 Sabine Hilt1*
Sabine Hilt1* Marta M. Alirangues Nuñez1
Marta M. Alirangues Nuñez1 Elisabeth S. Bakker2
Elisabeth S. Bakker2 Irmgard Blindow3Thomas A. Davidson4
Irmgard Blindow3Thomas A. Davidson4 Mikael Gillefalk1
Mikael Gillefalk1 Lars-Anders Hansson5Jan H. Janse2,6Annette B. G. Janssen2,7
Lars-Anders Hansson5Jan H. Janse2,6Annette B. G. Janssen2,7 Erik Jeppesen4,8Timm Kabus9Andrea Kelly10 Jan Köhler1Torben L. Lauridsen4,8Wolf M. Mooij2,11Ruurd Noordhuis12Geoff Phillips13
Erik Jeppesen4,8Timm Kabus9Andrea Kelly10 Jan Köhler1Torben L. Lauridsen4,8Wolf M. Mooij2,11Ruurd Noordhuis12Geoff Phillips13 Jacqueline Rücker14Hans-Heinrich Schuster15Martin Søndergaard4,8
Jacqueline Rücker14Hans-Heinrich Schuster15Martin Søndergaard4,8 Sven Teurlincx2Klaus van de Weyer16
Sven Teurlincx2Klaus van de Weyer16 Ellen van Donk2Arno Waterstraat17
Ellen van Donk2Arno Waterstraat17 Nigel Willby13
Nigel Willby13 Carl D. Sayer18
Carl D. Sayer18- 1Department of Ecosystem Research, Leibniz-Institute of Freshwater Ecology and Inland Fisheries, Berlin, Germany
- 2Departmnet of Aquatic Ecology, Netherlands Institute of Ecology (NIOO-KNAW), Wageningen, Netherlands
- 3Biological Station of Hiddensee, University of Greifswald, Greifswald, Germany
- 4Department of Bioscience, Aarhus University, Silkeborg, Denmark
- 5Department of Biology, Lund University, Lund, Sweden
- 6Netherlands Environmental Assessment Agency (PBL), Den Haag, Netherlands
- 7Water Systems and Global Change Group, Wageningen University and Research, Wageningen, Netherlands
- 8Sino-Danish Centre for Education and Research, University of Chinese Academy of Sciences, Beijing, China
- 9Institute of Applied Freshwater Ecology, Seddiner See, Germany
- 10Broads Authority, Norwich, United Kingdom
- 11Department of Aquatic Ecology and Water Quality Management, Wageningen University and Research, Wageningen, Netherlands
- 12Deltares, Delft, Netherlands
- 13Biological and Environmental Sciences, University of Stirling, Stirling, United Kingdom
- 14Department of Freshwater Conservation, Brandenburg University of Technology Cottbus-Senftenberg, Senftenberg, Germany
- 15Niedersächsischer Landesbetrieb für Wasserwirtschaft, Küsten- und Naturschutz, Sulingen, Germany
- 16Lanaplan, Nettetal, Germany
- 17Gesellschaft für Naturschutz und Landschaftsökologie, Kratzeburg, Germany
- 18Department of Geography, Environmental Change Research Centre, University College London, London, United Kingdom
Submerged macrophytes play a key role in north temperate shallow lakes by stabilizing clear-water conditions. Eutrophication has resulted in macrophyte loss and shifts to turbid conditions in many lakes. Considerable efforts have been devoted to shallow lake restoration in many countries, but long-term success depends on a stable recovery of submerged macrophytes. However, recovery patterns vary widely and remain to be fully understood. We hypothesize that reduced external nutrient loading leads to an intermediate recovery state with clear spring and turbid summer conditions similar to the pattern described for eutrophication. In contrast, lake internal restoration measures can result in transient clear-water conditions both in spring and summer and reversals to turbid conditions. Furthermore, we hypothesize that these contrasting restoration measures result in different macrophyte species composition, with added implications for seasonal dynamics due to differences in plant traits. To test these hypotheses, we analyzed data on water quality and submerged macrophytes from 49 north temperate shallow lakes that were in a turbid state and subjected to restoration measures. To study the dynamics of macrophytes during nutrient load reduction, we adapted the ecosystem model PCLake. Our survey and model simulations revealed the existence of an intermediate recovery state upon reduced external nutrient loading, characterized by spring clear-water phases and turbid summers, whereas internal lake restoration measures often resulted in clear-water conditions in spring and summer with returns to turbid conditions after some years. External and internal lake restoration measures resulted in different macrophyte communities. The intermediate recovery state following reduced nutrient loading is characterized by a few macrophyte species (mainly pondweeds) that can resist wave action allowing survival in shallow areas, germinate early in spring, have energy-rich vegetative propagules facilitating rapid initial growth and that can complete their life cycle by early summer. Later in the growing season these plants are, according to our simulations, outcompeted by periphyton, leading to late-summer phytoplankton blooms. Internal lake restoration measures often coincide with a rapid but transient colonization by hornworts, waterweeds or charophytes. Stable clear-water conditions and a diverse macrophyte flora only occurred decades after external nutrient load reduction or when measures were combined.
Introduction
Shallow lakes are the most abundant freshwater ecosystems on earth (Verpoorter et al., 2014). In their pristine state, they are often characterized by abundant submerged vegetation which can stabilize clear-water conditions (Scheffer et al., 1993) and plays a key role in the functioning of the ecosystem (Hilt et al., 2017). Several mechanisms contribute to a positive feedback between macrophytes and clear water conditions. As a consequence, shallow lakes are resistant to increasing nutrient loading up to a critical threshold, above which their macrophytes collapse and the lakes shift into a turbid, phytoplankton-dominated state (Scheffer et al., 1993). In recent centuries, excessive nutrient loading has resulted in a loss of macrophytes and shift to this turbid state in many temperate shallow lakes (e.g., Körner, 2002; Phillips et al., 2016).
Sayer et al. (2010a) suggested a typical pattern of lake macrophyte loss, defining a so-called “crashing” state lying between the stable clear-water state featuring a diverse plant community and the final turbid state lacking in macrophytes. This crashing state is characterized by the occurrence of only a few macrophyte species that can complete their life cycle during clear-water conditions in spring and early summer while later in summer, cyanobacteria blooms often occur. Eventually, the remaining macrophyte stands are also lost and give way to year-round phytoplankton dominance (Sayer et al., 2010a,b, 2016). Under these conditions, several ecosystem functions and services deteriorate, including biodiversity support, nutrient retention, provision of water of drinking or swimming quality (Hilt et al., 2017).
Hence, considerable efforts and financial resources have been devoted to the restoration of shallow lakes in many countries in recent decades (Jeppesen et al., 2005). The success of lake restoration in the long-term depends critically on the stable recovery of submerged macrophytes (Hilt et al., 2006). However, the turbid state is stabilized by feedback mechanisms that can prevent macrophyte re-colonization even at reduced nutrient loading. In theory, only the reduction of nutrient levels below a critical threshold or a significant reduction in the abundance of planktivorous and benthivorous fish (e.g., by biomanipulation or natural fish kills) will lead to a recovery of clear-water conditions and a return of macrophytes (Scheffer et al., 1993). In practice, reductions in the external nutrient load to shallow lakes often fail to deliver macrophyte recovery (Jeppesen et al., 2005). Similarly, biomanipulation of the fish community in turbid shallow lakes has produced variable effects on macrophytes in shallow lakes (Hansson et al., 1998; Bergman et al., 1999; Søndergaard et al., 2008; Jeppesen et al., 2012; Bernes et al., 2015; Sayer et al., 2016). Overall, the response of macrophyte communities to different types of lake restoration measures remains to be fully understood (Jeppesen et al., 2005; Bakker et al., 2013).
We hypothesize that (1) external lake restoration measures leading to nutrient load reduction in turbid temperate shallow lakes result in macrophyte re-establishment in a reversed sequence to the one described by Sayer et al. (2010a,b) for advancing eutrophication. An intermediate recovery state should occur where the water is clear in spring but dominated by phytoplankton and thus turbid in late summer, until, eventually, seasonally stable conditions characterized by high water clarity in both spring and summer would dominate (Figure 1). In contrast, lake internal measures such as biomanipulation or phosphorus precipitation are expected to result in transient clear-water conditions in spring and summer if either zooplankton is sufficiently released from fish predation or internal phosphorus loading from sediments is reduced enough to control summer phytoplankton. Such conditions are supposed to occur only temporarily in the absence of additional external nutrient load reduction (Figure 1). We hypothesize that (2) these contrasting types of restoration measures result in different macrophyte community composition and seasonal patterns in plant abundance. Specific macrophyte communities with short growth seasons should dominate during the intermediate recovery state following external nutrient loading reduction, while species with longer growing season requirements are predicted to temporarily establish following lake-internal measures (Figure 1). The establishment of stable clear conditions with a diverse macrophyte community is thus assumed to require both, external and internal measures (Figure 1).
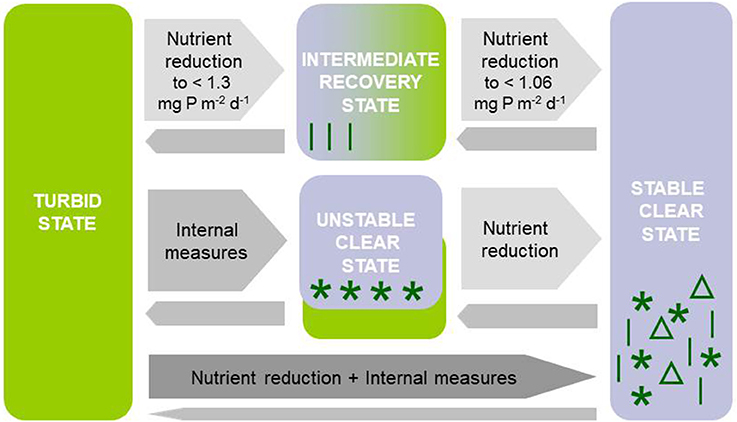
Figure 1. Response patterns of turbid north temperate shallow lakes to different restoration measures: (1) External restoration measures (reduction of external nutrient loading) are expected to lead to an intermediate recovery state with clear-water conditions in spring and turbid water in summer and specific macrophyte communities with short growth seasons and eventually stable clear conditions with a diverse macrophyte flora if nutrient loading is reduced sufficiently or additional internal measures are applied (reversed order as suggested for eutrophication by Sayer et al., 2010a,b). Thresholds in phosphorus (P) loading are based on simulations using PCLake (see Figure 5). (2) Lake-internal measures (biomanipulation, sediment suction dredging) leading to unstable clear-water conditions with specific macrophyte communities that may collapse resulting in a shift back to turbid conditions unless nutrient loading is reduced, or (3) a combination of external and internal restoration leading to stable clear-water conditions with an abundant and diverse macrophyte community.
To test these hypotheses, we analyse existing data on the water quality and submerged macrophytes of 49 temperate shallow lakes that had deteriorated to a turbid state and subsequently were subject to either external or internal restoration measures or both. In addition, we use an adapted version of the ecosystem model PCLake (Janse et al., 2008) to simulate the response of water clarity and macrophyte biomass to external nutrient load reduction and to detect any thresholds in nutrient loading for macrophyte recovery. Traits of the typical macrophyte species found after external and internal lake restoration measures are compared to provide a mechanistic understanding of the observed re-colonization patterns.
Materials and Methods
Literature and Data Search on Macrophyte Species Recovery
We started our literature review with the 22 shallow lakes described in detail in the study by Jeppesen et al. (2005) on the response of lakes to reduced external nutrient loading. However, only two of these 22 lakes were turbid before the nutrient load reduction (Müggelsee, Veluwemeer) and both showed an increase in macrophyte coverage following the intervention. The rest showed no change, macrophyte declines or lacked suitable data (Jeppesen et al., 2005). Therefore, we searched for more examples in published and unpublished studies on lakes in Germany, The Netherlands, Denmark, southern Sweden and UK where shallow lakes are abundant and experience similar eutrophication problems and climatic conditions. We selected lakes that had lost most of their submerged macrophytes during a turbid phase and subsequently had been subjected to either external restoration via nutrient load reduction (summarized in Table 1), or internal restoration using biomanipulation or sediment dredging (Table 2). We did not carry out a full systematic review of available data, but instead focussed on known lakes within the research network of the authors where at least partial recovery through restoration measures was evident.
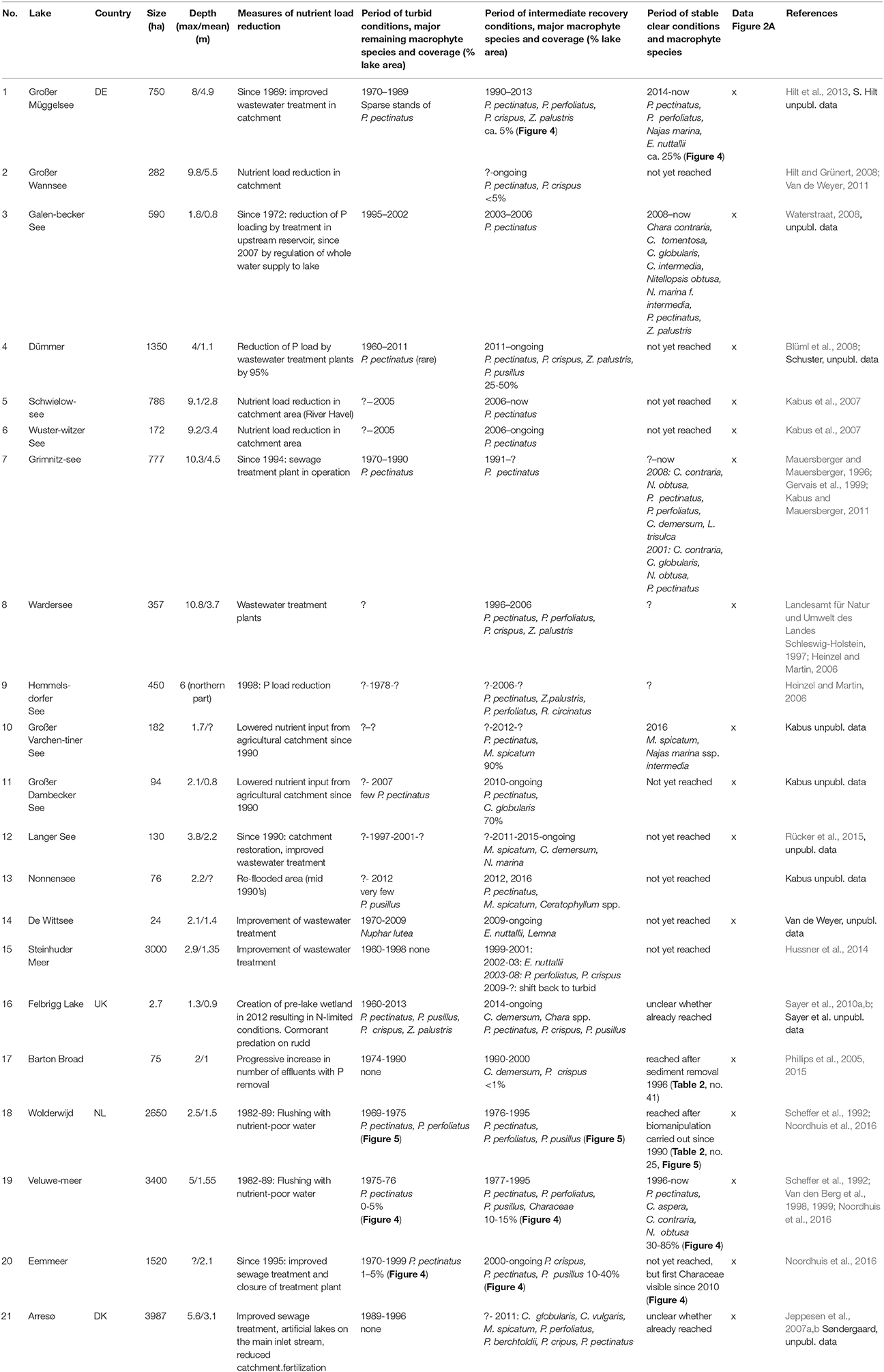
Table 1. The response of north temperate shallow lakes in Germany (DE), United Kingdom (UK), The Netherlands (NL), and Denmark (DK) to external nutrient load reduction (x: data on Secchi disk transparency and total phosphorus concentrations were available and used in Figure 2A).
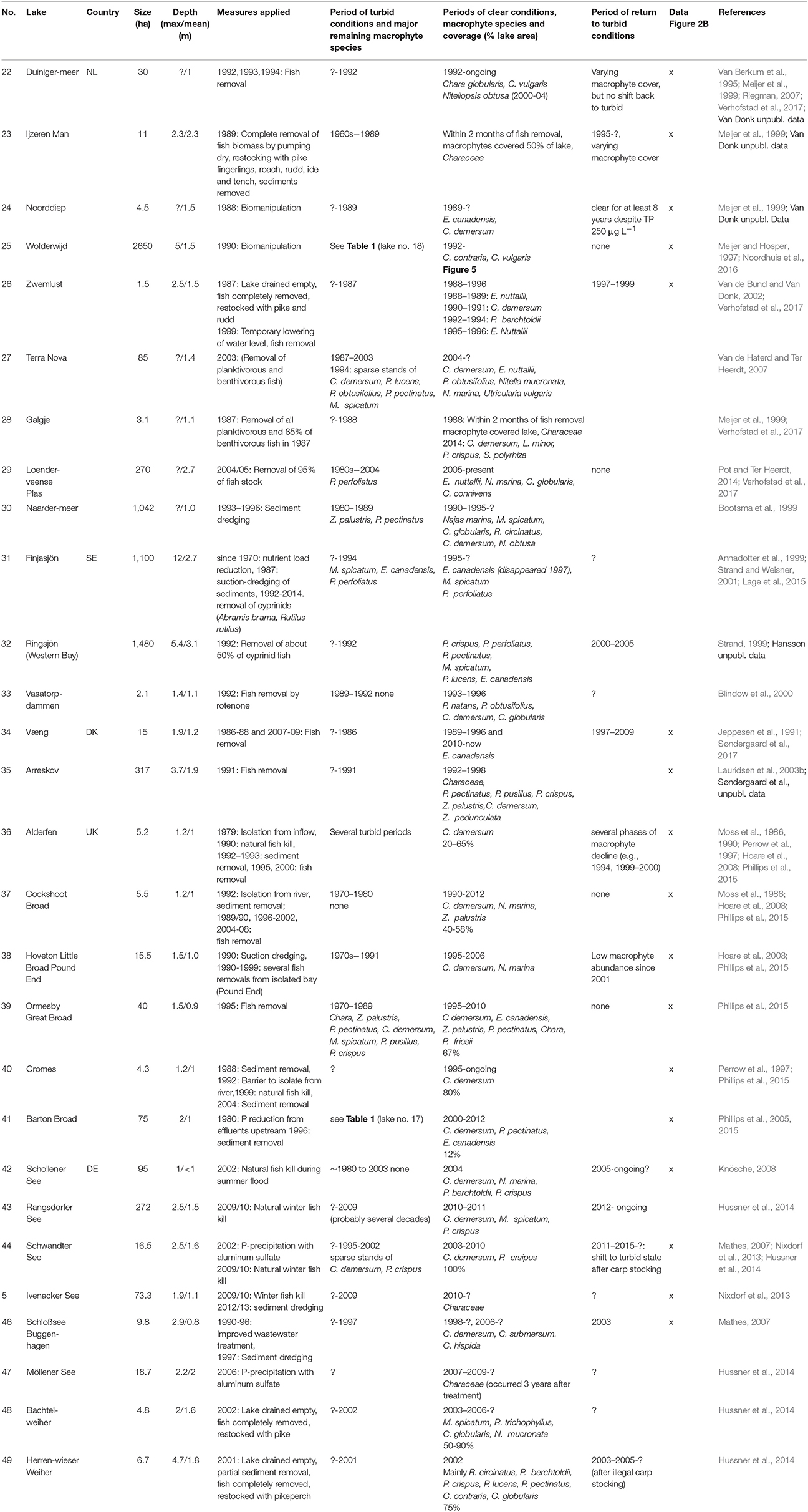
Table 2. The response of north temperate shallow lakes in The Netherlands (NL), Sweden (SE), Denmark (DK), United Kingdom (UK) ,and Germany (DE) to biomanipulation, natural fish kills or other lake-internal measures (x: data on Secchi disk transparency and total phosphorus concentrations were available and used in Figure 2B).
For all lakes, we retrieved information from the turbid period and its macrophyte assemblage, macrophyte composition after external/internal restoration, total phosphorus (TP) concentrations in the water and Secchi depth in spring (April-June) and summer (July–September) using published studies or questionnaire responses provided by co-authors. As is commonly the case, data on TP concentrations and Secchi depth were very diverse, ranging from multi-year weekly measurements to single values. The data were merged into a single value for each season and lake by using means (Figures 2A,B) and raw data are available as a supplement. We also analyzed the occurrence of dominant macrophyte species during the recovery period. To visualize potentially typical recovery patterns, the long-term changes in nutrient concentrations, water transparency and macrophyte occurrence are shown in more detail for three lakes restored only by reductions in external nutrient loading (Müggelsee, Veluwemeer, Eemmeer, for details see no. 1, 19 and 20 in Table 1) and for three lakes restored through biomanipulation of the fish community (Noorddiep, Wolderwijd, Zwemlust, for details see no. 24–26 in Table 2).
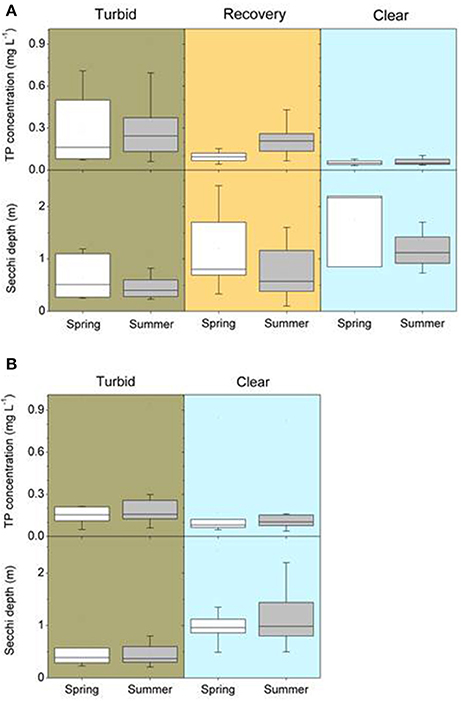
Figure 2. Total phosphorus (TP) concentrations and Secchi depth in spring (April–June) and summer (July–September) of different north temperate shallow lakes (A) before and after external nutrient load reductions during the turbid, the intermediate recovery and the clear-water state (for details see Table 1) and (B) before (turbid) and after (clear) biomanipulation or other lake-internal measures (for details see Table 2).
Mann-Whitney U-tests were performed to compare lake size, maximum and mean depths between lakes with different restoration measures (external nutrient load reduction vs. internal measures). Total phosphorus concentrations and Secchi disk transparency were compared between the different states (turbid, intermediate, clear) in lakes with external nutrient load reduction) using Kruskal-Wallis tests and subsequent posthoc comparisons (separately for different seasons). The same was done for lakes with internal measures using Mann-Whitney U tests. All statistical tests were run in SPSS.
PCLake Simulations
Simulating the response of water clarity and macrophyte biomass to external nutrient load reduction and detecting thresholds of nutrient loading for either intermediate recovery or clear state required adaptation of the established ecosystem model PCLake. This model has previously been used to estimate threshold responses of shallow lakes to nutrient loading (Janse et al., 2008; Janssen et al., 2017), and to simulate the response of temperate shallow lakes to climate warming (Mooij et al., 2007), to mowing of macrophytes (Kuiper et al., 2017) and—in a variant of the model with three plant species—to biomanipulation and herbivory (Janse et al., 1998). PCLake consists of a number of coupled ordinary differential equations that describe the most important biotic (submerged macrophytes, phytoplankton, detritivorous macrozoobenthos, zooplankton, zooplanktivorous fish, benthivorous fish, and piscivorous fish) and abiotic (detritus, inorganic material, dissolved phosphorus, ammonium, and nitrate) components of both the water column and the top-layer of the sediment in a non-stratifying shallow lake (Janse, 1997, 2005). All organic components (apart from predatory fish) are modeled in terms of dry weight (DW), nitrogen (N), and phosphorus (P), and hence the nutrient-to-dry-weight ratios of the organic components are variable. Internal fluxes of nutrients between the sediment layer and the pelagic zone, including internal loading, are accounted for and modeled dynamically.
For our simulations we used the default settings of a lake in PCLake. This default lake represents a relatively shallow lake with an average depth of 2 m and is relatively small with a maximum fetch of 1,000 m, an areal hydraulic loading of 20 mm days−1 (= 7.2 m year−1), no infiltration or seepage, no surrounding wetland zone, and a lightly clayish sediment (30% dry matter, of which 10% organic matter, and 10% fine mineral material) (Janse 2005). Due to small size and shallowness, the lake is dominated by macrophytes when nutrient loads are sufficiently low, but as nutrient loads increase the lake switches to a turbid state. This switch occurs rather suddenly due to the positive feedbacks in the model (Janse 2005) that lead to a critical transition (e.g., Scheffer and Carpenter, 2003). A common method to determine critical transitions is bifurcation analysis. In this approach the model is run to equilibrium several times, each with a different nutrient load. For each run, the yearly average phytoplankton chlorophyll-a concentration and macrophyte biomass are calculated. To assess the presence of hysteresis this procedure is repeated twice for each level of nutrient load, the first starting from a clear lake and the second from a turbid lake. Where the equilibrium outcomes of these two runs with identical nutrient load differ, hysteresis is inferred. Here we ran the model for nutrient loads ranging from 0.1 to 2.5 mg m−2 days−1 (0.4–9.0 kg ha−1 year−1) to cover a wide range of the eutrophication axis. The output of the bifurcation analysis is a load-response curve (or bifurcation plot) showing the effect of nutrient load on the biomass of primary producers. The point of a sudden switch marks the critical transition(s).
To simulate the influence of temperature and light on the response of different macrophyte species to nutrient load reduction we had to make two adjustments to the original formulations of PCLake, while maintaining the modeling of macrophytes as one functional group. First, the original power function for temperature limitation of macrophytes was replaced by a temperature optimum curve LT (-):
Here, σ (°C) is the temperature constant based on a Gaussian curve, T (°C) is the water temperature, Topt (°C) is the optimum temperature for macrophytes and Tref (°C) is the reference temperature used to normalize the limitation function to 1 (Janse, 2005). With this function the model is flexible to simulate macrophytes with different temperature optima. The second adjustment is the timing of root allocation which occurs in the autumn when macrophytes store energy to overwinter, for instance, in propagules. In the original PCLake, this timing was linked to a specific day in the year while submerged plants are known to respond to physiological and environmental cues, such as light availability, to determine timing of root allocation (Madsen, 1991). Hence, we decided to link the timing of root allocation to a minimum daily light availability for macrophytes, following Madsen (1991), Van Dijk and Van Vierssen (1991), and Van Dijk and Janse (1993). The available light for macrophytes is based on the light availability over the depth of the water column corrected for periphyton shading which is estimated by:
In which PAR (W m−2) is an approximation of the average Photosynthetic Available Radiation (PAR) for plant photosynthesis, PAR0 (W m−2) is the PAR available at the top of the macrophyte layer, PARbot (W m−2) is the PAR available at the bottom of the macrophyte layer, D (m) is the depth and εperiphyton is the shading by periphyton. In order to restrict complexity, we refrained from adding periphyton as an extra compartment to the model, but instead used the empirical relationship of Vadeboncoeur et al. (2006) to estimate periphyton chlorophyll-a biomass:
where c1 = 1.79 and c2 = 0.85 and TP is the in-lake total phosphorus concentration (mg m−3). The periphyton chlorophyll-a biomass is then used to estimate the shading effect:
where S is the specific light attenuation by periphyton (m g−1) and correction factors for light limitation f(L), temperature limitation f(T) and available plant surface area for periphyton growth f(α). The shading effect of periphyton affects the timing of root allocation through light availability to macrophytes as well as the light limitation of macrophyte shoots.
PCLake has previously been calibrated following a Bayesian approach to parameter estimation and uncertainty analysis using data from nearly 40 temperate shallow lakes (Janse et al., 2010). Although this calibration did not account for the specific effect of periphyton, the data used for model calibration most likely integrate this effect indirectly (Kuiper, 2016). By adding the effect of periphyton to PCLake implicitly, we thus have to technically recalibrate the model. This implies adjusting the parameter settings of the model, such that given the same boundary conditions, the model produces the same output. Therefore we have calibrated the adjusted model manually by lowering the half saturation light constant of vegetation and decreasing the parameter for dark respiration of vegetation (see Table 3 for new and calibrated parameter settings). PCLake is implemented in DATM (Mooij et al., 2014). For the full overview of parameter settings and model formulations, please see the DATM-file in the Supplementary Material.
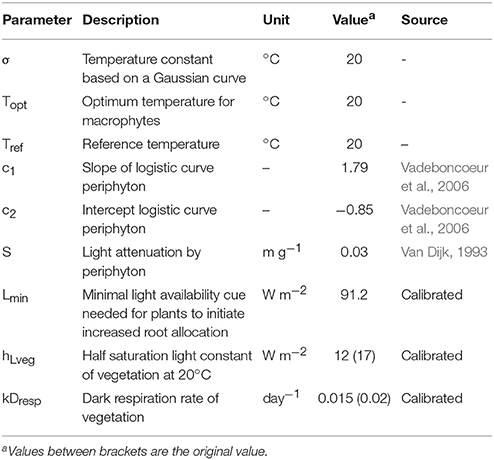
Table 3. Parameter settings for PCLake.
Results
Lake Water Quality following External and Internal Restoration
Our literature review provided information on water quality and macrophyte development in 21 turbid lakes that were subject to external nutrient loading reduction without additional in-lake measures (Table 1) and 28 lakes with in-lake restorative measures. Some of these measures were preceded or accompanied by external nutrient loading reduction (Table 2).
Lakes with internal measures were on average smaller than lakes with external nutrient load reduction alone, while maximum depths were higher but mean depths were similar (Table 4). Turbid conditions lasted from 1 year (Lake Veluwe) to 51 years (Dümmer). Often, however, exact timing and duration of the turbid period are unknown (Tables 1, 2). During the turbid phase, spring and summer TP concentrations were high (~ >0.15 mg L−1), while Secchi disk transparencies were low (~0.4 m), with considerable differences between lakes (Figures 2A,B, Table 4).
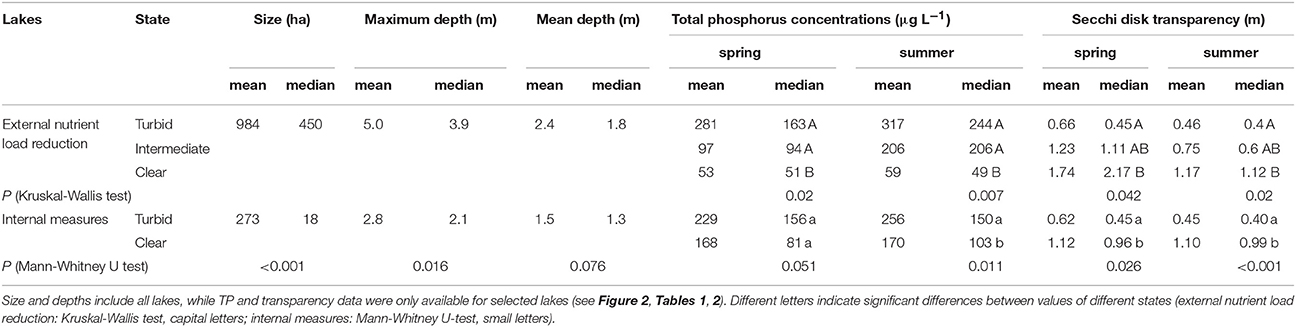
Table 4. Size, maximum and mean depths, total phosphorus concentrations and Secchi disk transparency in lakes after external nutrient load reduction or implementation of internal measures.
External nutrient loads were usually reduced following improved sewage treatment in the catchment. Lake Veluwe and Wolderwijd were flushed with nutrient-poor water (Table 1). The most commonly applied internal measure was biomanipulation in the form of removal of benthivorous and planktivorous fish (in some cases by draining/pumping the lake dry). In nine lakes, sediment removal was applied as an additional or the sole measure and in five lakes natural fish kills occurred during severe winters or as a result of a summer flood (Table 2), emulating the effects of planned biomanipulation. During the first years after nutrient load reduction, TP concentrations were lower than during the turbid period, but still about twice as high in summer as in spring. Spring water transparency was higher than during the turbid period, while summer values were still low (Table 4, Figure 2A). This phase has been found to last up to 20 years (e.g., Müggelsee), but often its start has not been recorded and/or lakes have not yet reached stable clear conditions (Table 1). Lakes Müggelsee, Veluwemeer, and Eemmeer show a similar intermediate recovery state with spring water transparencies being higher than during turbid summer conditions which lasted for about 20 years (Figure 3). A switch back from intermediate to turbid conditions has only been observed in Lake Steinhuder Meer, in this case about 10 years after macrophytes returned (Table 1).
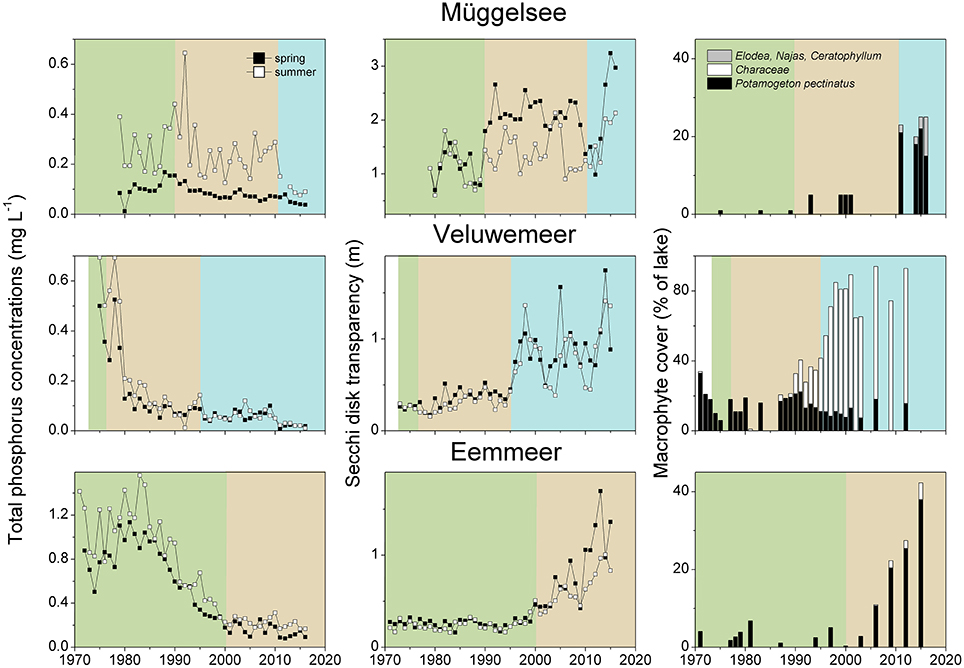
Figure 3. Total phosphorus (TP) concentrations and Secchi depth in spring (April–June) and summer (July–September) and macrophyte coverage in Lake Müggelsee; Lake Veluwe and Lake Eem during the turbid (green), the intermediate recovery (brown) and the clear-water (blue) state (for lake details see Table 1).
After implementation of in-lake measures, TP concentrations were at the same level as before, in both spring and summer, whereas water transparency in spring and summer was significantly higher than before restoration (Table 4, Figure 2B). These unstable clear conditions often only lasted for a few years and many lakes shifted back to turbid conditions (e.g., no. 23, 26, 32, 34, 36, 38, 42, 43, 44, 46, 49 in Table 2). Fish stock reductions in Wolderwijd, Zwemlust and Noorddiep led to clear conditions both in spring and summer (Figure 4). Lake Zwemlust shifted back to turbid conditions after 9 years, while Noorddiep stayed clear for at least 8 years with no further information on subsequent periods.
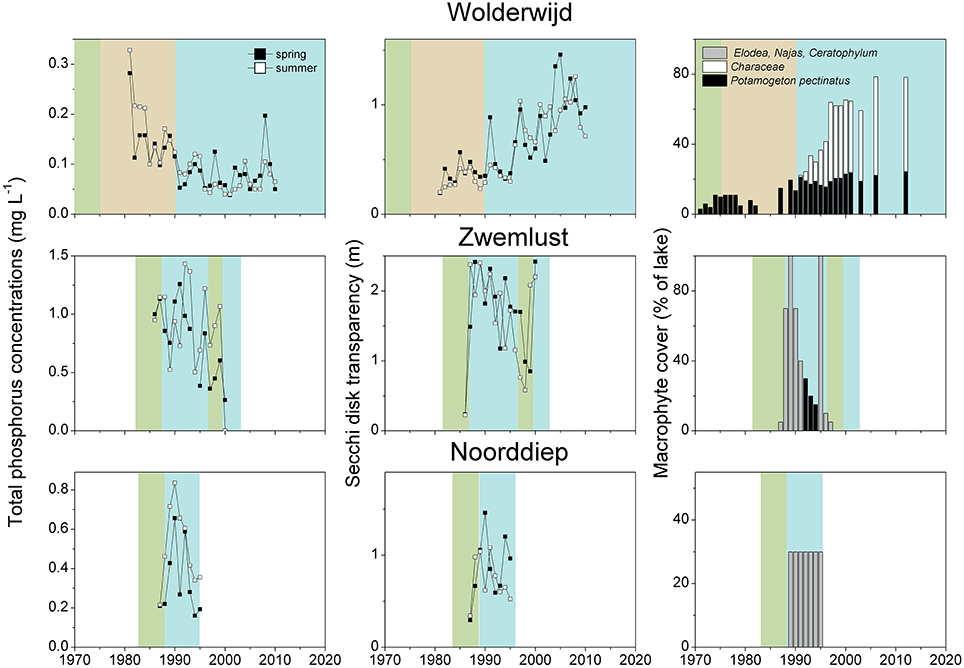
Figure 4. Total phosphorus (TP) concentrations and Secchi depth in spring (April–June) and summer (July–September) and macrophyte coverage in Lake Wolderwijd, Lake Zwemlust and Lake Noorddiep before (green and brown) and after (blue) biomanipulation (for lake details see Table 2). In Lake Zwemlust, P. berchtoldii occurred instead of P. pectinatus, and the coverage in Lake Noorddiep was only estimated based on the information that it was higher than 25% (Gulati and Van Donk, 2002).
Only six of the lakes with external nutrient load reduction (~25%, no. 1, 3, 7, 10, 18, and 19 in Table 1) reached stable clear-water conditions with lower ambient TP concentrations and higher Secchi depths than during the intermediate recovery state both in spring and summer (Figure 2A, Table 4). In four of the 28 lakes (14%, no. 25, 29, 37, 39 in Table 2, Lake Wolderwijd in Figure 4), stable, longer-term clear-water conditions with a diverse macrophyte flora were obtained after the application of in-lake measures. For several lakes, their longer-term development is not known (Table 2).
Model Simulations on Lake Response to External Nutrient Load Reduction
The results of the simulations using the adjusted PCLake model revealed three stages for lakes that undergo external nutrient load reduction: a turbid state, an intermediate recovery state and a clear state (Figure 5A). For the default lake in PCLake, the turbid state occurs if the P load exceeds 1.3 mg P m−2 days−1. The intermediate recovery state occurs between the two critical transitions that appear at a P load of 1.06 and 1.3 mg P m−2 days−1. If the P load drops below 1.06 mg P m−2 days−1 the lake turns into a clear state. The critical P loads for the intermediate recovery state are smaller in the adjusted model including periphyton than with the original formulation of PCLake (critical transition between approximately 1 and 2 mg P m−2 days−1, Janse et al., 2008). The periphyton effect thus reduces the threshold for intermediate recovery state of the default lake by about a quarter of what it would be without periphyton. The reason for this is that in case of hysteresis, the position of the highest critical nutrient load is mainly determined by macrophyte characteristics, while the position of the lowest critical nutrient load is mainly determined by phytoplankton characteristics. This is in agreement with the results of the sensitivity analysis of PCLake (Janse et al., 2010). Since periphyton negatively affects the performance of macrophytes, the highest critical nutrient load was reduced, while phytoplankton characteristics were unaltered and the lowest critical nutrient load (1 mg P m−2 days−1) thus did not change.
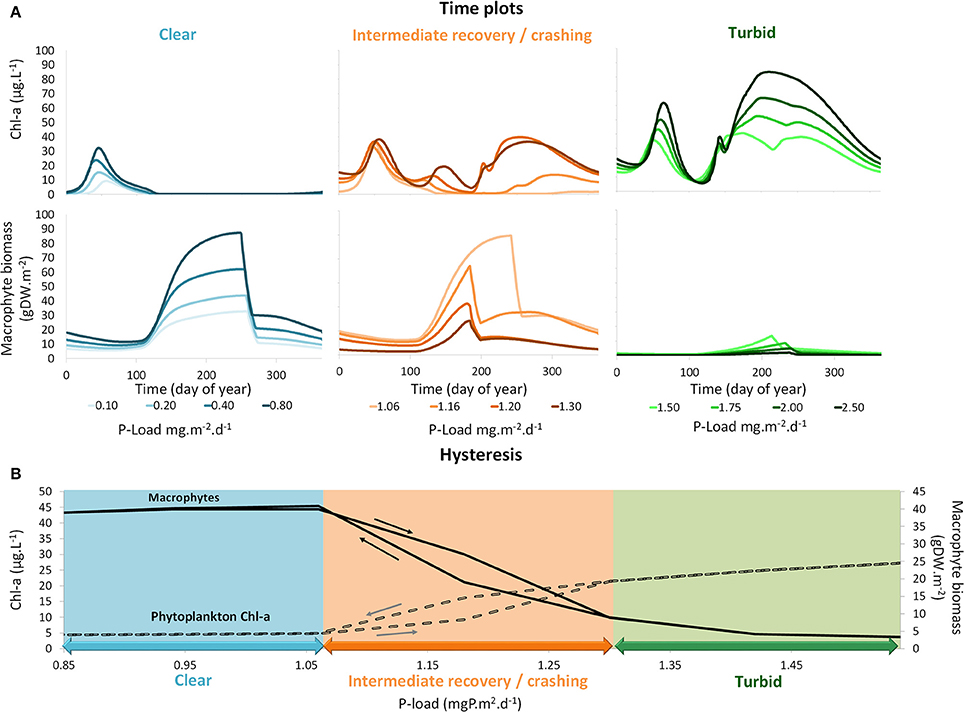
Figure 5. (A) Time series of simulation using PCLake of chlorophyll-a concentrations and macrophyte shoot biomass for different phosphorus (P) loadings within the clear, intermediate recovery and turbid states. Darker colors are associated with higher P loading simulations. (B) Hysteresis plots showing yearly mean simulated values of chlorophyll-a concentrations and macrophyte shoot biomass for different phosphorus (P) loadings within the clear, intermediate recovery and turbid states. Arrows denote the directions of the hysteresis effects.
The turbid state was characterized by lack of macrophytes and enhanced phytoplankton biomass with increasing nutrient loading (Figure 5A). The intermediate state was characterized by a changed phenology of the primary producers. With increasing nutrient loading, the phytoplankton summer peak shifted to earlier dates of the year and this advancement of phytoplankton was mirrored by an abbreviated macrophyte growing season (Figure 5A). The shorter growing season was a direct effect of the inclusion of periphyton shading in our adapted version of the model. The shading of macrophytes by periphyton was most severe at the peak of summer when the light input and water temperature are high. As a result, macrophyte growth was limited to the period just after the clear water phase in spring until the start of the summer phytoplankton bloom. The clear state was characterized by an increase of macrophyte biomass with increasing nutrient loading (Figure 5A). Phytoplankton production was restricted to the spring bloom peak and there was no summer bloom.
The bifurcation plot (Figure 5B) shows a less sudden transition of macrophytes and phytoplankton compared to the abrupt transition between the phytoplankton-dominated turbid state and the macrophyte-dominated clear state, often seen in bifurcation plots in literature (e.g., Janse et al., 2008). The gradual course of the bifurcation plot is due to the inclusion of periphyton shading of macrophytes. This shading permits high biomass of both macrophytes and phytoplankton within the same year during the crashing or intermediate recovery phase. Furthermore, the region of hysteresis is tilted, leading to a less abrupt and thus more realistic critical transition from phytoplankton dominance to macrophyte dominance and back again.
Macrophyte Species Recovery following External and Internal Restoration
During the turbid phase, three lakes with subsequent external nutrient load reductions were reported to lack macrophyte stands altogether and six and one lakes had sparse stands of Potamogeton pectinatus (also known as Stuckenia pectinata) and P. pusillus, respectively, while no information was available for the remaining lakes (Table 1). For lakes with internal measures, information on macrophyte species present during the turbid phase is available for 10 lakes. Apart from P. pectinatus and P. perfoliatus, plants such as Ceratophyllum demersum, M. spicatum or E. canadensis are mentioned, if indeed plant stands were present at all (Table 2).
During the intermediate recovery state P. pectinatus was the dominant macrophyte species in two thirds of the analyzed lakes with reduced external nutrient loading. Other pondweed species such as P. perfoliatus, P. crispus and P. pusillus or Zannichellia palustris were also found in several lakes during the intermediate recovery state, while other groups such as Characeae or Elodea species were much less common (Table 1, Figure 6). Lakes Müggelsee, Veluwemeer and Eemmeer were all dominated by P. pectinatus during the intermediate recovery state which lasted for about 20 years (Figure 3). Müggelsee and Veluwemeer seem to have entered a stable clear state with more diverse submerged vegetation (including Characeae in Veluwemeer) in 2011 and 1996, respectively, while Eemmeer has not yet reached that phase despite the recent detection of Characeae (Figure 3). Three other lakes also reached a stable clear state and were colonized by different species of Characeae and/or Najas marina, Elodea species and C. demersum (Table 1).
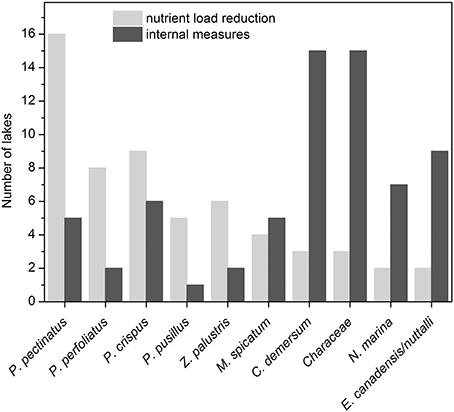
Figure 6. The 10 most common macrophyte taxa in north temperate shallow lakes during re-colonization after reduction of external nutrient loading (light gray) or implementation of internal measures (dark gray; for more details see Tables 1, 2).
The dominant macrophytes occurring after lake internal measures were Characeae, C. demersum, Elodea species or N. marina (Table 2, Figure 6). Often, lakes had only one or two dominant species, and in at least 10 cases, lakes switched back to turbid conditions and lost their macrophytes again (Table 2, Figure 4). The response of macrophytes occurred gradually in Wolderwijd, showing an increase in Characeae coverage, while in the much smaller Zwemlust and Noorddiep, macrophytes immediately covered large areas, E. nuttallii, E. canadensis and C. demersum dominating (Figure 4, Table 2). In four lakes, stable clear-water conditions were obtained after in-lake measures, all being characterized by Characeae dominance (Table 2, Figure 4).
Discussion
Our analysis of long-term data from 49 temperate shallow lakes during their recovery from a turbid phase reveals that both a reduction of the external nutrient loading and implementation of lake-internal measures often result in the occurrence of an intermediate state (Figure 1) that can last for several decades. External nutrient load reductions are often followed by the re-occurrence of a spring clear-water phase that opens a “window of opportunity” for macrophyte re-colonization, but with only a short growth period due to turbid conditions during summer. This pattern was confirmed by our model simulations (Figure 5). As hypothesized, macrophyte re-establishment following nutrient load reduction occurs in a reversed sequence to the one described for eutrophication by Sayer et al. (2010a,b). In contrast, lake internal measures such as fish or sediment removal often result in clear-water conditions during spring and summer. This clear-water state is, however, often only temporary and lakes frequently shift back to turbid conditions one or a few years after the restoration. Likely, the duration of the clear-water conditions is related to nutrient loading and the intensity of the restoration effort (Hansson et al., 1998). Only in a few examples have longer lasting clear-water conditions been observed. These required spring and summer TP concentrations below 0.05 mg L−1.
We also have evidence for our second hypothesis, namely that different types of restoration measures influence the macrophyte community composition. P. pectinatus and a few other pondweeds most often recolonise temperate shallow lakes with reduced external nutrient loading and dominate during the intermediate recovery state. The implementation of internal restoration measures results in the establishment of a different community, often consisting of a small selection from either hornwort (C. demersum), charophytes, water weeds (E. canadensis, E. nuttallii) or naiad (N. marina). Only in a few cases have lakes reached a state of clear-water conditions during spring and summer and with a more diverse macrophyte community.
Submerged Macrophyte Survival during the Turbid Phase
Whether and which macrophyte stands or propagules survive during the turbid phase depends on the occurrence of macrophyte species before the shift to turbid conditions and on the length and severity of the turbid phase (e.g., Vari and Toth 2017). Seed banks in shallow lake sediments have often been assumed to be insufficient for recovery of submerged vegetation by germination, due to low numbers of viable seedlings, strong seed dormancy, strict germination cues and the reliance of many species upon vegetative reproduction (Haag, 1983; Kautsky, 1990; Rodrigo et al., 2013; Baldridge and Lodge, 2014). In contrast, De Winton et al. (2000) and Verhofstad et al. (2017) have shown that seed banks from even the most degraded lakes are capable of an emergence response and thus offer a potential means to restore vegetation. In our survey, the duration of complete macrophyte loss was often unrecorded, but periods of several decades are common (Tables 1, 2). If macrophyte stands survived during this period and were recorded, these were often sparse stands of P. pectinatus, a species commonly associated with the crashing phase during eutrophication (Sayer et al., 2010a). This species survives in very shallow water even under phytoplankton dominance (Hilt et al., 2013) and its strongly apical growth form may allow it to survive at greater depths in turbid water. The shallow littoral, especially in larger lakes, is strongly disturbed by wave action and only species with high anchorage and breaking strength can survive under these conditions (Schutten et al., 2005). Potamogeton pectinatus has a high breaking strength (Brewer and Parker, 1990) and its phenotypic plasticity allows it to form short plants in shallower water (Idestam-Almquist and Kautsky, 1995). In contrast, other common species in eutrophic lakes such as E. canadensis or M. spicatum have been described as deep water species with lower tensile strength (Brewer and Parker, 1990), while C. demersum has no roots for anchorage. These species are thus less likely to persist through severely turbid states in very shallow littoral areas and to serve as remnant populations for re-colonization, at least in larger lakes. Nevertheless, they have been reported during turbid phases in four lakes included in our survey, most probably in wind-protected areas or bays.
Knowledge of the survival of propagules in sediments during turbid phases is limited. In general, charophyte oospores and macrophyte seeds have been found to survive up to 150 years (Kaplan and Muer, 1990; De Winton et al., 2000; Alderton et al., 2017). Germination tests with sediments have been suggested before implementing lake restoration measures to forecast the potential for macrophyte recovery from internal sources (Hilt et al., 2006), however, these are not routinely applied. Re-colonizing macrophyte clones of several species that originated from periods before eutrophication have been found (Sand-Jensen et al., 2008), but knowledge about the origin of re-colonizing macrophytes post-restoration remains scarce (Bakker et al., 2013).
Response of Macrophytes to Nutrient Load Reductions
Shallow temperate lakes often show a relatively rapid response to reductions in external phosphorus loading, characterized by a reduction in phytoplankton biomass during spring and early summer (Jeppesen et al., 2005). During late summer, however, the response is delayed because of sustained remobilisation of phosphorus from the sediment (Søndergaard et al., 2013). As a consequence, high phytoplankton and cyanobacterial abundance are often reasserted in summer (Sommer et al., 2012) resulting in turbid water and preventing macrophyte growth. In our survey, such conditions occurred at spring TP concentrations of around 0.1 mg L−1, while summer concentrations were twice as high.
The increased water transparency in spring and early summer seems to be exploited by certain macrophyte species, in our survey mainly P. pectinatus along with P. perfoliatus, P. crispus and Z. palustris. These macrophyte species are characterized by specific traits that may explain their prevalence. Firstly, they can compress their whole life cycle into the short clear-water period in spring and early summer due to early germination from tubers, turions or seeds, shortening time to peak biomass and allowing early formation of overwintering tubers and seeds (Table 5). Secondly, they can have short growth forms that can establish in very shallow habitats (e.g., Van Vierssen, 1982a). Thus, they are often the species that survive in turbid conditions in shallow margins (see Submerged Macrophyte Survival during the Turbid Phase) and then expand into deeper water with improvements in clarity during nutrient load reduction. Rhizomatic growth from remaining P. pectinatus stands has been shown, via microsatellite analyses, to be the dominant re-colonization mode in Lake Müggelsee; more recently established P. pectinatus stands had lower genotype diversity and were comprised of only a small subset of genotypes from shallower areas (Hilt et al., 2013). Thirdly, energy reserves in vegetative propagules such as tubers of P. pectinatus allow early onset of growth independent of light availability (Spencer, 1986). In addition, P. pectinatus can concentrate large parts of its biomass just under the water surface and thus survive in relatively turbid water (Van Wijk, 1988). An initial colonization of formerly turbid lakes with P. pectinatus during recovery has also been observed in deeper, stratifying lakes. Thus, in Lake Tegel (Germany), this species dominated for more than 20 years after the start of phosphorus stripping in the major inflow (Hilt et al., 2010).
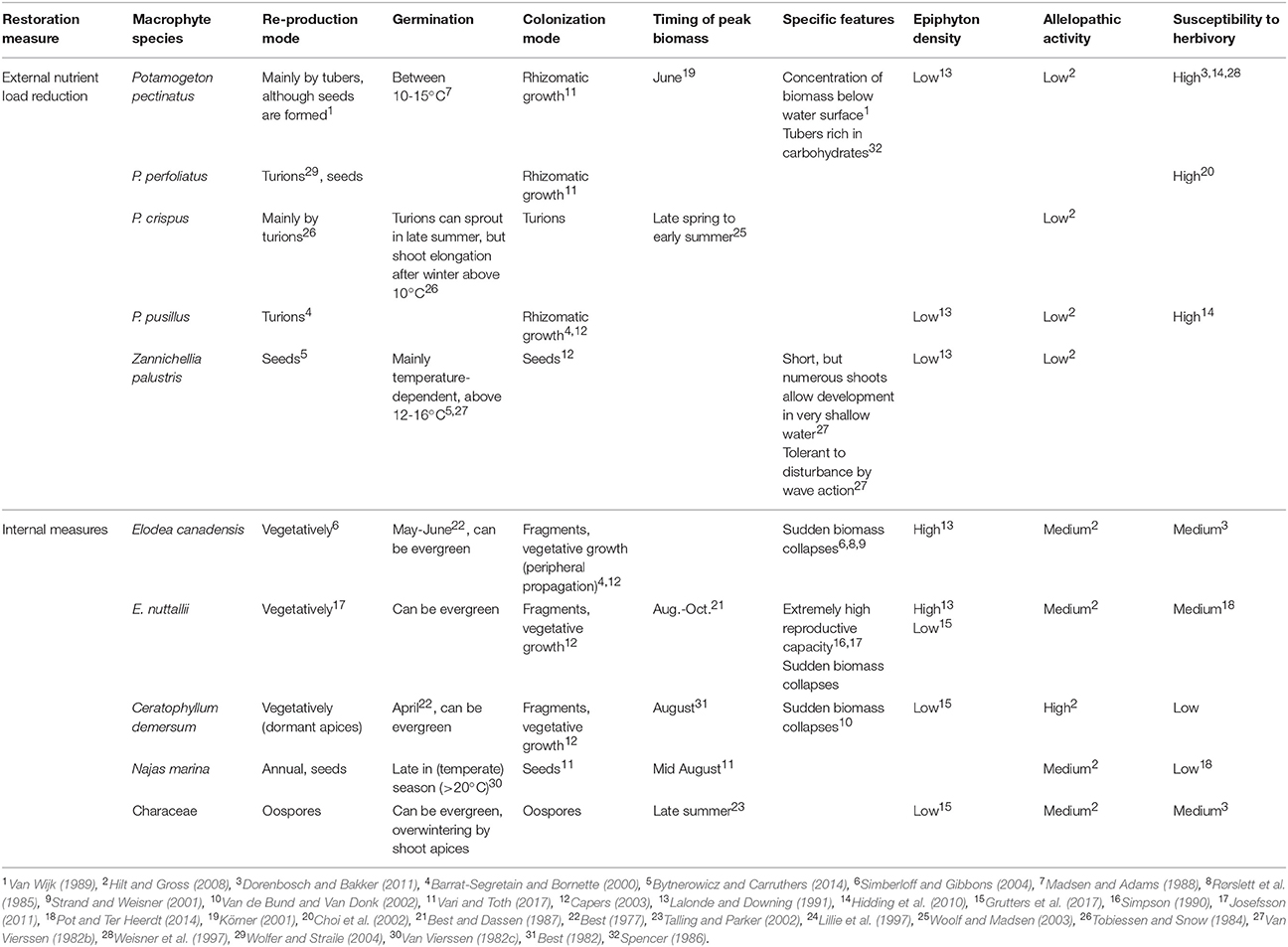
Table 5. Characteristics of macrophyte species/groups typically recolonising temperate shallow lakes after external nutrient load reduction or following implementation of lake-internal measures such as biomanipulation.
Usually, the maximum colonization depth of macrophytes during the intermediate recovery phase is low (around 1 m) and consequently, depending on the lake morphometry, only small parts of the lake bed might be covered. In contrast, very shallow lakes may reach over 50% cover (Table 1). This suggests that in “deeper” shallow lakes, macrophyte coverage during the intermediate phase may be insufficient to stabilize clear-water conditions during later summer. Based on the findings of Søndergaard et al. (2016) submerged macrophyte coverage on average needs to pass a threshold of 20% of lake area to markedly lower phytoplankton densities. In principle, small stands can be sufficient as a refuge for phytoplankton-grazing zooplankton against fish predation (Lauridsen et al., 1996; Portielje and Van der Molen, 1999). However, abundant colonial and filamentous cyanobacteria which often dominate the summer phytoplankton communities during lake recovery cannot be effectively controlled by zooplankton grazers (Wang et al., 2010 and references therein). Bottom-up stabilizing mechanisms of macrophytes on water clarity such as nutrient competition, increased sedimentation within stands and reduced sediment resuspension will be inefficient at low plant coverage (Blindow et al., 2014). Low coverage is, however, not the only reason why macrophytes in the intermediate recovery phase cannot stabilize clear-water conditions in late summer, as shown in the case of the very shallow Lake Dümmer, where cyanobacteria blooms still occurred in summer despite high macrophyte coverage.
Our model simulations suggest that high periphyton shading triggers macrophyte disappearance in summer. Periphyton shading, often accompanied by herbivory (Hidding et al., 2016), has been shown to impair macrophyte development in empirical studies (e.g., Jones et al., 2002; Jones and Sayer, 2003; Roberts et al., 2003) and is argued to be a major factor in the failure of macrophytes to establish even decades after the start of nutrient loading reduction, despite suitable water clarity for plant re-establishment in spring (Phillips et al., 2005). In our adapted PCLake model, periphyton biomass was dependent on TP concentrations in the water, based on the positive correlation between chlorophyll content of periphyton on hard substrata and TP in the water column (Vadeboncoeur et al., 2006). In eutrophic shallow, temperate lakes, periphyton is often top-down controlled by a cascading effect from omnivorous fish that feed on periphyton grazers such as snails and chironomid larvae (Jones and Sayer, 2003). Thus, nutrient load reductions will only reduce periphyton shading after the fish biomass built up during the turbid period has also been reduced, which may take 10–15 years (Jeppesen et al., 2005). Furthermore, the observed dominant macrophyte species in the intermediate recovery phase after nutrient load reduction (Table 1) show little or no allelopathic activity that might hamper periphyton growth (Table 5), thus making them more susceptible to shading by periphyton.
Cyanobacteria have been shown to potentially inhibit submerged macrophyte growth via allelopathy (Zheng et al., 2013), but whether this mechanism contributes to the disappearance of macrophytes during the recovery phase in summer is unknown. Most of the dominant macrophyte species during intermediate recovery after nutrient load reductions are also highly susceptible to herbivory due to their low content of polyphenols, low carbon to nitrogen ratio and low dry matter content (Elger and Willby, 2003; Dorenbosch and Bakker, 2011, Table 5). Periphyton shading may further increase the sensitivity of macrophytes to herbivory (Hidding et al., 2016). Finally, fine-leaved species such as P. pectinatus, P. pusillus and Z. palustris also suffer from leaf plucking by omnivorous fish during periods of low zooplankton abundance when those fish switch to macroinvertebrate prey found in the periphyton of macrophytes (Körner and Dugdale, 2003). Such leaf plucking by fish can lead to a considerable leakage of nutrients from injured macrophyte tissue, thereby further stimulating phytoplankton growth (Hansson et al., 1987). Overall, while being well-suited for survival during turbid phases and for exploiting the clear-water conditions in spring for re-colonization, other traits of macrophyte species typical of the intermediate recovery phase following nutrient load reduction prevent their survival during later summer (Table 5).
Stable clear-water conditions in spring and summer with more diverse macrophyte vegetation were observed when both spring and summer TP concentrations reached about 0.05 mg L−1. This value corresponds well with a threshold for low cyanobacterial abundance in shallow lakes found by Jeppesen et al. (2005) and Triest et al. (2016) and the average critical loading for shifts from turbid to clear conditions estimated for Dutch shallow lakes by Janse (2005) and in eastern England by Phillips et al. (2015). Whether external nutrient load reductions alone were responsible for the observed low in-lake TP concentrations in the lakes in our survey that reached stable clear-water conditions, however, remains questionable. It seems that in most cases additional changes in either the fish community (Lake Veluwe, Galenbecker See) and/or exotic mussel invasions (Lake Müggelsee, Eemmeer) contributed to the observed trend. In Lake Veluwe, several severe winters, an increase in bream (Abramis brama) fisheries between 1993 and 1997 and the increase in zebra mussel densities are all thought to have contributed to a break in the dominance of cyanobacteria, thus allowing for the prevalence of stable clear-water conditions with charophyte dominance since 1996 (Noordhuis et al., 2016, Figure 4). Once established, these dense charophyte beds provide more efficient stabilizing mechanisms for clear-water conditions than rooted angiosperms (Blindow et al., 2014). Characeae also successfully replaced P. pectinatus in Lake Wolderwijd after biomanipulation (Figure 4), while in Swedish Lake Krankesjön a similar development has been observed, the reasons for which are unknown (Blindow, 1992; Hargeby et al., 1994; Hansson et al., 2010). In Lake Müggelsee and Eemmeer, the additional influence of a sudden invasion of the quagga mussel (Dreissena rostriformis bugensis) in around 2013 might have contributed to a decline in TP concentrations and increased water transparencies (Figure 4, S. Hilt, unpublished, Noordhuis et al., 2016). This species can colonize soft substrates and thus cover much larger areas than those previously occupied by the zebra mussel (D. polymorpha) (Karatayev et al., 2015). In Lake Eemmeer, quagga mussels filtered the lake volume about five times a day in 2013 (Noordhuis et al., 2016).
Macrophyte recovery in Steinhuder Meer and Langer See deviates from the suggested pattern in that both are dominated by species more typical of lakes having undergone restoration with internal measures (Table 1). In Steinhuder Meer, a strong reduction of the fish population has been observed which was attributed to cormorant activities (Niedersächsischer Landesbetrieb für Wasserwirtschaft, Küsten- und Naturschutz, 2011). Cormorant effects on fish populations are also suggested for Felbrigg Lake (C. Sayer, unpublished). Through a natural increase in cormorants, the lake food web configuration was likely affected in ways comparable to those of lakes undergoing biomanipulation. This makes Steinhuder Meer and Felbrigg Lake cases of external load reduction with added unintentional internal measures (natural biomanipulation) possibly accelerating recovery. Therefore, these cases show closer correspondence to macrophyte community patterns of lakes having undergone internal measures. They also illustrate that parallel biological processes may be at play in recovering lakes that need to be considered in unison to understand the speed and trajectory of macrophyte recovery.
Response of Macrophytes to Biomanipulation in Shallow Temperate Lakes
In contrast to lakes undergoing only reduced external nutrient loading, submerged macrophytes often respond very quickly in shallow lakes subjected to biomanipulation by fish removal (Hansson et al., 1998; Bakker et al., 2013), even at rather high nutrient concentrations (Figure 2). Macrophytes colonizing these lakes are often “pioneer” species, such as Elodea or Ceratophyllum, re-colonizing from either seeds, oospores or fragments and characterized by high growth rates (Tables 2, 4). Similar species have also been recorded in lakes following natural fish kills (Sayer et al., 2016) or implementation of other in-lake restoration measures such as sediment dredging and phosphorus precipitation (Table 2). Fish removal may indirectly (due to more periphyton grazing invertebrates) reduce periphyton shading in summer, a major mechanism preventing macrophyte survival after nutrient load reduction (see Response of Macrophytes to Nutrient Load Reductions). The relevance of this process for macrophyte recovery after fish removal has not yet been directly tested, although mesocosm trials in which the density of periphyton grazers are manipulated produce predictable outcomes in terms of periphyton biomass and macrophyte composition (Elger et al., 2009). Excretion of allelopathic substances, which has been detected for many of the typical species that colonize after biomanipulation (Table 5), may also contribute to lower periphyton densities.
In general, macrophyte species typically occurring after introduction of in-lake restoration measures allow for a longer period of high macrophyte cover and dampen seasonal changes in phytoplankton abundance as described for “stable” lakes prior to major eutrophication (Sayer et al., 2010b). Both, Elodea and charophyte species can remain evergreen in temperate lakes (e.g., Søndergaard et al., 2017), thus extending their positive influence on water quality to seasons outside the influence of annual species.
In many cases, however, mass developments of monocultures occur. Monocultures of Elodea or Ceratophyllum species are often unstable in terms of interannual persistence (Table 5) and can collapse leading to a shift back to turbid conditions as in, for instance, Lakes Zwemlust, Væng, and Alderfen Broad (Table 2). Characeae seem less often involved in sudden collapses, although, exceptions are known, for example Schlosssee Buggenhagen (Table 2) or Lake Botshol (Rip et al., 2007). If lakes remain clear for several consecutive years, which is usually only the case at lower nutrient concentrations, a more diverse macrophyte community develops (Table 2, Lauridsen et al., 2003a). Lauridsen et al. (2003b) assumed that differences in the success of biomanipulation in Danish and Dutch shallow lakes might be attributable to variation in pioneer macrophyte species; thus, Elodea and Potamogeton species, typical for Danish lakes, were preferred over charophytes by macrophyte-grazing waterfowl (Weisner et al., 1997). Indeed, increasing top-down control of periphyton-grazing invertebrates by omnivorous fish, which increase in abundance in the period after a biomanipulation, may render macrophytes more susceptible to herbivory (Hidding et al., 2016).
Conclusions and Implications for Lake Management
Our analyses suggest that the composition of the macrophyte community and their seasonal abundance in shallow lakes during recovery from turbid, highly eutrophic conditions often depends on remnant macrophyte stands, the specific restoration measure applied and additional stochastic influences on water clarity such as winter fish kills, cormorant predation on fish or introduction of invasive filter-feeding mussel populations. In turn, the prevailing macrophyte community can influence lake water quality.
Reductions in external nutrient loading often result in the re-occurrence of spring clear-water phases exploitable by a few macrophyte species (mainly pondweeds) with specific traits. Resistance to wave action permits survival during the turbid phase in very shallow areas, in particular in larger lakes. During recovery these plants germinate early in spring from energy-rich vegetative propagules and complete their life cycle in early summer, when phytoplankton takes over. This intermediate recovery phase may, in some cases, last for several decades before a more diverse and abundant submerged macrophyte community develops that stabilizes clear-water conditions during the entire potential growing season (Figure 1, Table 1). Our model simulations suggest that, if the premature termination of macrophyte growth can be prevented, the summer phytoplankton peak responsible for turbid water and potentially harmful algae blooms will also be reduced. Simulations also revealed that at high periphyton shading, the intermediate recovery phase is shifted to lower nutrient loads compared with a scenario with lower periphyton shading. Therefore, if periphyton shading can be reduced external restoration measures could potentially be effective at a higher nutrient load. Macrophyte recovery during the intermediate recovery state might be facilitated by establishing exclosures to protect certain areas from herbivory by birds and/or predation of periphyton grazers by omnivorous fish, a lake-wide biomanipulation of fish, or internal measures, such as TP precipitation to lower water column TP concentrations in summer. Additional, usually unintended internal changes, such as reductions in fish abundance by commercial fisheries, natural fish kills or exotic mussel invasions can facilitate a shift to clearer conditions in summer and further aid the establishment of a more diverse macrophyte community.
In contrast, fish stock reductions, natural fish kills and sediment removal via suction-dredging can, by themselves, temporarily restore clear-water conditions in spring and summer, even at high nutrient concentrations and then allow rapid colonization by pioneer macrophyte species from in situ seeds, oospores or vegetative fragments. Fish stock reductions might thus be a suitable short-term management strategy, but fish removal needs to be frequently repeated.
Lasting macrophyte recovery can only be achieved in combination with reduced nutrient loading (Figure 1), a need that is further accentuated under future climate change scenarios, where cyanobacterial shading of macrophytes will likely be more severe (Kosten et al., 2012). Although global, political measures against ongoing climate warming are slow, local restoration efforts may reduce the combined stress from e.g. eutrophication and climate warming (Moss et al., 2011; Scheffer et al., 2015), and thereby serve as a buffer against further deterioration of macrophyte beds and the ecosystem services that derive from lakes and reservoirs (Urrutia-Cordero et al., 2016).
Author Contributions
SH conceived the presented idea, wrote the manuscript and performed the literature research. MA, EB, IB, TD, L-AH, EJ, TK, AK, JK, TL, RN, GP, JR, H-HS, MS, KvdW, EvD, AW, NW, and CS provided lake data. MG, JJ, AJ, WM, and ST performed the modeling. All authors contributed to discussions and the writing of different parts of the text.
Funding
MA and MG were supported by the German Research Foundation (DFG, grant no. SU 623/1-1 and GRK 2032/1, respectively). AJ is supported by the Netherlands Environmental Assessment Agency (PBL) and ST by STOWA (grant no. 443.269). L-AH was supported by the BiodivERsA ERA-net LIMNOTIP. EJ, MS, TD, and TL were supported by MARS (Managing Aquatic ecosystems and water Resources under multiple Stress) funded under the 7th EU Framework Programme (Contract No.: 603378). JR was supported by the German Ministry of Education and Research (project NITROLIMIT, grant no. 033L041 A).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling Editor is currently co-organizing a Research Topic with one of the authors EB, and confirms the absence of any other collaboration.
Acknowledgments
We thank all technicians of IGB Berlin responsible for the long-term measuring program in Lake Müggelsee. Antje Barsch, Nadine Baadke (Landesumweltamt Brandenburg) and Antje Köhler (Senat Berlin) provided data for selected German lakes. The Environment Agency provided water quality data and the Broads Authority macrophyte data for the lakes in the Broads National Park (UK). We acknowledge linguistic improvements by Anne Mette Poulsen. We thank two reviewers for their helpful comments.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2018.00194/full#supplementary-material
References
Alderton, E., Sayer, C. D., Davies, R., Lambert, S. J., and Axmacher, J. C. (2017). Buried alive: Aquatic plants survive in “ghost ponds” under agricultural fields. Biol. Conserv. 212, 105–110. doi: 10.1016/j.biocon.2017.06.004
Annadotter, H., Cronberg, G., Aagren, R., Lundstedt, B., Nilsson, P. A., and Ströbäck, S. (1999). Multiple techniques for lake restoration. Hydrobiologia 395, 77–85. doi: 10.1023/A:1017011132649
Bakker, E. S., Sarneel, S. M., Gulati, R. D., Liu, Z., and Van Donk, E. (2013). Restoring macrophyte diversity in shallow temperate lakes: biotic versus abiotic constraints. Hydrobiologia 710, 23–37. doi: 10.1007/s10750-012-1142-9
Baldridge, A. K., and Lodge, D. M. (2014). Long-term studies of crayfish-invaded lakes reveal limited potential for macrophyte recovery from the seed bank. Freshw. Sci. 33, 788–797. doi: 10.1086/677070
Barrat-Segretain, M. H., and Bornette, G. (2000). Regeneration and colonization abilities of aquatic plant fragments: effect of disturbance seasonality. Hydrobiologia 421, 31–39. doi: 10.1023/A:1003980927853
Bergman, E., Hansson, L.-A., Persson, A., Strand, J., Romare, P., Enell, M., et al. (1999). Synthesis of theoretical and empirical experiences from nutrient and cyprinid reductions in Lake Ringsjön. Hydrobiologia 404, 145–156. doi: 10.1023/A:1003788900521
Bernes, C., Carpenter, S. R., Gårdmark, A., Larsson, P., Persson, L., Skov, C., et al. (2015). What is the influence of a reduction of planktivorous and benthivorous fish on water quality in temperate eutrophic lakes? A systematic review. Environ. Evidence 4, 7. doi: 10.1186/s13750-015-0032-9
Best, E. P. H. (1977). Seasonal changes in mineral and organic components of Ceratophyllum demersum and Elodea canadensis. Aquat. Bot. 3, 337–348. doi: 10.1016/0304-3770(77)90038-9
Best, E. P. H. (1982). The aquatic macrophytes of Lake Vechten: species composition, spatial distribution and production. Hydrobiologia 95, 65–77. doi: 10.1007/BF00044477
Best, E. P. H., and Dassen, J. H. A. (1987). A seasonal study of growth characteristics and the levels of carbohydrates and proteins in Elodea nuttallii, Polygonum amphibium and Phragmites australis. Aquat. Bot. 28, 353–372. doi: 10.1016/0304-3770(87)90011-8
Blindow, I. (1992). Long- and short-term dynamics of submerged macrophytes in two shallow eutrophic lakes. Freshw. Biol. 28, 15–27. doi: 10.1111/j.1365-2427.1992.tb00558.x
Blindow, I., Hargeby, A., and Hilt, S. (2014). Facilitation of clear-water conditions in shallow lakes by macrophytes: differences between charophyte and angiosperm dominance. Hydrobiologia 737, 99–110. doi: 10.1007/s10750-013-1687-2
Blindow, I., Hargeby, A., Wagner, B. M. A., and Andersson, G. (2000). How important is the crustacean plankton for the maintenance of water clarity in shallow lakes with abundant submerged vegetation? Freshw. Biol. 44, 185–197. doi: 10.1046/j.1365-2427.2000.00552.x
Blüml, V., Körner, F., Marxmeier, U., Richter, M., and Schönheim, A. (2008). Entwicklung und aktuelle Situation der Verlandungsvegetation des Dümmers (Niedersachsen). Osnabr. Naturwiss. Mitteil. 33/34, 19–46.
Bootsma, M. C., Barendregt, A., and Van Alphen, J. C. A. (1999). Effectiveness of reducing external nutrient load entering a eutrophicated shallow lake ecosystem in the Naardermeer nature reserve, The Netherlands. Biol. Conserv. 90, 193–201. doi: 10.1016/S0006-3207(99)00027-0
Brewer, C. A., and Parker, M. (1990). Adaptations of macrophytes to life in moving water: upslope limits and mechanical properties of stems. Hydrobiologia 194, 133–142. doi: 10.1007/BF00028414
Bytnerowicz, T. A., and Carruthers, R. I. (2014). Temperature-dependent models of Zannichellia palustris seed germination for application in aquatic systems. Environ. Exp. Bot. 104, 44–53. doi: 10.1016/j.envexpbot.2014.03.006
Capers, R. S. (2003). Macrophyte colonization in a freshwater tidal wetland (Lyme, CT, USA). Aquat. Bot. 77, 325–338. doi: 10.1016/j.aquabot.2003.08.001
Choi, C., Bareiss, C., Walenciak, O., and Gross, E. M. (2002). Impact of polyphenols on growth of the aquatic herbivore Acentria ephemerella. J. Chem. Ecol. 28, 2223–2238. doi: 10.1023/A:1021049332410
De Winton, M. D., Clayton, J. S., and Champion, P. D. (2000). Seedling emergence from seed banks of 15 New Zealand lakes with contrasting vegetation histories. Aquat. Bot. 66, 181–194. doi: 10.1016/S0304-3770(99)00074-1
Dorenbosch, M., and Bakker, E. S. (2011). Herbivory in omnivorous fishes: effect of plant secondary metabolites and prey stoichiometry. Freshw. Biol. 56, 1783–1797. doi: 10.1111/j.1365-2427.2011.02618.x
Elger, A., and Willby, N. J. (2003). Leaf dry matter content as an integrative expression of plant palatability: the case of freshwater macrophytes. Func. Ecol. 17, 58–65. doi: 10.1046/j.1365-2435.2003.00700.x
Elger, A., Willby, N. J., and Cabello-Martinez, M. (2009). Invertebrate grazing during the regenerative phase affects the ultimate structure of macrophyte communities. Freshw. Biol. 54, 1246–1255 doi: 10.1111/j.1365-2427.2009.02171.x
Gervais, F., Berger, S., Schönfelder, T., and Rusche, R. (1999). Basic limnological characteristics of the shallow eutrophic lake Grimnitzsee (Brandenburg, Germany). Limnologica 29, 105–119. doi: 10.1016/S0075-9511(99)80058-9
Grutters, B. M. C., Gross, E. M., Van Donk, E., and Bakker, E. S. (2017). Periphyton density is similar on native and non-native plant species. Freshw. Biol. 62, 906–915. doi: 10.1111/fwb.12911
Gulati, R. D., and Van Donk, E. (2002). Lakes in the Netherlands, their origin, eutrophication and restoration: state-of-the-art review. Hydrobiologia 478, 73–106. doi: 10.1023/A:1021092427559
Haag, R. W. (1983). Emergence of seedlings of aquatic macrophytes from lake sediments. Can. J. Bot. 61, 148–156. doi: 10.1139/b83-014
Hansson, L.-A., Annadotter, H., Bergman, E., Hamrin, S. F., Jeppesen, E., Kairesalo, T., et al. (1998). Biomanipulation as an application of food-chain theory: constraints, synthesis, and recommendations for temperate lakes. Ecosystems 1, 558–574. doi: 10.1007/s100219900051
Hansson, L.-A., Johansson, L., and Persson, L. (1987). Effects of fish grazing on nutrient release and succession of primary producers. Limnol. Oceanogr. 32, 723–729. doi: 10.4319/lo.1987.32.3.0723
Hansson, L.-A., Nicolle, A., Brönmark, C., Hargeby, A., Lindström, Å., and Andersson, G. (2010). Waterfowl, macrophytes, and the clear water state of shallow lakes. Hydrobiologia 646, 101–109. doi: 10.1007/s10750-010-0169-z
Hargeby, A., Andersson, G., Blindow, I., and Johansson, S. (1994). Trophic web structure in a shallow eutrophic lake during a dominance shift from phytoplankton to submerged macrophytes. Hydrobiologia 279, 83–90. doi: 10.1007/BF00027843
Heinzel, K., and Martin, C. (2006). Monitoring der Qualitätskomponente Makrophyten für WRRL und FFH-RL in schleswig-holsteinischen Seen; Gutachten im Auftrag des Landesamt für Natur und Umwelt des Landes Schleswig-Holstein. Available online at: http://www.umweltdaten.landsh.de/nuis/wafis/seen/Berichte_Gutachten/Ufer_Unterwasservegetation/Bericht_Makrophyten_2006_WRRL_Heinzel_GFN.pdf.
Hidding, B., Bakker, E. S., Hootsmans, M. J. M., and Hilt, S. (2016). Synergy between shading and herbivory triggers plant loss and regime shifts in aquatic systems. Oikos 125, 1489–1495. doi: 10.1111/oik.03104
Hidding, B., Bakker, E. S., Keuper, F., De Boer, T., De Vries, P. P., and Nolet, B. A. (2010). Differences in tolerance of pondweeds and charophytes to vertebrate herbivores in a shallow Baltic estuary. Aquat. Bot. 93, 123–128. doi: 10.1016/j.aquabot.2010.04.002
Hilt, S., Adrian, R., Köhler, J., Monaghan, M. T., and Sayer, C. (2013). Clear, crashing, turbid and back – long-term changes of macrophyte assemblages in a shallow lake. Freshw. Biol. 58, 2027–2036. doi: 10.1111/fwb.12188
Hilt, S., Brothers, S., Jeppesen, E., Veraart, A., and Kosten, S. (2017). Translating regime shifts in shallow lakes into changes in ecosystem functions and services. Bioscience 67, 928–936. doi: 10.1093/biosci/bix106
Hilt, S., and Gross, E. M. (2008). Can allelopathically active submerged macrophytes stabilise clear-water states in shallow eutrophic lakes? Bas. Appl. Ecol. 9, 422–432. doi: 10.1016/j.baae.2007.04.003
Hilt, S., Gross, E. M., Hupfer, M., Morscheid, H., Mählmann, J., Melzer, A., et al. (2006). Restoration of submerged vegetation in shallow eutrophic lakes – guideline and state of the art in Germany. Limnologica 36, 155–171. doi: 10.1016/j.limno.2006.06.001
Hilt, S., and Grünert, U. (2008). Praxistest zur Bewertung von Makrophyten in Berliner Seen im Rahmen der Umsetzung der EU-Wasserrahmenrichtlinie. Report for Senate (Berlin), 1–58.
Hilt, S., Van de Weyer, K., Köhler, A., and Chorus, I. (2010). Submerged macrophyte responses to reduced phosphorus concentrations in two peri-urban lakes. Rest. Ecol. 18, 452–461. doi: 10.1111/j.1526-100X.2009.00577.x
Hoare, D., Phillips, G., and Perrow, M. (2008). Broads Lake Restoration Strategy. Appendix 4. Review of Biomanipulaton. Norwich: Broads Authority, 1–59.
Hussner, A., Gross, E. M., Van de Weyer, K., and Hilt, S. (2014). Handlungsempfehlung zur Abschätzung der Chancen einer Wiederansiedlung von Wasserpflanzen bei der Restaurierung von Flachseen Deutschlands. Arbeitsblatt: Deutsche Gesellschaft für Limnologie (DGL),
Idestam-Almquist, J., and Kautsky, L. (1995). Plastic responses in morphology of Potamogeton pectinatus L. to sediment and above-sediment conditions at two sites in the northern Baltic proper. Aquat. Bot. 52, 205–216. doi: 10.1016/0304-3770(95)00499-8
Janse, J. H. (1997). A model of nutrient dynamics in shallow lakes in relation to multiple stable states. Hydrobiologia 342, 1–8. doi: 10.1023/A:1017018812215
Janse, J. H. (2005). Model Studies on the Eutrophication of Shallow Lakes and Ditches, Dissertation, Wageningen University.
Janse, J. H., De Senerpont Domis, L. N., Scheffer, M., Lijklema, L., Van Liere, L., Klinge, M., et al. (2008). Critical phosphorus loading of different types of shallow lakes and the consequences for management estimated with the ecosystem model PCLake. Limnologica 38, 203–219. doi: 10.1016/j.limno.2008.06.001
Janse, J. H., Scheffer, M., Lijklema, L., Van Liere, L., Sloot, J. S., and Mooij, W. M. (2010). Estimating the critical phosphorus loading of shallow lakes with the ecosystem model PCLake: sensitivity, calibration and uncertainty. Ecol. Model. 221, 654–665. doi: 10.1016/j.ecolmodel.2009.07.023
Janse, J. H., Van Donk, E., and Aldenberg, T. (1998). A model study on the stability of the macrophyte-dominated state as affected by biological factors. Wat. Res. 32, 2696–2706. doi: 10.1016/S0043-1354(98)00049-9
Janssen, A. B. G., de Jager, V. C. L., Janse, J. H., Kong, X., Liu, S., Ye, Q., et al. (2017). Spatial identification of critical nutrient loads of large shallow lakes: implications for Lake Taihu (China). Wat. Res. 119, 276–287. doi: 10.1016/j.watres.2017.04.045
Jeppesen, E., Kristensen, P., Jensen, J. P., Søndergaard, M., Mortensen, E., and Lauridsen, T. (1991). Recovery resilience following a reduction in external phosphorus loading of shallow, eutrophic Danish lakes: duration, regulating factors and methods for overcoming resilience. Mem. Ist. Ital. Idrobiol. 48, 127–148.
Jeppesen, E., Søndergaard, M., Meerhoff, M., Lauridsen, T. L., and Jensen, J. P. (2007a). Shallow lake restoration by nutrient loading reduction – some recent findings and challenges ahead. Hydrobiologia 584, 239–252. doi: 10.1007/s10750-007-0596-7
Jeppesen, E., Meerhoff, M., Jacobsen, B. A., Hansen, R. S., Søndergaard, M., Jensen, J. P., et al. (2007b). Restoration of shallow lakes by nutrient control and biomanipulation - the successful strategy varies with lake size and climate. Hydrobiologia 581, 269–285. doi: 10.1007/s10750-006-0507-3
Jeppesen, E., Søndergaard, M., Jensen, J. P., Havens, K., Anneville, O., Carvalho, L., et al. (2005). Lake responses to reduced nutrient loading – an analysis of contemporary data from 35 European and North American long term studies. Freshw. Biol. 50, 1747–1771. doi: 10.1111/j.1365-2427.2005.01415.x
Jeppesen, E., Søndergaard, M., Lauridsen, T. L., Davidson, T. A., Liu, Z., Mazzeo, N., et al. (2012). Biomanipulation as a response tool to combat eutrophication: recent advances and future challenges. Adv. Ecol. Res. 47, 411–488. doi: 10.1016/B978-0-12-398315-2.00006-5
Jones, J. I., and Sayer, C. D. (2003). Does fish-invertebrate-periphyton cascade precipitate plant loss in shallow lakes? Ecology 84, 2155–2167. doi: 10.1890/02-0422
Jones, J. I., Young, J. O., Eaton, J. W., and Moss, B. (2002). The influence of nutrient loading, dissolved inorganic carbon and higher trophic levels on the interaction between submerged plants and periphyton. J. Ecol. 90, 12–24. doi: 10.1046/j.0022-0477.2001.00620.x
Josefsson, M. (2011). NOBANIS - Invasive Species Fact Sheet - Elodea canadensis, Elodea nuttallii and Elodea callitrichoides – NOBANIS. Available online at: www.nobanis.org. Date of Access (Accessed Nov 28, 2012).
Kabus, T., Schumann, M., Weiß, G., and Kalhoff, M. (2007). Beitrag zur Flora der Havel und angrenzender Flächen zwischen Potsdam und Pritzerbe. Verh. Bot. Ver. Berlin Brandenburg 140, 25–55.
Kabus, T., and Mauersberger, R. (2011). Liste und Rote Liste der Armleuchteralgengewächse (Characeae) des Landes Brandenburg. Natursch. Landschaftspfl. Brbg. 20, 1–32.
Kaplan, K., and Muer, T. (1990). Beobachtungen zum diasporenreservoir im Bereich ehemaliger Heideweiher. Florist. Rundbriefe 24, 38–45.
Karatayev, A. Y., Burlakova, L. E., and Padilla, D. K. (2015). Zebra versus quagga mussels: a review of their spread, population dynamics, and ecosystem impacts. Hydrobiologia 746, 97–112. doi: 10.1007/s10750-014-1901-x
Kautsky, L. (1990). Seed and tuber banks of aquatic macrophytes in thE Askö area, northern Baltic proper. Holarct. Ecol. 13, 143–148. doi: 10.1111/j.1600-0587.1990.tb00600.x
Knösche, R. (2008). Wiederfund von Najas marina L. spp. marina im Schollener See (Elbe-Havel-Winkel, Sachsen-Anhalt). Mitteil. Florist. Kart. Sachsen Anhalt 13, 41–51.
Körner, S. (2001). Development of submerged macrophytes in shallow Lake Müggelsee (Berlin, Germany) before and after its switch to the phytoplankton-dominated state. Arch. Hydrobiologia 152, 395–409. doi: 10.1127/archiv-hydrobiol/152/2001/395
Körner, S. (2002). Loss of submerged macrophytes in shallow lakes in North-Eastern Germany. Intern. Rev. Hydrobiologia 87, 375–384. doi: 10.1002/1522-2632(200207)87:4<375::AID-IROH375>3.0.CO;2-7
Körner, S., and Dugdale, T. (2003). Is roach herbivory preventing re-colonization of a shallow lake with submerged macrophytes? Hydrobiologia 506, 497–501. doi: 10.1023/B:HYDR.0000008561.67513.ec
Kosten, S., Huszar, V. L. M., Bécares, E., Costa, L., Van Donk, E., Hansson, L.-A., et al. (2012). Warmer climates boost cyanobacterial dominance in shallow lakes. Glob. Chang. Biol. 18, 118–126. doi: 10.1111/j.1365-2486.2011.02488.x
Kuiper, J. J. (2016). Making Eco Logic and Models work: An Integrative Approach to Lake Ecosystem Modelling, Dissertation, Wageningen University.
Kuiper, J. J., Verhofstad, M. J., Louwers, E. L., Bakker, E. S., Brederveld, R. J., Van Gerven, L. P. A., et al. (2017). Mowing submerged macrophytes in shallow lakes with alternative stable states: battling the good guys? Environ. Manage. 59, 619–634. doi: 10.1007/s00267-016-0811-2
Lage, S., Annadotter, H., Rasmussen, U., and Rydberg, S. (2015). Biotransfer of β-N-methylamino-L-alanine (BMAA) in a eutrophicated freshwater lake. Mar. Drugs 13, 1185–1201 doi: 10.3390/md13031185
Lalonde, S., and Downing, J. A. (1991). Epiphyton biomass is related to lake trophic status, depth, and macrophyte architecture. Can. J. Fish. Aquat. Sci. 48, 2285–2291. doi: 10.1139/f91-268
Landesamt für Natur und Umwelt des Landes Schleswig-Holstein (1997). Zustand und Belastungsquellen des Warder Sees. Flintbek: Landesamt für Natur und Umwelt des Landes Schleswig-Holstein.
Lauridsen, T. L., Jensen, J. P., Jeppesen, E., and Sondergaard, M. (2003a). Response of submerged macrophytes in Danish lakes to nutrient loading reductions and biomanipulation. Hydrobiologia 506, 641–649. doi: 10.1023/B:HYDR.0000008633.17385.70
Lauridsen, T. L., Sandsten, H., and Hald Møller, P. (2003b). Restoration of a shallow lake by introducing Potamogeton spp.: impact of waterfowl grazing. Lakes Reserv. Res. Manage. 8, 177–187. doi: 10.1111/j.1440-1770.2003.00224.x
Lauridsen, T., Pedersen, L. J., Jeppesen, E., and Søndergaard, M. (1996). The importance of macrophyte bed size for composition and horizontal migration of cladocerans in a shallow lake. J. Plankt. Res. 18, 2283–2294. doi: 10.1093/plankt/18.12.2283
Lillie, R. A., Budd, J., and Rasmussen, P. W. (1997). Spatial and temporal variability in biomass density of Myriophyllum spicatum L. in a northern temperate lake. Hydrobiologia 347, 69–74. doi: 10.1023/A:1003059000277
Madsen, J. D. (1991). Resource allocation at the individual plant level. Aquat. Bot. 41, 67–86. doi: 10.1016/0304-3770(91)90039-8
Madsen, J. D., and Adams, M. S. (1988). The germination of Potamogeton pectinatus tubers: environmental control by temperature and light. Can. J. Bot. 66, 2523–2526. doi: 10.1139/b88-343
Mathes, J. (2007). Erfahrungen mit Seentherapien in Mecklenburg-Vorpommern. Available online at: http://www.lake-jewel.net/download/Dateien/Seesanierung%20Mecklenburg-Vorpommern.pdf
Mauersberger, H., and Mauersberger, R. (1996). Die Seen im Biosphärenreservat Schorfheide-Chorin: eine ökologische Studie. Dissertation, Universität Greifswald.
Meijer, M. L., De Boois, I., Scheffer, M., Portielje, R., and Hosper, H. (1999). Biomanipulation in shallow lakes in The Netherlands: an evaluation of 18 case studies. Hydrobiologia 408, 13–30. doi: 10.1023/A:1017045518813
Meijer, M. L., and Hosper, H. (1997). Effects of biomanipulation in the large and shallow Lake Wolderwijd, The Netherlands. Hydrobiologia 342, 335–349. doi: 10.1023/A:1017088701621
Mooij, W. M., Brederveld, R. J., De Klein, J. J. M., DeAngelis, D. L., Downing, A. S., Faber, M., et al. (2014). Serving many at once: how a database approach can create unity in dynamical ecosystem modelling. Environ. Mod. Softw. 61, 266–273. doi: 10.1016/j.envsoft.2014.04.004
Mooij, W. M., Janse, J. H., De Senerpont Domis, L. N., Hülsmann, S., and Ibelings, B. W. (2007). Predicting the effect of climate change on temperate shallow lakes with the ecosystem model PCLake. Hydrobiologia 584, 443–454. doi: 10.1007/s10750-007-0600-2
Moss, B., Balls, H., Irvine, K., and Stansfield, J. (1986). Restoration of two lowland lakes by isolation from nutrient-rich water sources with and without removal of sediment. J. Appl. Ecol. 23, 391–414. doi: 10.2307/2404025
Moss, B., Kosten, S., Meerhoff, M., Battarbee, R. W., Jeppesen, E., Mazzeo, N., et al. (2011). Allied attack: climate change and nutrient pollution. Inland Wat. 1, 101–105. doi: 10.5268/IW-1.2.359
Moss, B., Stansfield, J., and Irvine, K. (1990). Problems in the restoration of a hypertrophic lake by diversion of a nutrient-rich inflow. Verh. Intern. Ver. Limnol. 24, 568–572. doi: 10.1080/03680770.1989.11898801
Nixdorf, B., Rücker, J., Dolman, A. M., Wiedner, C., Hilt, S., Kasprzak, P., et al. (2013). Prozessverständnis als Grundlage für die Gewässerbewirtschaftung – Fallbeispiele für Limitation, Konkurrenz, Gewässerstruktur und Nahrungsnetzsteuerung. Korr. Wasserwirtschaft 60, 693–701.
Niedersächsischer Landesbetrieb für Wasserwirtschaft, Küsten- und Naturschutz, NLWKN. (2011). Leitfaden Maßnahmenplanung Oberflächengewässer. Wasserrahmenrichtlinie Band 3 Steinhuder Meer. 1–33.
Noordhuis, R., Van Zuidam, B. G., Peeters, E. T. H. M., and Van Geest, G. J. (2016). Further improvements in water quality of the Dutch Border lakes: two types of clear states at different nutrient levels. Aquatic Ecol. 50, 521–539. doi: 10.1007/s10452-015-9521-8
Perrow, M. R., Schutten, J. H., Howes, J. R., Holzer, T., Madgwick, F. J., and Jowitt, A. J. D. (1997). Interactions between coot (Fulica atra) and submerged macrophytes: the role of birds in the restoration process. Hydrobiologia 342, 241–255. doi: 10.1023/A:1017007911190
Phillips, G., Bennion, H., Perrow, M., Sayer, C. D., Spears, B., and Willby, N. (2015). A Review of Lake Restoration Practices and their Performance in the Broads National Park, 1980-2013. Report for the Broads Authority, Norwich and Natural England. Available online at: http://www.broads-authority.gov.uk/news-and-publications/publications-and-reports/conservation-publications-and-reports/water-conservation-reports
Phillips, G., Kelly, A., Pitt, J. A., Sanderson, R., and Taylor, E. (2005). The recovery of a very shallow eutrophic lake, 20 years after the control of effluent derived phosphorus. Freshw. Biol. 50, 1628–1638. doi: 10.1111/j.1365-2427.2005.01434.x
Phillips, G. L., Willby, N., and Moss, B. (2016). Submerged macrophyte decline in shallow lakes: what have we learnt in the last forty years? Aquat. Bot. 135, 37–45. doi: 10.1016/j.aquabot.2016.04.004
Portielje, R., and Van der Molen, D. T. (1999). Relationships between eutrophication variables: from nutrient loading to transparency. Hydrobiologia 408, 375–387. doi: 10.1023/A:1017090931476
Pot, R., and Ter Heerdt, G. N. J. (2014). Succession dynamics of aquatic lake vegetation after restoration measures: increased stability after 6 years of development. Hydrobiologia 737, 333–345. doi: 10.1007/s10750-014-1835-3
Riegman, R. (2007). Evaluatie Rapport Vijftien jaar Actief Biologisch Beheer (ABB) in het Duinigermeer (in Dutch). Reest aen Wieden Waterschap.
Rip, W. J., Ouboter, R. L., and Los, H. L. (2007). Impact of climatic fluctuations on Characeae biomass in a shallow, restored lake in The Netherlands. Hydrobiologia 584, 415–424. doi: 10.1007/s10750-007-0608-7
Roberts, E., Kroker, J., Körner, S., and Nicklisch, A. (2003). The role of periphyton during the re-colonization of a shallow lake with submerged macrophytes. Hydrobiologia 506, 525–530. doi: 10.1023/B:HYDR.0000008560.73832.1c
Rodrigo, M. A., Rojo, C., Alonso-Guillén, J. L., and Vera, P. (2013). Restoration of two small Mediterranean lagoons: the dynamics of submerged macrophytes and factors that affect the success of revegetation. Ecol. Eng. 54, 1–15. doi: 10.1016/j.ecoleng.2013.01.022
Rørslett, B., Berge, D., and Johansen, S. W. (1985). Mass invasion of Elodea canadensis in a mesotrophic, South Norwegian lake – impact of water quality. Verh. Intern. Ver. Limnol. 22, 2920–2926. doi: 10.1080/03680770.1983.11897803
Rücker, J., Barsch, A., and Nixdorf, B. (2015). Besser, aber noch nicht gut“ – Ökologischer Zustand der Seen in Brandenburg 2014. Wasserwirtschaft 12, 41–47.
Sand-Jensen, K., Pedersen, N. L., Thorsgaard, I., Moeslund, B., Borum, J., and Brodersen, K. P. (2008). 100 years of vegetation decline and recovery in Lake Fure, Denmark. J. Ecol. 96, 206–271. doi: 10.1111/j.1365-2745.2007.01339.x
Sayer, C. D., Burgess, A., Kari, K., Davidson, T. A., and Rose, N. (2010a). Long-term dynamics of submerged macrophytes and algae in a small and shallow, eutrophic lake: implications for the stability of macrophyte-dominance. Freshw. Biol. 55, 565–583. doi: 10.1111/j.1365-2427.2009.02353.x
Sayer, C. D., Davidson, T. A., and Jones, J. I. (2010b). Seasonal dynamics of macrophytes and phytoplankton in shallow lakes: a eutrophication-driven pathway from plants to plankton? Freshw. Biol. 55, 500–513. doi: 10.1111/j.1365-2427.2009.02365.x
Sayer, C. D., Davidson, T. A., Rawcliffe, R., Langdon, P. G., Leavitt, P. R., Cockerton, G., et al. (2016). Consequences of fish kills for long-term trophic structure in shallow lakes: implications for theory and restoration. Ecosystems 16, 1289–1309. doi: 10.1007/s10021-016-0005-z
Scheffer, M., Barrett, S., Carpenter, S. R., Folke, C., Green, A. J., Holmgren, M., et al. (2015). Creating a safe operating space for iconic ecosystems. Science 347, 1317–1319. doi: 10.1126/science.aaa3769
Scheffer, M., and Carpenter, S. R. (2003). Catastrophic regime shifts in ecosystems: linking theory to observation. Trends Ecol. Evol. 18, 648–656. doi: 10.1016/j.tree.2003.09.002
Scheffer, M., De Redelijkheid, M. R., and Noppert, F. (1992). Distribution and dynamics of submerged vegetation in a chain of shallow eutrophic lakes. Aquat. Bot. 42, 199–216. doi: 10.1016/0304-3770(92)90022-B
Scheffer, M., Hosper, S. H., Meijer, M. L., Moss, B., and Jeppesen, E. (1993). Alternative equilibria in shallow lakes. Trends Ecol. Evol. 8, 275–279. doi: 10.1016/0169-5347(93)90254-M
Schutten, J., Dainty, J., and Davy, A. J. (2005). Root anchorage and its significance for submerged plants in shallow lakes. J. Ecol. 93, 556–571. doi: 10.1111/j.1365-2745.2005.00980.x
Simberloff, D., and Gibbons, L. (2004). Now you see them, now you don't! – population crashes of established introduced species. Biol. Invasions 6, 161–172. doi: 10.1023/B:BINV.0000022133.49752.46
Simpson, D. A. (1990). Displacement of Elodea canadensis Michx. by Elodea nuttallii (Planch.) St. John in the British Isles. Watsonia 18, 173–177.
Sommer, U., Adrian, R., De Senerpont Domis, L. N., Elser, J. J., Gaedke, U., Ibelings, B. W., et al. (2012). Beyond the Plankton Ecology Group (PEG) Model: mechanisms driving plankton succession. Ann. Rev. Ecol. Evol. Syst. 43, 429–448. doi: 10.1146/annurev-ecolsys-110411-160251
Søndergaard, M., Bjerring, R., and Jeppesen, E. (2013). Persistent internal phosphorus loading in shallow eutrophic lakes. Hydrobiologia 710, 95–107. doi: 10.1007/s10750-012-1091-3
Søndergaard, M., Larsen, S. E., Johansson, L. S., Lauridsen, T. L., and Jeppesen, E. (2016). Ecological classification of lakes: uncertainty and the influence of year-to-year variability. Ecol. Indic. 61, 248–257. doi: 10.1016/j.ecolind.2015.09.024
Søndergaard, M., Lauridsen, T. L., Johansson, L. S., and Jeppesen, E. (2017). Repeated fish removal to restore lakes: case study of Lake Væng, Denmark-two biomanipulations during 30 years of monitoring. Water 9, 43. doi: 10.3390/w9010043
Søndergaard, M., Liboriussen, L., Pedersen, A. R., and Jeppesen, E. (2008). Lake restoration by fish removal: short- and long-term effects in 36 Danish lakes. Ecosystems 11, 1291–1305. doi: 10.1007/s10021-008-9193-5
Spencer, D. F. (1986). Early growth of Potamogeton pectinatus L. in response to temperature and irradiance: morphology and pigment composition. Aquat. Bot. 26, 1–8. doi: 10.1016/0304-3770(86)90002-1
Strand, J. (1999). The development of submerged macrophytes in Lake Ringsjön after biomanipulation. Hydrobiologia 404, 113–121. doi: 10.1023/A:1003728730563
Strand, J. A., and Weisner, S. E. B. (2001). Dynamics of submerged macrophyte populations in response to biomanipulation. Freshw. Biol. 46, 1397–1408. doi: 10.1046/j.1365-2427.2001.00746.x
Talling, J. F., and Parker, J. E. (2002). Seasonal dynamics of phytoplankton and phytobenthos, and associated chemical interactions, in a shallow upland lake (Malham Tarn, northern England). Hydrobiologia 487, 167–181. doi: 10.1023/A:1022934705359
Tobiessen, P., and Snow, P. D. (1984). Temperature and light effects on the growth of Potamogeton crispus in Collins Lake, New York State. Can. J. Bot. 62, 2822–2826. doi: 10.1139/b84-376
Triest, L., Stiers, I., and Van Onsem, S. (2016). Biomanipulation as a nature-based solution to reduce cyanobacterial blooms. Aquat. Ecol. 50, 461–483. doi: 10.1007/s10452-015-9548-x
Urrutia-Cordero, P., Ekvall, M. K., and Hansson, L. A. (2016). Local food web management increases resilience and buffers against global change effects on freshwaters. Sci. Rep. 6:29542. doi: 10.1038/srep29542
Vadeboncoeur, Y., Kalff, J., Christoffersen, K., and Jeppesen, E. (2006). Substratum as a driver of variation in periphyton chlorophyll and productivity in lakes. J. N. Am. Benth. Soc. 25, 379–392. doi: 10.1899/0887-3593(2006)25[379:SAADOV]2.0.CO;2
Van Berkum, J. A., Klinge, M., and Grimm, M. P. (1995). Biomanipulation in the duinigermeer, first results. Neth. J. Aquat. Ecol. 29, 81–90. doi: 10.1007/BF02061791
Van de Bund, W. J., and Van Donk, E. (2002). Short-term and long-term effects of zooplanktivorous fish removal in a shallow lake: a synthesis of 15 years of data from Lake Zwemlust. Freshw. Biol. 47, 2380–2387. doi: 10.1046/j.1365-2427.2002.01006.x
Van de Haterd, R. J. W., and Ter Heerdt, G. N. J. (2007). Potential for the development of submerged macrophytes in eutrophicated shallow peaty lakes after restoration measures. Hydrobiologia 584, 277–290. doi: 10.1007/s10750-007-0593-x
Van den Berg, M. S., Scheffer, M., Coops, H., and Simons, J. (1998). The role of characean algae in the management of eutrophic shallow lakes. J. Phycol. 34, 750–756. doi: 10.1046/j.1529-8817.1998.340750.x
Van den Berg, M. S., Scheffer, M., Van Nes, E., and Coops, H. (1999). Dynamics and stability of Chara sp. and Potamogeton pectinatus in a shallow lake changing in eutrophication level. Hydrobiologia 408, 335–342. doi: 10.1023/A:1017074211970
Van de Weyer, K. (2011). Erfassung und Bewertung der Makrophytenvegetation in der Unteren Havel und im Wannsee im Jahr 2011. Report for Senate (Berlin). 1–111.
Van Dijk, G. M. (1993). Dynamics and attenuation characteristics of periphyton upon artificial substratum under various light conditions and some additional observations on periphyton upon Potamogeton pectinatus L. Hydrobiologia 252, 143–161. doi: 10.1007/BF00008152
Van Dijk, G. M., and Janse, J. H. (1993). Modeling resource allocation in Potamogeton pectinatus L. J. Aquat. Plant Manage. 31, 128–134.
Van Dijk, G. M., and Van Vierssen, W. (1991). Survival of a Potamogeton pectinatus L.population under various light conditions in a shallow eutrophic lake (Lake Veluwe) in The Netherlands. Aquat. Bot. 39, 121–129. doi: 10.1016/0304-3770(91)90027-3
Van Vierssen, W. (1982a). The ecology of communities dominated by Zannichellia taxa in Western Europe. 1. Characterization and autecology of the Zannichellia taxa. Aquat. Bot. 12, 103–155. doi: 10.1016/0304-3770(82)90010-9
Van Vierssen, W. (1982b). The ecology of communities dominated by Zannichellia taxa in Western Europe. II. Distribution, synecology and productivity aspects in relation to environmental factors. Aquat. Bot. 13, 385–483. doi: 10.1016/0304-3770(82)90073-0
Van Vierssen, W. (1982c). Some notes on the germination of seeds of Najas marina L. Aquat. Bot. 12, 201–203. doi: 10.1016/0304-3770(82)90015-8
Van Wijk, R. J. (1988). Ecological studies on Potamogeton pectinatus L. I. General characteristics, biomass production and life cycles under field conditions. Aquat. Bot. 31, 211–258. doi: 10.1016/0304-3770(88)90015-0
Van Wijk, R. J. (1989). Ecological studies on Potamogeton pectinatus L. 3. Reproductive strategies and germination ecology. Aquat. Bot. 33, 271–299. doi: 10.1016/0304-3770(89)90042-9
Vari, A., and Toth, V. R. (2017). Quantifying macrophyte colonisation strategies - A field experiment in a shallow lake (Lake Balaton, Hungary). Aquat. Bot. 136, 56–60. doi: 10.1016/j.aquabot.2016.09.006
Verhofstad, M. J. J. M., Alirangues Núñez, M. M., Reichman, E. P., van Donk, E., Lamers, L. P. M., and Bakker, E. S. (2017). Mass development of monospecific submerged macrophyte vegetation after the restoration of shallow lakes: roles of light, sediment nutrient levels, and propagule density. Aquat. Bot. 141, 29–38. doi: 10.1016/j.aquabot.2017.04.004
Verpoorter, C., Kutser, T., Seekell, D. A., and Tranvik, L. J. (2014). A global inventory of lakes based on high-resolution satellite imagery. Geophys. Res. Lett. 41, 6396–6402. doi: 10.1002/2014GL060641
Wang, X., Qin, B., Gao, G., and Paerl, H. W. (2010). Nutrient enrichment and selective predation by zooplankton promote Microcystis (Cyanobacteria) bloom formation. J. Plankt. Res. 32, 457–470. doi: 10.1093/plankt/fbp143
Waterstraat, A. (2008). Die Fischfauna des Galenbecker Sees und seiner Zu- und Abflüsse. In: 70 Jahre Naturschutzgebiet Galenbecker See. Staatliches Amt für Umwelt und Natur Ueckermünde. 27–34.
Weisner, S. E., Strand, J. A., and Sandsten, H. (1997). Mechanisms regulating abundance of submerged vegetation in shallow eutrophic lakes. Oecologia 109, 592–599. doi: 10.1007/s004420050121
Wolfer, S. R., and Straile, D. (2004). Spatio-temporal dynamics and plasticity of clonal architecture in Potamogeton perfoliatus. Aquat. Bot. 78, 307–318. doi: 10.1016/j.aquabot.2003.11.005
Woolf, T. E., and Madsen, J. D. (2003). Seasonal biomass and carbohydrate allocation patterns in southern Minnesota curlyleaf pondweed populations. J. Aquat. Plant Manage. 41, 113–118.
Keywords: aquatic plants, biomanipulation, eutrophication, lake restoration. nutrient load reduction, PCLake, plant traits, regime shift
Citation: Hilt S, Alirangues Nuñez MM, Bakker ES, Blindow I, Davidson TA, Gillefalk M, Hansson L-A, Janse JH, Janssen ABG, Jeppesen E, Kabus T, Kelly A, Köhler J, Lauridsen TL, Mooij WM, Noordhuis R, Phillips G, Rücker J, Schuster H-H, Søndergaard M, Teurlincx S, van de Weyer K, van Donk E, Waterstraat A, Willby N and Sayer CD (2018) Response of Submerged Macrophyte Communities to External and Internal Restoration Measures in North Temperate Shallow Lakes. Front. Plant Sci. 9:194. doi: 10.3389/fpls.2018.00194
Received: 28 September 2017; Accepted: 01 February 2018;
Published: 19 February 2018.
Edited by:
Janne Alahuhta, University of Oulu, FinlandReviewed by:
Rebecca Lester, Deakin University, AustraliaLudwig Triest, Vrije Universiteit Brussel, Belgium
Copyright © 2018 Hilt, Alirangues Nuñez, Bakker, Blindow, Davidson, Gillefalk, Hansson, Janse, Janssen, Jeppesen, Kabus, Kelly, Köhler, Lauridsen, Mooij, Noordhuis, Phillips, Rücker, Schuster, Søndergaard, Teurlincx, van de Weyer, van Donk, Waterstraat, Willby and Sayer. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sabine Hilt, hilt@igb-berlin.de