Age-related olfactory decline is associated with the BDNF val66met polymorphism: evidence from a population-based study
- 1 Department of Psychology, Stockholm University, Stockholm, Sweden
- 2 Stockholm Brain Institute, Stockholm, Sweden
- 3 Department of Pharmacology, Göteborg University, Göteborg, Sweden
- 4 Department of Radiation Sciences and Integrative Medical Biology, Umeå, Sweden
The present study investigates the effect of the brain-derived neurotrophic factor (BDNF) val66met polymorphism on change in olfactory function in a large scale, longitudinal population-based sample (n = 836). The subjects were tested on a 13 item force-choice odor identification test on two test occasions over a 5-year-interval. Sex, education, health-related factors, and semantic ability were controlled for in the statistical analyses. Results showed an interaction effect of age and BDNF val66met on olfactory change, such that the magnitude of olfactory decline in the older age cohort (70–90 years old at baseline) was larger for the val homozygote carriers than for the met carriers. The older met carriers did not display larger age-related decline in olfactory function compared to the younger group. The BDNF val66met polymorphism did not affect the rate of decline in the younger age cohort (45–65 years). The findings are discussed in the light of the proposed roles of BDNF in neural development and maintenance.
Introduction
Of perceptual and cognitive functions, olfactory abilities are among the most sensitive to age-related impairments (Doty et al., 1984 ; Stevens and Cain, 1987 ). According to normative studies, that used cued identification of common household odors as a measure of functional olfactory ability, a majority of community-dwelling elderly are impaired (Murphy et al., 2002 ; Brämerson et al., 2004 ). During the last two decades, the scientific understanding of the structure and function of the olfactory sense has developed greatly, in part due to an increasing knowledge about the genetics of the olfactory system (Keller and Vosshall, 2008 ). However, although normal genetic variability in the population is likely to influence olfactory abilities (Whissel-Buechy and Amoore, 1973 ), knowledge is sparse regarding specific genetic influences underlying individual differences in human olfaction (Keller et al., 2007 ). To our knowledge, no previous study has reported genetic effects on longitudinal change in general olfactory function. Here, we targeted the brain-derived neurotrophic factor (BDNF) as a potential moderator of age-associated olfactory impairment.
Brain-derived neurotrophic factor is a member of the neurotrophin family, representing growth factors that support neuronal survival, transmission, and synaptic plasticity in the central nervous system (CNS) (Poo, 2001 ). A specific single nucleotide polymorphism of the BDNF, the val66met, modulates intracellular trafficking and activity-dependent secretion of BDNF protein (Poo, 2001 ). The val66met is located at nucleotide 196 (G/A) in the pro-protein of BDNF and produces an amino acid substitution, valine to methionine. The met allele inhibits the intracellular trafficking and the regulated secretion of the BDNF protein at synapses (Egan et al., 2003 ). Several lines of evidence suggest that BDNF might be relevant for olfactory processing. It has been found to regulate the proliferation and survival of olfactory receptor neurons (ORN) in vivo (Simpson et al., 2002 ). In addition, the expression of BDNF is high in various parts of the CNS, including the hippocampus and the olfactory bulb (OB) although the level of expression may decrease with increasing age (Katoh-Semba et al., 1998 ). There is some support linking the polymorphism to olfactory functioning in animals. Bath et al. (2008) identified the BDNF val66met variant as a critical factor in the disruption of OB neurogenesis in adult BDNF and tyrosine receptor kinase (TkrB) knock-out mice. The results suggest that an absence of BDNF and its receptor TkrB results in olfactory impairment. While BDNF appears necessary for the integrity of olfactory function in mice, little is known about the possible influence of the BDNF val66met polymorphism on olfactory function in humans. Behavioral association studies have demonstrated a role of the val66met polymorphism on human cognitive function. For example, individual variance in episodic memory performance and hippocampal activity has been related to the allelic combination of val66met (Egan et al., 2003 ). Subjects with at least one met allele have lower declarative memory performance accompanied with reduced hippocampal engagement, and smaller hippocampal volume (Egan et al., 2003 ; Hariri et al. 2003 ; Pezawas et al., 2004 ; Bueller et al., 2006 ). Although studies have reported that the met allele is a risk factor for cognitive impairment also in older adults (Miyajima et al., 2008 ; Nagel et al., 2008 ; Raz et al., 2009 ), opposite findings of poorer cognitive performance in older val homozygotes compared to met carriers have been reported, suggesting that the met allele may protect against cognitive decline (Erickson et al., 2008 ; Harris et al., 2006 ).
To the best of our knowledge, no study has investigated the effects of val66met on human olfactory performance. BDNF has been linked to Alzheimer’s and Parkinson’s disorders, which are both characterized by olfactory impairment (Phillips et al., 1991 ; Mesholam et al., 1998 ; Siegel and Chauhan, 2000 ). Although evidence is mixed, the val/val carriers of the val66met polymorphism were associated with higher prevalence rates for Alzheimer’s disease (AD) (Ventriglia et al., 2002 ; Combarros et al., 2004 ). Since sub-clinical levels of AD-related neuropathology is probably present in a majority of elderly individuals (del Tredici and Braak, 2008 ), and since the disorder affects olfactory function also in preclinical stages (Bacon et al., 1998 ; Tabert et al., 2005 ; Olofsson et al., 2010 ), effects of neuropathology are related to olfactory identification ability in community-dwelling elderly (Wilson et al., 2007 ).
The general aim of this study was to investigate the possible relation between age-related decline in olfactory function and the BDNF val66met genotypes in a sample of community-dwelling adults. Although age-related deficits in olfactory function are well documented, there are remarkably few studies that have investigated longitudinal olfactory change in aging. Specifically, we wanted to investigate if BDNF val66met influences olfactory decline and if the change varied as a function of the individuals’ baseline age.
Materials and Methods
Participants
All participants were derived from the Betula project, an ongoing population-based longitudinal project focusing on memory, health, and aging. The Betula data is sampled from the population of Umeå, a city located in the north of Sweden with 110,000 habitants (Nilsson et al., 1997 , 2004 ). The participants gave their informed consent and the Betula Project obtained approval from the regional board of the ethics committee in Umeå, Sweden. The data analyzed in this study were obtained from two stratified random samples (S1 and S3), where odor identification had been tested two times over 5 years. In Betula, extensive psychological testing and health assessment is conducted every 5 years and the odor identification test was added in the third wave of testing (T3). The first test occasion for the odor identification assessment took place between the years 1998 to 2000 and 5 years later the subjects were retested with the same procedure. At T3, 1323 participants had been BDNF genotyped, completed the MMSE, the odor identification and the vocabulary tests. Five years later, 980 participants in the study sample completed the same tests at the fourth wave of data collection (T4), resulting in a drop-out rate of 26%. Among the drop-outs, 42% had died and 9% had become demented. When taking death and dementia under consideration the drop-out rate was reduced to 17%.
Participants with a dementia diagnosis 10 years after first olfactory testing were removed from the present sample (n = 29). Also, participants diagnosed with Parkinson’s disease (n = 2) were removed. Longitudinal studies indicate that a decline in global cognitive ability reliably predicts an impending dementia (Amieva et al., 2005 ; Bäckman et al., 2005 ; Small and Backman, 2007 ). Impending dementia is furthermore associated with impaired olfaction (Olofsson et al., 2009 ). To minimize influences from preclinical dementia on the results, subjects with an MMSE score below 24 in the two test occasions (n = 26) or exhibiting a decrease of the MMSE score by 3 points or more during the two test occasions (n = 86) were excluded.
The final sample comprised 836 cognitively intact subjects (447 females and 389 males) who were tested on both test occasions. With regard to the genetic information, the frequencies across the different genotypes of the BDNF val66met polymorphism correspond to the frequencies earlier reported (Egan et al., 2003 ). The subjects were grouped according to genotype: val/val homozygous carriers, val/met heterozygous carriers, and met/met homozygous carriers. Due to the low population frequency of the met/met genotype (<5%), homozygous met carriers were collapsed with the met heterozygote carriers into one group referred to as the met carriers (Hariri et al., 2003 ).
Reports based on cross-sectional as well as longitudinal data indicate that olfactory function shows an accelerated age-related impairment from ages around 65–70 years compared to younger ages (Doty et al., 1984 ; Murphy et al., 2002 ; Olofsson et al., 2010 ). To investigate the combined effects of age and BDNF genotype, the participants were divided into two age groups: middle-aged subjects ranging from 45 to 65 years, and old subjects ranging from 70 to 90 years of age. See Table 1 for descriptive data across age and genotype groups.
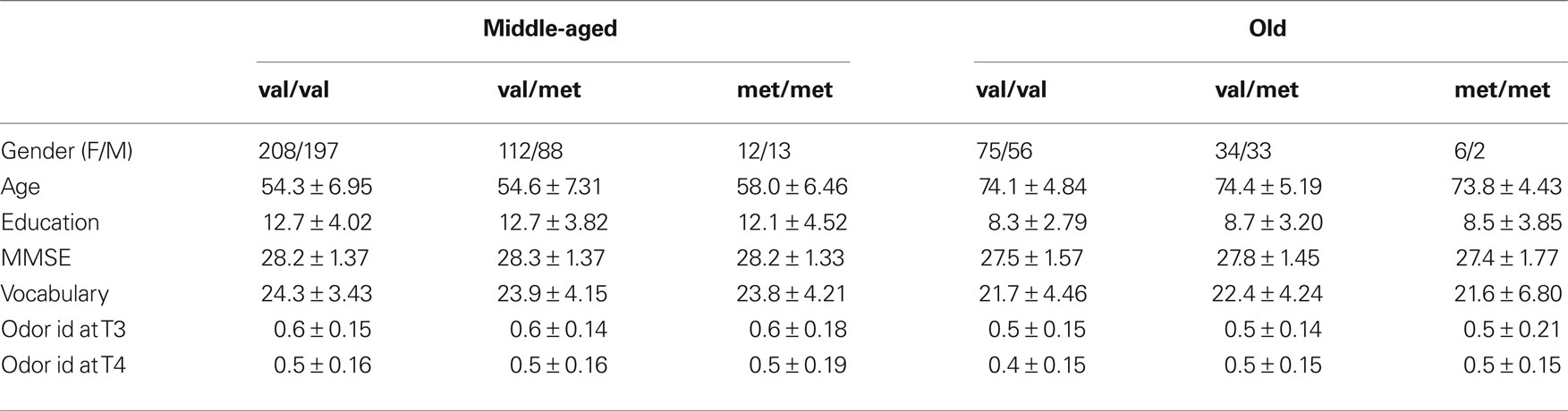
Table 1. Sample characteristics. Descriptive data across age and genotype groups (mean ± SD).
Procedure
Specific details regarding the test procedure have previously been reported (Nilsson et al., 1997 , 2004 ). In every test wave all participants were tested individually in two sessions, 1 week apart. The first test session was conducted by a nurse, and included blood sampling for genetic analyses, an extensive health examination, and some cognitive tests. Also, information regarding health-related and social factors was collected. In the second session, participants carried out a large battery of psychological tests, drawing on different cognitive domains. For the T3 and T4 this session also included the odor identification test. The test sessions lasted for approximately 2 h each, and the olfactory and psychological tests were administered by well-trained research assistants.
Olfactory and Cognitive Assessment
The Betula study includes a modified version of the Scandinavian Odor Identification Test (SOIT) (Nordin et al., 1998 ). The test consists of 13 odor stimuli specifically chosen to fit a Scandinavian population: almond (bitter), anise, apple, cinnamon, clove, juniper berry, lilac, lemon, orange, pine-needle, tar, vanilla, and violet. The selection of odors is empirically based on their identifiably, familiarity, intensity, and pleasantness. All odors are considered as predominantly olfactory stimulants and provide a good basis to generalize test performance to olfactory status. In order to minimize the cognitive demands, each stimulus is presented in conjunction with four force-choice response alternatives. The response alternatives were chosen based on a confusion matrix of identification responses. The SOIT has been demonstrated to constitute a valid and reliable test of odor identification ability (Nordin et al., 1998 ).
The liquid odorants were injected into a cotton pad and placed in an opaque, 80 ml glass jar. The stimuli were presented birhinally 1–2 cm under the participant’s nose for as long as required to accomplish the task. For each stimulus, the participant was provided with a written list of the four response alternatives and was instructed to choose the correct target odor name. To avoid ceiling effects, the distractor labels among the response alternatives were carefully chosen to be perceptually similar to the corresponding test odor. The stimulus order was randomized between subjects by randomly assigning one out of ten different stimulus orders to each subject. There was a 30-s inter-stimulus interval between stimuli to limit adaptation effects. The test took 10–15 min to administer.
As noted above, the MMSE was included as a screening test for global cognitive function and cognitive impairment. The MMSE is a brief 30-point questionnaire that assesses general cognitive function (Cockrell and Folstein, 1988 ).
A 30-item multiple-choice synonym test (SRB) was used to assess verbal knowledge that draws on semantic memory. Here, the participants were instructed to select a synonym for each target word out of five alternatives. A time limit was set to 7 min to complete the test. Since odor identification draws on semantic memory, the vocabulary test was included in the analyses to control for word knowledge that might influence odor identification performance (Larsson et al., 2004 ).
Health Assessment
The participants in the Betula study undergo an extensive health evaluation conducted by nurses in order to provide an overview the of health status of each participant. Dementia diagnoses are evaluated by a psychiatrist. An earlier study from the Betula project revealed that odor identification performance was negatively affected by diabetes and cardiovascular disorders (Olofsson et al., 2010 ). Consequently, these factors were controlled for statistically. Further, ear, nose, and throat disorders may affect olfactory functioning negatively (Deems et al., 1991 ) and were hence also controlled for statistically.
BDNF Genotyping
Genomic DNA was isolated from samples of whole blood and diluted to 2.5–5.0 ng/μl. Genotyping was performed using the Sequenom iPLEX® Gold assay and MassARRAY® MALDI-TOF mass spectrometry platform in accordance with the manufacturer’s instructions. Primers for PCR amplification (5′-ACGTTGGATGTTTTCTTCATTGGGCCGAAC-3′ and 5′-ACGTTGGATGCATCATTGGCTGACACTTTC-3′), unextendend primer (5′-CCAACAGCTCTTCTATCA-3′) and extension primers (5′-CCAACAGCTCTTCTATCAC-3′ and 5′-CCAACAGCTCTTCTATCAT-3′) were designed using the Sequenom MassARRAY® System Designer software.
Results
Intercorrelations Among Variables
In order to determine the relationships among the demographic, health, cognitive, genetic variables, and odor identification, zero-order Pearson correlations were calculated. The BDNF × age interaction term represents the residuals of the interaction effect after the main effects were partialled out. Thus, the interaction does not correlate with either age or BDNF status. The correlations among variables are displayed in Table 2 .
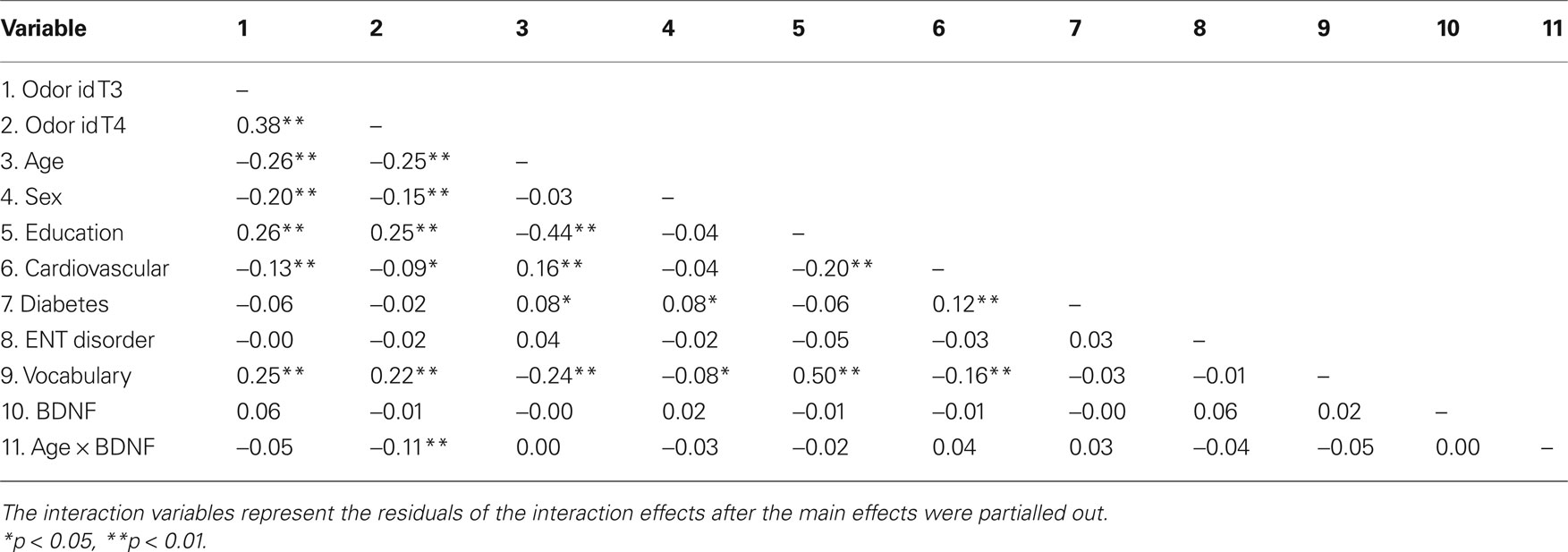
Table 2. Intercorrelations among variables.
The BDNF val66met did not correlate significantly with any variables. However, the interaction term comprising age and BDNF val66met correlated with odor identification performance at follow-up. Based on these observations, we carried out a hierarchical regression analysis to examine if the relation between age and BDNF val66met on odor identification at follow-up was related to a 5-year change in olfactory performance.
The allelic distribution of the BDNF val66met polymorphism fits the Hardy–Weinberg equilibrium for the middle-aged group (χ2 = 0.002, p = 0.96), and for the old group (χ2 = 0.05, p = 0.82). Also, the observed genotype frequencies did not differ between the two age groups (χ2 = 0.12, p = 0.94).
Regression Analyses
The data were analyzed with a hierarchical regression model comprising six blocks. Olfactory identification at follow-up was selected as the criterion measure. In the first block, the olfactory identification scores at baseline were added in order to control for baseline performance, and hence the variables of the following blocks would, if significant, predict olfactory change from T3 to T4. Then a block of demographic variables including age, sex, and years of formal education was added to the model. The third block consisted of health-related predictors, such as diabetes, cardiovascular disorders, and ear, nose, and throat disorders. The cognitive measure (i.e., vocabulary) was added in the fourth block. The fifth block contained information on the genetic variant of the BDNF val66met with two levels (i.e., the val/val homozygotes and the met carriers). Finally, in the sixth block, the age by BDNF val66met interaction term was included. The hierarchical regression model is illustrated in Table 3 .
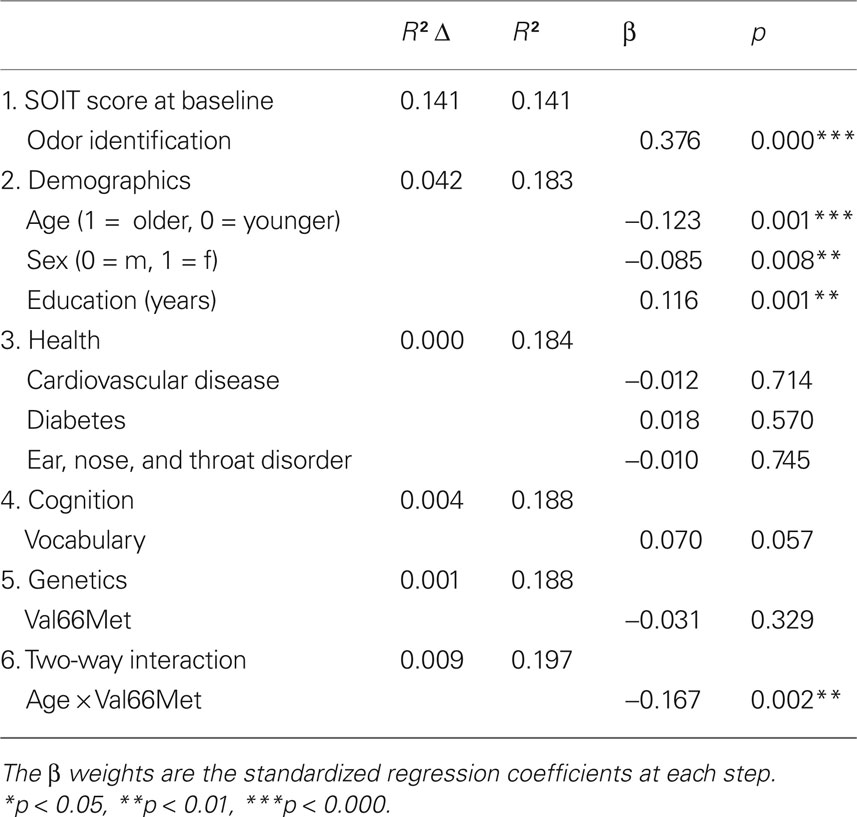
Table 3. Hierarchical regression analysis for predicting odor identification ability (SOIT) at 5-year follow-up (n = 836).
The predictor variables altogether accounted for 19.7% of the explanatory variance in odor identification performance at follow up. Odor identification proficiency at baseline (i.e., T3) was a significant predictor of odor identification at follow up (β = 0.376; p = 0.000) and explained 14.1% of the variance alone. In the second block, age, and education influenced olfactory decline, indicating that higher age and less years of formal education were related to faster olfactory decline. Sex affected the rate of the olfactory decline such that males declined more than females over the 5-year interval. None of the health predictors were related to olfactory decline. Semantic ability at baseline testing did not predict larger odor identification change. The genetic variable did not significantly contribute to odor identification change, suggesting that the BDNF val66met polymorphism alone has no general effect on odor identification decline in the age-range of 45–90 years at baseline. However, the interaction between age and val66met showed a highly significant contribution to olfactory decline (β = −0.167; p = .002). Importantly, the main variables in the age × val66met interaction were partialled out before the interaction term was added to the model.
The source of the interaction effect was investigated by splitting up the sample based on age cohort and BDNF val66met genotype. Four separate one-way ANOVAs were conducted with the standardized residuals from the regression model used above as a dependent variable. In the first and second ANOVAs, the sample was split based on age-cohorts and BDNF val66met was entered as between-group factor. The results showed that the effect of BDNF val66met on olfactory decline was non-significant in the middle-aged age cohort (p = 0.56), whereas BDNF val66met had a significant effect in the older age cohort [F(1,205) = 10.53, p = 0.001, η2 = 0.05]. Specifically, older val/val carriers displayed a significantly larger olfactory decline than the older met carriers. The third and fourth ANOVAs used age as a between-group factor. Separate analyses of the two genotype groups indicated a reliable age effect for the val homozygote carriers [F(1,534) = 18.44, p = 0.000, η2 = 0.03], with the older group exhibiting a larger olfactory decline. In contrast, no accelerated age-related decline in olfactory performance occurred in the older age groups among the met carriers (p = 0.62).
In order to explore the impact of age and BDNF val66met on the raw data scores for each test occasion, two separate between-group (Age × BDNF val66met status) ANOVAs were conducted, one on olfactory performance at baseline and the other on olfactory performance at follow-up. For the baseline measure, there was a significant effect of age [F(1,832) = 51.88, p = 0.000, η2 = 0.06], such that middle-aged participants outperformed the older adults. The main effect of BDNF status and the Age × BDNF interaction effect were not reliable (ps > 0.10) on olfactory baseline performance. For the follow-up olfactory test, there was a reliable age-effect, [F(1,832) = 36.25, p = 0.0001, η2 = 0.05] with poorer performance among the older than the middle-aged participants. Also, there was a main effect on BDNF status [F(1,832) = 4.24, p = 0.04, η2 = 0.01], such that val/val carriers outperformed met carriers on olfactory follow-up testing. The Age × BDNF interaction on olfactory follow-up performance was statistically significant [F(1,832) = 11.11, p = 0.001, η2 = 0.01] such that met carriers outperformed val/val carriers in the older age group whereas no difference was observed among the middle-aged adults.
Figure 1 displays raw data of the identification performance as a function of age and BDNF for baseline (A) and follow-up (B). Here, an impairment of the val/val carriers in the older age cohort is present in the follow-up testing. Figure 1 C illustrates the 5-year olfactory decline as a function of BDNF genotype across four age cohorts. In this graph the selective olfactory decline of the val/val carriers in the older age cohorts becomes apparent.
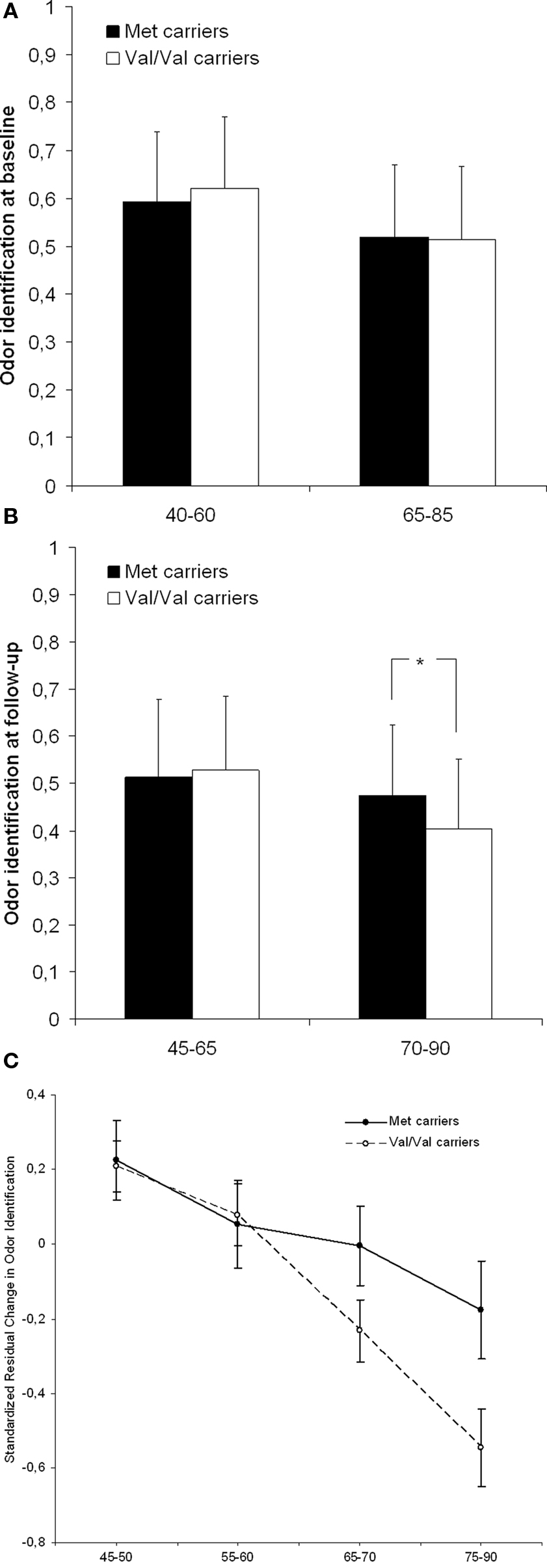
Figure 1. Odor identification performance (mean + SD) at baseline testing (A) and at follow-up testing (B) as a function of age and BDNF val66met status. (C) Displays the change in odor identification score (mean ± SE) from baseline to follow-up as a function of age and BDNF val66met status.
To explore whether the Age × BDNF val66met effect was driven by the met/met homozygotes (n = 33) in the collapsed met group the same regression analysis without the met/met carriers was conducted. The results showed that the Age × val66met interaction on olfactory decline remained significant (β = −0.176; p = 0.003) although met/met carriers were excluded from the sample. This observation suggests that met/met homozygosity is not of major influence for the observed interaction.
Also, to investigate whether the Age × BDNF interaction would be attenuated by dementia and cognitive decline, we performed an additional hierarchical regression analysis that included this group (n = 132) as a predictor variable. Dementia and cognitive decline had a significant effect on olfactory change (β = 0.150; p = 0.001). Interestingly, this factor did not affect the observed Age × BDNF effect on olfactory change (β = −0.214; p = 0.003), suggesting that dementia and cognitive decline are of minor importance for the Age × BDNF effect on olfactory change.
Discussion
The aim of this study was to explore the impact of the BDNF val66met polymorphism, a gene involved in neural proliferation and survival, on olfactory change using a large, population-based sample assessed longitudinally. The main findings suggest that the val66met affects the magnitude of the age-related decline in olfactory functioning. Specifically, the BDNF val66met did not affect the rate of decline in middle-aged subjects, whereas older val/val carriers showed a relatively higher age-related decline. The older carriers of the met allele displayed no accelerated decline over a 5-year-interval, whereas carriers of the val homozygote allele displayed an accelerated olfactory decline in the oldest age cohort compared to the middle-aged cohort.
Males exhibited a larger decrease in odor identification ability than females, corroborating previous work (Doty et al., 1984 ). Also, education influenced the rate of olfactory change, such that fewer years of education predicted a greater olfactory decrease. The included health-related factors, cardiovascular disease, diabetes, and ear, nose, and throat disorders proved unrelated to the rate of olfactory change.
It is important to note that the BDNF genotypes did not affect olfactory change overall but interacted with participants’ age. Specifically, the old (70–90 years of age) val/val carriers exhibited a steeper olfactory decline than the old met carriers. The current finding of an age by BDNF interaction on olfactory decline could be due to several reasons. It could be interpreted in the context of earlier findings on the met allele as a protective factor for cognitive decline (Harris et al., 2006 ; Erickson et al., 2008 ). However, it is worth noting that most of the previous literature investigating the effect of BDNF val66met on human memory functioning has shown that the met allele carriers were more impaired compared to val homozygotes carriers (Egan et al., 2003 ; Hariri et al., 2003 ; Miyajima et al., 2008 ; Nagel et al., 2008 ; Raz et al., 2009 ). Two studies focusing on older populations have reported a behavioral disadvantage of the val/val BDNF carriers, in congruence with the present results. Harris et al. (2006) studied the effect of BDNF val66met on cognitive function in an older sample (79-year olds) and found that the val/val genotype exerted negative effects on cognition in later life. Also, Erickson et al. (2008) established that the val/val genotype promoted faster rates of cognitive decline in old age. However, the effect of BDNF on olfactory performance is not necessarily related to the effects of BDNF on the cognitive profile. Also, it should be noted that the present data do not cover subjects in the earlier stages of the lifespan. Thus, it is possible that the influence of BDNF val66met on olfactory functioning in an earlier period is in the opposite direction.
Our findings suggest that the influence of the BDNF val66met genotypes on olfactory decline is only prevalent during the later stage of the lifespan. However, our data do not support the hypothesis that the steeper decline for val/val carriers is related to a preclinical AD state, at least within a 5-year follow-up interval. Association studies have suggested that BDNF is related to susceptibility of AD such that val/val carriers run an increased risk of developing AD, although other studies on AD and the BDNF val66met polymorphism have failed to demonstrate such an association (Combarros et al., 2004 ; Tsai et al., 2004 ). Olfactory dysfunction is related to AD and longitudinal studies have confirmed that preclinical AD cases exhibit impaired olfactory identification ability (Bacon et al., 1998 ). However, in the present study, dementia and preclinical dementia status did not affect the observed age × BDNF effect on olfactory decline. This finding suggests that the olfactory disadvantage among older val/val carriers is unrelated to AD, at least in a 5-year follow-up interval.
It is worth noting that the explanatory variance of the interaction between age and BDNF on odor change after accounting for demographic, health, and cognitive factors was approximately 1%, a finding that may be regarded as weak. However, the contribution of one SNP on a complex trait is expected to be small (Goldberg and Weinberger, 2004 ; Kovas and Plomin, 2006 ), and the size of the effect in the present study is in congruence with earlier studies reporting effects of BDNF on cognition (Egan et al., 2003 ; Erickson et al., 2008 ).
There are a number of potential mechanisms that may account for the evidenced influence of the BDNF on the rate of olfactory decline among older adults. Here we discuss two mechanisms that concern olfactory bulb neurogenesis and a genetically determined brain reserve.
Bath et al. (2008) reported that the BDNF val66met polymorphism modulates olfactory function through its involvement in olfactory bulb neurogenesis. Our results clearly highlight the different age-dependent roles played by the val and the met alleles for the integrity of olfactory function that may be driven by age-related changes in bulb neurogenesis (Zigova et al., 1998 ). However, it should be pointed out that it is yet unclear if supplement of BDNF can alter adult neurogenesis in this area (Galvao et al., 2008 ). Although several observations concerning neurogenesis in human adults report negative findings (Bhardwaj et al., 2006 ; Sanai et al., 2004 ), others have reported a preserved neurogenesis in the adult olfactory system (Bédard and Parent, 2004 ).
Speculatively, our findings may also be related to a genetically dependent brain reserve that regulates the number of neurons from birth (Katzman et al., 1988 ). BDNF could be one of the modifying genes of this reserve. Then, hypothetically, the val/val carriers may originally have started with less neurons and will consequently meet the threshold for olfactory deficits at an earlier stage. The lack of olfactory decline in the met carriers may then relate to a greater brain reserve among these individuals.
To summarize, the present findings provide new evidence that carriers of the val/val genotype show a marked aging-associated impairment in olfactory function, whereas the met allele of the BDNF val66met polymorphism may be protective against accelerated olfactory decline in the later stages of life.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by the Swedish Research Council Grant 421-2005-2113 to Maria Larsson and The Betula Study is funded by the Bank of Sweden Tercentenary Foundation (1988-0082:17), Swedish Council for Planning and Coordination of Research (D1988-0092, D1989-0115, D1990-0074, D1991-0258, D1992-0143, D1997-0756, D1997-1841, D1999-0739, B1999-474), Swedish Council for Research in the Humanities and Social Sciences (F377/1988-2000), the Swedish Council for Social Research (1988-1990: 88-0082, and 311/1991-2000), and the Swedish Research Council (315-2004-6977).
References
Amieva, H., Jacqmin-Gadda, H., Orgogozo, J.-M., Le Carret, N., Helmer, C., Letenneur, L., Barberger-Gateau, P., Fabrigoule, C., and Dartigues, J. F. (2005). The 9 year cognitive decline before dementia of the Alzheimer type: a prospective population-based study. Brain 128, 1093–1101.
Bäckman, L., Jones, S., Berger, A., Laukka, E. J., and Small, B. J. (2005). Cognitive impairment in preclinical Alzheimer’s disease: a meta-analysis. Neuropsychology 19, 520–531.
Bacon, A. W., Bondi, M. W., Salmon, D. P., and Murphy, C. (1998). Very early changes in olfactory functioning due to Alzheimer’s disease and the role of apolipoprotein E in olfaction. Ann. N. Y. Acad. Sci. 855, 723–731. (Olfaction and Taste XII: An International Symposium)
Bath, K. G., Mandairon, N., Jing, D., Rajagopal, R., Kapoor, R., Chen, Z., Khan., T., Proenca, C. C., Kraemer,.R., Cleland, T. A., Hempstead, B. L., Chao, M. V., and Lee, F. S. (2008). Variant brain-derived neurotrophic factor (Val66Met) alters adult olfactory bulb neurogenesis and spontaneous olfactory discrimination. J. Neurosci. 28, 2383–2393.
Bédard, A., and Parent, A. (2004). Evidence of newly generated neurons in the human olfactory bulb. Dev. Brain Res. 151, 159–168.
Bhardwaj, R. D., Curtis, M. A., Spalding, K. L., Buchholz, B. A., Fink, D., Björk-Eriksson, T., Nordborg, C., Gage, F. H., Druid, H., Eriksson, P. S, and Frisén, J. (2006). Neocortical neurogenesis in humans is restricted to development. Proc. Natl. Acad. Sci. U.S.A. 103, 12564–12568.
Brämerson, A., Johansson, L., Ek, L., Nordin, S., and Bende, M. (2004). Prevalence of olfactory dysfunction: the Skövde population-based study. Laryngoscope 114, 733–737.
Bueller, J., Aftab, M., Sen, S., Gomez-Hassan, D., Burmeister, M., and Zubieta, J. (2006). BDNF Val66Met allele is associated with reduced hippocampal volume in healthy subjects, Biol. Psychiatry 59, 812–815.
Cockrell, J. R., and Folstein, M. F. (1988). Mini-Mental State Examination (MMSE). Psychopharmacol. Bull. 24, 689–692.
Combarros, O., Infante, J., Llorca, J., and Berciano, J. (2004). Polymorphism at codon 66 of the brain-derived neurotrophic factor gene is not associated with sporadic Alzheimer’s disease. Dement. Geriatr. Cogn. Disord. 18, 55–58.
Deems, D. A., Doty, R. L., Settle, G., Mooregillon, V., Shaman, P., Mester, A. F., Kimmelman, C. P., Brightman, V. J., and Snow, J. B. (1991). Smell and taste disorders, a study of 750 patients from the University-of-Pennsylvania smell and taste center. Arch. Otolaryngol. 117, 519–528.
del Tredici, K., and Braak, H. (2008). Neurofibrillary changes of the Alzheimer type in very elderly individuals: neither inevitable nor benign – Commentary on “no disease in the brain of a 115-year-old woman”. Neurobiol. Aging 29, 1133–1136.
Doty, R., Shaman, P., Applebaum, S., Giberson, R., and Siksorski, L. (1984). Smell identification ability: changes with age. Science 226, 1441–1443.
Doty, R. L., Shaman, P., and Dann, M. (1984). Development of the University of Pennsylvania smell identification test: A standardized microencapsulated test of olfactory function. Physiol. Behav. 32, 489–502.
Egan, M. F., Kojima, M., Callicott, J. H., Goldberg, T. E., Kolachana, B. S., Bertolino, A., Zaitsev, E., Gold, B., Goldman, D., Dean, M., Lu B., and Weinberger, D. R. (2003). The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell 112, 257–269.
Erickson, K. I., Kim, J. S., Suever, B. L., Voss, M. W., Francis, B. M., and Kramer, A. F. (2008). Genetic contributions to age-related decline in executive function: a 10-year longitudinal study of COMT and BDNF polymorphisms. Front. Hum. Neurosci. 2, 11. doi:10.3389/neuro.09.011.2008.
Galvao, R., Garcia-Verdugo, J., and Alvarez-Buylla, A. (2008). Brain-derived neurotrophic Factor signaling does not stimulate subventricular zone neurogenesis in adult mice and rats. J. Neurosci. 28, 13368–13383.
Goldberg, T. E., and Weinberger, D. R. (2004). Genes and the parsing of cognitive processes. Trends Cogn. Sci. (Regul. Ed.) 8, 325–335.
Hariri, A. R., Goldberg, T. E., Mattay, V. S., Kolachana, B. S., Callicott, J. H., Egan, M. F., and Weinberger, D. R. (2003). Brain-derived neurotrophic factor val66met polymorphism affects human memory-related hippocampal activity and predicts memory performance. J. Neurosci. 23, 6690–6694.
Harris, S. E., Fox, H., Wright, A. F., Hayward, C., Starr, J. M., Whalley, L. J., and Deary, I. J. (2006). The brain-derived neurotrophic factor Val66Met polymorphism is associated with age-related change in reasoning skills. Mol. Psychiatry 11, 505–513.
Katoh-Semba, R., Semba, R., Takeuchi, I. K., and Kato, K. (1998). Age-related changes in levels of brain-derived neurotrophic factor in selected brain regions of rats, normal mice and senescence-accelerated mice: a comparison to those of nerve growth factor and neurotrophin-3. Neurosci. Res. 31, 227–234.
Katzman, R., Terry, R., DeTeresa, R., Brown, T., Davies, P., Fuld, P., Renbing, X., and Peck, A. (1988). Clinical, pathological, and neurochemical changes in dementia: a subgroup with preserved mental status and numerous neocortical plaques. Ann. Neurol. 23, 138–144.
Keller, A., and Vosshall, L. B. (2008). Better smelling through genetics: mammalian odor perception. Curr. Opin. Neurobiol. 18, 364–369.
Keller, A., Zhuang, H. Y., Chi, Q. Y., Vosshall, L. B., and Matsunami, H. (2007). Genetic variation in a human odorant receptor alters odour perception. Nature 449, 468–472.
Kovas, Y., and Plomin, R. (2006). Generalist genes: implications for the cognitive sciences. Trends Cogn. Sci. (Regul. Ed.) 10, 198–203.
Larsson, M., Nilsson, L., Olofsson, J. K., and Nordin, S. (2004). Demographic and cognitive predictors of cued odor identification: evidence from a population-based study. Chem. Sens. 29, 547–554.
Mesholam, R. I., Moberg, P. J., Mahr, R. N., and Doty, R. L. (1998). Olfaction in neurodegenerative disease: a meta-analysis of olfactory functioning in Alzheimer’s and Parkinson’s Diseases. Arch. Neurol. 55, 84–90.
Miyajima, F., Ollier, W., Mayes, A., Jackson, A., Thacker, N., Rabbitt, P., Pendleton, N., Horan, M., and Payton, A. (2008). Brain-derived neurotrophic factor polymorphism Val66Met influences cognitive abilities in the elderly. Genes Brain Behav. 7, 411–417.
Murphy, C., Schubert, C. R., Cruickshanks, K. J., Klein, B. E. K., Klein, R., and Nondahl, D. M. (2002). Prevalence of olfactory impairment in older adults. JAMA 288, 2307–2312.
Nagel, I. E., Chicherio, C., Li, S., Oertzen, T. V., Sander, T., Villringer, A., Heekeren, H. R., Bäckma, L., and Lindenberger, U. (2008). Human aging magnifies genetic effects on executive functioning and working memory. Front. Hum. Neurosci. 2, 1. doi:10.3389/neuro.09.001.2008.
Nilsson, L., Adolfsson, R., Backman, L., de Frias, C., Molander, B., and Nyberg, L. (2004). Betula: a prospective cohort study on memory, health and aging. Aging Neuropsychol. C. 11, 134–148.
Nilsson, L., Backman, L., Erngrund, K., Nyberg, L., Adolfsson, R., Bucht, G., Karlsson, S., Widing, M., and Winblad, B. (1997). The Betula prospective cohort study: memory, health and aging. Aging Neuropsychol. C. 4, 1–32.
Nordin, S., Bramerson, A., Liden, E., and Bende, M. (1998). The Scandinavian Odor-Identification Test: development, reliability, validity and normative data. Acta Oto-Laryngol. 118, 226–234.
Olofsson, J. K., Nordin, S., Wiens, S., Hedner, M., Nilsson, L., and Larsson, M. (2010). Odor identification impairment in carriers of ApoE-[var epsilon]4 is independent of clinical dementia. Neurobiol. Aging 31, 567–577.
Olofsson, J. K, Rönnlund. M., Nordin, S., Nyberg, L., Nilsson L.-G., and Larsson, M. (2009). Odor identification deficit as a predictor of five-year global cognitive change: interactive effects with age and ApoEε4. Behav. Genet. 39, 496–503.
Pezawas, L., Verchinsky, B. A., Mattay, V. S., Callicott, J. H., Kolachana, B. S., Straub, R. E., Egan, M. F., Meyer-Lindenberg, A., and Weinberger, D. R. (2004). The brain-derived neurotrophic factor val66met polymorphismand variation in human cortical morphology. J. Neurosci. 24, 10099–10102.
Phillips, H. S., Hains, J. M., Armanini, M., Laramee, G. R., Johnson, S. A., and Winslow, J. W. (1991). BDNF mRNA is decreased in the hippocampus of individuals with Alzheimer’s disease. Neuron 7, 695–702.
Raz, N., Rodrigue, K. M., Kennedy, K. M., and Land, S. (2009). Genetic and Vascular Modifiers of age-sensitive cognitive skills: effects of COMT, BDNF, ApoE and hypertension. Neuropsychology 23, 105–116.
Sanai, N., Tramontin, A. D., Quinones-Hinojosa, A., Barbaro, N. M., Gupta, N., Kunwar, S., Lawton, M. T., McDermott, M. W., Parsa, A. T., Manuel-García Verdugo, J., Berger, M. S., and Alvarez-Buylla, A. (2004). Unique astrocyte ribbon in adult human brain contains neural stem cells but lacks chain migration. Nature 427, 740–744.
Siegel, G., and Chauhan, N. (2000). Neurotrophic factors in Alzheimer’s and Parkinson’s disease brain. Brain Res. Rev. 33, 199–227.
Simpson, P., Miller, I., Moon, C., Hanlon, A., Liebl, D., and Ronnett, G. (2002). Atrial natriuretic peptide type C induces a cell-cycle switch from proliferation to differentiation in brain-derived neurotrophic factor- or nerve growth factor-primed olfactory receptor neurons. J. Neurosci. 22, 5536–5551.
Small, B., and Backman, L. (2007). Longitudinal trajectories of cognitive change in preclinical Alzheimer’s disease: a growth mixture modeling analysis. Cortex 43, 826–834.
Stevens, J. C., and Cain, W. S. (1987). Old-age deficits in the sense of smell as gauged by thresholds, magnitude matching, and odor identification. Psychol. Aging 2, 36–42.
Tabert, M. H., Liu, X., Doty, R. L., Serby, M., Zamora, D., Pelton, G. H., Marder, K., Albers, M. W., Stern, Y., and Devanand, D. P. (2005). A 10-item smell identification scale related to risk for Alzheimer’s disease. Ann. Neurol. 58, 155–160.
Tsai, S., Hong, C., Liu, H., Liu, T., Hsu, L., and Lin, C. (2004). Association analysis of brain-derived neurotrophic factor Val66Met polymorphisms with Alzheimer’s disease and age of onset. Neuropsychobiology 49, 10–12.
Ventriglia, M., Chiavetto, L., Benussi, L., Binetti, G., Zanetti, O., Riva, M., and Gennarelli, M. (2002). Association between the BDNF 196 A/G polymorphism and sporadic Alzheimer’s disease. Mol. Psychiatry 7, 136–137.
Whissell-Buechy, D., and Amoore, J. E. (1973). Letter: Odour-blindness to musk: simple recessive inheritance. Nature 245, 157–158.
Wilson, R. S., Arnold, S. E., Schneider, J. A., Tang, Y., and Bennett, D. A. (2007). The relationship between cerebral Alzheimer’s disease pathology and odour identification in old age. J. Neurol. Neurosurg. Psychiatry 78, 30–35.
Keywords: brain-derived neurotrophic factor, val66met, olfaction, odor identification, aging
Citation: Hedner M, Nilsson L-G, Olofsson JK, Bergman O, Eriksson E, Nyberg L, and Larsson M (2010) Age-related olfactory decline is associated with the BDNF val66met polymorphism: evidence from a population-based study. Front. Ag. Neurosci. 2:24. doi: 10.3389/fnagi.2010.00024
Received: 26 February 2010;
Paper pending published: 15 March 2010;
Accepted: 18 May 2010;
Published online: 07 June 2010
Edited by:
Thomas C. Foster, University of Florida, USAReviewed by:
Naftali Raz, Wayne State University, USARichard Doty, University of Pennsylvania, USA
Sarah Harris, University of Edinburgh, UK
Copyright: © 2010 Hedner, Nilsson, Olofsson, Bergman, Eriksson, Nyberg and Larsson. This is an open-access article subject to an exclusive license agreement between the authors and the Frontiers Research Foundation, which permits unrestricted use, distribution, and reproduction in any medium, provided the original authors and source are credited.
*Correspondence: Margareta Hedner, Department of Psychology, Stockholm University, Frescati Hagväg 14, SE - 106 91 Stockholm, Sweden. e-mail: margareta.hedner@psychology.su.se