Myricetin: A Dietary Molecule with Diverse Biological Activities



Abstract
:1. Introduction

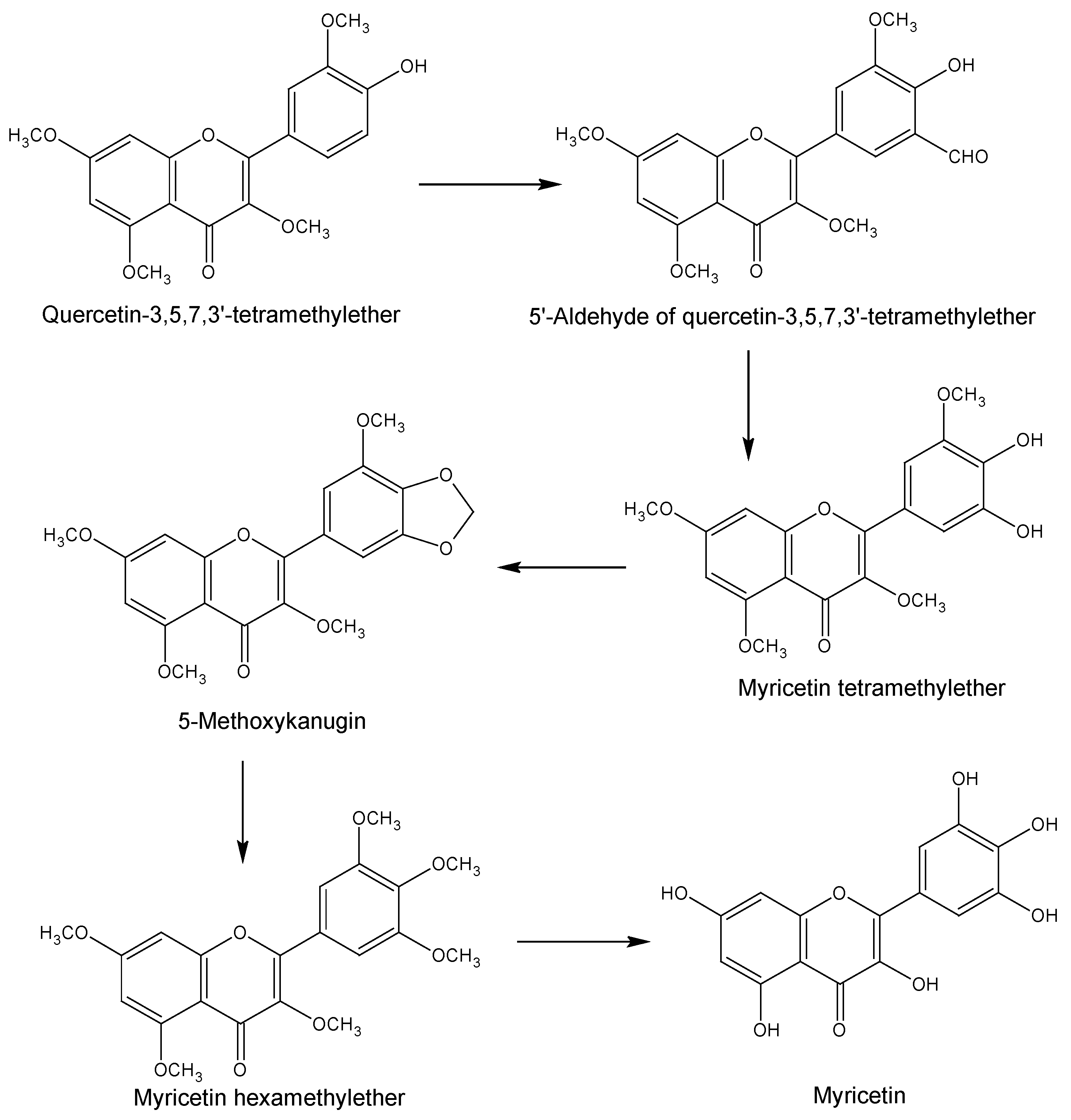
2. Chemical Synthesis


3. Pharmacological Applications
3.1. Anti-Oxidant Activity

{kind=link}
{kind=link}
{kind=link}
| Assay | Results | Control | Reference |
|---|---|---|---|
| DPPH | At 1 mg/mL inhibited DPPH radical by 71.5%. IC50 value was found to be 9 µg/mL | α-tocopherol (IC50 = 26 µg/mL) and BHT (IC50 = 30 µg/mL Trolox (1 mg/mL) inhibited DPPH radical by 61.5% | [20] |
| At 0.01 mM (3.2 µg/mL), 0.1 mM (32 μg/mL) and 1 mM (320 µg/mL) inhibited DPPH radical by 85.6%, 92.8% and 96.9%, respectively, whereas IC50 value was 4 µM (1.3 µg/mL) | β-Actin as internal control | [21] | |
| At 40 µg/mL inhibited DPPH radical by 78% | Rutin (85% inhibition) at 40 µg/mL | [22] | |
| Superoxide | Inhibited by 24.6%, 79.5% and 96.4% when applying concentrations of 0.001 mM (0.32 µg/mL), 0.01 mM (3.2 µg/mL) and 0.1 mM (32 µg/mL), respectively, while IC50 was calculated as 0.6 µM (0.2 µg/mL) | β-Actin as internal control | [21] |
| At 1.86 μg/mL, scavenged superoxide radicals in the nitroblue tetrazolium hypoxanthine/xanthine oxidase assay | Ascorbic acid (IC50 5.8 µg/mL) | [23] | |
| TEAC | Activity of 2.40 mM (764 μg/mL) trolox/mg sample after 20 min. The IC50 value was found to 22 µg/mL | Trolox (0.2 mg/mL) | [24] |
| ORAC | 1620 µmol trolox equivalent/g (515 mg/g) | - | [25] |
| FRAP | 590 µmol Fe2+/L at 10 µM (0.32 μg/mL) | Gallic acid | [26] |
| Ascorbic acid-induced lipid peroxidation | Inhibited in rat brain by 92%, 95% and 95% at 0.1, 1.0 and 4.0 mM myricetin, respectively (concentrations correspond to 32, 320 μg/mL and 1.3 mg/mL, respectively). | Ascorbic acid (0.1 or 1.0 mM) | [27] |
| Ferrous sulfate-induced lipid peroxidation | Inhibited in rat brain by 28%, 71% and 91% at 0.1, 1.0 and 4.0 mM myricetin, respectively (concentrations correspond to 32, 320 μg/mL and 1.3 mg/mL, respectively). | Ferrous sulfate (1.0 mM) | [27] |
| Oleic acid triglyceride | Inhibited oleic acid-induced triglyceride over-accumulation towards HepG2 cells by 24.8% with IC30 > 150 µM (47 µg/mL) | - | [28] |
| ROS | 34.5% inhibition with IC30 122.7 µM (39.0 µg/mL) | - | [28] |
| NO | At a dosage of 50 mg/kg, decreased NO production by 56.7, 31.4, 7.7, 48.9 and 53.4 ng/g tissue in the brain cortex, liver, kidney, blood and lungs, respectively, of intact rats. Together with lipopolysaccharide (10 mg/kg) and at the same concentration, decreased the level of NO production in these organs by 206.5, 1008.3, 337.0, 542.8 and 824.8 ng/g tissue, respectively | - | [29] |
| NO-scavenging capacity with kAOx/kPTIO value of 1.2 TEU | Trolox (7.3 TEU) | [30] | |
| Collagenase in human dermal fibroblasts | Inhibited by 12.7% and 29.6%.at myricetin concentration of 0.1 (32 μg/mL) and 0.2 mM (64 μg/mL), respectively | 1,10-phenanthroline (39.4% and 75.1%, respectively) | [21] |
| Peroxynitrite anions | Antioxidant effects against peroxynitrite anions, chemiluminescence initiated by peroxynitrite in rat liver homogenate and lucigenin chemiluminescence in aortic rings with IC50 values of 35, 20 and 32 μM, respectively | - | [31] |
3.2. Antiphotoaging Activity
3.3. Anticancer Activity
| Cell line/Enzyme | Effect of Myricetin | Reference |
|---|---|---|
| Activity on Cell Lines | ||
| Brain | ||
| U251, NCH89 and LN229 cells | No effect when alone, since the IC50 value for each cell line was found to be >200 µM. A combination of myricetin (150 µM) and TRAIL (50 ng/mL) yielded a synergistic activity and increased cell death in U251, NCH89 and LN229 by 59%, 65% and 52%, respectively. | [78] |
| Breast | ||
| MCF-7 | IC50 2.70 μg/mL compared to vinblastine (IC50 45.6 μg/mL) | [79] |
| Increased GSH content of cells and also increased the EROD reaction 2-fold at a concentration of 25 μM | [80] | |
| Cervix | ||
| HeLa cells | Cytotoxic with IC50 18.9 µg/mL | [81] |
| Colon | ||
| Epithelial adenocarcinoma cells | Proliferation of cells inhibited at 50 μM by decreasing COX-2 and cyclin D1 expression | [82] |
| HCT116 | Inhibited the proliferation of human colon carcinoma cells by halting the cell cycle in G2/M phase and inducing apoptosis; LD50 28.2 μM | [83] |
| COLO 205, COLO 320HSR, COLO 320DM, HT 29 and COLO 205-X | Inhibited the activation of MMP-2 enzyme in the cells with IC50 values of 7.82, 11.18, 11.56, 13.25 and 23.51 μM, respectively. It also suppressed TPA-induced MMP-2 protein expression in COLO 205 cells by blocking the translocation of PKCα from cytosol to membrane, phosphorylation of ERK1/2 protein and induction of c-Jun protein expression activated by TPA. | [84] |
| Leukemia | ||
| HL-60 | Alone, and in combination with piceatannol, induced apoptotic cell death through a ROS-independent cell death pathway. The effect was greater with the combined treatment | [85] |
| Anti-proliferative activity and the effect was enhanced with increasing concentration | [70] | |
| Prostate | ||
| LNCaP | IC50 value 2.10 μg/mL while taxol (IC50 0.08 μg/mL) used as standard | [79] |
| 22Rv1 | Inhibition of TCDD-induced EROD activity in cancer cells; IC50 value 3.0 μM | [86] |
| Uterus | ||
| RL95-2 endometrial cancer cells | Inhibition of CYP1 activity of cancer cells; IC50 values 3 μM and lower | [87] |
| Inhibition of enzyme/protein activity | ||
| Thioredoxin reductase (TrxR) from mammals | Inhibitory effect on enzyme, which is overexpressed in many aggressive tumours; IC50 value 0.62 μM. Attacks the reduced COOH-terminal of -Cys-Sec-Gly, the active site of TrxR | [88] |
| TrxR | At 50 μM, inhibited growth of A549 (human lung carcinoma) cells and reduced TrxR activity in the cell lysates, corresponding with the oxidization of thioredoxin | [88] |
| Mammalian DNA polymerases | IC50 values ranged from 21.3 to 40.9 μM. Human DNA topoisomerase II activity inhibited; IC50 27.5 μM | [83] |
| Phosphatidylinositol 3-kinase (PI3K) | Inhibited this enzyme (IC50 1.8 μM) that plays an important role in signal transduction and cell transformation. Also inhibited PKC and tyrosine kinase activity of EGF-R | [89,90] |
| E6, a primary oncoprotein of human papillomaviruses | Inhibited E6, responsible for cervical cancer by inhibiting GST-E6 and His-caspase 8 binding | [91] |
| CCAAT-enhancer-binding proteins-α, peroxisome proliferator-activated receptor-γ, lipoprotein lipase, fatty acid binding aP2 protein and adiponectin | At 30 μM, myricetin decreased mRNA levels of these enzymes. Inhibited adipogenesis in human adipose tissue-derived mesenchymal stem cells. | [92] |
| Multidrug resistance-associated protein MRP1 and MRP2 mediated vincristine efflux in MDCKII cells | Inhibitory effects with IC50 30.5 and 24.6 μM, respectively. At a concentration of 25 μM, it increased the sensitivity of the cells towards vincristine toxicity towards MRP1 and MRP2 cells with IC50 values of 7.6 and 5.8 μM, respectively | [93] |
3.4. Anti-Platelet Aggregation Activity
3.5. Antihypertensive Activity
3.6. Immunomodulatory Activity
3.7. Anti-Inflammatory Activity
3.8. Anti-Allergic Activity
3.9. Analgesic Activity
3.10. Activity Against Bone-Related Disorders
3.11. Activity against CNS Disorders
3.12. Hepatoprotective and Hypouricemic Activities
3.13. Activity against Cardiovascular Diseases
3.14. Activity against Eye Disorders
3.15. Antidiabetic and Anti-Obesity Activities
3.16. Antimicrobial Activity
3.17. Miscellaneous Activities
4. Toxicity Studies
5. Recommendations and Concluding Remarks
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Lau-Cam, C.A.; Chan, H.H. Flavonoids from Comptonia peregrine. Phytochemistry 1973, 12, 1829. [Google Scholar] [CrossRef]
- Jones, J.R.; Lebar, M.D.; Jinwal, U.K.; Abisambra, J.F.; Koren, J.; Blair, L.; O’Leary, J.C.; Davey, Z.; Trotter, J.; Johnson, A.G.; et al. The diarylheptanoid (+)-aR,11S-myricanol and two flavones from bayberry (Myrica cerifera) destabilize the microtubule-associated protein tau. J. Nat. Prod. 2011, 74, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Umadevi, I.; Daniel, M.; Sabnis, S.D. Chemotaxonomic studies on some members of Anacardiaceae. Proc. Plant Sci. 1988, 98, 205–208. [Google Scholar]
- El-Kader, A.M.A.; El-Readi, M.Z.; Ahmed, A.S.; Nafady, A.M.; Wink, M.; Ibraheim, Z.Z. Polyphenols from aerial parts of Polygonum bellardii and their biological activities. Pharm. Biol. 2013, 51, 1026–1034. [Google Scholar] [CrossRef] [PubMed]
- Hergert, H.L. The flavonoids of lodgepole pine bark. J. Org. Chem. 1956, 21, 534–537. [Google Scholar] [CrossRef]
- Chua, L.S.; Latiff, N.A.; Lee, S.Y.; Lee, C.T.; Sarmidi, M.R.; Aziz, R.A. Flavonoids and phenolic acids from Labisia pumila (Kacip Fatimah). Food Chem. 2011, 127, 1186–1192. [Google Scholar] [CrossRef] [PubMed]
- De Leo, M.; Braca, A.; Sanogo, R.; Cardile, V.; deTommasi, N.; Russo, A. Antiproliferative activity of Pteleopsis suberosa leaf extract and its flavonoid components in human prostate carcinoma cells. Planta Med. 2006, 72, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Kong, N.N.; Fang, S.T.; Wang, J.H.; Wang, Z.H.; Xia, C.H. Two new flavonoid glycosides from the halophyte Limonium franchetii. J. Asian Nat. Prod. Res. 2014, 16, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Lin, G.; Xie, Y.; Ma, P.; Li, G.; Meng, Q.; Wu, T. Preformulation studies of myricetin: A natural antioxidant flavonoid. Pharmazie 2014, 69, 19–26. [Google Scholar] [PubMed]
- Perkin, A.G.; Hummel, J.J. LXXVI-The colouring principle contained in the bark of Myrica nagi Part I. J. Chem. Soc. Trans. 1896, 69, 1287–1294. [Google Scholar] [CrossRef]
- Perkin, A.G. XXI.-Myricetin. Part II. J. Chem. Soc. Trans. 1902, 81, 203–210. [Google Scholar] [CrossRef]
- Perkin, A.G. CXCIII. Myricetin. Part III. J. Chem. Soc. Trans. 1911, 99, 1721–1725. [Google Scholar] [CrossRef]
- Lin, G.B.; Xie, Y.; Li, G.W. Research advances of myricetin. J. Int. Pharm. Res. 2012, 39, 483–487. [Google Scholar]
- Rao, K.V.; Seshadri, T.R. Nuclear oxidation in the flavone series—Part IV. New synthesis of herbacetin and hibiscetin. Proc. Indian Acad. Sci. A 1948, 25, 444–448. [Google Scholar]
- Tranchimand, S.; Tron, T.; Gaudin, C.; Iacazio, G. First chemical synthesis of three natural depsides involved in flavonol catabolism and related to quercetinase catalysis. Synth. Commun. 2006, 36, 587–597. [Google Scholar] [CrossRef]
- Dean, H.F.; Nierenstein, M. Attempts to synthesize myricetin. J. Am. Chem. Soc. 1925, 47, 1676–1684. [Google Scholar] [CrossRef]
- Kalff, J.; Robinson, R. XXVIII.-A synthesis of myricetin and of a galangin monomethyl ether occurring in galanga root. J. Chem. Soc. Trans. 1925, 127, 181–184. [Google Scholar] [CrossRef]
- Rao, K.V.; Seshadri, T.R. Nuclear oxidation in flavones and related compounds—Part XVI. A new synthesis of myricetin. Proc. Indian Acad. Sci. A 1948, 28, 210–215. [Google Scholar]
- Ong, K.C.; Khoo, H.E. Biological effects of myricetin. Gen. Pharmacol. 1997, 29, 121–126. [Google Scholar] [CrossRef]
- Qu, G.Z.; Si, C.L.; Wang, M.H. Antioxidant constituents from Leonurus japonicas. Nat. Prod. Sci. 2006, 12, 197–200. [Google Scholar]
- Sim, G.S.; Lee, B.C.; Ho, S.C.; Jae, W.L.; Kim, J.H.; Lee, D.H.; Kim, J.H.; Pyo, H.B.; Dong, C.M.; Oh, K.W.; et al. Structure activity relationship of antioxidative property of flavonoids and inhibitory effect on matrix metalloproteinase activity in UVA-irradiated human dermal fibroblast. Arch. Pharm. Res. 2007, 30, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Gendaram, O.; Choi, Y.H.; Kim, Y.S.; Ryu, S.Y. Anti-oxidative and antibacterial constituents from Sedum hybridum. Nat. Prod. Sci. 2011, 17, 279–284. [Google Scholar]
- Chaabi, M.; Beghidja, N.; Benayache, S.; Lobstein, A. Activity-guided isolation of antioxidant principles from Limoniastrum feei (Girard) Batt. Z. Naturforsch. C 2008, 63, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Mahjoub, M.A.; Ammar, S.; Edziri, H.; Mighri, N.; Bouraoui, A.; Mighri, Z. Anti-inflammatory and antioxidant activities of some extracts and pure natural products isolated from Rhus tripartitum (Ucria). Med. Chem. Res. 2010, 19, 271–282. [Google Scholar] [CrossRef]
- Yoshimura, M.; Yamakami, S.; Amakura, Y.; Yoshida, T. Diarylheptanoid sulfates and related compounds from Myrica rubra bark. J. Nat. Prod. 2012, 75, 1798–1802. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.B.; Rizvi, S.I. Ferric reducing and radical scavenging activities of selected important polyphenols present in foods. Int. J. Food Prop. 2012, 15, 702–708. [Google Scholar] [CrossRef]
- Ratty, A.K.; Das, N.P. Effects of flavonoids on nonenzymatic lipid peroxidation: Structure-activity relationship. Biochem. Med. Metab. Biol. 1988, 39, 69–79. [Google Scholar] [CrossRef]
- Zhang, D.; Xie, L.; Jia, G.; Cai, S.; Ji, B.; Liu, Y.; Wu, W.; Zhou, F.; Wang, A.; Chu, L.; et al. Comparative study on antioxidant capacity of flavonoids and their inhibitory effects on oleic acid-induced hepatic steatosis in vitro. Eur. J. Med. Chem. 2011, 46, 4548–4558. [Google Scholar] [CrossRef] [PubMed]
- Rostoka, E.; Baumane, L.; Isajevs, S.; Line, A.; Dzintare, M.; Svirina, D.; Sharipova, J.; Silina, K.; Kalvinsh, I.; Sjakste, N. Effects of kaempferol and myricetin on inducible nitric oxide synthase expression and nitric oxide production in rats. Basic Clin. Pharmacol. Toxicol. 2010, 106, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Sueishi, Y.; Hori, M. Nitric oxide scavenging rates of solubilized resveratrol and flavonoids. Nitric Oxide 2013, 29, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Valdez, L.B.; Álvarez, S.; Zaobornyj, T.; Boveris, A. Polyphenols and red wine as antioxidants against peroxynitrite and other oxidants. Biol. Res. 2004, 37, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, M.; Garcia-Carmona, F. Myricetin, an antioxidant flavonol, is a substrate of polyphenol oxidase. J. Sci. Food Agric. 1999, 79, 1993–2000. [Google Scholar] [CrossRef]
- Rusak, G.; Gutzeit, H.O.; Müller, J.L. Structurally related flavonoids with antioxidative properties differentially affect cell cycle progression and apoptosis of human acute leukemia cells. Nutr. Res. 2005, 25, 143–155. [Google Scholar] [CrossRef]
- Husain, S.R.; Cillard, J.; Cillard, P. Hydroxyl radical scavenging activity of flavonoids. Phytochemistry 1987, 26, 2489–2491. [Google Scholar] [CrossRef]
- Soucek, P.; Kondrová, E.; Heřmánek, J.; Stopka, P.; Boumendjel, A.; Ueng, Y.F.; Gut, I. New model system for testing effects of flavonoids on doxorubicin-related formation of hydroxyl radicals. Anticancer Drugs 2011, 22, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Sofie, E.; Prior, R.L. Peroxyl radical absorbing antioxidant activities of flavonoids. FASEB J. 1996, 10, A821. [Google Scholar]
- Ah Kang, K.; Wang, Z.H.; Zhang, R.; Piao, M.J.; Kim, K.C.; Kang, S.S.; Kim, Y.W.; Lee, J.; Park, D.; Hyun, J.W. Myricetin protects cells against oxidative stress-induced apoptosis via regulation of PI3K/Akt and MAPK signaling pathways. Int. J. Mol. Sci. 2010, 11, 4348–4360. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.H.; Ah Kang, K.; Zhang, R.; Piao, M.J.; Jo, S.H.; Kim, J.S.; Kang, S.S.; Lee, J.S.; Park, D.H.; Hyun, J.W. Myricetin suppresses oxidative stress-induced cell damage via both direct and indirect antioxidant action. Environ. Toxicol. Pharmacol. 2010, 29, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Duthie, S.J.; Collins, A.R.; Duthie, G.G.; Dobson, V.L. Quercetin and myricetin protect against hydrogen peroxide-induced DNA damage (strand breaks and oxidised pyrimidines) in human lymphocytes. Mutat. Res. 1997, 393, 223–231. [Google Scholar] [CrossRef]
- Duthie, S.J.; Dobson, V.L. Dietary flavonoids protect human colonocyte DNA from oxidative attack in vitro. Eur. J. Nutr. 1999, 38, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Miyajima, Y.; Kikuzaki, H.; Hisamoto, M.; Nakatani, N. Antioxidative polyphenols from berries of Pimenta dioica. Biofactors 2004, 22, 301–303. [Google Scholar] [CrossRef]
- Sahu, S.C.; Gray, G.C. Interactions of flavonoids, trace metals, and oxygen: Nuclear DNA damage and lipid peroxidation induced by myricetin. Cancer Lett. 1993, 70, 73–79. [Google Scholar] [CrossRef]
- Abalea, V.; Cillard, J.; Dubos, M.P.; Sergent, O.; Cillard, P.; Morel, I. Repair of iron-induced DNA oxidation by the flavonoid myricetin in primary rat hepatocyte cultures. Free Radic. Biol. Med. 1999, 26, 1457–1466. [Google Scholar] [CrossRef]
- Morel, I.; Abaléa, V.; Sergent, O.; Cillard, P.; Cillard, J. Involvement of phenoxyl radical intermediates in lipid antioxidant action of myricetin in iron-treated rat hepatocyte culture. Biochem. Pharmacol. 1998, 55, 1399–1404. [Google Scholar] [CrossRef]
- Bennett, C.J.; Caldwell, S.T.; McPhail, D.B.; Morrice, P.C.; Duthie, G.G.; Hartley, R.C. Potential therapeutic antioxidants that combine the radical scavenging ability of myricetin and the lipophilic chain of vitamin E to effectively inhibit microsomal lipid peroxidation. Bioorg. Med. Chem. 2004, 12, 2079–2098. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Li, Y.; Li, J.; Han, Q.; Ye, L.; Li, A. Myricetin affords protection against peroxynitrite-mediated DNA damage and hydroxyl radical formation. Food Chem. Toxicol. 2011, 49, 2439–2444. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.S.; Kim, A.K. Effect of myricetin combined with vitamin C or vitamin E on antioxidant enzyme system in murine melanoma cells. Korean J. Pharmacogn. 2004, 35, 357–363. [Google Scholar]
- Do Nascimento, A.J.; Henneberg, R.; Otuki, M.F.; Furman, A.E.F.; Hermann, P.; Leonart, M.S.S. Protective effect of favonoids against reactive oxygen species production in sickle cell anemia patients treated with hydroxyurea. Rev. Bras. Hematol. Hemoter. 2013, 35, 52–55. [Google Scholar]
- Robak, J.; Gryglewski, R.J. Flavonoids are scavengers of superoxide anions. Biochem. Pharmacol. 1988, 37, 837–841. [Google Scholar] [CrossRef]
- Costantino, L.; Rastelli, G.; Albasini, A. Inhibitory activity of flavonols towards the xanthine oxidase enzyme. Int. J. Pharm. 1992, 86, 17–23. [Google Scholar] [CrossRef]
- Qin, S.; Chen, J.; Tanigawa, S.; Hou, D.X. Microarray and pathway analysis highlight Nrf2/ARE-mediated expression profiling by polyphenolic myricetin. Mol. Nutr. Food Res. 2013, 57, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhang, X. Comparisons of cytoprotective effects of three flavonoids against human hepatocytes oxidative injury induced by hydrogen peroxide or carbon tetrachloride in vitro. J. Med. Plants Res. 2009, 3, 776–784. [Google Scholar]
- Laughton, M.J.; Halliwell, B.; Evans, P.J.; Hoult, J.R.S. Antioxidant and pro-oxidant actions of the plant phenolics quercetin, gossypol and myricetin. Effects on lipid peroxidation, hydroxyl radical generation and bleomycin-dependent damage to DNA. Biochem. Pharmacol. 1989, 38, 2859–2865. [Google Scholar] [CrossRef]
- Chobot, V.; Hadacek, F. Exploration of pro-oxidant and antioxidant activities of the flavonoid myricetin. Redox Rep. 2011, 16, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Duthie, G.; Morrice, P. Antioxidant capacity of flavonoids in hepatic microsomes is not reflected by antioxidant effects in vivo. Oxid. Med. Cell. Longev. 2012, 2012, 165127. [Google Scholar] [CrossRef] [PubMed]
- Fraga, C.G.; Martino, V.S.; Ferraro, G.E.; Coussio, J.D.; Boveris, A. Flavonoids as antioxidants evaluated by in vitro and in situ liver chemiluminescence. Biochem. Pharmacol. 1987, 36, 717–720. [Google Scholar] [CrossRef]
- Pandey, K.B.; Mishra, N.; Rizvi, S.I. Myricetin may provide protection against oxidative stress in type 2 diabetic erythrocytes. Z. Naturforsch. C 2009, 64, 626–630. [Google Scholar] [CrossRef] [PubMed]
- Oyama, Y.; Fuchs, P.A.; Katayama, N.; Noda, K. Myricetin and quercetin, the flavonoid constituents of Ginkgo biloba extract, greatly reduce oxidative metabolism in both resting and Ca2+-loaded brain neurons. Brain Res. 1994, 635, 125–129. [Google Scholar] [CrossRef]
- Teissedre, P.L.; Frankel, E.N.; Waterhouse, A.L.; Peleg, H.; German, J.B. Inhibition of in vitro human LDL oxidation by phenolic antioxidants from grapes and wines. J. Sci. Food Agric. 1996, 70, 55–61. [Google Scholar] [CrossRef]
- Sadasivam, K.; Kumaresan, R. Antioxidant behavior of mearnsetin and myricetin flavonoid compounds—A DFT study. Spectrochim. Acta A 2011, 79, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.J.; Mou, W.S.; Lin, F.R.; Xu, J.H.; Lei, Q.F.; Fang, W.J. Radical scavenging activity of myricetin. Acta PhysicoChim. Sin. 2013, 29, 1421–1432. [Google Scholar]
- Wang, L.; Tu, Y.C.; Lian, T.W.; Hung, J.T.; Yen, J.H.; Wu, M.J. Distinctive antioxidant and antiinflammatory effects of flavonols. J. Agric. Food Chem. 2006, 54, 9798–9804. [Google Scholar] [CrossRef] [PubMed]
- Mira, L.; Fernandez, M.T.; Santos, M.; Rocha, R.; Florêncio, M.H.; Jennings, K.R. Interactions of flavonoids with iron and copper ions: A mechanism for their antioxidant activity. Free Radic. Res. 2002, 36, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Justino, G.C.; Vieira, A.J.S.C. Antioxidant mechanisms of quercetin and myricetin in the gas phase and in solution- A comparison and validation of semi-empirical methods. J. Mol. Model. 2010, 16, 863–876. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.K.; Lee, K.W.; Kim, H.Y.; Oh, M.H.; Byun, S.; Lim, S.H.; Heo, Y.S.; Kang, N.J.; Bode, A.M.; Dong, Z.; et al. Myricetin suppresses UVB-induced wrinkle formation and MMP-9 expression by inhibiting Raf. Biochem. Pharmacol. 2010, 79, 1455–1461. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.H.; Huang, C.C.; Fang, J.Y.; Yang, C.; Chan, C.M.; Wu, N.L.; Kang, S.W.; Hung, C.F. Protective effects of myricetin against ultraviolet-B-induced damage in human keratinocytes. Toxicol. Vitro 2010, 24, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Sung, K.J.; Ki, W.L.; Byun, S.; Nam, J.K.; Sung, H.L.; Heo, Y.S.; Bode, A.M.; Bowden, G.T.; Hyong, J.L.; Dong, Z. Myricetin suppresses UVB-induced skin cancer by targeting Fyn. Cancer Res. 2008, 68, 6021–6029. [Google Scholar]
- Kim, W.; Yang, J.H.; Youn, H.; Yun, Y.J.; Seong, K.M.; Youn, B. Myricetin inhibits Akt survival signaling and induces bad-mediated apoptosis in a low dose ultraviolet (UV)-B-irradiated hacat human immortalized keratinocytes. J. Radiat. Res. 2010, 51, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Kumamoto, T.; Fujii, M.; Hou, D.X. Akt is a direct target for myricetin to inhibit cell transformation. Mol. Cell. Biochem. 2009, 332, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Mi, M.T.; Gu, Y.Y.; Yuan, J.L.; Ling, W.H.; Lin, H. Effects of flavonoids with different structures on proliferation of leukemia cell line HL-60. Chin. J. Cancer 2007, 26, 1309–1314. [Google Scholar]
- Romanouskaya, T.V.; Grinev, V.V. Cytotoxic effect of flavonoids on leukemia cells and normal cells of human blood. Bull. Exp. Biol. Med. 2009, 148, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Lazaro, M.L.; Willmore, E.; Austin, C.A. The dietary flavonoids myricetin and fisetin act as dual inhibitors of DNA topoisomerases I and II in cells. Mutat. Res. 2010, 696, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Constantinou, A.; Mehta, R.; Runyan, C.; Rao, K.; Vaughan, A.; Moon, R. Flavonoids as DNA topoisomerase antagonists and poisons: Structure-activity relationships. J. Nat. Prod. 1995, 58, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Kang, N.J.; Jung, S.K.; Lee, K.W.; Lee, H.J. Myricetin is a potent chemopreventive phytochemical in skin carcinogenesis. Ann. N. Y. Acad. Sci. 2011, 1229, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Kang, N.J.; Rogozin, E.A.; Kim, H.-G.; Cho, Y.Y.; Bode, A.M.; Lee, H.J.; Surh, Y.J.; Bowden, G.T.; Dong, Z. Myricetin is a novel natural inhibitor of neoplastic cell transformation and MEK1. Carcinogenesis 2007, 28, 1918–1927. [Google Scholar] [CrossRef] [PubMed]
- Kumamoto, T.; Fujii, M.; Hou, D.X. Myricetin directly targets JAK1 to inhibit cell transformation. Cancer Lett. 2009, 275, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Ichimatsu, D.; Nomura, M.; Nakamura, S.; Moritani, S.; Yokogawa, K.; Kobayashi, S.; Nishioka, T.; Miyamoto, K.I. Structure-activity relationship of flavonoids for inhibition of epidermal growth factor-induced transformation of JB6 CI 41 cells. Mol. Carcinog. 2007, 46, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Siegelin, M.D.; Gaiser, T.; Habel, A.; Siegelin, Y. Myricetin sensitizes malignant glioma cells to TRAIL-mediated apoptosis by down-regulation of the short isoform of FLIP and bcl-2. Cancer Lett. 2009, 283, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, M.R.; Said, A.; Tundis, R.; Hawas, U.W.; Rashed, K.; Menichini, F.; Frega, N.G.; Menichini, F. Antioxidant and antiproliferative activity of Diospyros lotus L. extract and isolated compounds. Plant Foods Hum. Nutr. 2009, 64, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, E.H.; Grant, M.H. The effect of the flavonoids, quercetin, myricetin and epicatechin on the growth and enzyme activities of MCF7 human breast cancer cells. Chem. Biol. Interact. 1998, 116, 213–228. [Google Scholar] [CrossRef]
- Mori, A.; Nishino, C.; Enoki, N.; Tawata, S. Cytotoxicity of plant flavanoids against HeLa cells. Phytochemistry 1988, 27, 1017–1020. [Google Scholar] [CrossRef]
- Alonso, S.G.; Collins, V.J.; Vauzour, D.; Mateos, A.R.; Corona, G.; Spencer, J.P.E. Inhibition of colon adenocarcinoma cell proliferation by flavonols is linked to a G2/M cell cycle block and reduction in cyclin D1 expression. Food Chem. 2012, 130, 493–500. [Google Scholar] [CrossRef]
- Shiomi, K.; Kuriyama, I.; Yoshida, H.; Mizushina, Y. Inhibitory effects of myricetin on mammalian DNA polymerase, topoisomerase and human cancer cell proliferation. Food Chem. 2013, 139, 910–918. [Google Scholar] [CrossRef] [PubMed]
- Ko, C.H.; Shen, S.C.; Lee, T.J.F.; Chen, Y.C. Myricetin inhibits matrix metalloproteinase 2 protein expression and enzyme activity in colorectal carcinoma cells. Mol. Cancer Ther. 2005, 4, 281–290. [Google Scholar] [PubMed]
- Morales, P.; Haza, A.I. Selective apoptotic effects of piceatannol and myricetin in human cancer cells. J. Appl. Toxicol. 2012, 32, 986–993. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, A.; Willett, K.L. Inhibition of human cytochrome CYP1 enzymes by flavonoids of St. John’s wort. Toxicology 2006, 217, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Master, Z.; Chaudhary, A.; Sutter, T.R.; Willett, K.L. Effects of flavonoids on CYP1 expression in RL95-2 endometrial carcinoma cells. Food Chem. 2012, 133, 912–922. [Google Scholar] [CrossRef]
- Lu, J.; Papp, L.V.; Fang, J.; Rodriguez-Nieto, S.; Zhivotovsky, B.; Holmgren, A. Inhibition of mammalian thioredoxin reductase by some flavonoids: Implications for myricetin and quercetin anticancer activity. Cancer Res. 2006, 66, 4410–4418. [Google Scholar] [CrossRef] [PubMed]
- Agullo, G.; Gamet-Payrastre, L.; Manenti, S.; Viala, C.; Rémésy, C.; Chap, H.; Payrastre, B. Relationship between flavonoid structure and inhibition of phosphatidylinositol 3-kinase: A comparison with tyrosine kinase and protein kinase C inhibition. Biochem. Pharmacol. 1997, 53, 1649–1657. [Google Scholar] [CrossRef]
- Walker, E.H.; Pacold, M.E.; Perisic, O.; Stephens, L.; Hawkins, P.T.; Wymann, M.P.; Williams, R.L. Structural determinants of phosphoinositide 3-kinase inhibition by wortmannin, LY294002, quercetin, myricetin, and staurosporine. Mol. Cell 2000, 6, 909–919. [Google Scholar] [CrossRef]
- Yuan, C.H.; Filippova, M.; Tungteakkhun, S.S.; Duerksen-Hughes, P.J.; Krstenansky, J.L. Small molecule inhibitors of the HPV16-E6 interaction with caspase 8. Bioorg. Med. Chem. Lett. 2012, 22, 2125–2129. [Google Scholar] [CrossRef] [PubMed]
- Bin, H.S.; Choi, U.K. Myricetin inhibits adipogenesis in human adipose tissue-derived mesenchymal stem cells. Food Sci. Biotechnol. 2012, 21, 1391–1396. [Google Scholar] [CrossRef]
- Van Zanden, J.J.; de Mul, A.; Wortelboer, H.M.; Usta, M.; van Bladeren, P.J.; Rietjens, I.M.C.M.; Cnubben, N.H.P. Reversal of in vitro cellular MRP1 and MRP2 mediated vincristine resistance by the flavonoid myricetin. Biochem. Pharmacol. 2005, 69, 1657–1665. [Google Scholar] [CrossRef] [PubMed]
- Durgo, K.; Vuković, L.; Rusak, G.; Osmak, M.; Čolić, J.F. Effect of flavonoids on glutathione level, lipid peroxidation and cytochrome P450 CYP1A1 expression in human laryngeal carcinoma cell lines. Food Technol. Biotechnol. 2007, 45, 69–79. [Google Scholar]
- Xu, R.; Zhang, Y.; Ye, X.; Xue, S.; Shi, J.; Pan, J.; Chen, Q. Inhibition effects and induction of apoptosis of flavonoids on the prostate cancer cell line PC-3 in vitro. Food Chem. 2013, 138, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Zheng, X.Y.; Ye, J.; Wu, T.T.; Wang, J.L.; Chen, W. Potential anticancer activity of myricetin in human T24 bladder cancer cells both in vitro and in vivo. Nutr. Cancer 2012, 64, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ling, Y.; Yu, H.; Ji, Y. Studies on mechanism of myricetin-induced apoptosis in human hepatocellular carcinoma HepG-2 cells. Zhongguo Zhongyao Zazhi 2010, 35, 1046–1050. [Google Scholar] [PubMed]
- Zhang, Q.; Zhao, X.H.; Wang, Z.J. Flavones and flavonols exert cytotoxic effects on a human oesophageal adenocarcinoma cell line (OE33) by causing G2/M arrest and inducing apoptosis. Food Chem. Toxicol. 2008, 46, 2042–2053. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhao, X.H.; Wang, Z.J. Cytotoxicity of flavones and flavonols to a human esophageal squamous cell carcinoma cell line (KYSE-510) by induction of G2/M arrest and apoptosis. Toxicol. Vitro 2009, 23, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhao, X.H. Flavone-and flavonol-caused apoptosis in human esophageal cancer cells by inducing the expression of PIG3. Prog. Biochem. Biophys. 2009, 36, 213–219. [Google Scholar] [CrossRef]
- Garcia, O.B.; Castillo, J.; Lorente, J.; Alcaraz, M.; Yanez, J.; Martinez, C.; Vicente, V.; Lozano, J.A. Antiproliferative activity of several phenolic compounds against melanoma cell lines: Relationship between structure and activity. Agro Food Ind. Hi-Tech 2005, 16, 30–34. [Google Scholar]
- Labbe, D.; Provencal, M.; Lamy, S.; Boivin, D.; Gingras, D.; Béliveau, R. The flavonols quercetin, kaempferol, and myricetin inhibit hepatocyte growth factor-induced medulloblastoma cell migration. J. Nutr. 2009, 139, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Shih, Y.W.; Wu, P.F.; Lee, Y.C.; Shi, M.D.; Chiang, T.A. Myricetin suppresses invasion and migration of human lung adenocarcinoma a549 cells: Possible mediation by blocking the erk signaling pathway. J. Agric. Food Chem. 2009, 57, 3490–3499. [Google Scholar] [CrossRef] [PubMed]
- Phillips, P.A.; Sangwan, V.; Borja-Cacho, D.; Dudeja, V.; Vickers, S.M.; Saluja, A.K. Myricetin induces pancreatic cancer cell death via the induction of apoptosis and inhibition of the phosphatidylinositol 3-kinase (PI3K) signaling pathway. Cancer Lett. 2011, 308, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Androutsopoulos, V.P.; Papakyriakou, A.; Vourloumis, D.; Spandidos, D.A. Comparative CYP1A1 and CYP1B1 substrate and inhibitor profile of dietary flavonoids. Bioorg. Med. Chem. 2011, 19, 2842–2849. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.E.; Ha, T.K.; Yoon, J.H.; Lee, J.S. Myricetin induces cell death of human colon cancer cells via BAX/BCL2-dependent pathway. Anticancer Res. 2014, 34, 701–706. [Google Scholar] [PubMed]
- Schutte, M.E.; van de Sandt, J.J.M.; Alink, G.M.; Groten, J.P.; Rietjens, I.M.C.M. Myricetin stimulates the absorption of the pro-carcinogen PhIP. Cancer Lett. 2006, 231, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.; Dobrzyńska, M.M.; Yu, T.W. Modulating effects of Silymarin and myricetin on food mutagens and doxorubicin in assays with different genetic endpoints. J. Environ. Pathol. Toxicol. Oncol. 1997, 16, 313–327. [Google Scholar]
- Tzeng, S.H.; Ko, W.C.; Ko, F.N.; Teng, C.M. Inhibition of platelet aggregation by some flavonoids. Thromb. Res. 1991, 64, 91–100. [Google Scholar] [CrossRef]
- Zang, B.X.; Jin, M.; Wu, W.; Chen, W.M.; Piao, Y.Z.; Li, J.R. Antagonistic effect of myricetin on platelet activing factor. Yaoxue Xuebao 2003, 38, 831–833. [Google Scholar]
- Melzig, M.F.; Henke, K. Inhibition of thrombin activity by selected natural products in comparison to neutrophil elastase. Planta Med. 2005, 71, 787–789. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.M.; Jin, M.; Wu, W.; Li, J.R.; Zang, B.X.; Piao, Y.Z. Inhibitory effect of flavonols of Carthamus tinctorius L. against rabbit platelet activation induced by platelet activating factor. Yaoxue Xuebao 2001, 36, 884–885. [Google Scholar]
- Liu, L.; Ma, H.; Yang, N.; Tang, Y.; Guo, J.; Tao, W.; Duan, J. A series of natural flavonoids as thrombin inhibitors: Structure-activity relationships. Thromb. Res. 2010, 126, e365–e378. [Google Scholar] [CrossRef] [PubMed]
- Landolfi, R.; Mower, R.L.; Steiner, M. Modification of platelet function and arachidonic acid metabolism by bioflavonoids. Structure-activity relations. Biochem. Pharmacol. 1984, 33, 1525–1530. [Google Scholar] [CrossRef]
- Robak, J.; Korbut, R.; Shridi, F.; Swies, J.; Rzadkowska-Bodalska, H. On the mechnaism of antiaggregatory effects of myricetin. Pol. J. Pharmacol. Pharm. 1988, 40, 337–340. [Google Scholar] [PubMed]
- Tong, Y.; Zhou, X.M.; Wang, S.J.; Yang, Y.; Cao, Y.L. Analgesic activity of myricetin isolated from Myrica rubra Sieb. et Zucc. Leaves. Arch. Pharm. Res. 2009, 32, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Borde, P.; Mohan, M.; Kasture, S. Effect of myricetin on deoxycorticosterone acetate (DOCA)-salt-hypertensive rats. Nat. Prod. Res. 2011, 25, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Godse, S.; Mohan, M.; Kasture, V.; Kasture, S. Effect of myricetin on blood pressure and metabolic alterations in fructose hypertensive rats. Pharm. Biol. 2010, 48, 494–498. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.Y.; Kim, S.H.; Cho, D.; Kim, T.S. Inhibition of interleukin-12 production in mouse macrophages via decreased nuclear factor-kappaB DNA binding activity by myricetin, a naturally occurring flavonoid. Arch. Pharm. Res. 2005, 28, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, R.; Andriambeloson, E.; Duarte, J.; Andriantsitohaina, R.; Jiménez, J.; Pérez-Vizcaino, F.; Zarzuelo, A.; Tamargo, J. Involvement of thromboxane A2 in the endothelium-dependent contractions induced by myricetin in rat isolated aorta. Br. J. Pharmacol. 1999, 127, 1539–1544. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.C.; Yoon, G.; Kwang, Y.L.; Hyun, J.C.; Bok, Y.K. Inhibition of interleukin-2 production by myricetin in mouse EL-4 T cells. Arch. Pharm. Res. 2007, 30, 1075–1079. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zeng, Y.Y.; Liu, L.; Ji, Y.H.; Xing, F.Y.; Zhao, J.X. The effects of myrictin on inhibition of mouse lymphocytes activated in vitro. Chin. Pharmacol. Bull. 2006, 22, 63–66. [Google Scholar]
- Grenier, D.; Chen, H.; Lagha, A.B.; Fournier-Larente, J.; Morin, M.P. Dual action of myricetin on Porphyromonas gingivalis and the inflammatory response of host cells: A promising therapeutic molecule for periodontal diseases. PLoS ONE 2015, 10, e0131758. [Google Scholar]
- Ishikawa, Y.T.; Goto, M.; Yamaki, K. Structure-activity relations of inhibitory effects of various flavonoids on lipopolysaccharide-induced prostaglandin E2 production in rat peritoneal macrophages: Comparison between subclasses of flavonoids. Phytomedicine 2006, 13, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.H.; Kim, D.H.; Kim, M.H.; Oh, M.H.; Kim, S.R.; Park, K.J.; Lee, M.W. Flavonoid constituents in the leaves of Myrica rubra sieb. et zucc. with anti-inflammatory activity. Arch. Pharm. Res. 2013, 36, 1533–1540. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.M.; Nam, J.K.; Jin, H.H.; Ki, W.L.; Hyong, J.L. Myricetin down-regulates phorbol ester-induced cyclooxygenase-2 expression in mouse epidermal cells by blocking activation of nuclear factor kappa B. J. Agric. Food Chem. 2007, 55, 9678–9684. [Google Scholar] [CrossRef] [PubMed]
- Venegas, G.G.; Luna, O.A.; Arroyo, J.A.V.; Bermúdez, C.H. Myricetin suppresses lipoteichoic acid-induced interleukin-1β and cyclooxygenase-2 expression in human gingival fibroblasts. Microbiol. Immunol. 2013, 57, 849–856. [Google Scholar] [CrossRef] [PubMed]
- Venegas, G.G.; Luna, O.A.; Cano, J.A.A.; Bermúdez, C.H. Myricetin blocks lipoteichoic acid-induced COX-2 expression in human gingival fibroblasts. Cell. Mol. Biol. Lett. 2014, 19, 126–139. [Google Scholar]
- Ko, S.Y. Myricetin suppresses LPS-induced MMP expression in human gingival fibroblasts and inhibits osteoclastogenesis by downregulating NFATc1 in RANKL-induced RAW 264.7 cells. Arch. Oral Biol. 2012, 57, 1623–1632. [Google Scholar] [CrossRef] [PubMed]
- Kuo, P.L. Myricetin inhibits the induction of anti-Fas IgM-, tumor necrosis factor-α- and interleukin-1 β-mediated apoptosis by Fas pathway inhibition in human osteoblastic cell line MG-63. Life Sci. 2005, 77, 2964–2976. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Choi, E.M. Myricetin inhibits IL-1β-induced inflammatory mediators in SW982 human synovial sarcoma cells. Int. Immunopharmacol. 2010, 10, 812–814. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Hong, T.; Dong, M.; Meng, Y.; Mu, J. Protective effect of myricetin in dextran sulphate sodium-induced murine ulcerative colitis. Mol. Med. Rep. 2013, 7, 565–570. [Google Scholar] [PubMed]
- Bai, H.W.; Zhu, B.T. Myricetin and quercetin are naturally occurring co-substrates of cyclooxygenases in vivo. Prostaglandins Leukot. Essent. Fatty Acids 2010, 82, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, K.C.P.; Figueiredo, C.A.V.; Figueredo, T.B.; Freire, K.R.L.; Santos, F.A.R.; Neves, N.M.A.; Silva, T.M.S.; Piuvezam, M.R. Anti-allergic effect of bee pollen phenolic extract and myricetin in ovalbumin-sensitized mice. J. Ethnopharmacol. 2008, 119, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Park, H.H.; Lee, S.; Son, H.Y.; Park, S.B.; Kim, M.S.; Choi, E.J.; Singh, T.S.K.; Ha, J.H.; Lee, M.G.; Kim, J.E.; et al. Flavonoids inhibit histamine release and expression of proinflammatory cytokines in mast cells. Arch. Pharm. Res. 2008, 31, 1303–1311. [Google Scholar] [CrossRef] [PubMed]
- Hagenacker, T.; Hillebrand, I.; Wissmann, A.; Büsselberg, D.; Schäfers, M. Anti-allodynic effect of the flavonoid myricetin in a rat model of neuropathic pain: Involvement of p38 and protein kinase C mediated modulation of Ca2+ channels. Eur. J. Pain 2010, 14, 992–998. [Google Scholar] [CrossRef] [PubMed]
- Hagenacker, T.; Hillebrand, I.; Büsselberg, D.; Schäfers, M. Myricetin reduces voltage activated potassium channel currents in DRG neurons by a p38 dependent mechanism. Brain Res. Bull. 2010, 83, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Cordova, M.M.; Werner, M.F.D.P.; Silva, M.D.D.; Ruani, A.P.; Pizzolatti, M.G.; Santos, A.R.S. Further antinociceptive effects of myricitrin in chemical models of overt nociception in mice. Neurosci. Lett. 2011, 495, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Hamamoto, R.; Uchiyama, S.; Ishiyama, K. Effects of flavonoid on calcium content in femoral tissue culture and parathyroid hormone-stimulated osteoclastogenesis in bone marrow culture in vitro. Mol. Cell. Biochem. 2007, 303, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.L.; Chang, J.K.; Tsai, C.H.; Chien, T.T.C.; Kuo, P.L. Myricetin induces human osteoblast differentiation through bone morphogenetic protein-2/p38 mitogen-activated protein kinase pathway. Biochem. Pharmacol. 2007, 73, 504–514. [Google Scholar] [CrossRef] [PubMed]
- Hyman, B.T.; van Hoesen, G.W.; Damasio, A.R.; Barnes, C.L. Alzheimer’s disease: Cell-specific pathology isolates the hippocam-pal formation. Science 1984, 225, 1168–1170. [Google Scholar] [CrossRef] [PubMed]
- Kanaan, N.M.; Himmelstein, D.S.; Ward, S.M.; Combs, B.; Binde, L.I. Tau Protein: Biology and Pathobiology. In Movement Disorders, 2nd ed.; Academic Press: New York, NY, USA, 2015; pp. 857–874. [Google Scholar]
- Duyckaerts, C.; Delatour, B.; Potier, M.C. Classification and basic pathology of Alzheimer disease. Acta Neuropathol. 2009, 118, 5–36. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Yoshiike, Y.; Takashima, A.; Hasegawa, K.; Naiki, H.; Yamada, M. Potent anti-amyloidogenic and fibril-destabilizing effects of polyphenols in vitro: Implications for the prevention and therapeutics of Alzheimer’s disease. J. Neurochem. 2003, 87, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Kim, T.D.; Paik, S.R.; Jeong, K.; Jung, S. Molecular simulations for anti-amyloidogenic effect of flavonoid myricetin exerted against Alzheimer’s β-amyloid fibrils formation. Bull. Korean Chem. Soc. 2008, 29, 1505–1509. [Google Scholar]
- Shimmyo, Y.; Kihara, T.; Akaike, A.; Niidome, T.; Sugimoto, H. Multifunction of myricetin on Aβ: Neuroprotection via a conformational change of Aβ and reduction of Aβ via the interference of secretases. J. Neurosci. Res. 2008, 86, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Chang, C.Y.; Wang, S.J.; Huang, S.K. Myricetin inhibits the release of glutamate in rat cerebrocortical nerve terminals. J. Med. Food 2015, 18, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Shimmyo, Y.; Kihara, T.; Akaike, A.; Niidome, T.; Sugimoto, H. Three distinct neuroprotective functions of myricetin against glutamate-induced neuronal cell death: Involvement of direct inhibition of caspase-3. J. Neurosci. Res. 2008, 86, 1836–1845. [Google Scholar] [CrossRef] [PubMed]
- Tucci, P.; Mhillaj, E.; Morgese, M.G.; Colaianna, M.; Zotti, M.; Schiavone, S.; Cicerale, M.; Trezza, V.; Campolongo, P.; Cuomo, V.; et al. Memantine prevents memory consolidation failure induced by soluble beta amyloid in rats. Front. Behav. Neurosci. 2014, 8, 332. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Yamada, M. Antioxidant compounds have potent anti-fibrillogenic and fibril-destabilizing effects for α-synuclein fibrils in vitro. J. Neurochem. 2006, 97, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Caruana, M.; Högen, T.; Levin, J.; Hillmer, A.; Giese, A.; Vassallo, N. Inhibition and disaggregation of α-synuclein oligomers by natural polyphenolic compounds. FEBS Lett. 2011, 585, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Jia, Y.H. Inhibition of catechol-o-methyltransferase (COMT) by myricetin, dihydromyricetin and myricitrin. Pharmazie 2014, 69, 183–186. [Google Scholar] [PubMed]
- Jimenez, M.F.M.; Reus, M.I.S.; Andres, D.; Cascales, M.; Benedi, J. Neuroprotective effect of fraxetin and myricetin against rotenone-induced apoptosis in neuroblastoma cells. Brain Res. 2004, 1009, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Laabich, A.; Manmoto, C.C.; Kuksa, V.; Leung, D.W.; Vissvesvaran, G.P.; Karliga, I.; Kamat, M.; Scott, I.L.; Fawzi, A.; Kubota, R. Protective effects of myricetin and related flavonols against A2E and light mediated-cell death in bovine retinal primary cell culture. Exp. Eye Res. 2007, 85, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Chen, J.; Zhang, W.; Fu, W.; Wu, G.; Wei, H.; Wang, Q.; Ruan, J. In vivo investigation on the potential of galangin, kaempferol and myricetin for protection of d-galactose-induced cognitive impairment. Food Chem. 2012, 135, 2702–2707. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.N.; Wang, W.; Ding, Y.; Xie, W.D.; Ma, C.; Du, L.J. A new flavonol glycoside and activity of compounds from the flower of Nymphaea candida. J. Asian Nat. Prod. Res. 2007, 9, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Mohan, M.; Jadhav, S.S.; Kasture, V.S.; Kasture, S.B. Effect of myricetin on behavioral paradigms of anxiety. Pharm. Biol. 2009, 47, 927–931. [Google Scholar] [CrossRef]
- Moonrungsee, N.; Shimamura, T.; Kashiwagi, T.; Jakmunee, J.; Higuchi, K.; Ukeda, H. An automated sequential injection spectrophotometric method for evaluation of tyramine oxidase inhibitory activity of some flavonoids. Talanta 2014, 122, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.G.; Wang, J.; Jiang, H.; Liu, T.W.; Xie, J.X. Myricetin reduces 6-hydroxydopamine-induced dopamine neuron degeneration in rats. Neuroreport 2007, 18, 1181–1185. [Google Scholar] [CrossRef] [PubMed]
- Franco, J.L.; Posser, T.; Missau, F.; Pizzolatti, M.G.; Santos, A.R.S.; Souza, D.O.; Aschner, M.; Rocha, J.B.T.; Dafre, A.L.; Farina, M. Structure-activity relationship of flavonoids derived from medicinal plants in preventing methylmercury-induced mitochondrial dysfunction. Environ. Toxicol. Pharmacol. 2010, 30, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Ma, Z.; Wang, J.; Xie, A.; Xie, J. Myricetin attenuated MPP+-induced cytotoxicity by anti-oxidation and inhibition of MKK4 and JNK activation in MES23.5 cells. Neuropharmacology 2011, 61, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Ma, Z.G.; Rowlands, D.K.; Gou, Y.L.; Fok, K.L.; Wong, H.Y.; Yu, M.K.; Tsang, L.L.; Mu, L.; Chen, L.; et al. Flavonoid myricetin modulates GABAA receptor activity through activation of Ca2+ channels and CaMK-II pathway. Evid. Based Complement. Altern. Med. 2012, 2012, 758097. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Choi, J.K.; Jung, C.H.; Koh, H.J.; Heo, P.; Shin, J.Y.; Kim, S.; Park, W.S.; Shin, H.J.; Kweon, D.H. SNARE-wedging polyphenols as small molecular botox. Planta Med. 2012, 78, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.G.; Liu, T.W. Myricetin facilitates potassium currents and inhibits neuronal activity of PVN neurons. Neurochem. Res. 2012, 37, 1450–1456. [Google Scholar] [CrossRef] [PubMed]
- Matic, S.; Stanic, S.; Bogojevic, D.; Vidakovic, M.; Grdovic, N.; Dinic, S.; Solujic, S.; Mladenovic, M.; Stankovic, N.; Mihailovic, M. Methanol extract from the stem of Cotinus coggygria Scop., and its major bioactive phytochemical constituent myricetin modulate pyrogallol-induced DNA damage and liver injury. Mutat. Res. 2013, 755, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Mo, S.F.; Zhou, F.; Lv, Y.Z.; Hu, Q.H.; Zhang, D.M.; Kong, L.D. Hypouricemic action of selected flavonoids in mice: Structure-activity relationships. Biol. Pharm. Bull. 2007, 30, 1551–1556. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Park, Y.S. Gene expression profiling of human umbilical vein endothelial cells exposed to myricetin. Biochip J. 2013, 7, 335–343. [Google Scholar] [CrossRef]
- Gan, C.L.; Liu, F.Z.; Jiang, S.X.; Cao, S.J.; Dong, N.W. Effects of five flavonols on [Ca2+]i in cardiomyocytes of rats. Chin. J. Epidemiol. 2007, 26, 624–626. [Google Scholar]
- Scarabelli, T.M.; Mariotto, S.; Azeim, S.A.; Shoji, K.; Darra, E.; Stephanou, A.; Scarabelli, C.C.; Marechal, J.D.; Knight, R.; Ciampa, A.; et al. Targeting STAT1 by myricetin and delphinidin provides efficient protection of the heart from ischemia/reperfusion-induced injury. FEBS Lett. 2009, 583, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.W.; Liu, F.Z.; Lu, J.K.; Liu, Y.C.; Yang, B.F. Effect of flavonols on caspase-3, Bcl-2 and bax expression in cardiomyocytes apoptosis. Chin. Pharm. J. 2007, 42, 749–753. [Google Scholar]
- Jimenez, R.; Zarzuelo, A.; Galisteo, M.; Duarte, J. Involvement of protein kinase C and Na+/K+-ATPase in the contractile response induced by myricetin in rat isolated aorta. Planta Med. 2002, 68, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, R.; Mohan, M.; Kasture, S.; Maxia, A.; Ballero, M. Cardioprotective potential of myricetin in isoproterenol-induced myocardial infarction in Wistar rats. Phytother. Res. 2009, 23, 1361–1366. [Google Scholar] [CrossRef] [PubMed]
- Lian, T.W.; Wang, L.; Lo, Y.H.; Huang, I.J.; Wu, M.J. Fisetin, morin and myricetin attenuate CD36 expression and oxLDL uptake in U937-derived macrophages. Biochim. Biophys. Acta 2008, 1781, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Angelone, T.; Pasqua, T.; di Majo, D.; Quintieri, A.M.; Filice, E.; Amodio, N.; Tota, B.; Giammanco, M.; Cerra, M.C. Distinct signalling mechanisms are involved in the dissimilar myocardial and coronary effects elicited by quercetin and myricetin, two red wine flavonols. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, G.; Khanna, A.K.; Sonkar, R.; Mishra, S.K.; Srivastava, S.; Lakshmi, V. Lipid lowering and antioxidant activity of flavones in triton treated hyperlipidemic rats. Med. Chem. Res. 2011, 20, 1622–1626. [Google Scholar] [CrossRef]
- Mohan, M.; Gupta, S.K.; Agnihotri, S.; Joshi, S.; Uppal, A.K. Anti-cataract effect of topical quercetin and myricetin in galactosemic rats. Med. Sci. Res. 1988, 16, 685–686. [Google Scholar]
- Hodges, L.C.; Kearse, E.C.; Green, K. Intraocular pressure-lowering activity of phenolic antioxidants in normotensive rabbits. Curr. Eye Res. 1999, 19, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Hollborn, M.; Grosche, A.; Reichenbach, A.; Wiedemann, P.; Bringmann, A.; Kohen, L. Effects of the vegetable polyphenols epigallocatechin-3-gallate, luteolin, apigenin, myricetin, quercetin, and cyanidin in primary cultures of human retinal pigment epithelial cells. Mol. Vis. 2014, 20, 242–258. [Google Scholar] [PubMed]
- Ong, K.C.; Khoo, H.E. Insulinomimetic effects of myricetin on lipogenesis and glucose transport in rat adipocytes but not glucose transporter translocation. Biochem. Pharmacol. 1996, 51, 423–429. [Google Scholar] [CrossRef]
- Ong, K.C.; Khoo, H.E. Effects of myricetin on glycemia and glycogen metabolism in diabetic rats. Life Sci. 2000, 67, 1695–1705. [Google Scholar] [CrossRef]
- Zelus, C.; Fox, A.; Calciano, A.; Faridian, B.S.; Nogaj, L.A.; Moffet, D.A. Myricetin inhibits islet amyloid polypeptide (IAPP) aggregation and rescues living mammalian cells from IAPP toxicity. Open Biochem. J. 2012, 6, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Liu, I.M.; Tzeng, T.F.; Liou, S.S. Myricetin ameliorates defective post-receptor insulin signaling via β-endorphin signaling in the skeletal muscles of fructose-fed rats. Evid. Based Complement. Altern. Med. 2011, 2011, 150752. [Google Scholar]
- Kang, S.J.; Park, J.H.Y.; Choi, H.N.; Kim, J.I. α-glucosidase inhibitory activities of myricetin in animal models of diabetes mellitus. Food Sci. Biotechnol. 2015, 24, 1897–1900. [Google Scholar] [CrossRef]
- Liu, I.M.; Tzeng, T.F.; Liou, S.S.; Lan, T.W. Improvement of insulin sensitivity in obese Zucker rats by myricetin extracted from Abelmoschus moschatus. Planta Med. 2007, 73, 1054–1060. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Dai, X.; Zhang, Z.; Li, Y. Myricetin attenuates hyperinsulinemia-induced insulin resistance in skeletal muscle cells. Eur. Food Res. Technol. 2012, 234, 873–881. [Google Scholar] [CrossRef]
- Lee, K.H.; Choi, E.M. Myricetin, a naturally occurring flavonoid, prevents 2-deoxy-d-ribose induced dysfunction and oxidative damage in osteoblastic MC3T3-E1 cells. Eur. J. Pharmacol. 2008, 591, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ozcan, F.; Ozmen, A.; Akkaya, B.; Aliciguzel, Y.; Aslan, M. Beneficial effect of myricetin on renal functions in streptozotocin-induced diabetes. Clin. Exp. Med. 2012, 12, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Liu, I.M.; Liou, S.S.; Cheng, J.T. Mediation of β-endorphin by myricetin to lower plasma glucose in streptozotocin-induced diabetic rats. J. Ethnopharmacol. 2006, 104, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, N.; Ashokkumar, N. Myricetin modulates streptozotocin-cadmium induced oxidative stress in long term experimental diabetic nephrotoxic rats. J. Funct. Foods 2013, 5, 1466–1477. [Google Scholar] [CrossRef]
- Ding, Y.; Dai, X.Q.; Zhang, Z.F.; Li, Y. Upregulation of Wnt signal pathway by myricetin attenuates β-cell dysfunction induced by cytokines. Prog. Biochem. Biophys. 2013, 40, 50–56. [Google Scholar] [CrossRef]
- Williams, L.K.; Li, C.; Withers, S.G.; Brayer, G.D. Order and disorder: Differential structural impacts of myricetin and ethyl caffeate on human amylase, an antidiabetic target. J. Med. Chem. 2012, 55, 10177–10186. [Google Scholar] [CrossRef] [PubMed]
- Gebhardt, R. Variable Influence of Kaempferol and Myricetin on in vitro Hepatocellular Cholesterol Biosynthesis. Planta Med. 2003, 69, 1071–1074. [Google Scholar] [PubMed]
- Chang, C.J.; Tzeng, T.F.; Liou, S.S.; Chang, Y.S.; Liu, I.M. Myricetin increases hepatic peroxisome proliferator-activated receptor protein expression and decreases plasma lipids and adiposity in rats. Evid. Based Complement. Altern. Med. 2012, 2012, 787152. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Wu, C.D. Compounds from Syzygium aromaticum possessing growth inhibitory activity against oral pathogens. J. Nat. Prod. 1996, 59, 987–990. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, P.; Sakharkar, M.K.; Lim, C.S.; Tang, T.H.; Sakharkar, K.R. Activity and interactions of antibiotic and phytochemical combinations against Pseudomonas aeruginosa in vitro. Int. J. Biol. Sci. 2010, 6, 556–568. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.D.; Chin, Y.P.; Lee, M.H. Antimicrobial activity of antibiotics in combination with natural flavonoids against clinical extended-spectrum β-lactamase (ESBL)-producing Klebsiella pneumonia. Phytother. Res. 2005, 19, 612–617. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, L.; Wahidulla, S.; Devi, P. Antibacterial phenolics from the mangrove Lumnitzera racemosa. Indian J. Mar. Sci. 2010, 39, 294–298. [Google Scholar]
- Griep, M.A.; Blood, S.; Larson, M.A.; Koepsell, S.A.; Hinrichs, S.H. Myricetin inhibits Escherichia coli DnaB helicase but not primase. Bioorg. Med. Chem. 2007, 15, 7203–7208. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.X.; Lee, S.F. Activity of plant flavonoids against antibiotic-resistant bacteria. Phytother. Res. 2001, 15, 39–43. [Google Scholar] [CrossRef]
- Kang, S.S.; Kim, J.G.; Lee, T.H.; Oh, K.B. Flavonols inhibit sortases and sortase-mediated Staphylococcus aureus clumping to fibrinogen. Biol. Pharm. Bull. 2006, 29, 1751–1755. [Google Scholar] [CrossRef] [PubMed]
- Naz, S.; Siddiqi, R.; Ahmad, S.; Rasool, S.A.; Sayeed, S.A. Antibacterial activity directed isolation of compounds from Punica granatum. J. Food Sci. 2007, 72, M341–M345. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.K.; Thakur, J.; Prakash, O.; Khan, F.; Saikia, D.; Gupta, M.M. Screening of flavonoids for antitubercular activity and their structure-activity relationships. Med. Chem. Res. 2013, 22, 2706–2716. [Google Scholar] [CrossRef]
- Ono, K.; Nakane, H.; Fukushima, M.; Chermann, J.C.; Barré-Sinoussi, F. Differential inhibitory effects of various flavonoids on the activities of reverse transcriptase and cellular DNA and RNA polymerases. Eur. J. Biochem. 1990, 190, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Veljkovic, V.; Mouscadet, J.F.; Veljkovic, N.; Glisic, S.; Debyser, Z. Simple criterion for selection of flavonoid compounds with anti-HIV activity. Bioorg. Med. Chem. Lett. 2007, 17, 1226–1232. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.X.; Qi, J.-B.; Cheng, K.J.; Hu, C.Q. Anti-HIV chemical constituents of aerial parts of Caragana rosea. Yaoxue Xuebao 2007, 42, 179–182. [Google Scholar]
- Chu, S.C.; Hsieh, Y.S.; Lin, J.Y. Inhibitory effects of flavonoids on moloney murine leukemia virus reverse transcriptase activity. J. Nat. Prod. 1992, 55, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.S.; Lee, J.; Lee, J.M.; Kim, Y.; Chin, Y.W.; Jee, J.G.; Keum, Y.S.; Jeong, Y.J. Identification of myricetin and scutellarein as novel chemical inhibitors of the SARS coronavirus helicase, nsP13. Bioorg. Med. Chem. Lett. 2012, 22, 4049–4054. [Google Scholar] [CrossRef] [PubMed]
- Keum, Y.S.; Jeong, Y.J. Development of chemical inhibitors of the SARS coronavirus: Viral helicase as a potential target. Biochem. Pharmacol. 2012, 84, 1351–1358. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.C.; Jung, H.Y.; Harikishore, A.; Kwon, O.D.; Yoon, H.S.; Kim, K.T.; Choi, B.H. The flavonoid myricetin reduces nocturnal melatonin levels in the blood through the inhibition of serotonin N-acetyltransferase. Biochem. Biophys. Res. Commun. 2013, 440, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Thiyagarajah, P.; Kuttan, S.C.; Lim, S.C.; Teo, T.S.; Das, N.P. Effect of myricetin and other flavonoids on the liver plasma membrane Ca2+ pump kinetics and structure-function relationships. Biochem. Pharmacol. 1991, 41, 669–675. [Google Scholar] [CrossRef]
- Pereira, T.A.; Das, N.P. Assay of liver cytosol lipoxygenase by differential pulse polarography. Anal. Biochem. 1991, 197, 96–100. [Google Scholar] [CrossRef]
- Yuksel, S.; Yesilada, E.; Gulbay, G.; Kurtoglu, E.; Serap Savaci, S. Protective effect of myricetin against e2-induced genotoxic damage in human lymphocytes. Fresen. Environ. Bull. 2012, 21, 1022–1026. [Google Scholar]
- Panickar, K.S.; Anderson, R.A. Mechanisms underlying the protective effects of myricetin and quercetin following oxygen-glucose deprivation-induced cell swelling and the reduction in glutamate uptake in glial cells. Neuroscience 2011, 183, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Barlas, N.; Ozer, S.; Karabulut, G. The estrogenic effects of apigenin, phloretin and myricetin based on uterotrophic assay in immature Wistar albino rats. Toxicol. Lett. 2014, 226, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Yang, Z.S.; Wen, C.C.; Chang, Y.S.; Wang, B.C.; Hsiao, C.A.; Shih, T.L. Evaluation of the structure-activity relationship of flavonoids as antioxidants and toxicants of zebrafish larvae. Food Chem. 2012, 134, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.D.; Liu, L.; Guo, W.; Meydani, M. Chemical structure of flavonols in relation to modulation of angiogenesis and immune-endothelial cell adhesion. J. Nutr. Biochem. 2006, 17, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Canada, A.T.; Watkins, W.D.; Nguyen, T.D. The toxicity of flavonoids to guinea pig enterocytes. Toxicol. Appl. Pharmacol. 1989, 99, 357–361. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, Y.M.; Xu, J.; Hua, R. Periphery analgesic effect of myricetin on a rat model of inflammatory pain. Chin. Pharmacol. Bull. 2015, 31, 1108–1111. [Google Scholar]
- Smith, C.; Halliwell, B.; Aruoma, O.I. Protection by albumin against the pro-oxidant actions of phenolic dietary components. Food Chem. Toxicol. 1992, 30, 483–489. [Google Scholar] [CrossRef]
- Hakkinen, S.H.; Karenlampi, S.O.; Mykkanen, H.M.; Torronen, A.R. Influence of domestic processing and storage on flavonol contents in berries. J. Agric. Food Chem. 2000, 48, 2960–2965. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semwal, D.K.; Semwal, R.B.; Combrinck, S.; Viljoen, A. Myricetin: A Dietary Molecule with Diverse Biological Activities. Nutrients 2016, 8, 90. https://doi.org/10.3390/nu8020090
Semwal DK, Semwal RB, Combrinck S, Viljoen A. Myricetin: A Dietary Molecule with Diverse Biological Activities. Nutrients. 2016; 8(2):90. https://doi.org/10.3390/nu8020090
Chicago/Turabian StyleSemwal, Deepak Kumar, Ruchi Badoni Semwal, Sandra Combrinck, and Alvaro Viljoen. 2016. "Myricetin: A Dietary Molecule with Diverse Biological Activities" Nutrients 8, no. 2: 90. https://doi.org/10.3390/nu8020090