 Chuang Li
Chuang Li Caixia Liu
Caixia Liu Na Li
Na Li- 1Department of Obstetrics and Gynecology, Shengjing Hospital of China Medical University, Shenyang, China
- 2Key Laboratory of Maternal-Fetal Medicine of Liaoning Province, Shenyang, China
Growing evidence indicates that gut microbiota could be closely associated with a variety of adverse pregnancy outcomes (APOs), but a causal link between gut microbiome and APOs has yet to be established. Therefore, in this study, we comprehensively investigated the relationship between gut microbiota and APOs to identify specific causal bacteria that may be associated with the development and occurrence of APOs by conducting a two-sample Mendelian randomization (MR) analysis. The microbiome genome-wide association study (GWAS) from the MiBioGen consortium was used as exposure data, and the GWAS for six common APOs was used as outcome data. Single-nucleotide polymorphisms (SNPs) that significantly correlated to exposure, data obtained from published GWAS, were selected as instrumental variables (IVs). We used the inverse variance-weighted (IVW) test as the main MR analysis to estimate the causal relationship. The Mendelian randomization pleiotropy residual sum and outlier (MR-PRESSO) and MR-Egger regression were used to confirm the presence of horizontal pleiotropy and to exclude outlier SNPs. We performed Cochran's Q test to assess the heterogeneity among SNPs associated with each bacterium. The leave-one-out sensitivity analysis was used to evaluate whether the overall estimates were affected by a single SNP. Our analysis shows a causal association between specific gut microbiota and APOs. Our findings offer novel insights into the gut microbiota-mediated development mechanism of APOs.
Introduction
Adverse pregnancy outcomes (APOs) include all pathological pregnancy and childbirth complications. Many public health and medical interventions have been implemented to reduce APOs, yet the incidence of APOs remains high. APOs are a leading cause of maternal, fetal, and neonatal deaths; severe maternal morbidity; and maternal intensive care unit admissions (Rana et al., 2019). APOs are widely recognized as major public health problems due to short- and long-term adverse impacts on mothers and infants.
The gut microbiota is a massive complex community of microbial species living in the digestive tract that not only participates in the digestion of food and the absorption of nutrients but also is involved in physiological regulation of the host through the production of hormonally active substances (Luo et al., 2022). Increasing evidence indicates that microbiome dysbiosis might play an important role in the pathogenesis of APOs (Qi et al., 2021). Zheng et al. (2020) found that dynamics in gut microbiota during the first half of pregnancy differed significantly between gestational diabetes mellitus (GDM) and normoglycemic pregnant women. Another case–control study showed that pregnant women with pre-eclampsia (PE) had a high abundance of Fusobacterium and Veillonella and a relatively low abundance of Faecalibacterium and Akkermansia compared with normotensive pregnant women (Chen C. et al., 2020; Chen X. et al., 2020). Yin et al. (2021) reported that women with preterm birth (PTB) showed a distinct gut microbiome dysbiosis compared with those who delivered at term. Opportunistic pathogens, particularly Porphyromonas, Streptococcus, Fusobacterium, and Veillonella, were enriched, while Coprococcus and Gemmiger were markedly depleted in the preterm group (Yin et al., 2021). However, whether these associations are causal has not been established because of potential biases including residual confounding factors and reverse causality.
Mendelian randomization (MR) is an approach integrating summary data of the genome-wide association study (GWAS) and resembles a randomized controlled trial by leveraging allelic randomization during meiosis and subsequent irreversible exposure to genotype at conception (Lawlor et al., 2008; Ellervik et al., 2019; Ference et al., 2019). Using genetic variants as instrumental variables (IVs), the MR design is typically less likely to be affected by residual confounding factors and reverse causation than conventional observational analysis, thereby strengthening the causal relationship between exposure and outcomes (Chen C. et al., 2020; Chen J. et al., 2020; Yuan and Larsson, 2022).
Exploring causal associations between gut microbiota and APOs not only deepens our understanding of pathogenesis derived from enteric bacteria but also facilitates the development of personalized treatments for pregnancy complications based on dietary or microbiome interventions (Di Simone et al., 2020). Elucidating the causal inference on the gut microbiome and its association with APOs is warranted. Thus, in this study, we conducted a two-sample MR analysis to evaluate the causal association between gut microbiota and six common APOs, namely, postpartum hemorrhage (PPH), abruptio placenta (AP), spontaneous abortion (SAB), premature rupture of membrane (PROM), gestational diabetes mellitus (GDM), and pre-eclampsia (PE) or eclampsia (Figure 1). To our knowledge, this is the first study assessing the causal role of gut microbiome in the development of pregnancy complications.
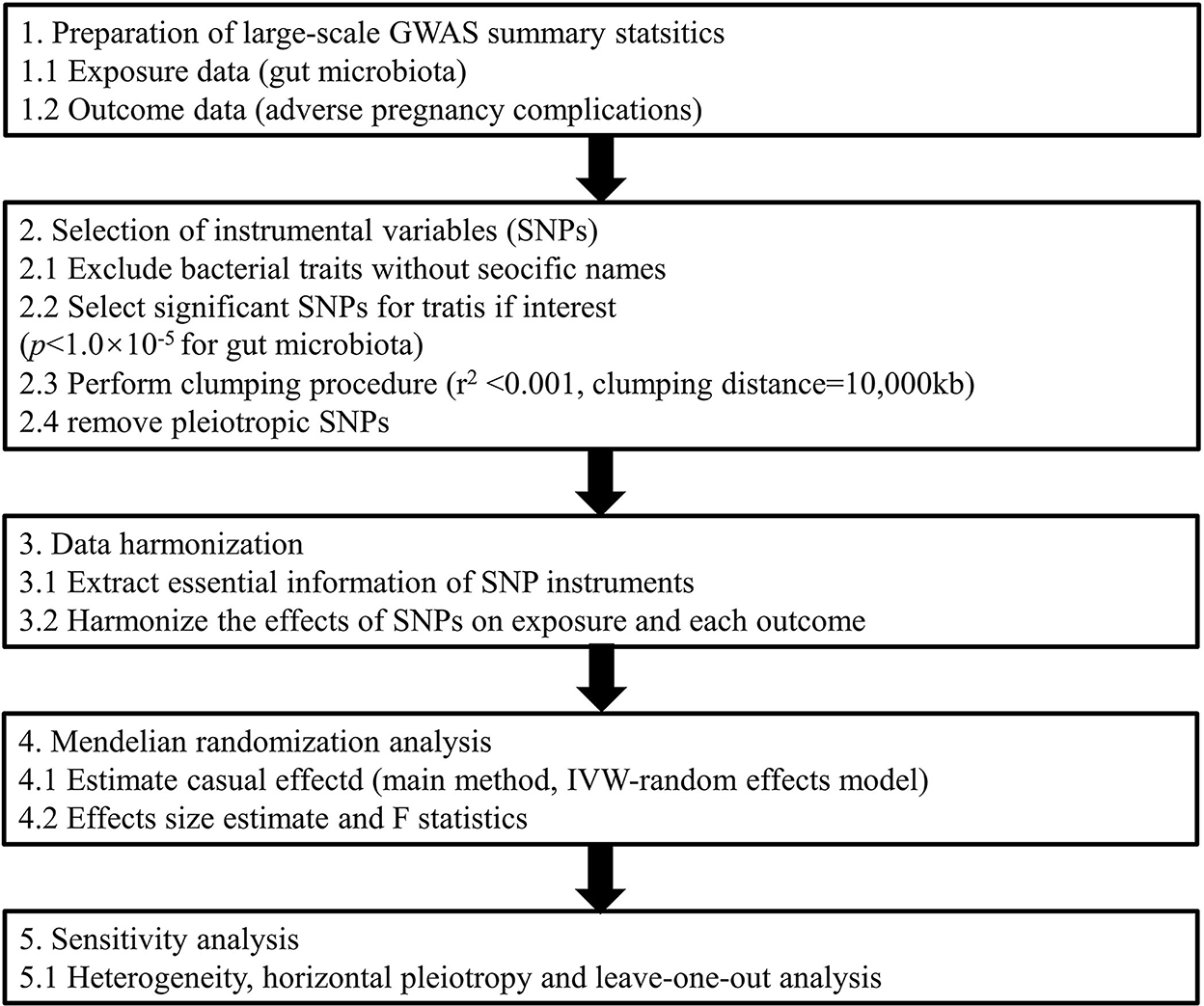
Figure 1. Study design of the present MR study on the associations of gut microbiota and adverse pregnancy complications. SNP, single-nucleotide polymorphism; IVW, inverse variance-weighted; MR, Mendelian randomization.
Materials and methods
Data sources
We obtained the GWAS dataset of human gut microbiome from the MiBioGen consortium (Kurilshikov et al., 2021). The MiBioGen study is the largest, multi-ethnic, and genome-wide analysis to investigate host –genetics– microbiome associations (Wang et al., 2018). This study coordinated 16S rRNA gene sequencing profiles and genotyping data of 18,340 participants from 24 population-based cohorts comprising 211 bacterial taxa (Kurilshikov et al., 2021), among which 15 microbial taxa without specific species name (unknown family or genus) were excluded, resulting in a total of 196 bacterial taxa being included in the present study.
GWAS summary data on PPH (3,670 cases and 98,626 controls), AP (294 cases and 104,247 controls), SAB (9,113 cases and 89,340 controls), PROM (3,011 cases and 104,247 controls), GDM (5,687 cases and 117,892 controls), and PE or eclampsia (3,903 cases and 114,735 controls) were obtained from results of the GWAS on the FinnGen consortium R5. Detailed data on the involved cohorts, genotypes, endpoint definition, and association test in the FinnGen consortium are available on the FinnGen webpage.
Instrumental variable selection
The accuracy of the conclusion on the causal effect of enteric microbiota on APOs was ensured by the following quality control steps used for the section of genetic predictors associated with microbiome features. We selected instrumental variables (IVs) with the locus-wide significance level (p < 1 × 10−5) to obtain a more comprehensive result (Sanna et al., 2019). The impact of linkage disequilibrium (LD) among included genetic variants on the results was avoided by excluding the SNPs using the PLINK clumping method (r2 > 0.001 and clump window <10,000 kb). We then extracted the corresponding data of the selected SNPs from the GWAS outcome data. When SNPs related to exposure were absent in the GWAS outcome data, the proxy SNPs with high LD (r2 > 0.80) would be selected. We removed palindromic SNPs to ensure the effects of SNPs on exposure correspond to the same allele as the effects of SNPs on the outcome during the harmonizing process.
Mendelian randomization analysis
MR analysis is a method used to estimate the magnitude of a causal effect of a phenotype on an outcome by using genetic variants such as IVs (Burgess et al., 2011; Davey Smith and Hemani, 2014). There are three important assumptions of MR analysis, which are depicted in Figure 2. The first assumption is that variants should be associated with exposure. The second assumption is that variants should not be associated with any confounders. The third assumption is that the association of genetic variants with outcomes should only be through exposure (Burgess and Thompson, 2015; Yuan S. et al., 2021).
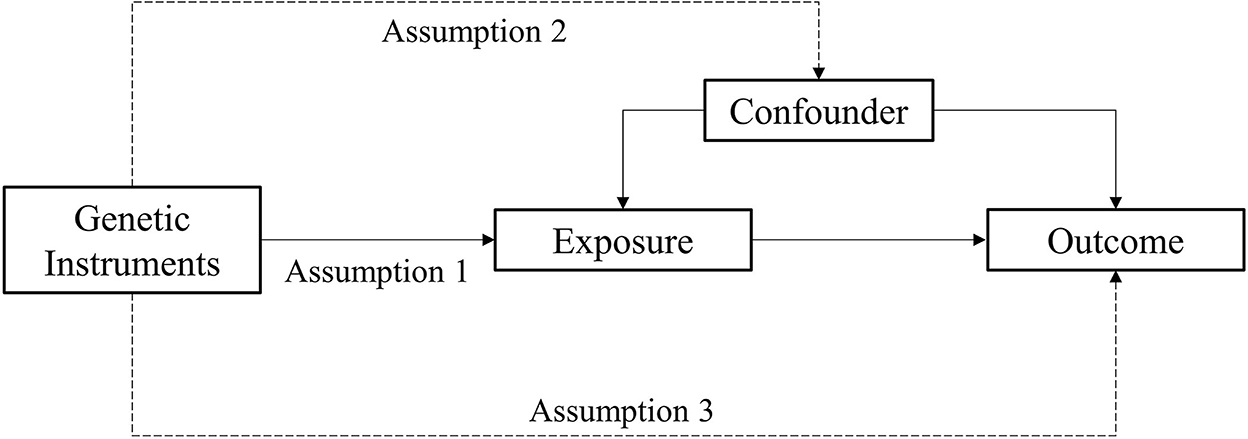
Figure 2. Assumption of the Mendelian randomization study.
The two-sample IVW method was used as the main MR analysis to estimate the causal relationship of gut microbiota with APOs (Burgess et al., 2013). We performed sensitivity analysis in order to assess the significance of our results. We used the MR-Egger regression and MR-PRESSO to confirm the presence of horizontal pleiotropy (Bowden et al., 2015; Verbanck et al., 2018). Using MR-Egger regression, we can assess whether genetic instruments have pleiotropic effects on the outcome (Bowden et al., 2015). Compared with MR-PRESSO, the MR-Egger method has less precision and statistical power. The MR-PRESSO can detect horizonal pleiotropy and eliminate the effects of pleiotropy by conducting outlier removal (Verbanck et al., 2018). If horizontal pleiotropy was detected among the selected SNPs, the analyses were repeated after removing these pleiotropic SNPs. We performed Cochran's Q test to evaluate the heterogeneity among SNPs associated with each microbial taxon (Egger et al., 1997). The leave-one-out sensitivity analysis was used to assess the effects of a single SNP on the overall estimates. The strength of the selected IVs was assessed using F statistics, which determine if estimates of the causal associations are affected by weak instrument bias. We excluded weak IVs with F statistics <10 (Pierce et al., 2011). All MR analyses were performed in R (version 4.1.2) using R package TwoSampleMR (Hemani et al., 2018) and MRPRESSO (Verbanck et al., 2018).
Results
Selection of instrumental variables
After ensuring quality control, we selected 61 SNPs related to five bacterial taxa for PPH, 109 SNPs related to 10 bacterial taxa for AP, 33 SNPs related to four bacterial taxa for SAB, 81 SNPs related to seven bacterial taxa for PROM, 155 SNPs related to 14 bacterial taxa for GDM, 132 SNPs related to 11 bacterial taxa for PE or eclampsia, and 44 SNPs related to four bacterial taxa for PTB (Supplementary Table S1). The F statistics for IVs significantly associated with gut microbiota were all larger than 10, indicating that the estimates were less likely to suffer weak instrument bias.
Causal effects of gut microbiota on adverse pregnancy outcomes
PPH
The estimates of the IVW test indicated that the genetically predicted relative abundance of five genera, namely, Deltaproteobacteria, Butyricimonas, Marvinbryantia, Oscillibacter, and Peptococcus, was associated with an increased or reduced risk of PPH (Table 1; Figure 3) (Chen C. et al., 2020). Specifically, a higher genetically predicted abundance of Deltaproteobacteria was associated with a higher risk of PPH (OR: 1.26, 95% CI: 1.00–1.59, p = 0.047). A higher genetically predicted abundance of Butyricimonas was associated with a reduced risk of PPH (OR: 0.79, 95% CI: 0.63–0.98, p = 0.036). The genetically predicted abundance of Marvinbryantia was also associated with a reduced risk of PPH (OR: 0.76, 95% CI: 0.59–0.98, p = 0.033). The genetically predicted abundance of Oscillibacter was negatively related to the risk of PPH (OR: 0.80, 95% CI: 0.64–0.99, p = 0.042). The higher genetically predicted abundance of Peptococcus was also linked to a reduced risk of PPH (OR: 0.84, 95% CI: 0.72–0.97, p = 0.024).
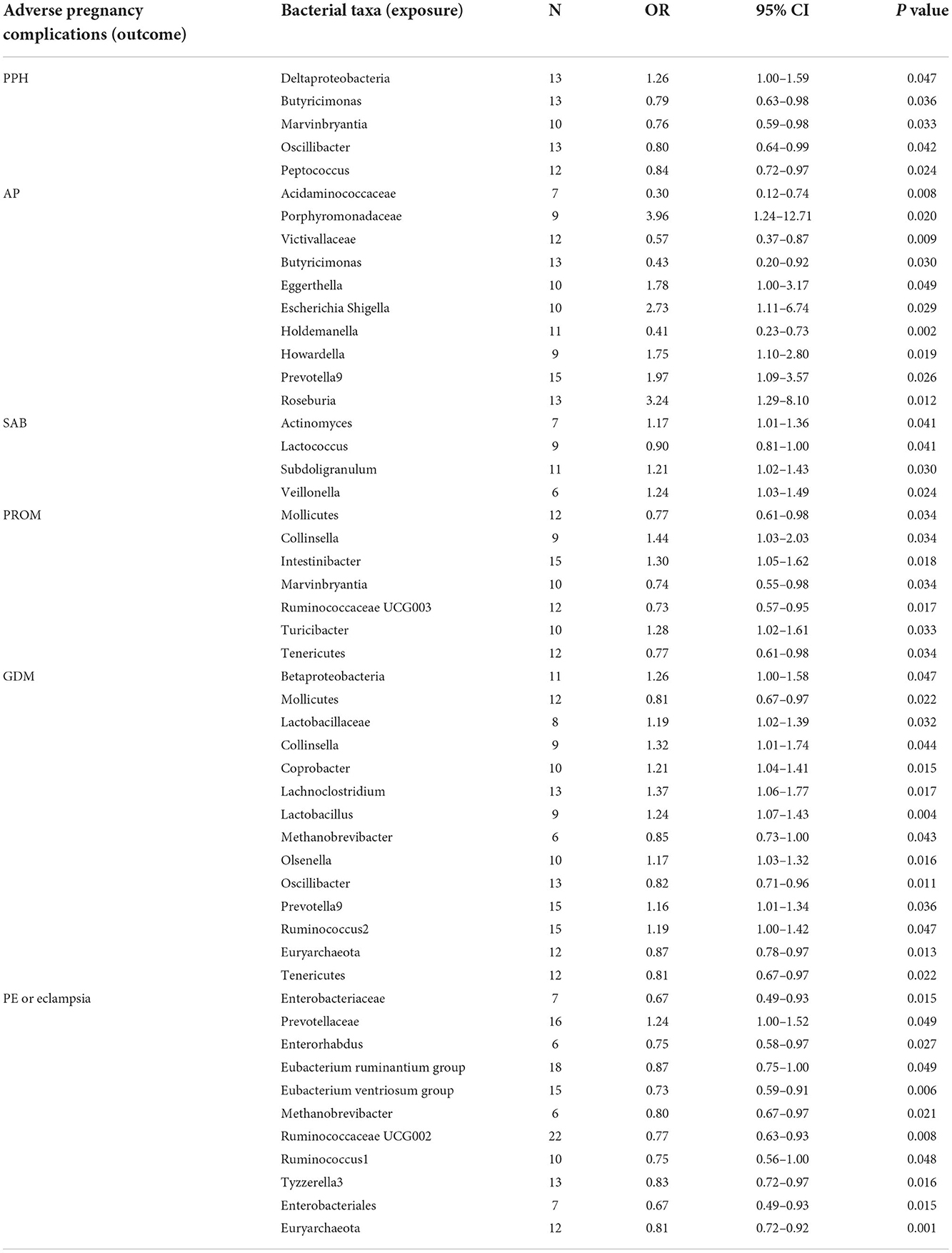
Table 1. Causal associations between gut microbiota and the risk of adverse pregnancy complications by using the IVW method.
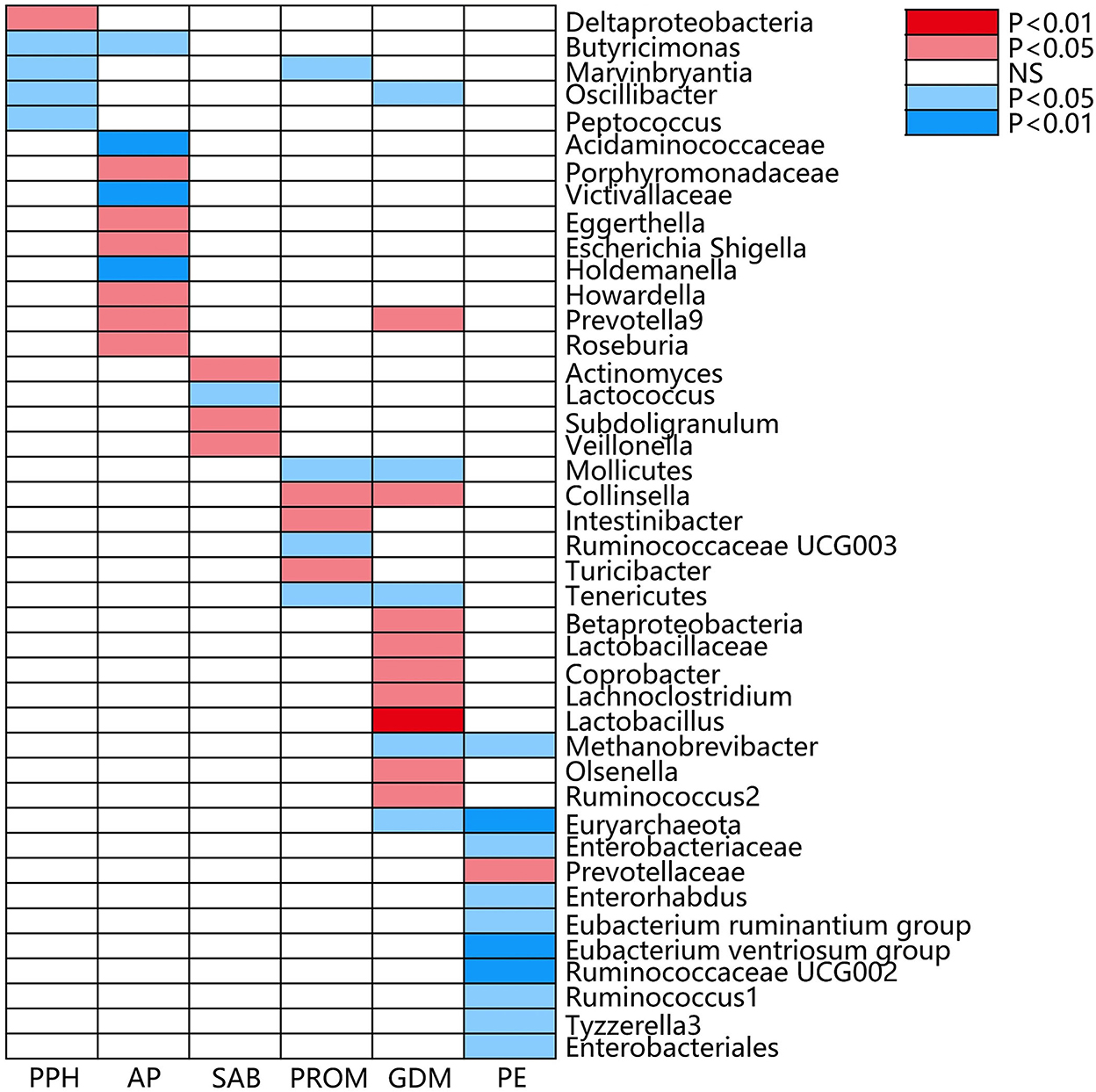
Figure 3. Causal associations of gut microbial genera on adverse pregnancy outcomes (APOs) identified at the nominal significance from the IVW method (P < 0.05/0.01). Red represents the risk bacterial traits for APOs, blue represents the protective bacterial traits for APOs, and white represents no causal bacterial traits for APOs. PPH, postpartum hemorrhage; AP, abruptio placentae; SAB, spontaneous abortion; PROM, premature rupture of membrane; GDM, gestational diabetes mellitus; PE, pre-eclampsia; NS, no significant association.
AP
The estimates of the IVW test suggested that the genetically predicted relative abundance of four genera, namely, Acidaminococcaceae (OR: 0.30, 95% CI: 0.12–0.74, p = 0.008), Victivallaceae (OR: 0.57, 95% CI: 0.37–0.87, p = 0.009), Butyricimonas (OR: 0.43, 95% CI: 0.20–0.92, p = 0.030), and Holdemanella (OR: 0.41, 95% CI: 0.23–0.73, p = 0.002), was negatively associated with the risk of AP. The genetically predicted relative abundance of a different five genera, namely, Porphyromonadaceae (OR: 3.96, 95% CI: 1.24–12.71, p = 0.020), Eggerthella (OR: 1.78, 95% CI: 1.00–3.17, p = 0.049), Escherichia Shigella (OR: 2.73, 95% CI: 1.11–6.74, p = 0.029), Howardella (OR: 1.75, 95% CI: 1.10–2.80, p = 0.019), Prevotella9 (OR: 1.97, 95% CI: 1.09–3.57, p = 0.026), and Roseburia (OR: 3.24, 95% CI: 1.29–8.10, p = 0.012), was positively associated with the risk of AP.
SAB
A higher genetically predicted abundance of Actinomyces (OR: 1.17, 95% CI: 1.01–1.36, p = 0.041), Subdoligranulum (OR: 1.21, 95% CI: 1.02–1.43, p = 0.030), and Veillonella (OR: 1.24, 95% CI: 1.03–1.49, p = 0.024) was significantly associated with the risk of SAB. There was a tendency toward a protective effect of increasing genetically predicted abundance of Lactococcus on the risk of SAB (OR: 0.90, 95% CI: 0.81–1.00, p = 0.041).
PROM
The genetically predicted relative abundance of three genera, namely, Collinsella (OR: 1.44, 95% CI: 1.03–2.03, p = 0.034), Intestinibacter (OR: 1.30, 95% CI: 1.05–1.62, p = 0.018), and Turicibacter (OR: 1.28, 95% CI: 1.02–1.61, p = 0.033), was associated with an increased risk of PROM. By contrast, there was a tendency toward a protective effect of increasing genetically predicted abundance of Mollicutes (OR: 0.77, 95% CI: 0.61–0.98, p = 0.034), Marvinbryantia (OR: 0.74, 95% CI: 0.55–0.98, p = 0.034), Ruminococcaceae UCG003 (OR: 0.73, 95% CI: 0.57–0.95, p = 0.017), and Tenericutes (OR: 0.77, 95% CI: 0.61–0.98, p = 0.034) on the risk of PROM.
GDM
The estimates of the IVW test suggested that the genetically predicted relative abundance of five genera, namely, Mollicutes (OR: 0.81, 95% CI: 0.67–0.97, p = 0.022), Methanobrevibacter (OR: 0.85, 95% CI: 0.73–1.00, p = 0.043), Oscillibacter (OR: 0.82, 95% CI: 0.71–0.96, p = 0.011), Euryarchaeota (OR: 0.87, 95% CI: 0.78–0.97, p = 0.013), and Tenericutes (OR: 0.81, 95% CI: 0.67–0.97, p = 0.022), was in negatively associated with the risk of GDM, whereas the genetically predicted relative abundance of nine other genera, namely, Betaproteobacteria (OR: 1.26, 95% CI: 1.00–1.58, p = 0.047), Lactobacillaceae (OR: 1.19, 95% CI: 1.02–1.39, p = 0.032), Collinsella (OR: 1.32, 95% CI: 1.01–1.74, p = 0.044), Coprobacter (OR: 1.21, 95% CI: 1.04–1.41, p = 0.015), Lachnoclostridium (OR: 1.37, 95% CI: 1.06–1.77, p = 0.017), Lactobacillus (OR: 1.24, 95% CI: 1.07–1.43, p = 0.004), Olsenella (OR: 1.17, 95% CI: 1.03–1.32, p = 0.016), Prevotella9 (OR: 1.16, 95% CI: 1.01–1.34, p = 0.036), and Ruminococcus2 (OR: 1.19, 95% CI: 1.00–1.42, p = 0.047), was positively associated with the risk of GDM.
PE or eclampsia
The genetically predicted abundance of Prevotellaceae (OR: 1.24, 95% CI: 1.00–1.52, p = 0.049) was associated with an increased risk of PE or eclampsia. By contrast, there was a tendency toward a protective effect of increasing genetically predicted abundance of Enterobacteriaceae (OR: 0.67, 95% CI: 0.49–0.93, p = 0.015), Enterorhabdus (OR: 0.75, 95% CI: 0.58–0.97, p = 0.027), Eubacterium ruminantium (OR: 0.87, 95% CI: 0.75–1.00, p = 0.049), Eubacterium ventriosum (OR: 0.73, 95% CI: 0.59–0.91, p = 0.006), Ruminococcaceae UCG002 (OR: 0.77, 95% CI: 0.63–0.93, p = 0.008), Ruminococcus1 (OR: 0.75, 95% CI: 0.56–1.00, p = 0.048), Tyzzerella3 (OR: 0.83, 95% CI: 0.72–0.97, p = 0.016), Enterobacterales (OR: 0.67, 95% CI: 0.49–0.93, p = 0.015), and Euryarchaeota (OR: 0.81, 95% CI: 0.72–0.92, p = 0.001) on PE or eclampsia.
Sensitivity analysis
We did not find pleiotropic effects among the selected SNPs using the MR-Egger and MR-PROSSO global test (p > 0.05) (Supplementary Tables S2, S3). No evidence of heterogeneity was observed between the selected IVs and APOs (Supplementary Table S4). In addition, we found no causal relationships between identified bacterial taxa and APOs that were driven by any single SNP (Supplementary Table S5).
Discussion
In the present study, we found causal relationships between genetically predicted abundance of specific bacterial taxa and six common pregnancy complications (PPH, AP, SAB, PROM, GDM, and PE or eclampsia) using large-scale GWAS summary data.
Considerable interest in the potential molecular mechanisms of the gut microbiome in the pathogenesis of PPH was evidenced by an abundance of prior studies. The gut microbiome composition was found to be involved in the pathogenesis of PPH by regulating inflammation, which are closely associated with development of PPH (Farhana et al., 2015; Yang et al., 2021). We successfully identified that Butyricimonas, Marvinbryantia, Oscillibacter, and Peptococcus were negatively associated with the risk of PPH. Butyricimonas, Marvinbryantia, and Oscillibacter can produce butyrate, which participates in regulating gut permeability and induces the inflammatory response (Jang et al., 2020; Chen B. D. et al., 2021; Chen Z. et al., 2021; Liu et al., 2022). Prior studies have reported that the abundance of Peptococcus increases when intestinal inflammation occurs, indicating its pro-inflammatory effects (Sun et al., 2022). The exact mechanism of Peptococcus on the pathogenesis of PPH warrants verification studies. This study also suggested that increased abundance of Deltaproteobacteria was causally related to a higher risk of PPH. Deltaproteobacteria is a common pathogenic bacterium, which initiates inflammation by activating the Notch signaling pathway and induces the expression of pro-inflammatory factors including pro-IL-1β and SOCS3 (Jin et al., 2016; Singh et al., 2021).
Emerging evidence has shown the role of perturbations in mitochondrial biogenesis, oxidative phosphorylation, and oxidative stress pathways in the pathogenesis of AP (Workalemahu et al., 2018). The relationship between gut microbiota dysbiosis and AP remains unclear. This study found that the abundance of Butyricimonas and Holdemanella reduced the risk of AP. The association may be mediated by the short-chain fatty acid butyrate as Butyricimonas and Holdemanella are involved in its production (Zagato et al., 2020; Jing et al., 2021). Butyrate has a key role in maintaining mucosal integrity, reducing inflammation via macrophage function, and regulating oxidative stress (Hu et al., 2020; Jing et al., 2021). We also showed that abundance of Eggerthella and Escherichia Shigella was positively associated with the risk of AP. Eggerthella is a pro-inflammatory genus, which has been associated with the depletion of butyrate (Nikolova et al., 2021). Escherichia Shigella can also mediate inflammation and oxidative stress by regulating the lipopolysaccharide/TLR4/NF-kB signaling pathway (Peng et al., 2020). This study demonstrated that Roseburia and Porphyromonadaceae, butyrate-producing bacteria, were positively related to the risk of AP (Wang et al., 2022). It can suggest that a single microbial taxon may not affect the development of AP, and the overall role of gut microbiota may have a more important role in the onset of AP. We showed that some novel gut bacteria including Acidaminococcaceae, Victivallaceae, Howardella, and Prevotella9 were closely associated with the pathogenesis of AP, but the exact mechanism of these gut microbiome on the development of AP warrants additional investigations.
We discovered that the abundance of Actinomyces, Subdoligranulum, and Veillonella was associated with an increased risk of SAB, while a high abundance of Lactococcus was related to a lower risk of SAB. Immunologic dysfunction and thrombophilia are known key factors in the pathogenesis of SAB (Liu et al., 2021a). A previous study showed that the relative abundance of Actinomyces and Subdoligranulum is closely related to multiple autoimmune diseases, including systemic lupus erythematosus and chronic spontaneous urticaria, suggesting that these bacteria play an important role in the host immune response (Chen B. D. et al., 2021; Liu et al., 2021). The protective effect of Lactococcus on SAB was likely due to lactic acid bacteria reducing autoimmunity and recovering immune homeostasis (Xu et al., 2020). Increased thrombophilia is one of the clinical signs of SAB (de Jong et al., 2014). Veillonella may influence the host trimethylamine N-oxide (TMAO) level by mediating carnitine production and metabolism, which contribute to platelet hyperreactivity and thrombosis (Yang et al., 2019). The exact the mechanism of gut microbiota leading to SAB remains poorly understood and warrants additional studies.
Although exact mechanisms underlying the occurrence of PROM are not fully elucidated, oxidative stress and inflammation have been reported to play an important role (Liu et al., 2021b). Our study revealed that the genetically predicted abundance of Turicibacter was positively correlated to the risk of PROM. Animal studies have reported that the abundance of Turicibacter was negatively associated with propionic acid and butyric acid, which can inhibit the inflammatory response (Wan et al., 2021). We have found that an increased abundance of Marvinbryantia and Ruminococcaceae, which are known to produce short-chain fatty acid, was negatively associated with the risk of PROM (Chen B. D. et al., 2021; Chen Z. et al., 2021).
Previous studies on GDM or post-GDM women with impaired glucose tolerance discovered a broad range of gut microbiota dysbiosis, which was associated with Collinsella (Crusell et al., 2018), Olsenella (Crusell et al., 2018), Prevotella9 (Hasain et al., 2020), and Ruminococcus2 (Crusell et al., 2018). On the other hand, beneficial butyrate-producing bacteria including Methanobrevibacter (Kuang et al., 2017) and Oscillibacter (Crusell et al., 2018) were depleted. A prior study also indicated that individuals with diminished insulin sensitivity had a lower abundance of Tenericutes, suggesting it might be associated with the occurrence and development of GDM (Yuan X. et al., 2021). Larsen et al. found that the relative abundance of Betaproteobacteria was higher in the type 2 diabetes group and positively correlated with the plasma glucose level (Larsen et al., 2010). These results were consistent with the present study. The mechanisms through which these bacteria exert beneficial or detrimental effects on the pathogenesis of GDM deserve to be studied.
Our study also showed that the abundance of Eubacterium ruminantium, Eubacterium ventriosum, Ruminococcaceae UCG002, Ruminococcus1, and Enterobacterales was negatively linked to the risk of PE or eclampsia. Prior studies indicated that chronic peripheral and placental inflammation were critical to the onset of PE (Black and Horowitz, 2018). Eubacterium, Ruminococcaceae, and Enterobacterales are involved in the production of short-chain fatty acids, such that their depletion is causally linked to inflammation (Tran et al., 2019; Zhuang et al., 2020). Wang et al. (2019) showed that fecal microbiota showed a significant reduction of Ruminococcus in the PE group, which was important in maintaining gut homeostasis. This study also demonstrated that some bacteria such as Prevotellaceae, Enterobacteriaceae, Enterorhabdus, and Euryarchaeota were involved in the pathogenesis of PE. Future studies are warranted to elucidate the functional significance of these bacteria in the onset of PE.
The advantages of this study as follows: First, the MR design diminishes the residual confounding and reverse causality and thereby improves the causal inference in relationships of gut microbiota and APOs. Second, we comprehensively studied up to six common APOs. Third, the identified causal associations in our MR analysis may provide candidate microbial taxa for subsequent functional studies and help in the development of new approaches for the prevention and treatment of pregnancy complications by targeting specific gut bacteria.
The limitations of our study must also be acknowledged. First, SNPs obtained based on the genome-wide statistical significance threshold (5 × 10−8) were too limited for further study; therefore, we included the SNPs that met the locus-wide significance level (1 × 10−5) in this study. Second, while the GWAS summary data in this study were obtained mostly from European populations, only a small amount of the gut microbiome data was taken from other races, which may partially bias our findings. Third, we cannot exclude the possible diet–gene or gene–environment interactions on outcomes, which may influence the observed results.
In conclusion, this MR study comprehensively explores causal effects of gut microbiota on APOs for the first time. Our findings provide new insights into the prevention, course of disease, and treatment of APOs by targeting specific bacteria taxa. Further studies are warranted to identify the exact mechanism of enteric microbiota–pregnancy complication associations.
Data availability statement
Summary data used for this study can be accessed through the following links: microbiota, https://mibiogen.gcc.rug.nl/; post-partum hemorrhage, https://r5.finngen.fi/pheno/O15_POSTPART_HEAMORRH; abruptio placenta, https://r5.finngen.fi/pheno/O15_PLAC_PREMAT_SEPAR; spontaneous abortion, https://r5.finngen.fi/pheno/O15_ABORT_SPONTAN; premature rupture of membrane, https://r5.finngen.fi/pheno/O15_MEMBR_PREMAT_RUPT; gestational diabetes mellitus, https://r5.finngen.fi/pheno/GEST_DIABETES; pre-eclampsia or eclampsia, https://r5.finngen.fi/pheno/O15_PRE_OR_ECLAMPSIA.
Ethics statement
This study was approved by Ethics Committee of Shengjing Hospital of China Medical University.
Author contributions
CLi and CLiu designed the research. CLi collected and analyzed the data and drafted the manuscript. CLiu and NL supervised the study. CLi, CLiu, and NL were involved in writing the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the National Key Research and Development Program of China (2021YFC2701503).
Acknowledgments
We thank the FinnGen consortium and MiBioGen consortium for providing GWAS summary statistics data for our analysis.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2022.1059281/full#supplementary-material
Abbreviations
MR, Mendelian randomization; APO, Adverse pregnancy outcome; GWAS, Genome-wide association study; IV, Instrumental variable; SNP, Single-nucleotide polymorphism; IVW, Inverse variance-weighted; MR-PRESSO, Mendelian randomization pleiotropy residual sum and outlier; GDM, Gestational diabetes mellitus; PE, Pre-eclampsia; PTB, Preterm birth; PPH, Postpartum hemorrhage; AP, Abruptio placenta; SAB, Spontaneous abortion; PROM, Premature rupture of membrane; LD, Linkage disequilibrium.
References
Black, K. D., and Horowitz, J. A. (2018). Inflammatory markers and preeclampsia: a systematic review. Nurs. Res. 67, 242–251. doi: 10.1097/NNR.0000000000000285
Bowden, J., Davey Smith, G., and Burgess, S. (2015). Mendelian randomization with invalid instruments: effect estimation and bias detection through Egger regression. Int. J. Epidemiol. 44, 512–525. doi: 10.1093/ije/dyv080
Burgess, S., Butterworth, A., and Thompson, S. G. (2013). Mendelian randomization analysis with multiple genetic variants using summarized data. Genet. Epidemiol. 37, 658–665. doi: 10.1002/gepi.21758
Burgess, S., and Thompson, S. G. (2015). Multivariable Mendelian randomization: the use of pleiotropic genetic variants to estimate causal effects. Am. J. Epidemiol. 181, 251–260. doi: 10.1093/aje/kwu283
Burgess, S., Thompson, S. G., and CRP CHD Genetics Collaboration. (2011). Avoiding bias from weak instruments in Mendelian randomization studies. Int. J. Epidemiol. 40, 755–764. doi: 10.1093/ije/dyr036
Chen, B. D., Jia, X. M., Xu, J. Y., Zhao, L. D., Ji, J. Y., Wu, B. X., et al. (2021). An autoimmunogenic and proinflammatory profile defined by the gut microbiota of patients with untreated systemic lupus erythematosus. Arthritis & rheumatology 73, 232–243. doi: 10.1002/art.41511
Chen, C., Chen, H., Zhang, Y., Thomas, H. R., Frank, M. H., He, Y., et al. (2020). TBtools: an integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 13, 1194–1202. doi: 10.1016/j.molp.2020.06.009
Chen, J., Bacelis, J., Sole-Navais, P., Srivastava, A., Juodakis, J., Rouse, A., et al. (2020). Dissecting maternal and fetal genetic effects underlying the associations between maternal phenotypes, birth outcomes, and adult phenotypes: a mendelian-randomization and haplotype-based genetic score analysis in 10,734 mother-infant pairs. PLoS Med. 17, e1003305. doi: 10.1371/journal.pmed.1003305
Chen, X., Li, P., Liu, M., Zheng, H., He, Y., Chen, M. X., et al. (2020). Gut dysbiosis induces the development of pre-eclampsia through bacterial translocation. Gut 69, 513–522. doi: 10.1136/gutjnl-2019-319101
Chen, Z., Radjabzadeh, D., Chen, L., Kurilshikov, A., Kavousi, M., Ahmadizar, F., et al. (2021). Association of insulin resistance and type 2 diabetes with gut microbial diversity: a microbiome-wide analysis from population studies. JAMA Netw Open 4, e2118811. doi: 10.1001/jamanetworkopen.2021.18811
Crusell, M., Hansen, T. H., Nielsen, T., Allin, K. H., Rühlemann, M. C., Damm, P., et al. (2018). Gestational diabetes is associated with change in the gut microbiota composition in third trimester of pregnancy and postpartum. Microbiome. 6, 89. doi: 10.1186/s40168-018-0472-x
Davey Smith, G., and Hemani, G. (2014). Mendelian randomization: genetic anchors for causal inference in epidemiological studies. Hum. Mol. Genet. 23, R89–98. doi: 10.1093/hmg/ddu328
de Jong, P. G., Kaandorp, S., Di Nisio, M., Goddijn, M., and Middeldorp, S. (2014). Aspirin and/or heparin for women with unexplained recurrent miscarriage with or without inherited thrombophilia. Cochrane Database Syst Rev CD004734. doi: 10.1002/14651858.CD004734.pub4
Di Simone, N., Santamaria Ortiz, A., Specchia, M., Tersigni, C., Villa, P., Gasbarrini, A., et al. (2020). Recent insights on the maternal microbiota: impact on pregnancy outcomes. Front. Immunol. 11, 528202. doi: 10.3389/fimmu.2020.528202
Egger, M., Smith, G. D., and Phillips, A. N. (1997). Meta-analysis: principles and procedures. BMJ 315, 1533–1537. doi: 10.1136/bmj.315.7121.1533
Ellervik, C., Roselli, C., Christophersen, I. E., Alonso, A., Pietzner, M., Sitlani, C. M., et al. (2019). Assessment of the relationship between genetic determinants of thyroid function and atrial fibrillation: a mendelian randomization study. JAMA Cardiol 4, 144–152. doi: 10.1001/jamacardio.2018.4635
Farhana, M., Tamura, N., Mukai, M., Ikuma, K., Koumura, Y., Furuta, N., et al. (2015). Histological characteristics of the myometrium in the postpartum hemorrhage of unknown etiology: a possible involvement of local immune reactions. J. Reprod. Immunol. 110, 74–80. doi: 10.1016/j.jri.2015.04.004
Ference, B. A., Ray, K. K., Catapano, A. L., Ference, T. B., Burgess, S., Neff, D. R., et al. (2019). Mendelian randomization study of ACLY and cardiovascular disease. N. Engl. J. Med. 380, 1033–1042. doi: 10.1056/NEJMoa1806747
Hasain, Z., Mokhtar, N. M., Kamaruddin, N. A., Mohamed Ismail, N. A., Razalli, N. H., Gnanou, J. V., et al. (2020). Gut microbiota and gestational diabetes mellitus: a review of host-gut microbiota interactions and their therapeutic potential. Front. Cell. Infect. Microbiol. 10, 188. doi: 10.3389/fcimb.2020.00188
Hemani, G., Zheng, J., Elsworth, B., Wade, K. H., Haberland, V., Baird, D., et al. (2018). The MR-Base platform supports systematic causal inference across the human phenome. Elife 7. doi: 10.7554/eLife.34408
Hu, S., Kuwabara, R., de Haan, B. J., Smink, A. M., and de Vos, P. (2020). Acetate and butyrate improve β-cell metabolism and mitochondrial respiration under oxidative stress. Int. J. Mol. Sci. 21. doi: 10.3390/ijms21041542
Jang, J. H., Yeom, M. J., Ahn, S., Oh, J. Y., Ji, S., Kim, T. H., et al. (2020). Acupuncture inhibits neuroinflammation and gut microbial dysbiosis in a mouse model of Parkinson's disease. Brain Behav. Immun. 89, 641–655. doi: 10.1016/j.bbi.2020.08.015
Jin, C., Zeng, Z., Fu, Z., and Jin, Y. (2016). Oral imazalil exposure induces gut microbiota dysbiosis and colonic inflammation in mice. Chemosphere. 160, 349–358. doi: 10.1016/j.chemosphere.2016.06.105
Jing, Y., Yu, Y., Bai, F., Wang, L., Yang, D., Zhang, C., et al. (2021). Effect of fecal microbiota transplantation on neurological restoration in a spinal cord injury mouse model: involvement of brain-gut axis. Microbiome. 9, 59. doi: 10.1186/s40168-021-01007-y
Kuang, Y. S., Lu, J. H., Li, S. H., Li, J. H., Yuan, M. Y., He, J. R., et al. (2017). Connections between the human gut microbiome and gestational diabetes mellitus. Gigascience. 6, 1–12. doi: 10.1093/gigascience/gix058
Kurilshikov, A., Medina-Gomez, C., Bacigalupe, R., Radjabzadeh, D., Wang, J., Demirkan, A., et al. (2021). Large-scale association analyses identify host factors influencing human gut microbiome composition. Nat. Genet. 53, 156–165. doi: 10.1038/s41588-020-00763-1
Larsen, N., Vogensen, F. K., van den Berg, F. W., Nielsen, D. S., Andreasen, A. S., Pedersen, B. K., et al. (2010). Gut microbiota in human adults with type 2 diabetes differs from non-diabetic adults. PLoS ONE. 5, e9085. doi: 10.1371/journal.pone.0009085
Lawlor, D. A., Harbord, R. M., Sterne, J. A., Timpson, N., and Davey Smith, G. (2008). Mendelian randomization: using genes as instruments for making causal inferences in epidemiology. Stat. Med. 27, 1133–1163. doi: 10.1002/sim.3034
Liu, R., Peng, C., Jing, D., Xiao, Y., Zhu, W., Zhao, S., et al. (2021). Biomarkers of gut microbiota in chronic spontaneous urticaria and symptomatic dermographism. Front. Cell. Infect. Microbiol. 11, 703126. doi: 10.3389/fcimb.2021.703126
Liu, X., Tong, X., Zou, Y., Lin, X., Zhao, H., Tian, L., et al. (2022). Mendelian randomization analyses support causal relationships between blood metabolites and the gut microbiome. Nat. Genet. 54, 52–61. doi: 10.1038/s41588-021-00968-y
Liu, Y., Chen, H., Feng, L., and Zhang, J. (2021a). Interactions between gut microbiota and metabolites modulate cytokine network imbalances in women with unexplained miscarriage. NPJ Biofilms Microb. 7, 24. doi: 10.1038/s41522-021-00199-3
Liu, Y., Wu, M., Song, L., Bi, J., Wang, L., Chen, K., et al. (2021b). Association between prenatal rare earth elements exposure and premature rupture of membranes: results from a birth cohort study. Environ. Res. 193, 110534. doi: 10.1016/j.envres.2020.110534
Luo, Q., Hu, Y., Chen, X., Luo, Y., Chen, J., and Wang, H. (2022). Effects of gut microbiota and metabolites on heart failure and its risk factors: a two-sample mendelian randomization study. Front Nutr 9, 899746. doi: 10.3389/fnut.2022.899746
Nikolova, V. L., Hall, M., Hall, L. J., Cleare, A. J., Stone, J. M., and Young, A. H. (2021). Perturbations in gut microbiota composition in psychiatric disorders: a review and meta-analysis. JAMA Psychiatry 78, 1343–1354. doi: 10.1001/jamapsychiatry.2021.2573
Peng, L., Gao, X., Nie, L., Xie, J., Dai, T., Shi, C., et al. (2020). Astragalin attenuates dextran sulfate sodium (DSS)-induced acute experimental colitis by alleviating gut microbiota dysbiosis and inhibiting NF-κB activation in mice. Front. Immunol. 11, 2058. doi: 10.3389/fimmu.2020.02058
Pierce, B. L., Ahsan, H., and Vanderweele, T. J. (2011). Power and instrument strength requirements for Mendelian randomization studies using multiple genetic variants. Int. J. Epidemiol. 40, 740–752. doi: 10.1093/ije/dyq151
Qi, X., Yun, C., Pang, Y., and Qiao, J. (2021). The impact of the gut microbiota on the reproductive and metabolic endocrine system. Gut Microbes 13, 1–21. doi: 10.1080/19490976.2021.1894070
Rana, S., Lemoine, E., Granger, J. P., and Karumanchi, S. A. (2019). Preeclampsia: pathophysiology, challenges, and perspectives. Circ. Res. 124, 1094–1112. doi: 10.1161/CIRCRESAHA.118.313276
Sanna, S., van Zuydam, N. R., Mahajan, A., Kurilshikov, A., Vich Vila, A., Võsa, U., et al. (2019). Causal relationships among the gut microbiome, short-chain fatty acids and metabolic diseases. Nat. Genet. 51, 600–605. doi: 10.1038/s41588-019-0350-x
Singh, S. B., Coffman, C. N., Carroll-Portillo, A., Varga, M. G., and Lin, H. C. (2021). Notch signaling pathway is activated by sulfate reducing bacteria. Front. Cell. Infect. Microbiol. 11, 695299. doi: 10.3389/fcimb.2021.695299
Sun, L., Guo, L., Xu, G., Li, Z., Appiah, M. O., Yang, L., et al. (2022). Quercetin reduces inflammation and protects gut microbiota in broilers. Molecules 27. doi: 10.3390/molecules27103269
Tran, T., Corsini, S., Kellingray, L., Hegarty, C., Le Gall, G., Narbad, A., et al. (2019). APOE genotype influences the gut microbiome structure and function in humans and mice: relevance for Alzheimer's disease pathophysiology. FASEB J. 33, 8221–8231. doi: 10.1096/fj.201900071R
Verbanck, M., Chen, C. Y., Neale, B., and Do, R. (2018). Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nat. Genet. 50, 693–698. doi: 10.1038/s41588-018-0099-7
Wan, F., Zhong, R., Wang, M., Zhou, Y., Chen, Y., Yi, B., et al. (2021). Caffeic acid supplement alleviates colonic inflammation and oxidative stress potentially through improved gut microbiota community in mice. Front. Microbiol. 12, 784211. doi: 10.3389/fmicb.2021.784211
Wang, J., Gu, X., Yang, J., Wei, Y., and Zhao, Y. (2019). Gut microbiota dysbiosis and increased plasma LPS and TMAO levels in patients with preeclampsia. Front. Cell. Infect. Microbiol. 9, 409. doi: 10.3389/fcimb.2019.00409
Wang, J., Kurilshikov, A., Radjabzadeh, D., Turpin, W., Croitoru, K., Bonder, M. J., et al. (2018). Meta-analysis of human genome-microbiome association studies: the MiBioGen consortium initiative. Microbiome 6, 101. doi: 10.1186/s40168-018-0479-3
Wang, X., Yu, C., Liu, X., Yang, J., Feng, Y., Wu, Y., et al. (2022). Fenofibrate ameliorated systemic and retinal inflammation and modulated gut microbiota in high-fat diet-induced mice. Front. Cell. Infect. Microbiol. 12, 839592. doi: 10.3389/fcimb.2022.839592
Workalemahu, T., Enquobahrie, D. A., Gelaye, B., Thornton, T. A., Tekola-Ayele, F., Sanchez, S. E., et al. (2018). Abruptio placentae risk and genetic variations in mitochondrial biogenesis and oxidative phosphorylation: replication of a candidate gene association study. Am. J. Obstet. Gynecol. 219, e1–617.e17. doi: 10.1016/j.ajog.2018.08.042
Xu, H., Cao, J., Li, X., Lu, X., Xia, Y., Fan, D., et al. (2020). Regional differences in the gut microbiota and gut-associated immunologic factors in the ileum and cecum of rats with collagen-induced arthritis. Front. Pharmacol. 11, 587534. doi: 10.3389/fphar.2020.587534
Yang, S., Li, X., Yang, F., Zhao, R., Pan, X., Liang, J., et al. (2019). Gut microbiota-dependent marker TMAO in promoting cardiovascular disease: inflammation mechanism, clinical prognostic, and potential as a therapeutic target. Front. Pharmacol. 10, 1360. doi: 10.3389/fphar.2019.01360
Yang, X., He, Z., Hu, R., Yan, J., Zhang, Q., Li, B., et al. (2021). Dietary β-carotene on postpartum uterine recovery in mice: crosstalk between gut microbiota and inflammation. Front. Immunol. 12, 744425. doi: 10.3389/fimmu.2021.744425
Yin, C., Chen, J., Wu, X., Liu, Y., He, Q., Cao, Y., et al. (2021). Preterm birth is correlated with increased oral originated microbiome in the gut. Front. Cell. Infect. Microbiol. 11, 579766. doi: 10.3389/fcimb.2021.579766
Yuan, S., and Larsson, S. C. (2022). Coffee and caffeine consumption and risk of kidney stones: a mendelian randomization study. Am. J. Kidney Dis. 79, 9–14.e1. doi: 10.1053/j.ajkd.2021.04.018
Yuan, S., Liu, J., and Larsson, S. C. (2021). Smoking, alcohol and coffee consumption and pregnancy loss: a Mendelian randomization investigation. Fertil. Steril. 116, 1061–1067. doi: 10.1016/j.fertnstert.2021.05.103
Yuan, X., Chen, R., McCormick, K. L., Zhang, Y., Lin, X., and Yang, X. (2021). The role of the gut microbiota on the metabolic status of obese children. Microb. Cell Fact. 20, 53. doi: 10.1186/s12934-021-01548-9
Zagato, E., Pozzi, C., Bertocchi, A., Schioppa, T., Saccheri, F., Guglietta, S., et al. (2020). Endogenous murine microbiota member Faecalibaculum rodentium and its human homologue protect from intestinal tumour growth. Nat Microbiol 5, 511–524. doi: 10.1038/s41564-019-0649-5
Zheng, W., Xu, Q., Huang, W., Yan, Q., Chen, Y., Zhang, L., et al. (2020). Gestational diabetes mellitus is associated with reduced dynamics of gut microbiota during the first half of pregnancy. mSystems. 5, e00109–20. doi: 10.1128/mSystems.00109-20
Keywords: Mendelian randomization, instrumental variable, gut microbiota, adverse pregnancy outcomes, causal relationship
Citation: Li C, Liu C and Li N (2022) Causal associations between gut microbiota and adverse pregnancy outcomes: A two-sample Mendelian randomization study. Front. Microbiol. 13:1059281. doi: 10.3389/fmicb.2022.1059281
Received: 01 October 2022; Accepted: 11 November 2022;
Published: 16 December 2022.
Edited by:
Dongsheng Che, Jilin Agricultural University, ChinaCopyright © 2022 Li, Liu and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Na Li, lina_naonao@126.com