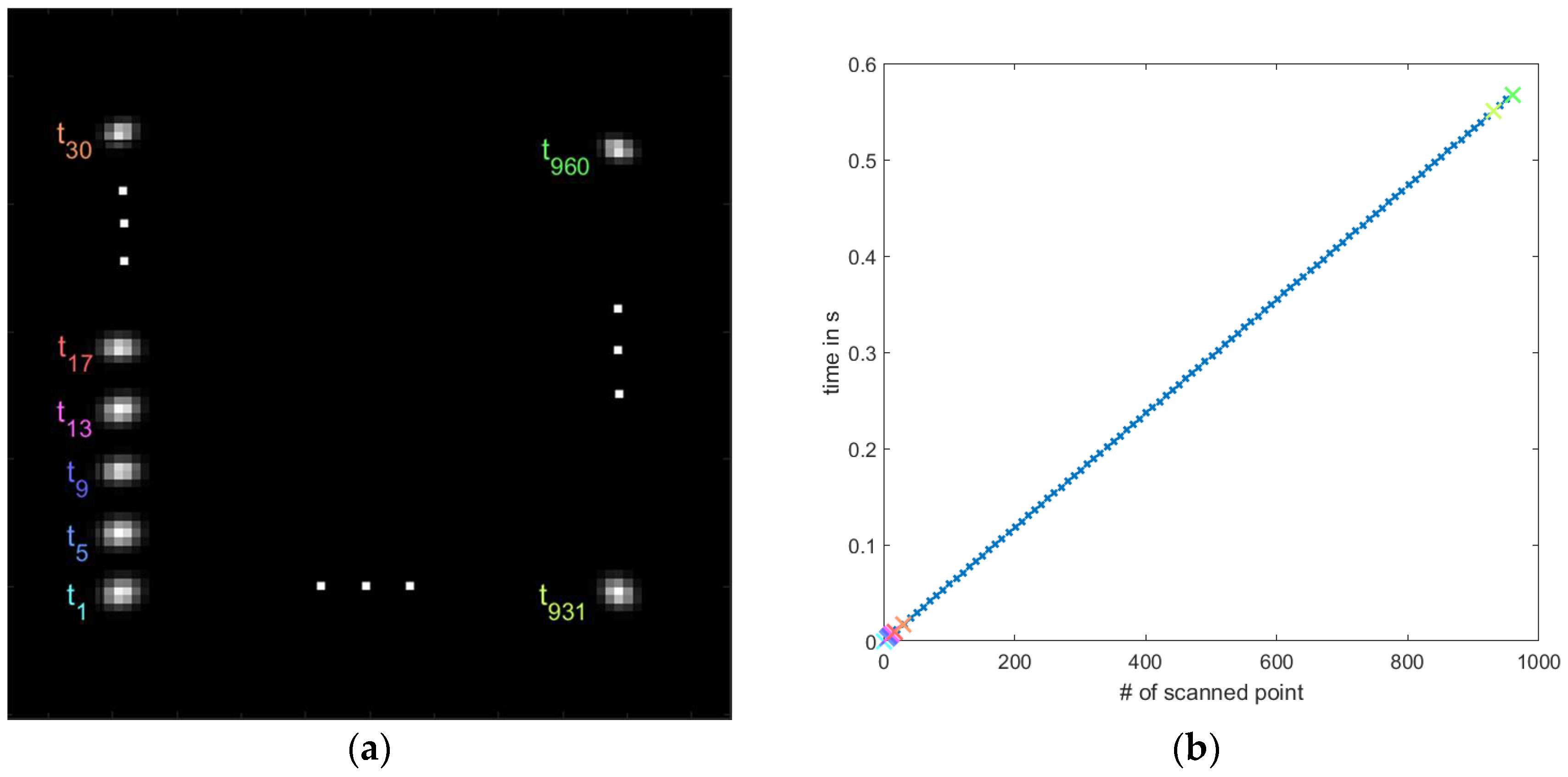
3.1.2. Pattern Update Rate
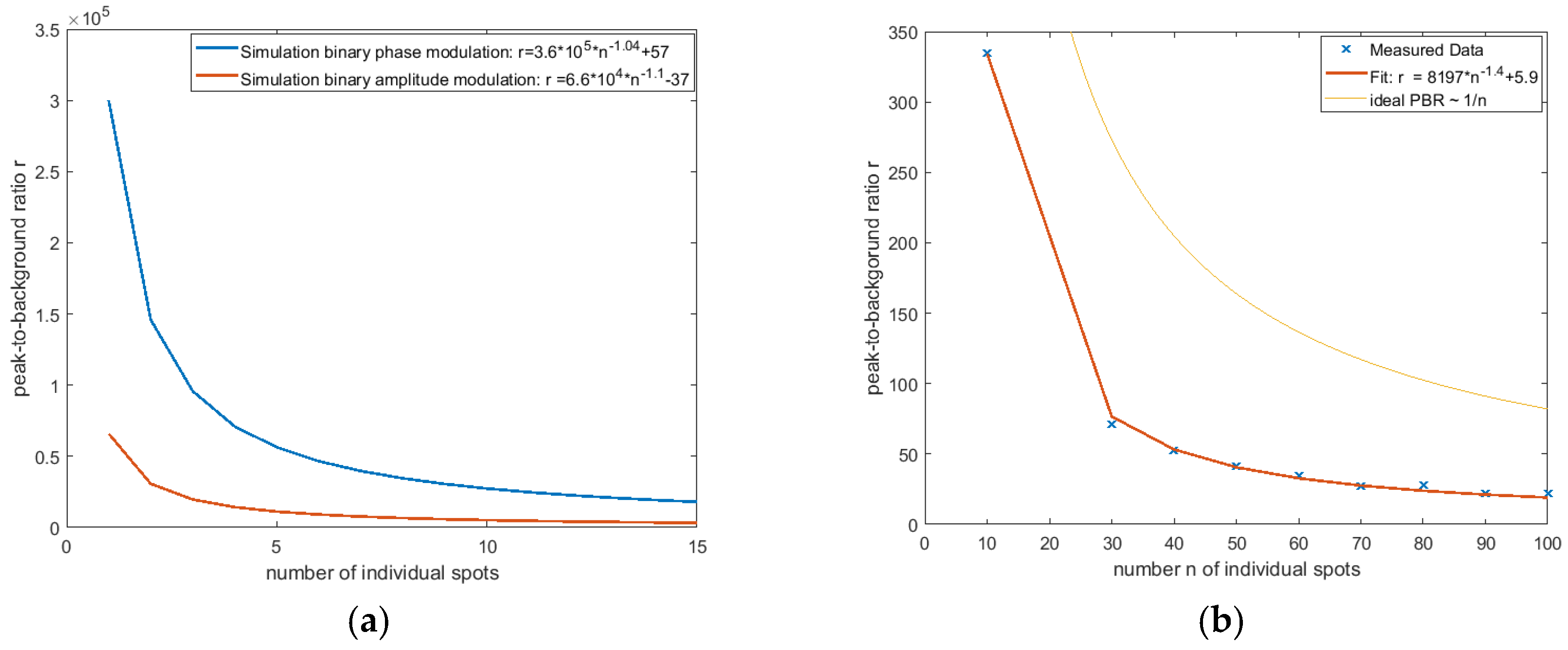
3.1.3. Generation of Multiple Foci
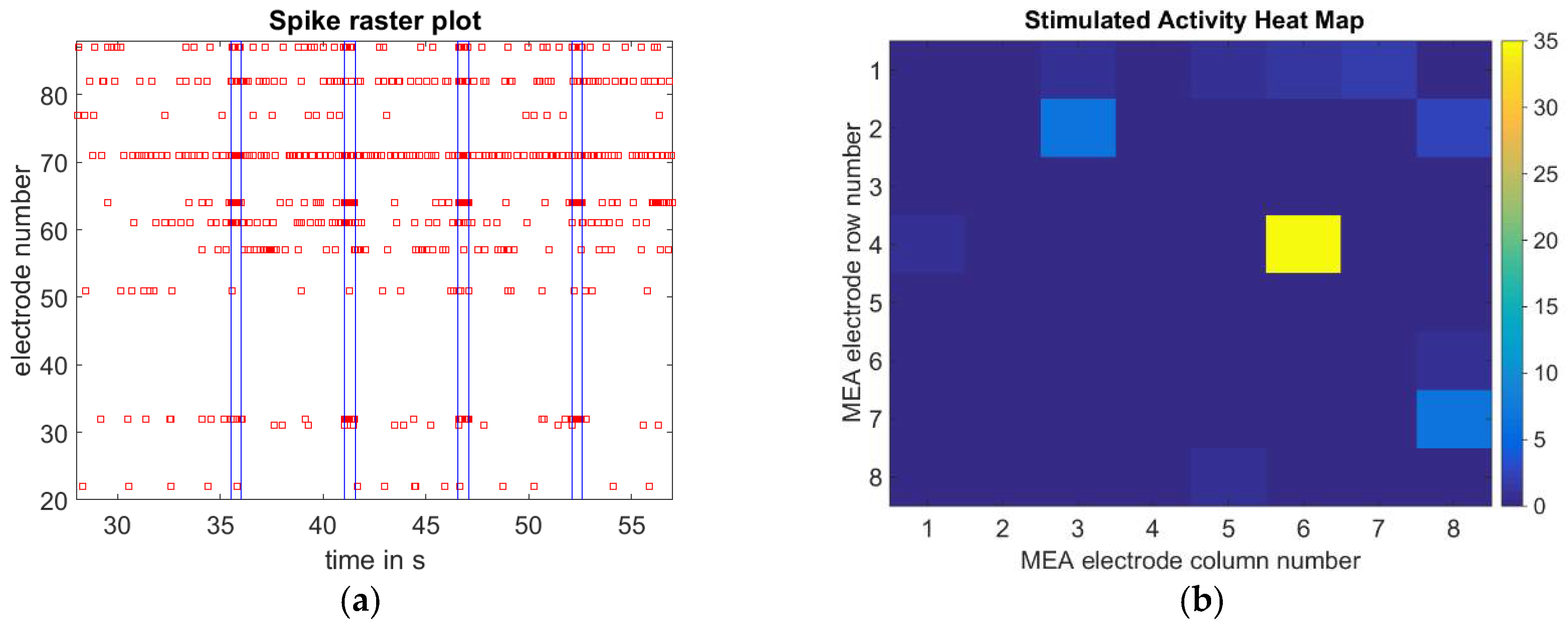
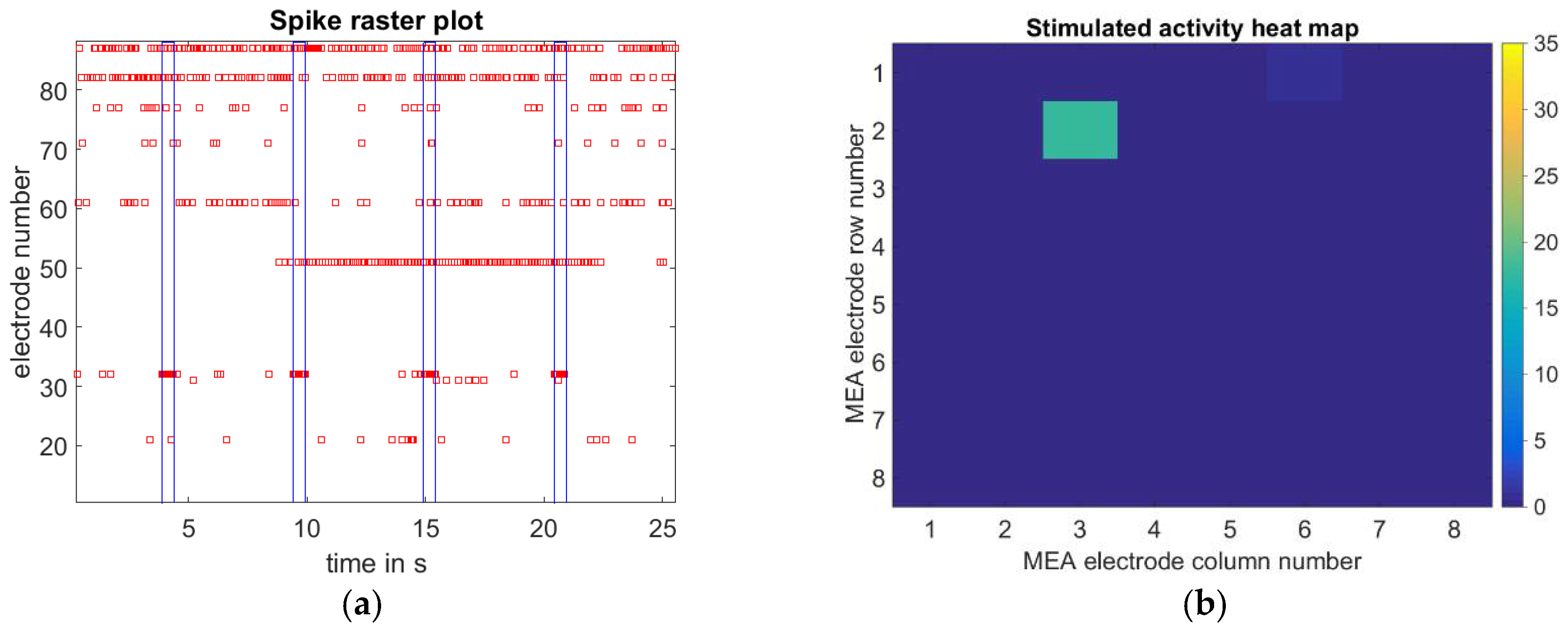
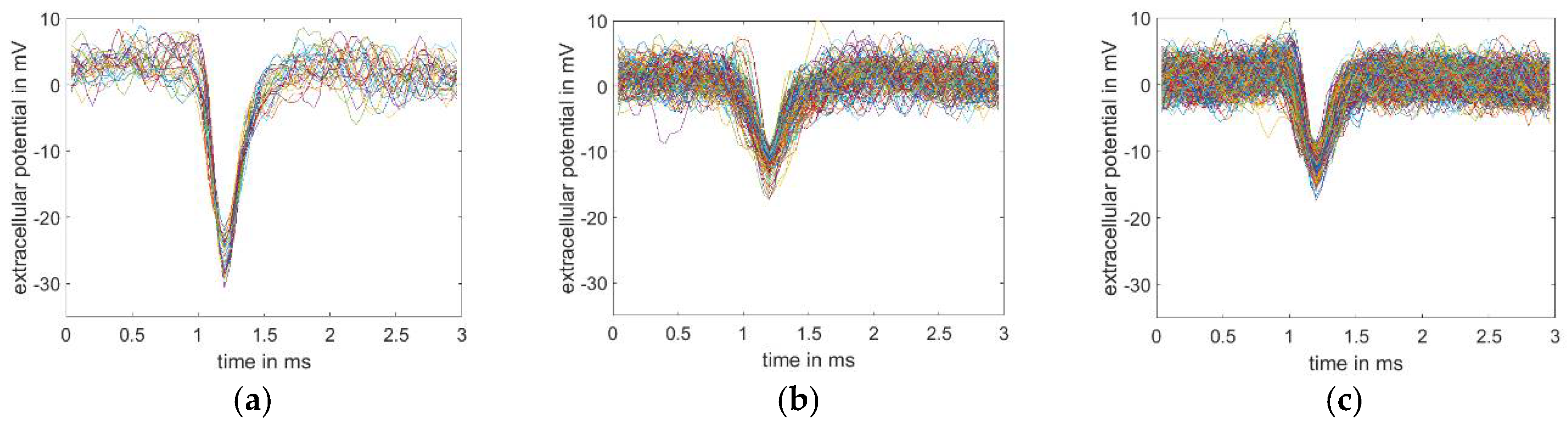
3.2. Single Cell Stimulation
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Betzel, R.F.; Byrge, L.; He, Y.; Goñi, J.; Zuo, X.-N.; Sporns, O. Changes in structural and functional connectivity among resting-state networks across the human lifespan. NeuroImage 2014, 102 Pt 2, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Van den Heuvel, M.P.; Sporns, O. Rich-Club Organization of the Human Connectome. J. Neurosci. 2011, 31, 15775–15786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schröter, M.; Paulsen, O.; Bullmore, E.T. Micro-connectomics: Probing the organization of neuronal networks at the cellular scale. Nat. Rev. Neurosci. 2017, 18, 131–146. [Google Scholar] [CrossRef] [PubMed]
- Tijms, B.M.; Wink, A.M.; de Haan, W.; van der Flier, W.M.; Stam, C.J.; Scheltens, P.; Barkhof, F. Alzheimer’s disease: Connecting findings from graph theoretical studies of brain networks. Neurobiol. Aging 2013, 34, 2023–2036. [Google Scholar] [CrossRef] [PubMed]
- Sporns, O.; Chialvo, D.R.; Kaiser, M.; Hilgetag, C.C. Organization, development and function of complex brain networks. Trends Cogn. Sci. 2004, 8, 418–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marom, S.; Shahaf, G. Development, learning and memory in large random networks of cortical neurons: Lessons beyond anatomy. Q. Rev. Biophys. 2002, 35, 63–87. [Google Scholar] [CrossRef] [PubMed]
- Nabavi, S.; Fox, R.; Proulx, C.D.; Lin, J.Y.; Tsien, R.Y.; Malinow, R. Engineering a memory with LTD and LTP. Nature 2014, 511, 348–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attarwala, H. TGN1412: From Discovery to Disaster. J. Young Pharm. JYP 2010, 2, 332–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, R.S.G.; da Silva, A.B.; Whittaker, R.G.; Woodhall, G.L.; Cunningham, M.O. Human brain slices for epilepsy research: Pitfalls, solutions and future challenges. J. Neurosci. Methods 2016, 260, 221–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busskamp, V.; Lewis, N.E.; Guye, P.; Ng, A.H.M.; Shipman, S.L.; Byrne, S.M.; Sanjana, N.E.; Murn, J.; Li, Y.; Li, S.; et al. Rapid neurogenesis through transcriptional activation in human stem cells. Mol. Syst. Biol. 2014, 10, 760. [Google Scholar] [CrossRef] [PubMed]
- Lam, R.S.; Töpfer, F.M.; Wood, P.G.; Busskamp, V.; Bamberg, E. Functional Maturation of Human Stem Cell-Derived Neurons in Long-Term Cultures. PLoS ONE 2017, 12, e0169506. [Google Scholar] [CrossRef] [PubMed]
- Boyden, E.S.; Zhang, F.; Bamberg, E.; Nagel, G.; Deisseroth, K. Millisecond-timescale, genetically targeted optical control of neural activity. Nat. Neurosci. 2005, 8, 1263–1268. [Google Scholar] [CrossRef] [PubMed]
- Sakmann, B.; Neher, E. Patch Clamp Techniques for Studying Ionic Channels in Excitable Membranes. Annu. Rev. Physiol. 1984, 46, 455–472. [Google Scholar] [CrossRef] [PubMed]
- Staley, K.J.; Otis, T.S.; Mody, I. Membrane properties of dentate gyrus granule cells: Comparison of sharp microelectrode and whole-cell recordings. J. Neurophysiol. 1992, 67, 1346–1358. [Google Scholar] [CrossRef] [PubMed]
- Shahaf, G.; Marom, S. Learning in Networks of Cortical Neurons. J. Neurosci. 2001, 21, 8782–8788. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Wang, L.-P.; Boyden, E.S.; Deisseroth, K. Channelrhodopsin-2 and optical control of excitable cells. Nat. Methods 2006, 3, 785–792. [Google Scholar] [CrossRef] [PubMed]
- Busskamp, V.; Duebel, J.; Balya, D.; Fradot, M.; Viney, T.J.; Siegert, S.; Groner, A.C.; Cabuy, E.; Forster, V.; Seeliger, M.; et al. Genetic Reactivation of Cone Photoreceptors Restores Visual Responses in Retinitis Pigmentosa. Science 2010, 329, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, V.H.; Gehrt, A.; Reuter, K.; Jing, Z.; Jeschke, M.; Mendoza Schulz, A.; Hoch, G.; Bartels, M.; Vogt, G.; Garnham, C.W.; et al. Optogenetic stimulation of the auditory pathway. J. Clin. Investig. 2014, 124, 1114–1129. [Google Scholar] [CrossRef] [PubMed]
- Tzur, A.; Kafri, R.; LeBleu, V.S.; Lahav, G.; Kirschner, M.W. Cell Growth and Size Homeostasis in Proliferating Animal Cells. Science 2009, 325, 167–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.T.; Gu, S.; Ma, P.; Watanabe, M.; Rollins, A.M.; Jenkins, M.W. Optical stimulation enables paced electrophysiological studies in embryonic hearts. Biomed. Opt. Express 2014, 5, 1000–1013. [Google Scholar] [CrossRef] [PubMed]
- Packer, A.M.; Russell, L.E.; Dalgleish, H.W.P.; Häusser, M. Simultaneous all-optical manipulation and recording of neural circuit activity with cellular resolution in vivo. Nat. Methods 2015, 12, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Packer, A.M.; Peterka, D.S.; Hirtz, J.J.; Prakash, R.; Deisseroth, K.; Yuste, R. Two-photon optogenetics of dendritic spines and neural circuits. Nat. Methods 2012, 9, 1202–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prakash, R.; Yizhar, O.; Grewe, B.; Ramakrishnan, C.; Wang, N.; Goshen, I.; Packer, A.M.; Peterka, D.S.; Yuste, R.; Schnitzer, M.J.; et al. Two-photon optogenetic toolbox for fast inhibition, excitation and bistable modulation. Nat. Methods 2012, 9, 1171–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papagiakoumou, E. Optical developments for optogenetics. Biol. Cell 2013, 105, 443–464. [Google Scholar] [CrossRef] [PubMed]
- Bañas, A.; Palima, D.; Villangca, M.; Aabo, T.; Glückstad, J. GPC Light Shaper for speckle-free one- and two-photon contiguous pattern excitation. Opt. Express 2014, 22, 5299–5311. [Google Scholar] [CrossRef] [PubMed]
- Papagiakoumou, E.; Anselmi, F.; Bègue, A.; de Sars, V.; Glückstad, J.; Isacoff, E.Y.; Emiliani, V. Scanless two-photon excitation of channelrhodopsin-2. Nat. Methods 2010, 7, 848. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, P.J.; Daria, V.R.; Glückstad, J. Four-dimensional optical manipulation of colloidal particles. Appl. Phys. Lett. 2005, 86, 074103. [Google Scholar] [CrossRef]
- Czarske, J.W.; Haufe, D.; Koukourakis, N.; Büttner, L. Transmission of independent signals through a multimode fiber using digital optical phase conjugation. Opt. Express 2016, 24, 15128–15136. [Google Scholar] [CrossRef] [PubMed]
- Koukourakis, N.; Fregin, B.; König, J.; Büttner, L.; Czarske, J.W. Wavefront shaping for imaging-based flow velocity measurements through distortions using a Fresnel guide star. Opt. Express 2016, 24, 22074–22087. [Google Scholar] [CrossRef] [PubMed]
- Papagiakoumou, E.; de Sars, V.; Oron, D.; Emiliani, V. Patterned two-photon illumination by spatiotemporal shaping of ultrashort pulses. Opt. Express 2008, 16, 22039–22047. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Salter, P.S.; Roider, C.; Jesacher, A.; Strauss, J.; Heberle, J.; Schmidt, M.; Booth, M.J. Four-dimensional light shaping: Manipulating ultrafast spatiotemporal foci in space and time. Light Sci. Appl. 2018, 7, 17117. [Google Scholar] [CrossRef]
- Reutsky-Gefen, I.; Golan, L.; Farah, N.; Schejter, A.; Tsur, L.; Brosh, I.; Shoham, S. Holographic optogenetic stimulation of patterned neuronal activity for vision restoration. Nat. Commun. 2013, 4, 1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klapper, S.D.; Sauter, E.J.; Swiersy, A.; Hyman, M.A.E.; Bamann, C.; Bamberg, E.; Busskamp, V. On-demand optogenetic activation of human stem-cell-derived neurons. Sci. Rep. 2017, 7, 14450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vellekoop, I.M. Feedback-based wavefront shaping. Opt. Express 2015, 23, 12189–12206. [Google Scholar] [CrossRef] [PubMed]
- Cottrell, D.M.; Davis, J.A.; Hedman, T.R.; Lilly, R.A. Multiple imaging phase-encoded optical elements written as programmable spatial light modulators. Appl. Opt. 1990, 29, 2505–2509. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.-B.; Ma, J.-S.; Su, P.; Liang, X.-T. Binary hologram generation based on discrete wavelet transform. Opt. Int. J. Light Electron Opt. 2016, 127, 558–561. [Google Scholar] [CrossRef]
- Tsang, P.W.M.; Poon, T.-C. Novel method for converting digital Fresnel hologram to phase-only hologram based on bidirectional error diffusion. Opt. Express 2013, 21, 23680–23686. [Google Scholar] [CrossRef] [PubMed]
- Han, X. In Vivo Application of Optogenetics for Neural Circuit Analysis. ACS Chem. Neurosci. 2012, 3, 577–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayling, O.G.S.; Harrison, T.C.; Boyd, J.D.; Goroshkov, A.; Murphy, T.H. Automated light-based mapping of motor cortex by photoactivation of channelrhodopsin-2 transgenic mice. Nat. Methods 2009, 6, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.Y.; Lin, M.Z.; Steinbach, P.; Tsien, R.Y. Characterization of Engineered Channelrhodopsin Variants with Improved Properties and Kinetics. Biophys. J. 2009, 96, 1803–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quiroga, R.Q.; Nadasdy, Z.; Ben-Shaul, Y. Unsupervised Spike Detection and Sorting with Wavelets and Superparamagnetic Clustering. Neural Comput. 2004, 16, 1661–1687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- jm303 Spike Sorting—University of Leicester. Available online: https://www2.le.ac.uk/departments/engineering/research/bioengineering/neuroengineering-lab/spike-sorting (accessed on 22 June 2018).










© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmieder, F.; Klapper, S.D.; Koukourakis, N.; Busskamp, V.; Czarske, J.W. Optogenetic Stimulation of Human Neural Networks Using Fast Ferroelectric Spatial Light Modulator—Based Holographic Illumination. Appl. Sci. 2018, 8, 1180. https://doi.org/10.3390/app8071180
Schmieder F, Klapper SD, Koukourakis N, Busskamp V, Czarske JW. Optogenetic Stimulation of Human Neural Networks Using Fast Ferroelectric Spatial Light Modulator—Based Holographic Illumination. Applied Sciences. 2018; 8(7):1180. https://doi.org/10.3390/app8071180
Chicago/Turabian StyleSchmieder, Felix, Simon D. Klapper, Nektarios Koukourakis, Volker Busskamp, and Jürgen W. Czarske. 2018. "Optogenetic Stimulation of Human Neural Networks Using Fast Ferroelectric Spatial Light Modulator—Based Holographic Illumination" Applied Sciences 8, no. 7: 1180. https://doi.org/10.3390/app8071180