 Niccolo’ Meriggi
Niccolo’ Meriggi Monica Di Paola
Monica Di Paola Duccio Cavalieri
Duccio Cavalieri Irene Stefanini
Irene Stefanini- 1Department of Biology, University of Florence, Florence, Italy
- 2Department of Life Sciences and Systems Biology, University of Turin, Turin, Italy
Over the last few years, an increasing number of studies have reported the existence of an association between the budding yeast Saccharomyces cerevisiae and insects. The discovery of this relationship has called into question the hypothesis that S. cerevisiae is unable to survive in nature and that the presence of S. cerevisiae strains in natural specimens is the result of contamination from human-related environments. S. cerevisiae cells benefit from this association as they find in the insect intestine a shelter, but also a place where they can reproduce themselves through mating, the latter being an event otherwise rarely observed in natural environments. On the other hand, insects also take advantage in hosting S. cerevisiae as they rely on yeasts as nutriment to properly develop, to localize suitable food, and to enhance their immune system. Despite the relevance of this relationship on both yeast and insect ecology, we are still far from completely appreciating its extent and effects. It has been shown that other yeasts are able to colonize only one or a few insect species. Is it the same for S. cerevisiae cells or is this yeast able to associate with any insect? Similarly, is this association geographically or topographically limited in areas characterized by specific physical features? With this review, we recapitulate the nature of the S. cerevisiae-insect association, disclose its extent in terms of geographical distribution and species involved, and present YeastFinder, a cured online database providing a collection of information on this topic.
Saccharomyces cerevisiae in Natural Environments
Saccharomyces cerevisiae is widely used in the industry for winemaking, brewery, and bakery, as animal and human food supplement or probiotic (Palma et al., 2015), and for biofuel, flavorings, pharmaceuticals, and enzymes production (e.g., invertases, lactases) (Parapouli et al., 2020). The physiology and genetics of this yeast have been studied in depth, as well as molecular mechanisms shared with other eukaryotes (Resnick and Cox, 2000). Despite this broad range of applications, the natural diffusion and evolution of this yeast remained unexplored until recently. The ability to overgrow other microorganisms in fermentable substrates and the widespread use of S. cerevisiae in the industry of fermented products lead to the hypothesis that this yeast has been domesticated and is confined to human activities (Gallone et al., 2016). However, over the last few decades, this hypothesis was compromised by the isolation of strains from natural environments (e.g., soil, barks, and water) and by the observation of the existence of strains genetically different from those used in the industry (Liti, 2015). A new hypothesis was proposed: a neutral model in which S. cerevisiae is functionally adapted to a range of different environments (Goddard and Greig, 2015). Despite improving our understanding of the natural spread of the budding yeast, these new findings did not clarify (i) how the yeast can survive when nutrient sources are lacking, (ii) where it was before humans started using it to ferment food, and (iii) how can this not airborne microorganism move among different environmental sources. The identification of the association between the budding yeast and social wasps helped fill in several of the gaps in our knowledge on S. cerevisiae natural cycle. Previous studies have already shown some aspects of yeast-insect associations (e.g., Stefanini, 2018 and Blackwell, 2017). This review recapitulates the current knowledge exclusively on associations involving S. cerevisiae also addressing new aspects: the extent and geographical distribution.
Saccharomyces cerevisiae-Insects Association: the Yeast Side of the Coin
Social wasps and hornets are omnivorous insects that move around a broad foraging area and visit substrates that can be colonized by S. cerevisiae strains. Among the substrates visited by wasps, grape skin is the main source of environmental S. cerevisiae strains, even if this yeast represents only a minor component of the residing microbial communities. Polistes dominula (social wasps) and Vespa crabro (hornets) host in their intestines S. cerevisiae cells all year long, thus providing an environment in which yeasts can reside and survive in the seasonal period with less access to sugary sources (Stefanini et al., 2012). Insects can spread S. cerevisiae cells among environmental substrates and, also, share them within the colony, passing it among adults and to larvae (Stefanini et al., 2012). Thus, the capability of spreading yeast cells increases exponentially with the increase of the insect colony. This has pivotal importance for the diffusion of S. cerevisiae cells in the wild, especially considering that the increase in the demographic rate of the wasp colony occurs at the same time of grape ripening (Stefanini et al., 2012). Hence, the high frequency of S. cerevisiae cells isolated after the ripening period correlates these insects to the dispersion of yeasts in the vineyard (Stefanini et al., 2012). In addition to social wasps and hornets, other insects bear and spread S. cerevisiae cells, as proven in laboratory conditions for Drosophila spp. (Christiaens et al., 2014) and confirmed in the wild for bees. In fact, S. cerevisiae strains isolated from vineyard specimens are highly similar to strains isolated from bees caught in the same geographic region, suggesting that insects are responsible for the local dispersion of yeast cells (Goddard et al., 2010). Interestingly, the genetic and phenotypic diversity of S. cerevisiae does not affect the capability of different yeast strains to survive in the insect intestines (Dapporto et al., 2016; Ramazzotti et al., 2019) or to produce volatile metabolites attracting insects (Palanca et al., 2013). Hence, the whole genetic and phenotypic variability of S. cerevisiae can potentially attract and be vectored among natural specimens thanks to insects (Stefanini et al., 2012; Dapporto et al., 2016).
Saccharomyces cerevisiae mating is infrequent in nature, possibly because wild yeast cells are mainly diploid and hence need to face sporulation and germination to be able to mate with other strains, conditions that rarely occur in nature (Cubillos et al., 2009). Contrarily, yeast mating does occur within insect intestines (Stefanini et al., 2016; Figure 1). S. cerevisiae spores can survive in the intestinal tract of Drosophila melanogaster, and, by passing through the insect intestines, the sporal ascus is broken and hence the mating among yeast strains is facilitated (Reuter et al., 2007). The differences among chemical and physiological characteristics of different tracts of the insect intestine could offer a series of environmental changes (Engel and Moran, 2013) promoting yeast sporulation and germination, and hence mating, as shown by experiments carried out in the laboratory (Stefanini et al., 2016). Hence, the intestine not only promotes the yeast ascus break, but also diploid yeast cells sporulation and yeast spores germination, and thus allowing the mating among potentially any yeast strain and ploidy (Stefanini et al., 2016).
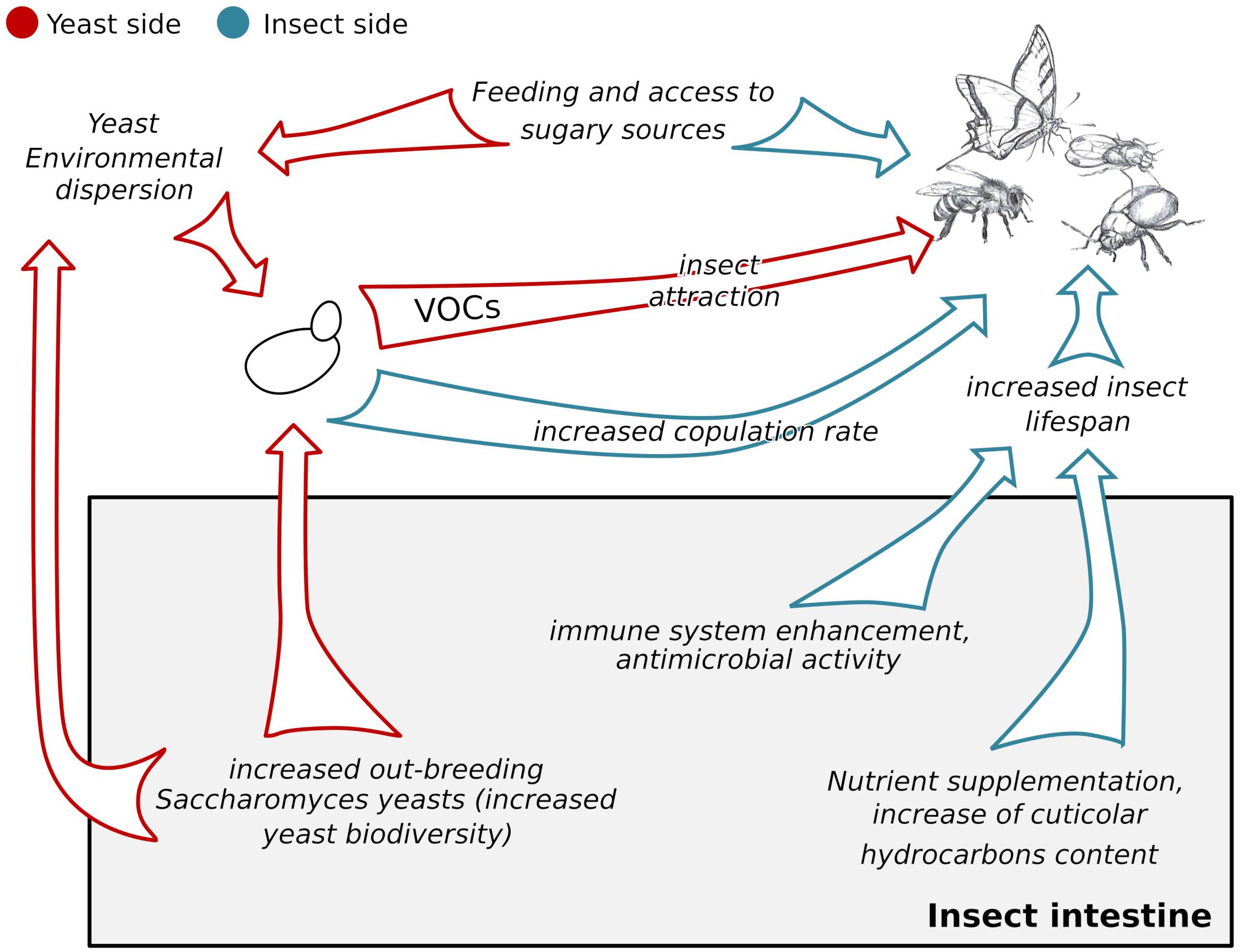
Figure 1. Schematic representation of the impacts of the S. cerevisiae-insects association on the yeast and on the insect. Red arrows indicate the impacts of the association on yeasts, blue arrows indicate the impacts of the association on insects. The gray box indicates effects of the association occurring within the insect intestines, both on S. cerevisiae and on the insects. VOCs = Volatile Organic Compounds.
Saccharomyces cerevisiae-Insects Association: the Insect Side of the Coin
The S. cerevisiae-insect association has beneficial effects not only on the yeast, but also on its counterpart: insects (Figure 1).
The capability to detect food is fundamental for insect survival to obtain the nourishment and find an environment suitable for oviposition. S. cerevisiae attracts various insect species including D. melanogaster (Becher et al., 2012), Vespula germanica, and V. vulgaris (Babcock et al., 2017) to food, which is otherwise less appealing. The main features making this yeast capable of enticing insects are the presence of functional mitochondria (Schiabor et al., 2014) and the capability of producing volatile compounds, such as isoamyl acetate and ethyl acetate (Christiaens et al., 2014), which are produced at high levels by yeasts isolated from wasp intestines (Dapporto et al., 2016). Overall, these findings show that the presence of fermenting S. cerevisiae cells is a strong cue used by insects to detect sugary substrates. It is fair to consider that, besides S. cerevisiae, other microorganisms present on sugary substrates, such as Hanseniaspora spp. and Gluconobacter spp. (Bueno et al., 2019), and vectored by insects (Palanca et al., 2013; Quan and Eisen, 2018) could produce aromas attracting insects. Interestingly, species-specific attractions have been observed. While D. melanogaster is attracted by S. cerevisiae, D. simulans is indifferent to this yeast species (Gunther et al., 2019). At the same time, D. melanogaster and the subgenus Sophophora are preferentially attracted to baits seeded with Hanseniaspora uvarum than to S. cerevisiae and forest-dwelling Drosophila species (e.g., D. tripunctata and the guarani group) are more attracted by S. cerevisiae than by H. uvarum (Batista et al., 2017). The differential preferences of Drosophila spp. towards different yeast species may mirror what happens in the yeast–insect–morning glory ecosystem, where beetles and yeasts (mostly Metschnikowia spp. and Candida spp.) reciprocally influence the occupancy of associations-specific niches (Stefanini, 2018).
In the laboratory, insect rearing is based on the use of media providing the full range of nutrients to support larval development but also appealing to female adults and promoting egg deposition (Piper, 2017). To this aim, it is a common practice to use media including S. cerevisiae (Becher et al., 2012; Grangeteau et al., 2018). In fact, the yeast supports larval development mostly by providing nicotinic acid, vitamin B, pantothenic acid, inositol, choline, beta-alanine, and pimelic acid (Tatum, 1941). The presence of S. cerevisiae in the larval diet also defines the fitness and behavior of juvenile and adult insects (Grangeteau et al., 2018). Providing live S. cerevisiae cells to Drosophila larvae improves the copulation rate, increases the cuticular hydrocarbon content, extends the insect life, and makes adults preferring food supplemented with the yeast, compared to a diet based on yeast extracts or lacking the yeast (Grangeteau et al., 2018; Murgier et al., 2019). Transcriptional analyses carried out on D. melanogaster adults developed from germ-free larvae showed the over-expression of genes involved in several metabolic pathways, if the insect diet was supplemented with live S. cerevisiae compared to single or multiple bacterial species (Elya et al., 2016). Despite this effect being observed only in intestinal cells and not at the whole-body level, hence suggesting the impact of yeast on insects is only local, it has to be considered that over the developmental process genes expression is rapidly regulated by multiple factors. Thus, time-course transcriptional analyses would be required to appreciate at the molecular level the impact of S. cerevisiae on insect development. The impact of S. cerevisiae on development and related traits varies according to the yeast species. For instance, even if a substrate supplemented with the budding yeast improves the survival of adult mosquitoes (Culex pipiens) compared to substrates supplemented with other yeast species, it is not preferred for oviposition by gravid C. pipiens females (Díaz-Nieto et al., 2016).
As in bigger animals, even in insects the fate and impact of the encountered microorganisms are often determined by the host immune system. The insect immune system includes a cellular and a humoural component (Lemaitre and Hoffmann, 2007). Although several receptors on immune cells have been described as responsible for the recognition of microorganisms, their role in the response to S. cerevisiae and yeasts in general is still unclear (Lu and St Leger, 2016). Conversely, an entire pathway of the humoral response is responsible for the insect’s reaction to yeasts: the Toll signaling pathway (Roh et al., 2009). When triggered by yeast cell wall β-glucans and proteases, this pathway induces the expression of Drosomycin, an antimicrobial agent (Gottar et al., 2006). Insects can also fight potential yeast pathogens through the Duox response pathway, which induces the production of not-specific antimicrobial reactive oxygen species (Hoang et al., 2015). To note, the vast majority of information on the insect immune response to S. cerevisiae has been obtained by using the Drosophila spp. model that presents perturbations in the Toll pathway (Alarco et al., 2004), making this insect susceptible to the budding yeast (Lionakis, 2011). Alternatively, the use of Galleria mellonella, naturally susceptible to S. cerevisiae, yielded fundamental information that could not be gathered with fruit flies. For instance, G. mellonella provided the first insights on the capability of a pre-exposure to S. cerevisiae cells or glucans to protect the insect against a subsequent infection with a lethal dose of Candida albicans (Bergin et al., 2006). This immune-enhancing elicited by S. cerevisiae has been recently confirmed in the social wasps Polistes dominula, which become more resistant to Escherichia coli infections upon pre-immunization with the yeast (Meriggi et al., 2019).
Biogeography and Diffusion
Insects can colonize habitats with extremely different characteristics and are considered, as defined by E.O. Wilson, “the little things that run the world” (Wilson, 1987). The large number of insect species makes it impossible to comprehensively analyze their biodiversity. Of the estimated 6 million species, only 1 million are known (Larsen et al., 2017). For a matter of clarity, we will report here information on insects broadly grouped. We have, however, created a detailed on-line database that can be browsed by the reader (YeastFinder,1). S. cerevisiae has been identified, through isolation or metabarcoding, in the intestine or on the body of several insects all over the World (Figure 2). Notably, studies carried out so far lack consistency in the methods adopted for yeast isolation, and this may greatly impact the capability of identifying S. cerevisiae associated with insects. However, in this review we will neglect the heterogeneity of the adopted methods, leaving the exploration of this topic to dedicated future studies. Interestingly, S. cerevisiae has not been found in insects caught in South Africa (SAf), Ecuador, Thailand, Indonesia, Nigeria (the only study on termites), Iran, Japan (J), Malaysia, and Central America (CA: Panama, Costa Rica, and Guatemala). It is worth to consider that the investigations carried out in Ecuador and Malaysia (Freitas et al., 2013), Thailand (Saksinchai et al., 2015), Indonesia (Basukriadi et al., 2010), Nigeria (Adelabu et al., 2019), and Iran (Siavoshi et al., 2018) are related to individual studies, hence the lack of identification of S. cerevisiae could be ascribed to the procedure adopted for yeast isolation. On the other hand, multiple studies failed in identifying S. cerevisiae in association with beetles, honeybees and mosquitoes, butterflies, mites, and moths collected in J (Toki et al., 2012; Ninomiya et al., 2013), SAf (de Vega et al., 2012; Steyn et al., 2016), and CA (Lachance et al., 2001a, b, 2006, Suh and Blackwell, 2006; Suh et al., 2006, 2007; Rivera et al., 2009; Urbina et al., 2013; Ravenscraft et al., 2018). The lack of identification of S. cerevisiae in insects of these areas could indicate an unusual situation that is worth to be further investigated. All the insects investigated in New Zealand, Taiwan, and Seychelles Islands bore S. cerevisiae (Figure 2). However, the number of cases studied in these areas is low (n = 6), and hence this observation may be poorly representative of the real situation.
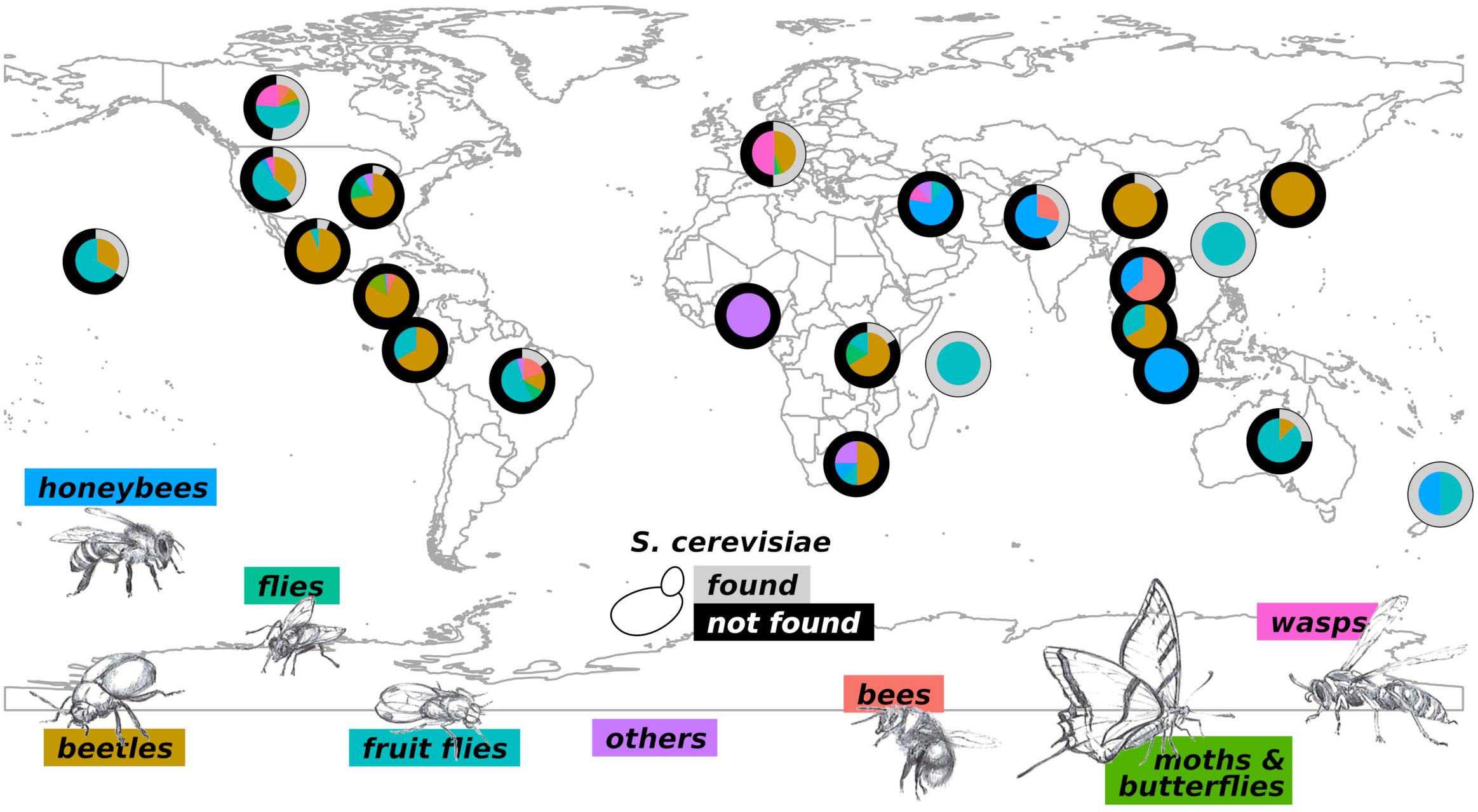
Figure 2. Biogeography and extent of the S. cerevisiae-insect association. For each geographical location, the outer ring of the pie chart shows the percentages of insects in which S. cerevisiae has been (gray) or has not been (black) identified. The inner parts of pie charts show the types of insects investigated in the corresponding area and the relative proportions; colors are indicated for each insect group at the bottom of the figure. The group “others” refers to insects poorly represented: ants, lacewings, termites, mites, and mosquitoes.
Multiple studies investigating various insects in other locations (shown in Figure 2) indicated an even geographical distribution of S. cerevisiae. The budding yeast has not been isolated from ants, lacewings, termites, mites, and mosquitoes (the group “others” in Figure 2), which were, however, poorly investigated (two species in different locations at most) (Carreiro et al., 1997; Lachance et al., 2003; Suh et al., 2005; Nguyen et al., 2006; Steyn et al., 2016; Siavoshi et al., 2018). Similarly, S. cerevisiae has not been found in butterflies (n = 11 species in different locations) and moths (n = 4) (Suh et al., 2006; Witzgall et al., 2012; Ravenscraft et al., 2018). Interestingly, only the 0.03% of beetles, which have been widely investigated (n = 236), bear S. cerevisiae (Kurtzman and Robnett, 1998; Lachance et al., 2001a, b, 2006; Six, 2003; Suh and Blackwell, 2004, Suh et al., 2005, 2006, 2007, 2013; Delalibera et al., 2005; Nguyen et al., 2006; Rosa et al., 2007; Rivera et al., 2009; de Vega et al., 2012; Hui et al., 2012; Toki et al., 2012; Freitas et al., 2013; Kaltenpoth and Steiger, 2013; Ninomiya et al., 2013; Urbina et al., 2013; Cline et al., 2014; Ren et al., 2014, 2015; Liu et al., 2016; Tanahashi and Hawes, 2016; Wang et al., 2016; Briones-Roblero et al., 2017; Chai et al., 2019). Similarly, bees only accidentally bear S. cerevisiae, with only 1 occurrence over 21 reported cases (Sandhu and Waraich, 1985; Lachance et al., 2003; Rosa et al., 2003; Daniel et al., 2013; Charron et al., 2014; Saksinchai et al., 2015). Conversely, S. cerevisiae has been found in a large portion of investigated flies, fruit flies, honey-bees, and wasps (29, 57, 20, and 71%, respectively) (Phaff and Knapp, 1956; Batra et al., 1973; Sandhu and Waraich, 1985; Morais et al., 1993, 1994; Rosa et al., 1994; Lachance et al., 1995, 2003, 2006; Suh et al., 2005; Nguyen et al., 2006, 2007; Basukriadi et al., 2010; Goddard et al., 2010; Chandler et al., 2012; de Vega et al., 2012; Hamby et al., 2012; Stefanini et al., 2012; Freitas et al., 2013; Buser et al., 2014; Charron et al., 2014; Lam and Howell, 2015; Saksinchai et al., 2015; Batista et al., 2017; Deutscher et al., 2017; Jimenez et al., 2017; Piper et al., 2017; Quan and Eisen, 2018; Siavoshi et al., 2018; dos Santos et al., 2019; Meriggi et al., 2019; Park et al., 2019). A few possible scenarios could explain the higher occurrence of S. cerevisiae in these groups of insects: (i) they are more prone to visit human-related environments, such as wineries and vineyards, that are likely to host higher amounts of S. cerevisiae cells, (ii) they are more attracted by substrates inhabited by the budding yeast compared to other insects, (iii) diet and physical-chemical intestine conditions facilitate the housing of S. cerevisiae.
Conclusion and Perspectives
According to the reports gathered for this review, it appears that every group of insects can bear S. cerevisiae, but only further and more detailed studies investigating a higher number of a broader range of insect species, as well as the standardization of isolation and identification methodologies, will consolidate this observation. Also, further studies on the geographical extent of this phenomenon would allow evaluating the existence of different physiological characteristics among insect species that favor or prevent the instauration of the association with the budding yeast. Aiming at this, it will be fundamental to also include groups neglected so far. For instance, planthoppers, mosquitoes, and spiders have not or have only poorly been investigated, albeit they could represent an unprecedented source of information as they visit and forage on a broad range of environmental sources. In addition, S. cerevisiae has been shown to have an impact on spiders’ behavior and health (Tietjen et al., 1987; Patt et al., 2012), and, especially considering that spiders are mostly carnivores, exploring this association would provide insightful information on the role of this yeast in prey hunting and interactions among species.
Overall, the reports published so far depict a tangled relationship between insect and yeast, in which various factors define the insect attraction to yeasts and the impact of this yeast on insect health. Understanding the factors responsible for the attraction of insects by yeasts, also by further exploring the differences among insect species has also important applications. A better understanding of the factors regulating this complex field will provide relevant information potentially useful to ideate approaches to use S. cerevisiae as a promoter of insect health or as a pest control. For instance, dissecting the capability of enhancing the host immune reaction against pathogenic microorganisms would be very useful in the fight against the worldwide decline of honeybees and pollinators (Wagner, 2020).
Only further studies will allow us to fully unravel the influence of S. cerevisiae on insects, and the potential applications of strains isolated from this natural source.
Author Contributions
IS ideated the review. NM and IS gathered the data. DC, MD, NM, and IS wrote the manuscript. All authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to thank Miss Marta Mocciaro for drawing the insect’s images. This work was supported by the University of Turin (Local Research Funds). NM, MD, and DC were supported by the University of Florence and by the Regione Toscana POR FSE 2014-2020, VESPATER project.
Footnotes
References
Adelabu, B., Kareem, S. O., Adeogun, A. I., and Wakil, S. M. (2019). Optimization of cellulase enzyme from sorghum straw by yeasts isolated from plant feeding-termite Zonocerus variegatus. Food Appl. Biosci. J. 7, 81–101.
Alarco, A. M., Marcil, A., Chen, J., Suter, B., Thomas, D., and Whiteway, M. (2004). Immune-deficient Drosophila melanogaster: a model for the innate immune response to human fungal pathogens. J. Immunol. 172, 5622–5628. doi: 10.4049/jimmunol.172.9.5622
Babcock, T., Gries, R., Borden, J., Plamero, L., Mattiacci, A., Masciocchi, M., et al. (2017). Brewer’s yeast, Saccharomyces cerevisiae, enhances attraction of two invasive yellowjackets (Hymenoptera: Vespidae) to dried fruit and fruit powder. J. Insect Sci. 17:91.
Basukriadi, A., Sjamsuridzal, W., and Beristama Putra, B. (2010). Molecular identification and diversity of yeasts associated with Apis cerana foraging on flowers of Jatropha integerrima. Microbiol. Indones. 4, 44–48. doi: 10.5454/mi.4.1.9
Batista, M. R., Uno, F., Chaves, R. D., Tidon, R., Rosa, C. A., and Klaczko, L. B. (2017). Differential attraction of drosophilids to banana baits inoculated with Saccharomyces cerevisiae and Hanseniaspora uvarum within a Neotropical forest remnant. PeerJ 5:e3063. doi: 10.7717/peerj.3063
Batra, L. R., Batra, S. W. T., and Bohart, G. E. (1973). The mycoflora of domesticated and wild bees (Apoidea). Mycopathol. Mycol. Appl. 49, 13–44. doi: 10.1007/bf02057445
Becher, P. G., Flick, G., Rozpêdowska, E., Schmidt, A., Hagman, A., Lebreton, S., et al. (2012). Yeast, not fruit volatiles mediate Drosophila melanogaster attraction, oviposition and development. Funct. Ecol.26, 822–828. doi: 10.1111/j.1365-2435.2012.02006.x
Bergin, D., Murphy, L., Keenan, J., Clynes, M., and Kavanagh, K. (2006). Pre-exposure to yeast protects larvae of Galleria mellonella from a subsequent lethal infection by Candida albicans and is mediated by the increased expression of antimicrobial peptides. Microbes Infect. 8, 2105–2112. doi: 10.1016/j.micinf.2006.03.005
Blackwell, M. (2017). Made for each other: ascomycete yeasts and insects. Microbiol. Spectr. 5, 1–18. doi: 10.1128/microbiolspec.FUNK-0081-2016
Briones-Roblero, C. I., Rodríguez-Díaz, R., Santiago-Cruz, J. A., Zúñiga, G., and Rivera-Orduña, F. N. (2017). Degradation capacities of bacteria and yeasts isolated from the gut of Dendroctonus rhizophagus (Curculionidae: Scolytinae). Folia Microbiol. 62, 1–9. doi: 10.1007/s12223-016-0469-4
Bueno, E., Martin, K. R., Raguso, R. A., Mcmullen, J. G., Hesler, S. P., Loeb, G. M., et al. (2019). Response of wild spotted wing Drosophila (Drosophila suzukii) to microbial volatiles. J. Chem. Ecol. doi: 10.1007/s10886-019-01139-4 [Epub ahead of print].
Buser, C. C., Newcomb, R. D., Gaskett, A. C., and Goddard, M. R. (2014). Niche construction initiates the evolution of mutualistic interactions. Ecol. Lett. 17, 1257–1264. doi: 10.1111/ele.12331
Carreiro, S. C., Pagnocca, F. C., Bueno, O. C., Júnior, M. B., Hebling, M. J. A., and da Silva, O. A. (1997). Yeasts associated with nests of the leaf-cutting ant Atta sexdens rubropilosa Forel, 1908. Antonie Van Leeuwenhoek 71, 243–248.
Chai, C. Y., Huang, L. N., Cheng, H., Liu, W. J., and Hui, F. L. (2019). Metschnikowia baotianmanensis f.a., sp. nov., a new yeast species isolated from the gut of the rhinoceros beetle Allomyrina dichotoma. Int. J. Syst. Evol. Microbiol. 69, 3087–3092. doi: 10.1099/ijsem.0.003593
Chandler, J. A., Eisen, J. A., and Kopp, A. (2012). Yeast communities of diverse Drosophila species: comparison of two symbiont groups in the same hosts. Appl. Environ. Microbiol. 78, 7327–7336. doi: 10.1128/aem.01741-12
Charron, G., Leducq, J. B., Bertin, C., Dubé, A. K., and Landry, C. R. (2014). Exploring the northern limit of the distribution of Saccharomyces cerevisiae and Saccharomyces paradoxus in North America. FEMS Yeast Res. 14, 281–288. doi: 10.1111/1567-1364.12100
Christiaens, J. F., Franco, L. M., Cools, T. L., De Meester, L., Michiels, J., Wenseleers, T., et al. (2014). The fungal aroma gene ATF1 promotes dispersal of yeast cells through insect vectors. Cell Rep. 9, 425–432. doi: 10.1016/j.celrep.2014.09.009
Cline, A. R., Skelley, P. E., Kinnee, S. A., Rooney-Latham, S., Winterton, S. L., Borkent, C. J., et al. (2014). Interactions between a sap beetle, sabal palm, scale insect, filamentous fungi and yeast, with discovery of potential antifungal compounds. PLoS One 9:e89295. doi: 10.1371/journal.pone.0089295
Cubillos, F. A., Vasquez, C., Faugeron, S., Ganga, A., and Martínez, C. (2009). Self-fertilization is the main sexual reproduction mechanism in native wine yeast populations. FEMS Microbiol. Ecol. 67, 162–170. doi: 10.1111/j.1574-6941.2008.00600.x
Daniel, H. M., Rosa, C. A., Sao Thiago-Calac, P. S., Antonini, J., Bastos, E. M. A. F., Evrard, P., et al. (2013). Starmerella neotropicalis f.a., sp. nov., a yeast species found in bees and pollen. Int. J. Syst. Evol. Microbiol. 63, 3896–3903. doi: 10.1099/ijs.0.055897-0
Dapporto, L., Stefanini, I., Rivero, D., Polsinelli, M., Capretti, P., De Marchi, P., et al. (2016). Social wasp intestines host the local phenotypic variability of Saccharomyces cerevisiae strains. Yeast 33, 277–287. doi: 10.1002/yea.3173
de Vega, C., Guzman, B., Lachance, M. A., Steenhuisen, S. L., Johnson, S. D., and Herrera, C. M. (2012). Metschnikowia proteae sp. nov., a nectarivorous insect-associated yeast species from Africa. Int. J. Syst. Evol. Microbiol. 62, 2538–2545. doi: 10.1099/ijs.0.040790-0
Delalibera, I., Handelsman, J., and Raffa, K. F. (2005). Contrasts in cellulolytic activities of gut microorganisms between the wood borer, Saperda vestita (Coleoptera: Cerambycidae), and the bark beetles, Ips pini and Dendroctonus frontalis (Coleoptera: Curculionidae). Environ. Entomol. 34, 541–552.
Deutscher, A. T., Reynolds, O. L., and Chapman, T. A. (2017). Yeast: an overlooked component of Bactrocera tryoni (Diptera: Tephritidae) larval gut microbiota. J. Econ. Entomol. 110, 298–300. doi: 10.1093/jee/tow262
Díaz-Nieto, L. M., D’Alessio, C., Perotti, M. A., and Berón, C. M. (2016). Culex pipiens development is greatly influenced by native bacteria and exogenous yeast. PLoS One 11:e0153133. doi: 10.1371/journal.pone.0153133
dos Santos, T. T., de Oliveira, D. P., Ramos Cabette, H. S., and de Morais, P. B. (2019). The digestive tract of Phylloicus (Trichoptera: Calamoceratidae) harbours different yeast taxa in Cerrado streams, Brazil. Symbiosis 77, 147–160. doi: 10.1007/s13199-018-0577-9
Elya, C., Zhang, V., Ludington, W. B., and Eisen, M. B. (2016). Stable host gene expression in the gut of adult Drosophila melanogaster with different bacterial mono-associations. PLoS One 11:e0167357. doi: 10.1371/journal.pone.0167357
Engel, P., and Moran, N. A. (2013). The gut microbiota of insects–diversity in structure and function. FEMS Microbiol. Rev. 37, 699–735. doi: 10.1111/1574-6976.12025
Freitas, L. F., Barriga, E. J. C., Barahona, P. P., Lachance, M. A., and Rosa, C. A. (2013). Kodamaea transpacifica fa, sp. nov., a yeast species isolated from ephemeral flowers and insects in the Galápagos Islands and Malaysia: further evidence for ancient human transpacific contacts. Int. J. Syst. Evol. Microbiol. 63, 4324–4329. doi: 10.1099/ijs.0.052282-0
Gallone, B., Steensels, J., Prahl, T., Soriaga, L., Saels, V., Herrera-Malaver, B., et al. (2016). Domestication and divergence of Saccharomyces cerevisiae beer yeasts. Cell 166, 1397–1410.
Goddard, M. R., Anfang, N., Tang, R., Gardner, R. C., and Jun, C. (2010). A distinct population of Saccharomyces cerevisiae in New Zealand: evidence for local dispersal by insects and human-aided global dispersal in oak barrels. Environ. Microbiol. 12, 63–73. doi: 10.1111/j.1462-2920.2009.02035.x
Goddard, M. R., and Greig, D. (2015). Saccharomyces cerevisiae: a nomadic yeast with no niche? FEMS Yeast Res. 15:fov009.
Gottar, M., Gobert, V., Matskevich, A. A., Reichhart, J. M., Wang, C., Butt, T. M., et al. (2006). Dual detection of fungal infections in Drosophila via recognition of glucans and sensing of virulence factors. Cell 127, 1425–1437. doi: 10.1016/j.cell.2006.10.046
Grangeteau, C., Yahou, F., Everaerts, C., Dupont, S., Farine, J. P., BEney, L., et al. (2018). Yeast quality in juvenile diet affects Drosophila melanogaster adult life traits. Sci. Rep. 8:13070. doi: 10.1038/s41598-018-31561-9
Gunther, C. S., Knight, S. J., Jones, R., and Goddard, M. R. (2019). Are Drosophila preferences for yeasts stable or contextual? Ecol. Evol. 9, 8075–8086. doi: 10.1002/ece3.5366
Hamby, K. A., Hernández, A., Boundy-Mills, K., and Zalom, F. G. (2012). Associations of yeasts with spotted wing Drosophila (Drosophila suzukii; Diptera: Drosophilidae) in cherries and raspberries. Appl. Environ. Microbiol. 78, 4869–4873.
Hoang, D., Kopp, A., and Chandler, J. A. (2015). Interactions between Drosophila and its natural yeast symbionts – Is Saccharomyces cerevisiae a good model for studying the fly–yeast relationship? PeerJ 3:e1116. doi: 10.7717/peerj.1116
Hui, F. L., Niu, Q. H., Ke, T., and Liu, Z. (2012). Candida ficus sp. nov., a novel yeast species from the gut of Apriona germari larvae. Int. J. Syst. Evol. Microbiol. 62, 2805–2809. doi: 10.1099/ijs.0.044214-0
Jimenez, S. I., Carroll, C., Babock, T., Derstine, N., Hadwin, A., Moore, M., et al. (2017). Yeasts harbored by vespine wasps in the Pacific Northwest. Environ. Entomol. 46, 217–225. doi: 10.1093/ee/nvw173
Kaltenpoth, M., and Steiger, S. (2013). Unearthing carrion beetles’ microbiome: characterization of bacterial and fungal hindgut communities across the Silphidae. Mol. Ecol. 23, 1251–1267. doi: 10.1111/mec.12469
Kurtzman, C. P., and Robnett, C. J. (1998). Three new insect-associated species of the yeast genus Candida. Can. J. Microbiol. 44, 965–973. doi: 10.1139/cjm-44-10-965
Lachance, M. A., Anderson, T. M., and Starmer, W. T. (2006). A new subclade of haplontic Metschnikowia species associated with insects of morning glory flowers in Africa and description of Metschnikowia aberdeeniae sp. nov. Int. J. Syst. Evol. Microbiol. 56, 1141–1145. doi: 10.1099/ijs.0.64195-0
Lachance, M. A., Bowles, J. M., Chavarria Diaz, M. M., and Janzen, D. H. (2001a). Candida cleridarum, Candida tilneyi and Candida powellii, three new yeast species isolated from insects associated with flowers. Int. J. Syst. Evol. Microbiol. 51, 1201–1207. doi: 10.1099/00207713-51-3-1201
Lachance, M. A., Bowles, J. M., and Starmer, W. T. (2003). Metschnikowia santaceciliae, Candida hawaiiana, and Candida kipukae, three new yeast species associated with insects of tropical morning glory. FEMS Yeast Res. 3, 97–103. doi: 10.1111/j.1567-1364.2003.tb00144.x
Lachance, M. A., Gilbert, D. G., and Starmer, W. T. (1995). Yeast communities associated with Drosophila species and related flies in an eastern oak-pine forest: a comparison with western communities. J. Ind. Microbiol. 14, 484–494. doi: 10.1007/bf01573963
Lachance, M. A., Starmer, W. T., Rosa, C. A., Bowles, J. M., Barker, J. S., and Janzen, D. H. (2001b). Biogeography of the yeasts of ephemeral flowers and their insects. FEMS Yeast Res. 1, 1–8. doi: 10.1016/s1567-1356(00)00003-9
Lam, S. S., and Howell, K. S. (2015). Drosophila-associated yeast species in vineyard ecosystems. FEMS Microbiol. Lett. 362:fnv170. doi: 10.1093/femsle/fnv170
Larsen, B. B., Miller, E. C., Rhodes, M. K., and Wiens, J. J. (2017). Inordinate fondness multiplied and redistributed: the number of species on earth and the new pie of life. Q. Rev. Biol. 92, 229–265. doi: 10.1086/693564
Lemaitre, B., and Hoffmann, J. (2007). The host defense of Drosophila melanogaster. Annu. Rev. Immunol. 25, 697–743.
Lionakis, M. S. (2011). Drosophila and Galleria insect model hosts: new tools for the study of fungal virulence, pharmacology and immunology. Virulence 2, 521–527. doi: 10.4161/viru.2.6.18520
Liti, G. (2015). The fascinating and secret wild life of the budding yeast S. cerevisiae. eLife 4:e05835. doi: 10.7554/eLife.05835
Liu, X. J., Yi, Z. H., Ren, Y. C., Li, Y., and Hui, F. L. (2016). Five novel species in the Lodderomyces clade associated with insects. Int. J. Syst. Evol. Microbiol. 66, 4881–4889. doi: 10.1099/ijsem.0.001446
Lu, H. L., and St Leger, R. J. (2016). “Insect immunity to entomopathogenic fungi” in Genetics and Molecular Biology of Entomopathogenic Fungi, eds B. Lovett R. J. S. Leger? (New York, NY: Elsevier), 251–285. doi: 10.1016/bs.adgen.2015.11.002
Meriggi, N., Di Paola, M., Vitali, F., Rivero, D., Cappa, F., Turillazzi, F., et al. (2019). Saccharomyces cerevisiae induces immune enhancing and shapes gut microbiota in social wasps. Front. Microbiol. 10:2320. doi: 10.3389/fmicb.2019.02320
Morais, B. P., Hagler, A. N., Rosa, C. A., Mendonca-Hagler, L. C., and Klaczko, L. B. (1993). Yeasts associated with Drosophila in tropical forests of Rio de Janeiro, Brazil. Can. J. Microbiol. 38, 1150–1155. doi: 10.1139/m92-188
Morais, P. B., Rosa, C. A., Hagler, A. N., and Mendonca-Hagler, L. C. (1994). Yeast communities of the cactus Pilosocereus arrabidae as resources for larval and adult stages of Drosophila serido. Antonie Van Leeuwenhoek 66, 313–317. doi: 10.1007/bf00882766
Murgier, J., Everaerts, C., Farine, J.-P., and Ferveur, J.-F. (2019). Live yeast in juvenile diet induces species-specific effects on Drosophila adult behaviour and fitness. Sci Rep. 9:8873. doi: 10.1038/s41598-019-45140-z
Nguyen, N. H., Suh, S. O., and Blackwell, M. (2007). Five novel Candida species in insect-associated yeast clades isolated from Neuroptera and other insects. Mycologia 99, 842–858. doi: 10.3852/mycologia.99.6.842
Nguyen, N. H., Suh, S. O., Erbil, C. K., and Blackwell, M. (2006). Metschnikowia noctiluminum sp. nov., Metschnikowia corniflorae sp. nov., and Candida chrysomelidarum sp. nov., isolated from green lacewings and beetles. Mycol. Res. 110, 346–356. doi: 10.1016/j.mycres.2005.11.010
Ninomiya, S., Mikata, K., Kajimura, H., and Kawasaki, H. (2013). Two novel ascomycetous yeast species, Wickerhamomyces scolytoplatypi sp. nov. and Cyberlindnera xylebori sp. nov., isolated from ambrosia beetle galleries. Int. J. Syst. Evol. Microbiol. 63, 2706–2711. doi: 10.1099/ijs.0.050195-0
Palanca, L., Gaskett, A. C., Günther, C. S., Newcomb, R. D., and Goddard, M. R. (2013). Quantifying variation in the ability of yeasts to attract Drosophila melanogaster. PLoS One 8:e75332. doi: 10.1371/journal.pone.0075332
Palma, M. L., Zamith-Miranda, D., Martins, F. S., Bozza, F. A., Nimrichter, L., Montero-Lomeli, M., et al. (2015). Probiotic Saccharomyces cerevisiae strains as biotherapeutic tools: Is there room for improvement? Appl. Microbiol. Biotechnol. 99, 6563–6570. doi: 10.1007/s00253-015-6776-x
Parapouli, M., Vasileiadis, A., Afendra, A.-S., and Hatziloukas, E. (2020). Saccharomyces cerevisiae and its industrial applications. AIMS Microbiol. 6, 1–31. doi: 10.3934/microbiol.2020001
Park, R., Dzialo, M. C., Spaepen, S., Nsabimana, D., Gielens, K., Devriese, H., et al. (2019). Microbial communities of the house fly Musca domestica vary with geographical location and habitat. Microbiome 7:147. doi: 10.1186/s40168-019-0748-9
Patt, J. M., Pfannenstiel, R. S., Meikle, W. J., and Adamczyk, J. J. (2012). Supplemental diets containing yeast, sucrose, and soy powder enhance the survivorship, growth, and development of prey-limited cursorial spiders. Biol. Control. 63, 237–245. doi: 10.1016/j.biocontrol.2011.02.004
Phaff, H. J., and Knapp, E. P. (1956). The taxonomy of yeast found in exudates of certain trees and other natural breeding sites of some species of Drosophila. Antonie van Leeuwenhoek 22, 117–130. doi: 10.1007/bf02538319
Piper, A. M., Farnier, K., Linder, T., Speight, R., and Cunningham, J. P. (2017). Two gut-associated yeasts in a Tephritid fruit fly have contrasting effects on adult attraction and larval survival. J. Chem. Ecol. 43, 891–901. doi: 10.1007/s10886-017-0877-1
Piper, M. D. W. (2017). Using artificial diets to understand the nutritional physiology of Drosophila melanogaster. Curr. Opin. Insect Sci. 23, 104–111. doi: 10.1016/j.cois.2017.07.014
Quan, A. S., and Eisen, M. B. (2018). The ecology of the Drosophila-yeast mutualism in wineries. PLoS One 13:e0196440. doi: 10.1371/journal.pone.0196440
Ramazzotti, M., Stefanini, I., Di Paola, M., De Filippo, C., Rizzetto, L., Berná, L., et al. (2019). Population genomics reveals evolution and variation of Saccharomyces cerevisiae in the human and insects gut. Environ. Microbiol. 22, 50–71. doi: 10.1111/1462-2920.14422
Ravenscraft, A., Berry, B., Hammer, T., Peay, K., and Boggs, C. (2018). Structure and function of the bacterial and fungal gut microbiota of neotropical butterflies. Ecol. Monogr. 89:e01346. doi: 10.1002/ecm.1346
Ren, Y. C., Liu, S. T., Li, Y., and Hui, F. L. (2015). Pichia dushanensis sp. nov. and Hyphopichia paragotoi sp. nov., two sexual yeast species associated with insects and rotten wood. Int. J. Syst. Evol. Microbiol. 65, 2875–2881. doi: 10.1099/ijs.0.000349
Ren, Y. C., Wang, Y., Chen, L., Ke, T., and Hui, F. L. (2014). Wickerhamiella allomyrinae fa, sp. nov., a yeast species isolated from the gut of the rhinoceros beetle Allomyrina dichotoma. Int. J. Syst. Evol. Microbiol. 64, 3856–3861. doi: 10.1099/ijs.0.068403-0
Resnick, M. A., and Cox, B. S. (2000). Yeast as an honorary mammal. Mutat. Res. 451, 1–11. doi: 10.1016/s0027-5107(00)00036-1
Reuter, M., Bell, G., and Greig, D. (2007). Increased outbreeding in yeast in response to dispersal by an insect vector. Curr. Biol. 17, R81–R83.
Rivera, F. N., Gonzalez, E., Gomez, Z., Lopez, N., Hernandez-Rodriguez, C., Berkov, A., et al. (2009). Gut-associated yeast in bark beetles of the genus Dendroctonus Erichson (Coleoptera: Curculionidae: Scolytinae). Biol. J. Linn. Soc. 98, 325–391.
Roh, K. B., Kim, C. H., Lee, H., Kwon, H. M., Park, J. W., Ryu, J. H., et al. (2009). Proteolytic cascade for the activation of the insect toll pathway induced by the fungal cell wall component. J. Biol. Chem. 284, 19474–19481. doi: 10.1074/jbc.m109.007419
Rosa, C. A., Lachance, M. A., Silva, J. O. C., Teixeira, A. C. P., Marini, M. M., Antonini, Y., et al. (2003). Yeast communities associated with stingless bees. FEMS Yeast Res. 4, 271–275. doi: 10.1016/S1567-1356(03)00173-9
Rosa, C. A., Lachance, M. A., Teixeira, L. C., Pimenta, R. S., and Morais, P. B. (2007). Metschnikowia cerradonensis sp. nov., a yeast species isolated from ephemeral flowers and their nitidulid beetles in Brazil. Int. J. Syst. Evol. Microbiol. 57, 161–165. doi: 10.1099/ijs.0.64624-0
Rosa, C. A., Morais, P. B., Hagler, A. N., Mendonsa-Hagler, L. C., and Monteiro, R. F. (1994). Yeast communities of the cactus Pilosocereus urruhidae and associated insects in the sandy coastal plains of southeastern Brazil. Antonie Van Leeuwenhoek 65, 55–62. doi: 10.1007/bf00878279
Saksinchai, S., Suzuki, M., Chantawannakul, P., Ohkuma, M., and Lumyong, S. (2015). A novel ascosporogenous yeast species, Zygosaccharomyces siamensis, and the sugar tolerant yeasts associated with raw honey collected in Thailand. Fungal Divers. 52, 123–139. doi: 10.1007/s13225-011-0115-z
Sandhu, D. K., and Waraich, M. K. (1985). Yeasts associated with pollinating bees and flower nectar. Microb. Ecol. 11, 51–58. doi: 10.1007/bf02015108
Schiabor, K. M., Quan, A. S., and Eisen, M. B. (2014). Saccharomyces cerevisiae mitochondria are required for optimal attractiveness to Drosophila melanogaster. PLoS One 9:e113899. doi: 10.1371/journal.pone.0113899
Siavoshi, F., Sahraee, M., Ebrahimi, H., Sarrafnejad, A., and Saniee, P. (2018). Natural fruits, flowers, honey, and honeybees harbor Helicobacter pylori-positive yeasts. Helicobacter 23:e12471. doi: 10.1111/hel.12471
Six, D. L. (2003). A comparison of mycangial and phoretic fungi of individual mountain pine beetles. Can. J. For. Res. 33, 1331–1334. doi: 10.1139/x03-047
Stefanini, I. (2018). Yeast-insect associations: it takes guts. Yeast 35, 315–330. doi: 10.1002/yea.3309
Stefanini, I., Dapporto, L., Berná, L., Polsinelli, M., Turillazzi, S., and Cavalieri, D. (2016). Social wasps are a Saccharomyces mating nest. Proc. Natl. Acad. Sci. U.S.A. 113, 2247–2251. doi: 10.1073/pnas.1516453113
Stefanini, I., Dapporto, L., Legras, J. L., Calabretta, A., Di Paola, M., De Filippo, C., et al. (2012). Role of social wasps in Saccharomyces cerevisiae ecology and evolution. Proc. Natl. Acad. Sci. U.S.A. 109, 13398–13403. doi: 10.1073/pnas.1208362109
Steyn, A., Roets, F., and Botha, A. (2016). Yeasts associated with Culex pipiens and Culex theileri mosquito larvae and the effect of selected yeast strains on the ontogeny of Culex pipiens. Microb. Ecol. 71, 747–760. doi: 10.1007/s00248-015-0709-1
Suh, S. O., and Blackwell, M. (2004). Three new beetle-associated yeast species in the Pichia Guilliermondii clade. FEMS Yeast Res. 5, 87–95. doi: 10.1016/j.femsyr.2004.06.001
Suh, S. O., and Blackwell, M. (2006). Three new asexual arthroconidial yeasts, Geotrichum carabidarum sp. nov., Geotrichum histeridarum sp. nov., and Geotrichum cucujoidarum sp. nov., isolated from the gut of insects. Mycol. Res. 110, 220–228. doi: 10.1016/j.mycres.2005.08.008
Suh, S. O., Houseknecht, J. L., Gujjari, P., and Zhou, J. J. (2013). Scheffersomyces parashehatae fa, sp. nov., Scheffersomyces xylosifermentans fa, sp. nov., Candida broadrunensis sp. nov. and Candida manassasensis sp. nov., novel yeasts associated with wood-ingesting insects, and their ecological and biofuel implications. Int. J. Syst. Evol. Microbiol. 63, 4330–4339. doi: 10.1099/ijs.0.053009-0
Suh, S. O., Nguyen, N. H., and Blackwell, M. (2005). Nine new Candida species near C. membranifaciens isolated from insects. Mycol. Res. 109, 1045–1056. doi: 10.1017/S0953756205003254
Suh, S. O., Nguyen, N. H., and Blackwell, M. (2006). A yeast clade near Candida kruisii uncovered: nine novel Candida species associated with basidioma-feeding beetles. Mycol. Res. 110, 1379–1394. doi: 10.1016/j.mycres.2006.09.009
Suh, S. O., Nguyen, N. H., and Blackwell, M. (2007). Yeasts isolated from plant-associated beetles and other insects: seven novel Candida species near Candida albicans. FEMS Yeast Res. 8, 88–102. doi: 10.1111/j.1567-1364.2007.00320.x
Tanahashi, M., and Hawes, C. J. (2016). The presence of a mycangium in European Sinodendron cylindricum (Coleoptera: Lucanidae) and the associated yeast symbionts. J. Insect. Sci. 16:76. doi: 10.1093/jisesa/iew054
Tatum, E. L. (1941). Vitamin B requirements of Drosophila melanogaster. Proc. Natl. Acad. Sci. U.S.A. 27, 193–197. doi: 10.1073/pnas.27.4.193
Tietjen, E. J., Ayyagari, L. R., and Uetz, G. W. (1987). Symbiosis between social spiders and yeast: the role in prey attraction. Psyche A J. Entomol. 94, 151–158. doi: 10.1155/1987/67258
Toki, W., Tanahashi, M., Togashi, K., and Fukatsu, T. (2012). Fungal farming in a non-social beetle. PLoS One 7:e41893. doi: 10.1371/journal.pone.0041893
Urbina, H., Schuster, J., and Blackwell, M. (2013). The gut of Guatemalan passalid beetles: a habitat colonized by cellobiose-and xylose-fermenting yeasts. Fungal Ecol. 6, 339–355. doi: 10.1016/j.funeco.2013.06.005
Wagner, D. L. (2020). Insect declines in the Anthropocene. Annu. Rev. Entomol. 65, 457–480. doi: 10.1146/annurev-ento-011019-025151
Wang, Y., Ren, Y. C., Zhang, Z. T., Ke, T., and Hui, F. L. (2016). Spathaspora allomyrinae sp. nov., a D-xylose-fermenting yeast species isolated from a scarabeid beetle Allomyrina dichotoma. Int. J. Syst. Evol. Microbiol. 66, 2008–2012. doi: 10.1099/ijsem.0.000979
Wilson, E. O. (1987). The little things that run the World (The importance and conservation of invertebrates). Conserv. Biol. 1, 344–346. doi: 10.1111/j.1523-1739.1987.tb00055.x
Keywords: Saccharomyces cerevisiae, insect, yeast-insect association, biogeography, Saccharomyces cerevisiae evolution, Saccharomyces cerevisiae yeast ecology
Citation: Meriggi N, Di Paola M, Cavalieri D and Stefanini I (2020) Saccharomyces cerevisiae – Insects Association: Impacts, Biogeography, and Extent. Front. Microbiol. 11:1629. doi: 10.3389/fmicb.2020.01629
Received: 03 May 2020; Accepted: 22 June 2020;
Published: 14 July 2020.
Edited by:
Nathalie Connil, Université de Rouen, FranceReviewed by:
Emilia Garcia-Moruno, Council for Agricultural and Economics Research (CREA), ItalyAgustin Aranda, University of Valencia, Spain
Flaviano S. Martins, Federal University of Minas Gerais, Brazil
Copyright © 2020 Meriggi, Di Paola, Cavalieri and Stefanini. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Irene Stefanini, irene.stefanini@unito.it