 Wu Qu
Wu Qu Tan Liu2
Tan Liu2 Jing Zhao
Jing Zhao- 1School of Life Sciences, Xiamen University, Xiamen, China
- 2College of Ocean and Earth Sciences, Xiamen University, Xiamen, China
- 3The Department of Laboratory Medicine, The First Affiliated Hospital of Xiamen University, Xiamen, China
Malachite green (MG) is an organic contaminant and the effluents with MG negatively influence the health and balance of the coastal and marine ecosystem. The diverse and abundant microbial communities inhabiting in mangroves participate actively in various ecological processes. Metagenomic sequencing from mangrove sediments was applied to excavate the resources MG-degradation genes (MDGs) and to assess the potential of their corresponding enzymes. A data set of 10 GB was assembled into 33,756 contigs and 44,743 ORFs were predicted. In the data set, 666 bacterial genera and 13 pollutant degradation pathways were found. Proteobacteria and Actinobacteria were the most dominate phyla in taxonomic assignment. A total of 44 putative MDGs were revealed and possibly derived from 30 bacterial genera, most of which belonged to the phyla of Proteobacteria and Bacteroidetes. The MDGs belonged to three gene families, including peroxidase genes (up to 93.54% of total MDGs), laccase (3.40%), and p450 (3.06%). Of the three gene families, three representatives (Mgv-rLACC, Mgv-rPOD, and Mgv-rCYP) which had lower similarities to the closest sequences in GenBank were prokaryotic expressed and their enzymes were characterized. Three recombinant proteins showed different MG-degrading activities. Mgv-rPOD had the strongest activity which decolorized 97.3% of MG (300 mg/L) within 40 min. In addition, Mgv-rPOD showed a more complete process of MG degradation compared with other two recombinant proteins according to the intermediates detected by LC-MS. Furthermore, the high MG-degrading activity was maintained at low temperature (20°C), wider pH range, and the existence of metal ions and chelating agent. Mgv-rLACC and Mgv-rCYP also removed 63.7% and 54.1% of MG (20 mg/L) within 24 h, respectively. The results could provide a broad insight into discovering abundant genetic resources and an effective strategy to access the eco-friendly way for preventing coastal pollution.
Introduction
The application of synthetic dyes is extensive in the fields of textile, pharmaceutical, cosmetics, paper making, solar cells, and acrylic industries (Chang and Lin, 2001). Due to various risks on environment and human health, strategies for the removal of dyes have attracted increasing attention from health professionals and environmentalists (Srivastava et al., 2004; Venil and Lakshmanaperumalsamy, 2010; Gopinathan et al., 2015). Malachite green (MG), in particular, is one typical of synthetic dyes and is widely used in dyeing of silk, leather, and paper, which has been shown to harm humans and animals because of toxicity, mutagenicity, and carcinogenicity (Srivastava et al., 2004; Gopinathan et al., 2015). MG is a triphenylmethane dye and highly soluble in water (Yong et al., 2015). MG can be metabolized into leucomalachite green (LMG) (Yong et al., 2015) and remains in fish muscles, fat, and organs with a half-life of about 10 days and even longer (Chen and Miao, 2010). MG is also persistent in the environment and its half-life in sediment can range from 12.9 to 50.34 days (Jiang, 2011). Despite the global ban on MG in aquaculture, 1–15% of dyes are estimated being discharged within effluents, which seriously inhibit survival, development, and reproductive of aquatic organisms (Chang et al., 2016). The removal or reduction of MG can be processed by chemicals methods or biological treatments. Chemicals methods, such as photodegradation (Algubury, 2016; Pathania et al., 2016; Liang et al., 2017), have some disadvantages of high costs and secondary pollutant generation (Tayabali et al., 2017). Bioremediation strategy may constitute an alternative approach to conventional physicochemical methods, benefiting from the potential of indigenous microorganisms to metabolize anthropogenic compounds (Grosser et al., 1991; Tayabali et al., 2017).
Mangrove ecosystems constitute 60–70% of the coastline in the tropical and subtropical regions on Earth, which receive nutrient-rich aquaculture effluent from nearby farming activities and accumulated organic contaminnat from industrial wastewater discharge (Gomes Gomes et al., 2008; Giri et al., 2011). The highly productive and diverse microbial community living in tropical and subtropical mangrove ecosystems continuously transforms varied nutrients into sources of nitrogen, phosphorus, and other nutrients that can be used by the plants (Ouyang et al., 2017; Yang et al., 2017). Due the ability to absorb waste and pollutants, mangroves are considered as significant sinks for pollutants from freshwater discharges as well as from contaminated tidal water (Marchand et al., 2016). Therefore, mangrove sediments are suitable for exploring MG degrading microorganisms because of input of carbon in the form of litter which then acts as a substrate for decomposition by microbe. The bioremediation potential of microorganisms isolated from hydrocarbon-contaminated environments was as active as or even higher than those originating from non-contaminated sediments (Jones et al., 2011). For now, a few culturable microorganisms with the degradation activity of dyestuff pollutant are isolated from mangrove sediment microbiome. Two strains, P1 and D1, isolated from mangrove sediments previously, were found with the decolorization ability against a variety of dyes, including nitomill brill crimson, methyl red, and nitro green B (Srinivasan et al., 2014). A mangrove-derived strain, Aplanochytrium sp., was reported with MG degrading activity which could remove 86.32% of MG within 5.5 days (Gomathi et al., 2013). Despite global advancement in understanding the microbial diversity in mangrove sediments, more than 90% of environmental microorganisms remain unculturable (Amann et al., 1990; Schloss and Handelsman, 2005; Kimura, 2006). MG degradation potential could be difficult to be assessed and utilized by using traditional culture-dependent method.
Nowadays, culture-independent metagenomic library has successfully used for discovering of novel biosynthetic gene from diverse environments. Lac15, a laccase isolated from marine bacterial metagenome, could degrade several industrial dyes belonged to reactive azo class under alkalescent conditions (Fang et al., 2011). Besides, a novel bacterial laccase Lac21 was isolated from metagenomic library of the South China Sea. Lac21 could remove 80% of Reactive Deep Blue M-2GE (50 mg/L) within 24 h (Fang et al., 2012). However, the reliance on relatively low-throughput of clone libraries combined with activity-based screening could limit screening output (Pope and Moran, 2010; Hess et al., 2011). In addition, metagenomic library focuses on the individual functional gene, not conveniently provides the capacity and diversity of the functional gene in a certain environmental sample. Sequence-based screening of metagenomics combined with the databases, such as Kyoto Encyclopedia of Genes and Genomes (KEGG), Cluster of Orthologous Groups of Proteins (COG), etc., provides a high-performance method on screening the sequences and abundance of potential functional genes (Hugenholtz and Tyson, 2008; Simon and Daniel, 2009). For now, several MDGs have been isolated from MG-degrading strains and metagenomics libraries, including laccase (LACC) (Murugesan et al., 2009), peroxidase (POD) (Ulson de Souza et al., 2007), cytochrome p450 (CYP) (Jefferson and Jones, 2003), triphenylmethane reductase (TMD) (Kim et al., 2008) and triphenylmethane dye oxidase (TpmD) (Ren et al., 2006). However, MDGs resources and abilities from different environment, such as mangrove sediment, were still cryptical. Moreover, the enzymatic properties of MDGs from varied gene families, such as degrading activity, stability against complex conditions, and more radical degradation products, are still strongly desired.
In this study, the culture independent metagenomic method was applied in understanding bacterial abundance and diversity, assessing the gene families, and revealing the novel MDGs genes of the mangrove sediment along South China Sea. Based on the metabolism pathway analysis, some potential MDGs from different gene families were selected and prokaryotic expressed. The biochemical characterization of the corresponding enzymes of MDGs, including temperature, pH, metal ions, metal-chelator, salinity, ionic detergent, and degradation pathway, were tested to further analyze their adaptability and potential to serve as tool enzymes in MG bioremediation.
Materials and Methods
Sampling and Environment DNA (eDNA) Extraction
The sediment samples were collected from mangrove forest located in Longhai City, Fujian, China (Zini Mangrove Nature Reserve, 24°20′N, 117°45′E). About 50 g of sediments were collected in a 50 mL of sterile centrifugal tube and stored on ice. Stored the sediments samples in ultra-low temperature freezer (-80°C) after they were brought back to lab.
eDNA was extracted using the chemical lysis and enzyme digestion method. It was purified with low melting-point agarose [Sangon Biotech (Shanghai) Co., Ltd., China] and agarase (Takara Biotechnology Co., Ltd., Japan). Three replications of extraction and purification were performed and pooled together to avoid the extraction biases.
Sequencing and Bioinformatics Analysis of Metagenome
eDNA sample was used for shotgun paired-end library construction. DNA sample was break into the fragments with the length of about 400 bp using ultrasonic breakers (Covaris, United States). End-repair, adaptor jointing, and purification of the DNA fragments were performed to construct the paired-end library. The concentration of the library was measured by Qubit 2.0 Fluorometer (Invitrogen, United States). High-throughput sequencing using Illumina HiSeq 2500 (Illumina Inc., United States) was performed by Shanghai Majorbio Bio-Pharm Technology Co. (China). Approximately 10 GB of data were generated and the raw reads containing “N” or adaptors were removed from the dataset and the clean reads were retaining for further analysis. The raw data of Illumina sequencing has been deposited in Sequence Read Archive (SRA) database1 under the accession numbers SRR5824292.
All clean reads were assembled using SOAPdenovo with Kmer of 43–47. The best Kmer was identified on the basis of contig numbers, contig N50, contig length, etc. MetaGeneMark was used to predict the open reading frames (ORFs) based on the contigs and singletons obtained. ORFs were aligned using BLAST+ in Nr database of NCBI (cut-off E-value of 1e-5) and were annotated with the functional information. The putative MDGs mentioned before were filtered out according to the gene annotation. The bacteria origin of putative MDGs was predicted based on sequence homology with the protein sequences in NCBI Nr database using BLAST+ (cut-off E-value of 1e-5). COGs functional classification was conducted in STRING database2 and KEGG3 was used to obtain the biological pathways with a BLAST algorithm in KEGG database. All reads were aligned in SSU rRNA database of SILVA (cut-off E-value of 1e-5) to investigate taxonomic assignment of sampling site.
Full-Length Amplification of mgv-laccase and mgv-p450 Genes
Due to relatively lower similarities (about 50% similarities) with known sequences compared with other putative MDGs, three representative MDGs belonging to POD, LACC, and CYP, respectively, were chosen to be expressed in E. coli BL21 (DE3) cells and their MG-degrading activities were further detected. Three genes were labeled as mgv-peroxidase, mgv-laccase, and mgv-p450, respectively. Among them, mgv-laccase and mgv-p450, which had not complete ORFs, were amplified by TAIL-PCR with Genome Walking Kit (Takara Biotechnology Co., Ltd., Japan) according to the instructions. In brief, eDNA solution was used as the template for TAIL-PCR. Thermal asymmetric cycle was performed for three times, and the programs and reagent doses were mixed strictly according to the instructions. The degenerate primers for thermal asymmetric amplification were provided in the kit, and the specific primers (Table 1) for mgv-laccase and mgv-p450 were synthesized by Invitrogen (Shanghai) Co., Ltd. (United States). The possible DNA fragments were retrieved with TIANgel Midi Purification Kit (Tiangen Biotech Co., Ltd., China) after 1% agarose gel electrophoresis, and were cloned into pMD19-T Vector (Takara Biotechnology Co., Ltd., Japan). Then, DNA sequencing was conducted by using 3730xl DNA Analyzer (Thermo Fisher Scientific Co., Ltd., United States). Sequences were assembled with DNAman (Version 6.0.3.99) and ORFs of mgv-laccase and mgv-p450 were predicted in ORF Finder4.
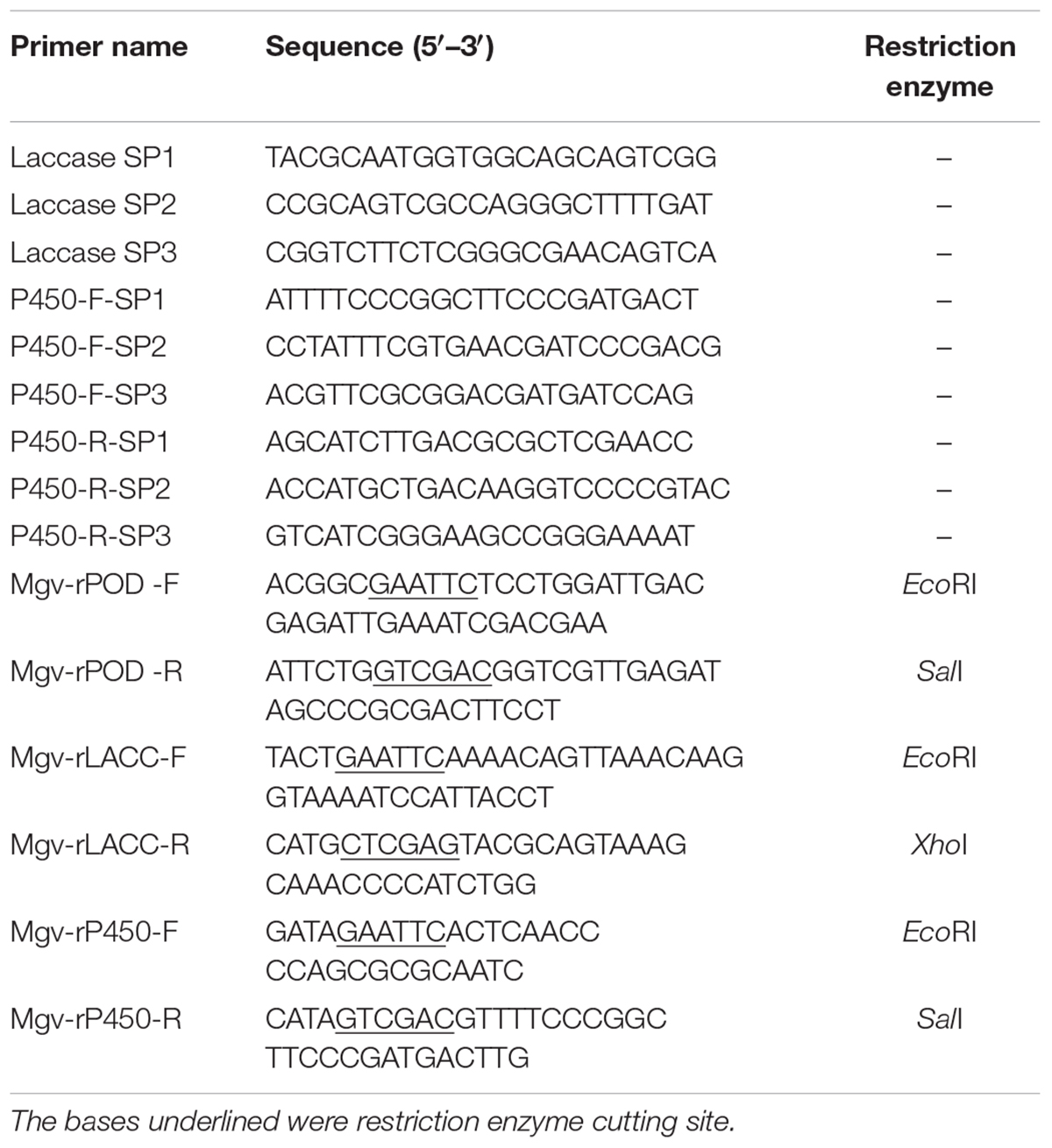
TABLE 1. Primers for TAIL-PCR and expression of malachite green-degrading genes (MDGs).
Expression and Purification of MDGs
Three putative MDGs, mgv-laccase, mgv-p450, and mgv-peroxidase, were cloned by primers (Table 1) designed according to ORFs. The PCR product was ligated into pET-32a vector (Novagen Inc., United States) and transformed into E. coli BL21 (DE3) cells. Recombination proteins were purified through nickel-affinity chromatography column. The three recombinant proteins were named Mgv-rPOD, Mgv-rLACC, and Mgv-rCYP, respectively. Sequences of mgv-laccase, mgv-p450, and mgv-peroxidase have been deposited in GenBank database under the accession numbers MF461728, MF461729, and MF461730, respectively.
Biochemical Analysis of MG-Degrading Activity of Recombinant Proteins
The MG-degrading activity of recombinant proteins was measured by the decolorization efficiency of MG with the formulae as follows: Decolorization efficiency (%) = (A - B)/A × 100, where A is the initial absorbance of MG and B is the final absorbance of MG at a wavelength of 622 nm. To value the cost of MG degradation, same quality (100 μg) of the three proteins were used in decolorization experiment.
The effect of temperature on the decolorizing efficiency was determined by measuring the decolorization percentage of MG at 10°C, 20°C, 30°C, 40°C, 50°C, 60°C, and 70°C, respectively. Similarly, the effect of pH was determined in the buffer solution of citric acid/sodium citrate (pH 3.0–6.0), phosphate (pH 6.0–8.0), Tris-HCl (pH 8.0–10.0). The effect of metal ions was determined with ZnSO4, MgSO4, MnSO4, NiSO4, FeSO4, CuSO4, Al2(SO4)3, and CaSO4 (1 mM). The effect of the potential inhibitors was determined with NaCl (1, 10, and 100 mM), EDTA (1, 10, and 100 mM) and SDS (0.1, 0.5, and 1%).
Determination of MG Biodegradation Intermediates
The method for the determination of MG biodegradation intermediates referred to the previous studies (Du et al., 2011, 2013; Wang et al., 2012; Yang et al., 2015). To determine the MG biodegradation intermediates, 20 mg/L MG was mixed with the three recombination proteins and incubated at 30°C for 12 h. The products were desalted with C18 solid phase extraction column (Waters Sep-pak, United States). All the samples were analyzed using LC-MS (UPLC-Tof MS System, Waters, United States). The mobile phase contained H2O and acetonitrile. The initial proportion of acetonitrile was 5% and reached 100% within 15 min. The injection volume was 10 μl, and other parameters were set at the default settings.
Results
Metagenomic Analysis
Approximately 10 GB of data was generated and 9.4 GB was remained as clean data. Subsequently, 33,756 contigs were assembled with best Kmer of 45 (Supplementary Table S1) and 44,743 ORFs were predicted. Based on SSU rRNA genes from metagenomic sequencing, microbial community showed great diversity on phylum level, mainly including Proteobacteria (50.19%) and Actinobacteria (17.03%), followed by other minor groups (Figure 1A). On genus level, a total of 666 bacterial genera were found in our datasets. Due to the lack of adequate contig length, 70.57% genus of total microbes remained unknown (Figure 1B). Twenty-three COG categories (Supplementary Figure S3) and 258 pathways in KEGG (Supplementary Table S2) were found. Thirteen metabolic pathways of common pollutant, such as styrene, ethylbenzene, aromatic compounds, chlorinated cyclohexane, and benzene degradation, were also detected in KEGG analysis.
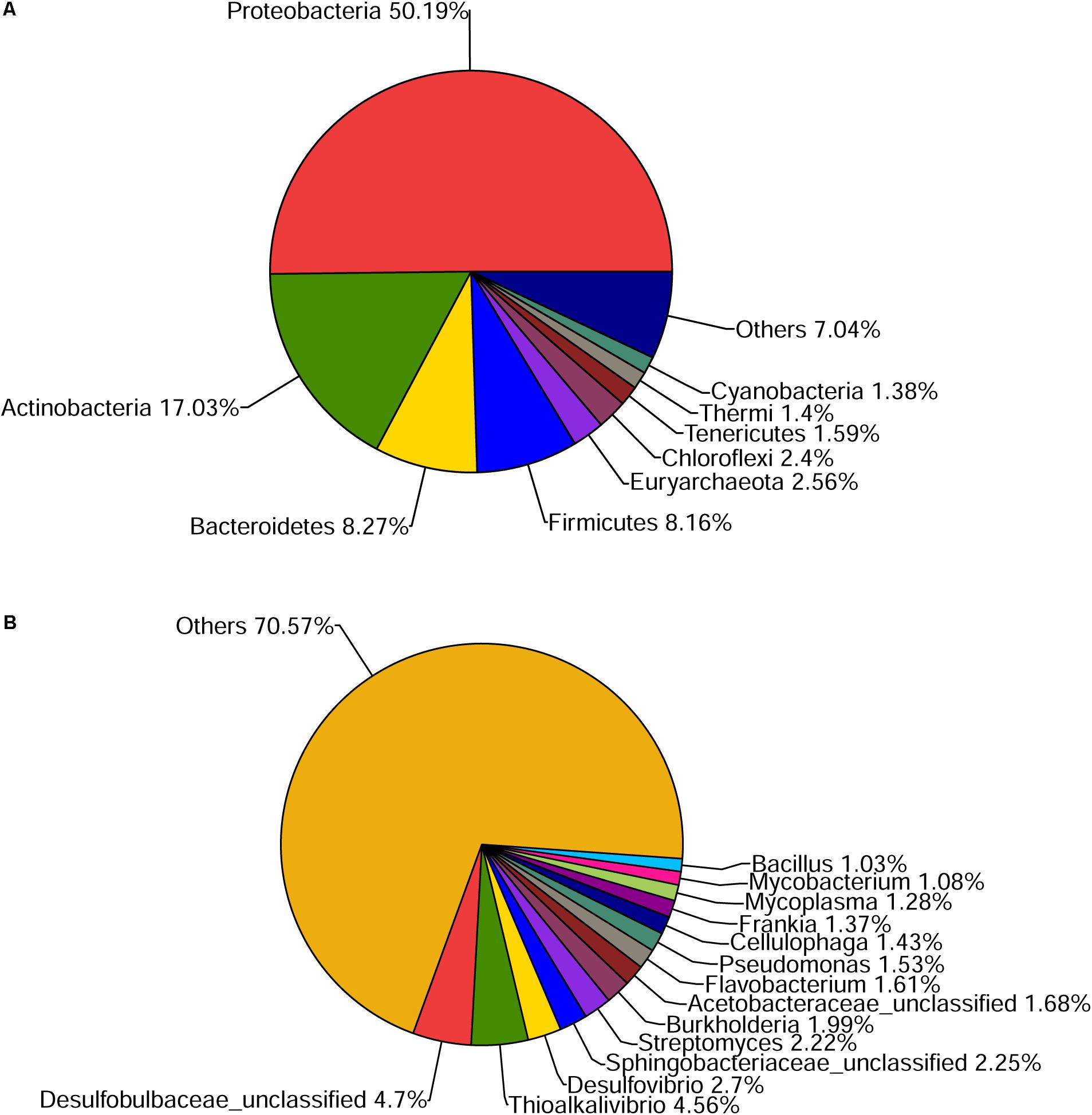
FIGURE 1. Community composition of mangrove sediment microbiome on phylum (A) and genus level (B) revealed by the SSU rRNA genes in the dataset.
Scanning, Cloning, and Expression of Putative MDGs
Forty-four putative MDGs were found in 33,756 contigs with 55–75% identities in Nr database and belonged to three gene families (Supplementary Table S4 and Figure 2A). Peroxidase genes possessed dominant proportion (up to 93.54% of 44 putative MDGs), followed by LACC (3.40%) and CYP (3.06%) (Figure 2B). Of them, seven genes were unclassified because of their unavailable TaxIDs and the remaining 37 genes were assigned to 30 bacterial genera (Figure 2C). Combined with the analysis of microbial communities, the corresponding functional genes related to the pollutant degradation and their possible bacterial origin were listed in Supplementary Table S5. Amongst these microorganisms, most (18/30) genera belonged to Proteobacteria, followed by Bacteroidetes (8/30), Actinobacteria (1/30), Spirochaetes (1/30), Thaumarchaeota (1/30), and Cyanobacteria (1/30) (Supplementary Table S5).
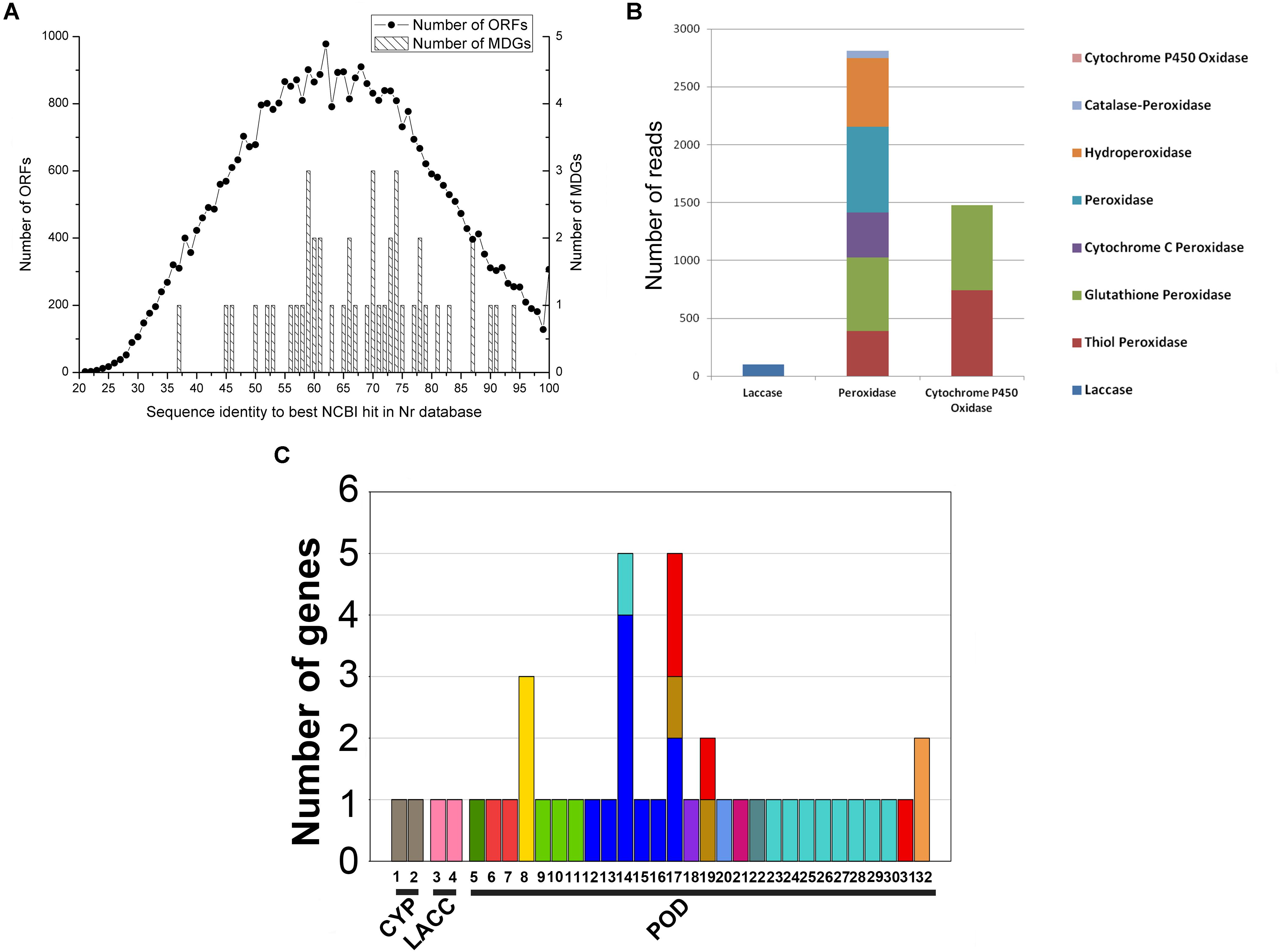
FIGURE 2. Bioinformatics analysis of putative malachite green-degrading genes (MDGs). (A) Distribution of best identity in Nr database of all open reading frames (ORFs) and putative MDGs. (B) The number of 44 putative MDGs in the dataset of mangrove sediment microbiome. (C) The bacteria origin of putative MDGs. The numbers on the X-axis indicated the genera of Nitrososphaera, Acidovorax, Desulfuromonas, Chromobacterium, Geopsychrobacter, Cellvibrio, Shewanella, Simiduia, Oscillatoria, Thiothrix, Thiocapsa, Draconibacterium, Frankia, Nitrosomonas, Sedimenticola, Bizionia, NO-TaxID, Formosa, norank, Leucothrix, Magnetococcus, Spirochaeta, Rhodanobacter, Gemmobacter, Kangiella, Gaetbulibacter, Mariniradius, Zooshikilla, Imtechella, Salinibacter, Gelidibacter, and Marinobacter, respectively.
Phylogenetic tree based on the amino acid sequences showed that mgv-laccase clustered within LACC of Pelobacter seleniigenes (Supplementary Figure S1A), mgv-p450 gathered with CYP of Salinisphaera shabanensis (Supplementary Figure S1B) and mgv-peroxidase was close to POD from Salinibacter ruber (Supplementary Figure S1C).
Reasonable ORFs were found in both mgv-laccase and mgv-p450 by TAIL-PCR and sequencing. The length of complete mgv-laccase gene and mgv-p450 gene was 795 bp and 627 bp, respectively. Mgv-rPOD, Mgv-rLACC and Mgv-rCYP were purified with molecular weight of 40 kDa (Figure 3A), 54 kDa (Figure 4A), and 43 kDa (Figure 5A), respectively.
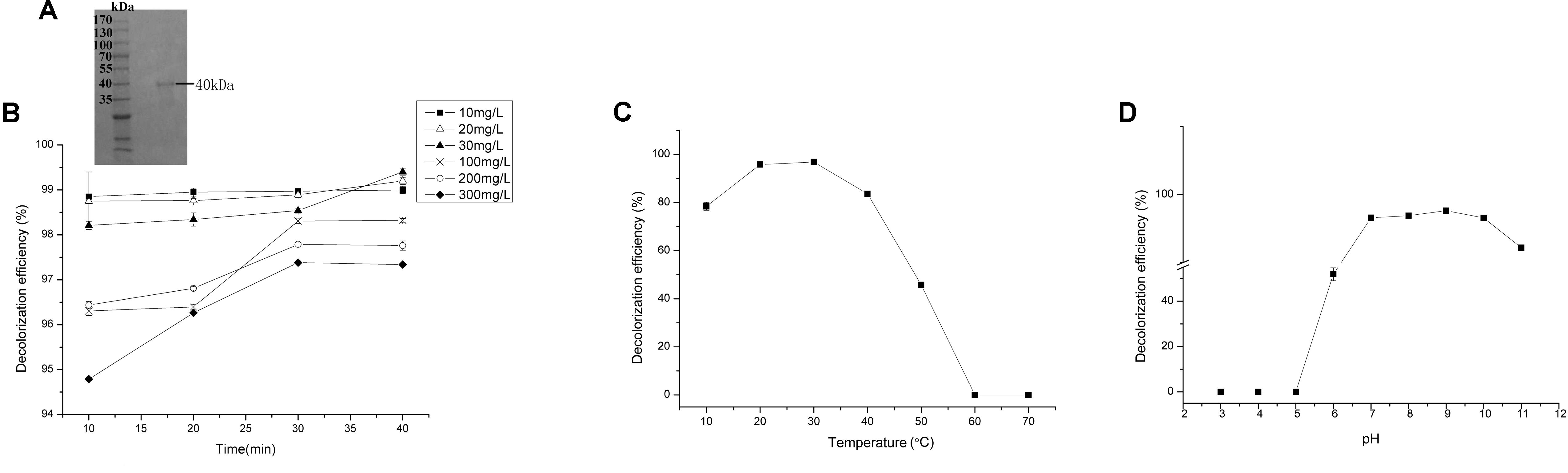
FIGURE 3. Malachite green (MG) degradation characteristics of Mgv-rPOD. (A) SDS-PAGE result of Mgv-rPOD. (B) The effect of MG concentration on the MG decolorization efficiency of Mgv-rPOD. (C) The effect of temperature on the MG decolorization efficiency of Mgv-rPOD. (D) The effect of pH on the MG decolorization efficiency of Mgv-rPOD.
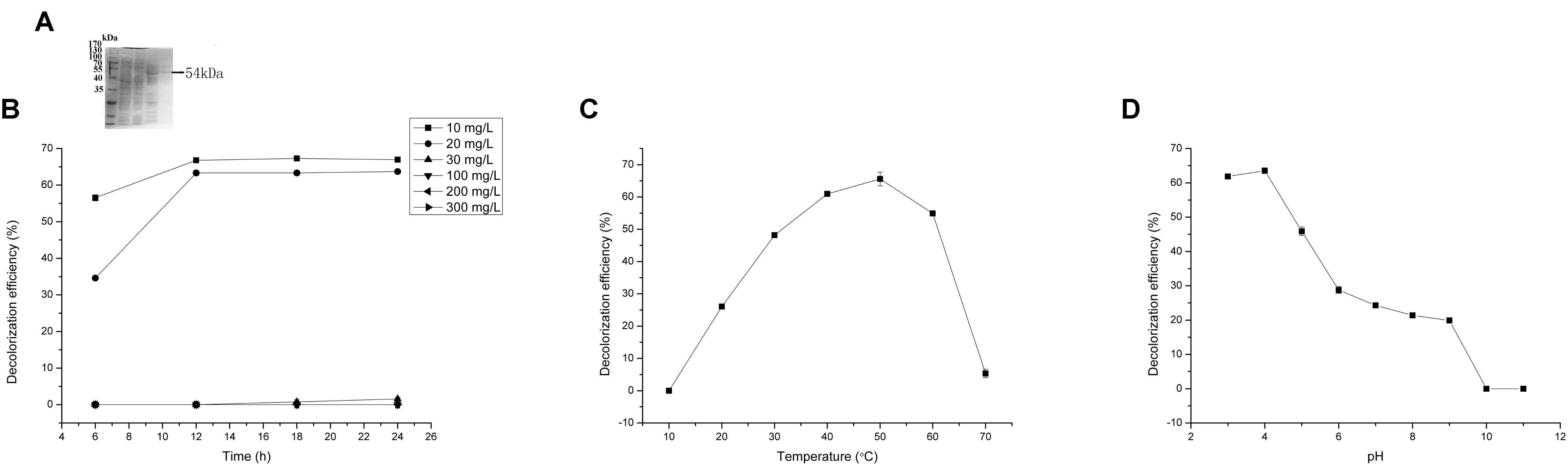
FIGURE 4. Malachite green (MG) degradation characteristics of Mgv-rLACC. (A) SDS-PAGE result of Mgv-rLACC. (B) The effect of MG concentration on the MG decolorization efficiency of Mgv-rLACC. (C) The effect of temperature on the MG decolorization efficiency of Mgv-rLACC. (D) The effect of pH on the MG decolorization efficiency of Mgv-rLACC.
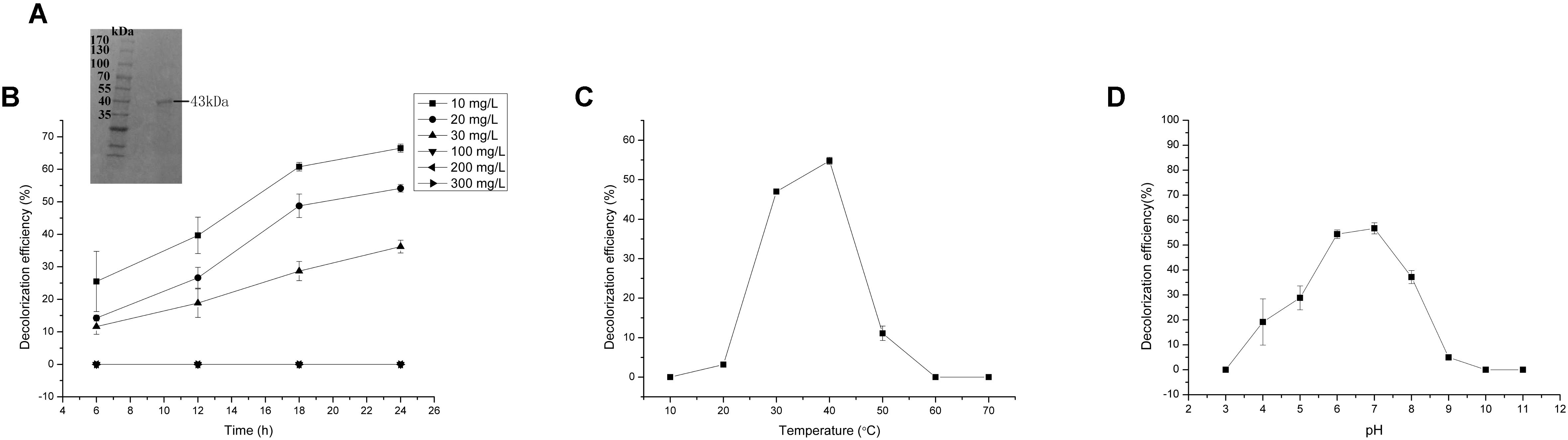
FIGURE 5. Malachite green (MG) degradation characteristics of Mgv-rCYP. (A) SDS-PAGE result of Mgv-rCYP. (B) The effect of MG concentration on the MG decolorization efficiency of Mgv-rCYP. (C) The effect of temperature on the MG decolorization efficiency of Mgv-rCYP. (D) The effect of pH on the MG decolorization efficiency of Mgv-rCYP.
MG-Degrading Characteristics Analysis
Mgv-rPOD showed strong MG-degrading activity. After incubated with Mgv-rPOD for only 40 min, the MG decolorization efficiency could reach 99.0%, 99.2%, 99.4%, 96.3%, 97.8%, and 97.3% in 10 mg/L, 20 mg/L, 30 mg/L, 100 mg/L, 200 mg/L, and 300 mg/L of MG, respectively (Figure 3B). The Mgv-rPOD had the optimum temperature 30°C for color removal with 96.9% of MG-decolorizing efficiency and still maintained the high MG-degrading activity at 20°C (Figure 3C). Also, experimental results showed that high MG decolorizing activity was often between 7.0 and 10.0 and the observed optimum pH for the reaction was 9.0 (Figure 3D). Cu2+ had obviously negative effect, which caused the loss of 65.9% MG-degrading activity of Mgv-rPOD. Meanwhile, lots of bubbles were generated in the reaction with Cu2+. Besides, the use of other metal ions did not show much variation in the activity of decolorizing MG (Figure 6A). Different concentrations of EDTA (1, 10, and 100 mM) and NaCl (1, 10, and 100 mM) could not affect the decolorizing efficiency. However, there was a sharp decrease with 0.1% SDS and an absence of MG-degrading activity with 1% SDS (Figure 6B).
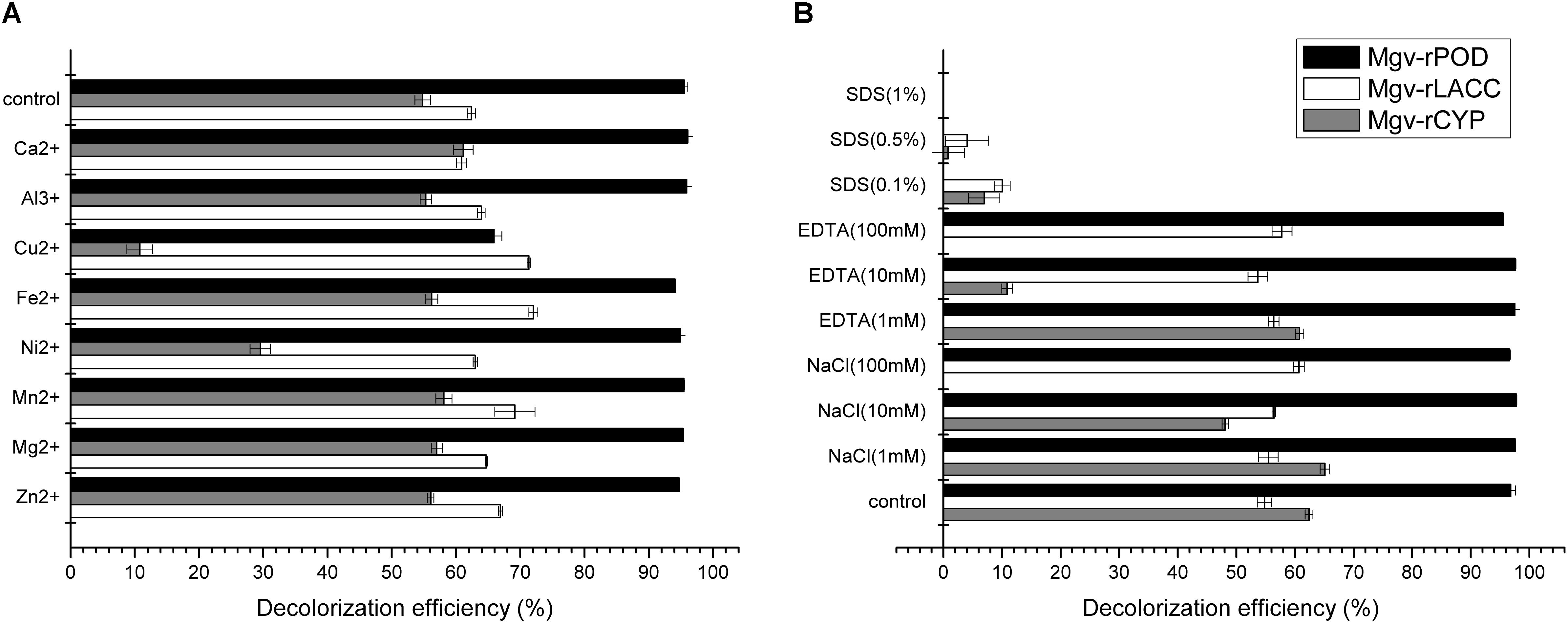
FIGURE 6. The effect of metal ions (A) and potential inhibitors (B) on the MG decolorization efficiency of Mgv-rPOD, Mgv-rLACC, and Mgv-rCYP.
Mgv-rLACC could decolorize 66.9% and 63.7% of MG at a concentration of 10 mg/L and 20 mg/L (Figure 4B) in 24 h. However, in higher concentration of MG, Mgv-rLACC did not show obvious MG-decolorizing characteristic. The optimum pH of Mgv-rLACC was 4.0 (Figure 4D) and the optimum temperature was 50°C (Figure 4C). 10 mM and 100 mM sodium chloride additive dramatically inhibited the decolorizing activity of MG, up to 48.1% and 0%, respectively (Figure 6B). Metal-chelators, EDTA, had a negative impact on Mgv-rLACC (Figure 6B) due to the lack of Cu2+ which may serve as an electron transfer during MG degradation. There were no significant differences observed in the variation of metal ions (Figure 6A).
With increasing of the MG concentration, the Mgv-rCYP showed the trend of gradual reduction. The rates of MG degradation incubated with MG concentration of 10 mg/L, 20 mg/L, and 30 mg/L for 24 h were 66.5%, 54.1%, and 36.2%, respectively (Figure 5B). As Mgv-rLACC, no decolorizing phenomenon was detected with higher concentration of MG (Figure 5B). The optimum pH of Mgv-rCYP for MG degradation was 7.0 (Figure 5D) and the optimum temperature was 40°C (Figure 5C). Cu2+, Ni2+, and SDS could inhibit MG-degrading activities significantly (Figures 6A,B). MG-degrading characteristics of three recombination proteins were comparatively analyzed (Supplementary Table S3).
Intermediates of MG Degradation by the Three Recombination Proteins
By using LC-MS, different intermediates were detected in the MG-degrading process of three enzymes, respectively. Three intermediates from Mgv-rLACC according to the m/z values, were desmethyl-MG (m/z 315), didesmethyl-MG (m/z 302) and tetradesmethyl-LMG (m/z 274). Mgv-rCYP degraded MG into two products, didesmethyl-MG (m/z 302) and hydroxyl(didesmethyl)-MG (m/z 318). The intermediate products from Mgv-rPOD were the most diverse, including Desmethyl-MG (m/z 315), didesmethyl-MG (m/z 302), hydroxyl(didesmethyl)-MG (m/z 318), tetradesmethyl-LMG (m/z 274), hydroxyl-MG (m/z 346), Michler’s ketone (m/z 269), 4-(dimethylamino) benzophenone (m/z 226), and 4-(methylamino) benzophenone (m/z 212) (Figure 7 and Supplementary Figure S2).
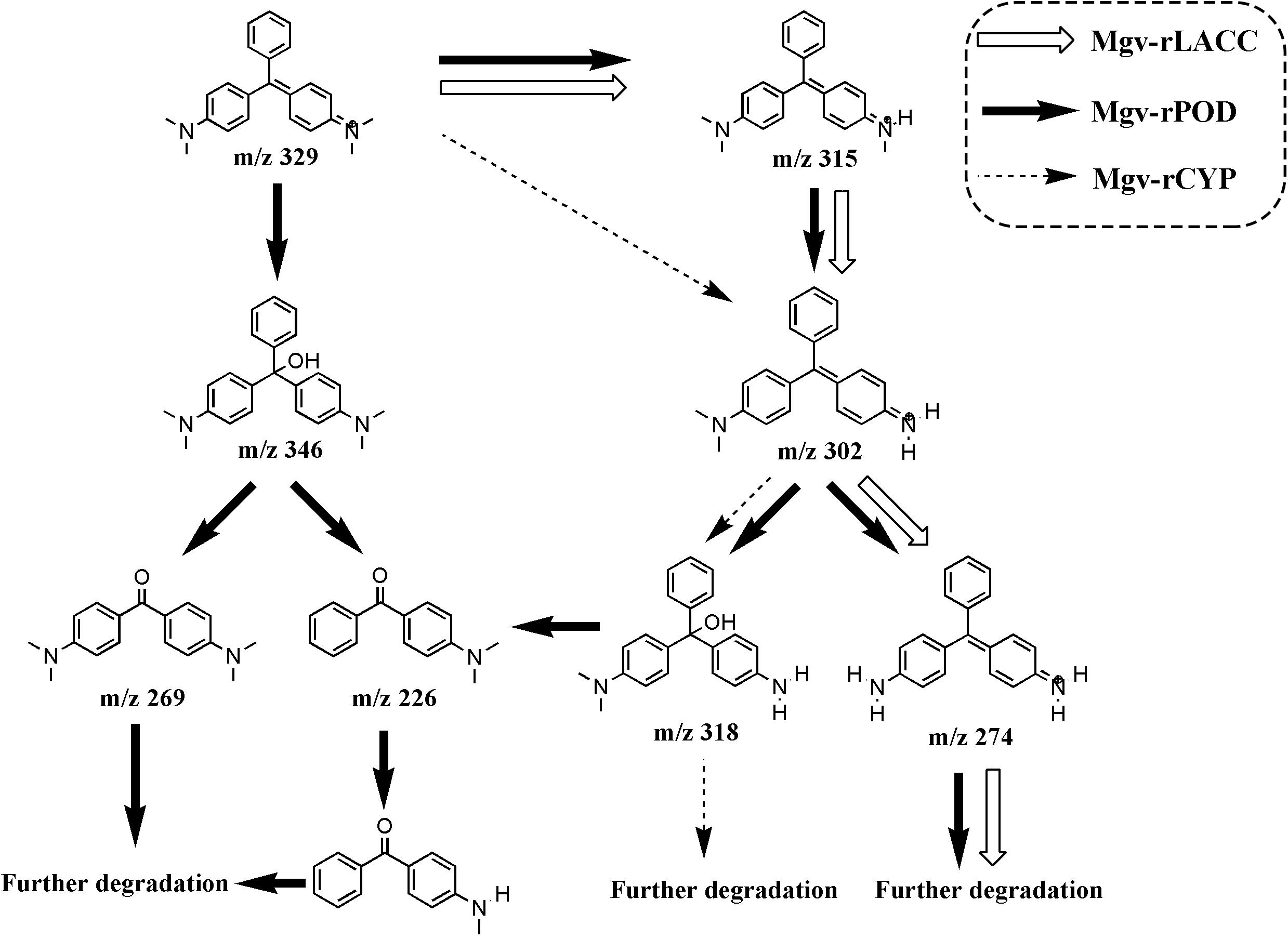
FIGURE 7. The putative MG degradation pathway deduced from the products of Mgv-rPOD, Mgv-rLACC, and Mgv-rCYP. The intermediates connected by the black, white and dashed arrows indicated the MG-degrading pathway of Mgv-rPOD, Mgv-rLACC, and Mgv-rCYP, respectively.
Discussion
Advanced metagenomic sequencing has been widely used in studying the structures and functions of microbial communities (Andreote et al., 2012; Soares et al., 2017). There were evidences from metagenomics analysis showed that abundant microorganisms and genes existing in mangrove environment could be implicated in the process of pollutant degrading (Andreote et al., 2012; Gonçalves et al., 2015). In this study, a broad diversity of bacterial and MDGs profiles were confirmed in mangrove sediments. However, the actual bioactivities of the functional genes were still unclear due to the lack of characteristics of complete gene products. Metagenomics based TAIL-PCR was an effective and relatively low-cost method for obtaining complete genes. The results that MDGs from different gene families were expressed and the corresponding MG-degrading characteristics were tested to further prove the validity of the strategy.
Mangroves are one among the world’s most productive ecosystems and are of ecological, economic, and societal importance (Mumby et al., 2004; Donato et al., 2011). Microbes play a key role in maintaining this productivity and conserve this ecosystem (Andreote et al., 2012). From our dataset of mangrove sediments, 666 bacterial genera (Figure 1) and 13 pollutant degradation pathways (Supplementary Table S2) were found. Genes that were found more frequently in one community than others are assumed to endow beneficial function on that community (Hugenholtz and Tyson, 2008). It was implied that microbes in the mangrove could constitute a large gene pool related to pollutant transformation with potential biotechnological and environmental implications. Besides, the apparent associations between various bacterial taxa and functional genes were analyzed (Figure 2D). Thirteen genera were identified to be the original genera of the 44 MDGs. Of them, 12 genera have been reported to be involved in organic pollutant degradation. Nitrosomonas could be the most active genera with the most diverse MDGs, including four glutathione peroxidase genes and one peroxidase gene. It was reported that the genus had significant activities to degrade halogenated aliphatic (Wahman et al., 2005), trichloroethylene (Suttinun et al., 2010) and PAHs (Chang et al., 2002). The Shewanella genus could also remove MG, methyl violet B (Chen et al., 2010), naphthylamine sulfonic azo dye (Hong et al., 2007) and naphthol green B dye (Xiao et al., 2012). This information provided important clues to discover microbial and genetic resources from the mangroves. According to the previous studies, lots of members of Proteobacteria and Bacteroidetes harbor many functional genes, including biodegradation genes, phenol degradation genes and PAHs degradation genes (Klankeo et al., 2009; Fang et al., 2013; Zhao et al., 2016). However, the functions of Proteobacteria and Bacteroidetes for MG degradation have been poorly studied. In our work, 18/30 and 8/30 of those bacterial origin, respectively, belonged to the phyla of Proteobacteria and Bacteroidetes, which indicated that Proteobacteria and Bacteroidetes also played a crucial role of MG degradation in mangrove sediments.
Forty-four genes were predicted to be related to MG-degrading and belonged to three gene classes (LACC, POD, and CYP, Figure 2C). Relative lower amino acid identities from 50% to 70% (Figure 2B) suggested most genes were new sequences which were not reported in other environments. The diversity and richness of functional gene represented by the metagenomes implied the potential of mangrove microorganisms in environment restoration. Three MDGs from three gene families were expressed and the MG-degrading characteristics from the corresponding recombinant proteins at varied testing levels were verified. Mgv-rPOD showed the highest MG-degrading activity among the three MG-degrading enzymes (Figure 3). Due to the usual low temperature of natural seawater (about 10–35°C), the low-temperature adaptability of Mgv-rPOD which kept a high degradation activity (95.8%) at 20°C, has great advantage applying in marine environment. A strong anti-interference ability to environmental factors, including the adaptabilities to metal ions (except Cu2+), metal-chelator (represented by EDTA), and the stability in different salinity further showed that Mgv-rPOD could be a promising bio-resource for removing MG from complex wastewaters. In addition, Mgv-rPOD had advantages in MG-degrading activity over not only the enzymes from this study but also other peroxidases reported in previous works. A manganese peroxidase, which showed the MG-degrading activity, was discovered from the white rot fungus Irpex lacteus F17. The manganese peroxidase could efficiently degrade 96% of MG at the concentration of 200 mg/L within 1 h; however, only about 75% of MG was degraded by using this enzyme at the MG concentration of 300 mg/L (Yang et al., 2016). By comparison, Mgv-rPOD could degrade more than 97% of MG within 30 min at the concentration of 300 mg/L, and the concentration was higher than most MG-degrading enzymes (Li et al., 2009; Saravanakumar et al., 2013; Zhang et al., 2013). Besides, many MG-degrading enzymes could only work at a narrow pH range. For instance, Saravanakumar et al. (2013) isolated manganese peroxidase isozyme H4 from Phanerochaete chrysosporium. However, about 50% MG-degrading activity of the enzymes was lost when the working pH was deviated from the optimum pH 4.5 (Saravanakumar et al., 2013). Unfortunately, it is very difficult to ensure the pH of nature environment when this enzyme was used in the remediation of MG pollution. Conversely, Mgv-rPOD possessed a wider pH range (7.0–11.0) for MG degradation, which suggested that Mgv-rPOD was more suitable for practice abatement of MG pollution. The possible mechanisms of MG degradation by Mgv-rPOD, Mgv-rLACC, and Mgv-rCYP were speculated according to the intermediates detected by LC-MS (Figure 7). All the processes of MG-degrading by the three recombination proteins were begin with a series of N-demethylation reactions, which was consistent with the former reports (Kedderis and Hollenberg, 1983; Cha et al., 2001; Murugesan et al., 2009). The products degrading by Mgv-rLACC and Mgv-rCYP were retained the triphenyl structure, and no further intermediates were detected, which was consistent with the previous study (Cha et al., 2001; Murugesan et al., 2009). The products of Mgv-rPOD suggested a further process of MG degradation. Mgv-rPOD not only mediated the N-demethylation but could also disconnect the triphenyl structure. Therefore, Mgv-rPOD was an attractive option for the remediation of MG pollution due to the high MG-degrading activity, high anti-interference, wide pH working condition, and the more complete degradation process.
Conclusion
Three recombinant proteins, especially Mgv-rPOD, were discovered and characterized as promising enzymes for the remediation of MG pollution in nature environment. Meanwhile, metagenomics provided an efficient path to view the microbial community structure and the functional (metabolic) potential of microbial community. The study further confirmed not-yet-cultivated bacteria from mangrove environments were a potential source for novel biocatalysts.
Author Contributions
WQ and JZ designed this work and wrote this paper. WQ, TL, and JZ performed the experiments in this work. DW and TL collected the sediment samples. GH revised this paper.
Funding
This research was supported by grants from the Scientific Research Project of Xiamen Southern Ocean Center (Grant No. 17GZP007NF03) and China Ocean Mineral Resources R&D Association (Grant No. DY-125-22-QY-18).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.02187/full#supplementary-material
Footnotes
- ^ https://www.ncbi.nlm.nih.gov/sra
- ^ http://string-db.org/
- ^ http://www.genome.jp/kegg/
- ^ http://www.ncbi.nlm.nih.gov/projects/gorf/
References
Algubury, H. (2016). The effect of coupled titanium dioxide and cobalt oxide on photo catalytic degradation of malachite green. Int. J. Chemtech Res. 9, 227–235.
Amann, R. I., Binder, B. J., Olson, R. J., Chisholm, S. W., Devereux, R., and Stahl, D. A. (1990). Combination of 16S rRNA-targeted oligonucleotide probes with flow cytometry for analyzing mixed microbial populations. Appl. Environ. Microbiol. 56, 1919–1925.
Andreote, F. D., Jiménez, D. J., Chaves, D., Dias, A. C., Luvizotto, D. M., Diniandreote, F., et al. (2012). The microbiome of Brazilian mangrove sediments as revealed by metagenomics. PLOS One 7:e38600. doi: 10.1371/journal.pone.0038600
Cha, C. J., Doerge, D. R., and Cerniglia, C. E. (2001). Biotransformation of malachite green by the fungus Cunninghamella elegans. Appl. Environ. Microbiol. 67, 4358–4360. doi: 10.1128/AEM.67.9.4358-4360.2001
Chang, G. R., Chen, H. S., and Lin, F. Y. (2016). Analysis of banned veterinary drugs and herbicide residues in shellfish by liquid chromatography-tandem mass spectrometry (LC/MS/MS) and gas chromatography-tandem mass spectrometry (GC/MS/MS). Mar. Pollut. Bull. 113, 579–584. doi: 10.1016/j.marpolbul.2016.08.080
Chang, J. S., and Lin, C. Y. (2001). Decolorization kinetics of a recombinant Escherichia coli strain harboring azo-dye-decolorizing determinants from Rhodococcus sp. Biotechnol. Lett. 23, 631–636. doi: 10.1023/A:1010306114286
Chang, S. W., Hyman, M. R., and Williamson, K. J. (2002). Cooxidation of naphthalene and other polycyclic aromatic hydrocarbons by the nitrifying bacterium. Nitrosomonas europaea. Biodegradation 13, 373–381. doi: 10.1023/A:1022811430030
Chen, C. H., Chang, C. F., and Liu, S. M. (2010). Partial degradation mechanisms of malachite green and methyl violet B by Shewanella decolorationis NTOU1 under anaerobic conditions. J. Hazard. Mater. 177, 281–289. doi: 10.1016/j.jhazmat.2009.12.030
Chen, G., and Miao, S. (2010). HPLC determination and ms confirmation of malachite green, gentian violet, and their leuco metabolite residues in channel catfish muscle. J. Agric. Food Chem. 58, 7109–7114. doi: 10.1021/jf9043925
Donato, D. C., Kauffman, J. B., Murdiyarso, D., Kurnianto, S., Stidham, M., and Kanninen, M. (2011). Mangroves among the most carbon-rich forests in the tropics. Nat. Geosci. 4, 293–297. doi: 10.1038/ngeo1123
Du, L. N., Wang, S., Li, G., Wang, B., Jia, X. M., Zhao, Y. H., et al. (2011). Biodegradation of malachite green by Pseudomonas sp. strain DY1 under aerobic condition: characteristics, degradation products, enzyme analysis and phytotoxicity. Ecotoxicology 20, 438–446. doi: 10.1007/s10646-011-0595-3
Du, L. N., Zhao, M., Li, G., Xu, F. C., Chen, W. H., and Zhao, Y. H. (2013). Biodegradation of malachite green by Micrococcus sp. strain BD15: biodegradation pathway and enzyme analysis. Int. Biodeterior. Biodegradation 78, 108–116. doi: 10.1016/j.ibiod.2012.12.011
Fang, H., Cai, L., Yu, Y., and Zhang, T. (2013). Metagenomic analysis reveals the prevalence of biodegradation genes for organic pollutants in activated sludge. Bioresour. Technol. 129, 209–218. doi: 10.1016/j.biortech.2012.11.054
Fang, Z., Li, T., Wang, Q., Zhang, X., Peng, H., Fang, W., et al. (2011). A bacterial laccase from marine microbial metagenome exhibiting chloride tolerance and dye decolorization ability. Appl. Microbiol. Biotechnol. 89, 1103–1110. doi: 10.1007/s00253-010-2934-3
Fang, Z. M., Li, T. L., Chang, F., Zhou, P., Fang, W., Hong, Y. Z., et al. (2012). A new marine bacterial laccase with chloride-enhancing, alkaline-dependent activity and dye decolorization ability. Bioresour. Technol. 111, 36–41. doi: 10.1016/j.biortech.2012.01.172
Giri, C., Ochieng, E., Tieszen, L. L., Zhu, Z., Singh, A., Loveland, T., et al. (2011). Status and distribution of mangrove forests of the world using earth observation satellite data. Glob. Ecol. Biogeogr. 20, 154–159. doi: 10.1111/j.1466-8238.2010.00584.x
Gomathi, V., Saravanakumar, K., and Kathiresan, K. (2013). Bio removal of malachite green by mangrove-derived Aplanochytrium sp. KGA2512. Afr. J. Microbiol. Res. 7, 3056–3065. doi: 10.5897/AJMR12.843
Gomes Gomes, C. M., Borges, L. R., Paranhos, R., Pinto, F. N., Mendonçahagler, L. C. S., and Smalla, K. (2008). Exploring the diversity of bacterial communities in sediments of urban mangrove forests. FEMS Microbiol. Ecol. 66, 96–109. doi: 10.1111/j.1574-6941.2008.00519.x
Gonçalves, A. C., Dos Santos, A. C., Dos Santos, T. F., Pessoa, T. B., Dias, J. C., and Rezende, R. P. (2015). High yield of functional metagenomic library from mangroves constructed in fosmid vector. Genet. Mol. Res. 14, 11841–11847. doi: 10.4238/2015.October.2.17
Gopinathan, R., Kanhere, J., and Banerjee, J. (2015). Effect of malachite green toxicity on non target soil organisms. Chemosphere 120, 637–644. doi: 10.1016/j.chemosphere.2014.09.043
Grosser, R. J., Warshawsky, D., and Vestal, J. R. (1991). Indigenous and enhanced mineralization of pyrene, benzo[a]pyrene, and carbazole in soils. Appl. Environ. Microbiol. 57, 3462–3469.
Hess, M., Sczyrba, A., Egan, R., Kim, T. W., Chokhawala, H., Schroth, G., et al. (2011). Metagenomic discovery of biomass-degrading genes and genomes from cow rumen. Science 331, 463–467. doi: 10.1126/science.1200387
Hong, Y., Guo, J., Xu, Z., Mo, C., Xu, M., and Sun, G. (2007). Reduction and partial degradation mechanisms of naphthylaminesulfonic azo dye amaranth by Shewanella decolorationis S12. Appl. Microbiol. Biotechnol. 75, 647–654. doi: 10.1007/s00253-007-0838-7
Hugenholtz, P., and Tyson, G. W. (2008). Microbiology: metagenomics. Nature 455, 481–483. doi: 10.1038/455481a
Jefferson, J., and Jones, J. O. F. I. I. I. (2003). Decolorization of malachite green and crystal violet by waterborne pathogenic mycobacteria. Antimicrob. Agents Chemother. 47, 2323–2326. doi: 10.1128/AAC.47.7.2323-2326.2003
Jiang, L. (2011). Microbiodegradation of Two Aquacultural Drugs in the Sediment. Nanjing: Nanjing University.
Jones, E. J. W., Perry, R. G., and Wild, J. L. (2011). Geology of the hebridean margin of the Rockall Trough. Proc. R. Soc. Edinb. 88, 27–51.
Kedderis, G. L., and Hollenberg, P. F. (1983). Characterization of the N-demethylation reactions catalyzed by horseradish peroxidase. J. Biol. Chem. 258, 8129–8138.
Kim, M. H., Kim, Y., Park, H. J., Lee, J. S., Kwak, S. N., Jung, W. H., et al. (2008). Structural insight into bioremediation of triphenylmethane dyes by Citrobacter sp. triphenylmethane reductase. J. Biol. Chem. 283, 31981–31990. doi: 10.1074/jbc.M804092200
Kimura, N. (2006). Metagenomics: access to unculturable microbes in the environment. Microbes Environ. 21, 201–215. doi: 10.1264/jsme2.21.201
Klankeo, P., Nopcharoenkul, W., and Pinyakong, O. (2009). Two novel pyrene-degrading Diaphorobacter sp. and Pseudoxanthomonas sp. isolated from soil. J. Biosci. Bioeng. 108, 488–495. doi: 10.1016/j.jbiosc.2009.05.016
Li, L. T., Hong, Q., Yan, X., Fang, G. H., Ali, S. W., and Li, S. P. (2009). Isolation of a malachite green-degrading Pseudomonas sp. MDB-1 strain and cloning of the tmr2 gene. Biodegradation 20, 769–776. doi: 10.1007/s10532-009-9265-z
Liang, S. X., Jia, Z., Zhang, W. C., Wang, W. M., and Zhang, L. C. (2017). Rapid malachite green degradation using Fe 73.5 Si 13.5 B 9 Cu 1 Nb 3 metallic glass for activation of persulfate under UV–Vis light. Mater. Des. 119, 244–253. doi: 10.1016/j.matdes.2017.01.039
Marchand, C., Fernandez, J. M., and Moreton, B. (2016). Trace metal geochemistry in mangrove sediments and their transfer to mangrove plants (New Caledonia). Sci. Total Environ. 562, 216–227. doi: 10.1016/j.scitotenv.2016.03.206
Mumby, P. J., Edwards, A. J., Ariasgonzález, J. E., Lindeman, K. C., Blackwell, P. G., Gall, A., et al. (2004). Mangroves enhance the biomass of coral reef fish communities in the Caribbean. Nature 427, 533–536. doi: 10.1038/nature02286
Murugesan, K., Yang, I. H., Kim, Y. M., Jeon, J. R., and Chang, Y. S. (2009). Enhanced transformation of malachite green by laccase of Ganoderma lucidum in the presence of natural phenolic compounds. Appl. Microbiol. Biotechnol. 82, 341–350. doi: 10.1007/s00253-008-1819-1
Ouyang, X., Lee, S. Y., and Connolly, R. M. (2017). The role of root decomposition in global mangrove and saltmarsh carbon budgets. Earth Sci. Rev. 166, 53–63. doi: 10.1016/j.earscirev.2017.01.004
Pathania, D., Katwal, R., Sharma, G., Naushad, M., Khan, M. R., and Al-Muhtaseb, A. A. H. (2016). Novel guar gum/Al 2 O 3 nanocomposite as an effective photocatalyst for the degradation of malachite green dye. Int. J. Biol. Macromol. 87, 366–374. doi: 10.1016/j.ijbiomac.2016.02.073
Pope, P. B., and Moran, N. A. (2010). Adaptation to herbivory by the Tammar wallaby includes bacterial and glycoside hydrolase profiles different from other herbivores. Proc. Natl. Acad. Sci. U.S.A. 107, 14793–14798. doi: 10.1073/pnas.1005297107
Ren, S., Guo, J., Zeng, G., and Sun, G. (2006). Decolorization of triphenylmethane, azo, and anthraquinone dyes by a newly isolated Aeromonas hydrophila strain. Appl. Microbiol. Biotechnol. 72, 1316–1321. doi: 10.1007/s00253-006-0418-2
Saravanakumar, T., Palvannan, T., Kim, D. H., and Park, S. M. (2013). Manganese peroxidase H4 isozyme mediated degradation and detoxification of triarylmethane dye malachite green: optimization of decolorization by response surface methodology. Appl. Biochem. Biotechnol. 171, 1178–1193. doi: 10.1007/s12010-013-0220-4
Schloss, P. D., and Handelsman, J. (2005). Metagenomics for studying unculturable microorganisms: cutting the Gordian knot. Genome Biol. 6:229. doi: 10.1186/gb-2005-6-8-229
Simon, C., and Daniel, R. (2009). Achievements and new knowledge unraveled by metagenomic approaches. Appl. Microbiol. Biotechnol. 85, 265–276. doi: 10.1007/s00253-009-2233-z
Soares, F. L., Marcon, J., Khakhum, N., Cerdeira, L. T., Ottoni, J. R., Domingos, D. F., et al. (2017). A novel multifunctional β-N-acetylhexosaminidase revealed through metagenomics of an oil-spilled mangrove. Bioengineering 4:62. doi: 10.3390/bioengineering4030062
Srinivasan, G. P., Sikkanthar, A., Elamaran, A., Delma, C. R., and Subramaniyan, K. (2014). Biodegradation of carcinogenic textile azo dyes using bacterial isolates of mangrove sediment. J. Coast. Life Med. 2, 154–162.
Srivastava, S., Sinha, R., and Roy, D. (2004). Toxicological effects of malachite green. Aquat. Toxicol. 66, 319–329. doi: 10.1016/j.aquatox.2003.09.008
Suttinun, O., Müller, R., and Luepromchai, E. (2010). Cometabolic degradation of trichloroethene by Rhodococcus sp. strain L4 immobilized on plant materials rich in essential oils. Appl. Environ. Microbiol. 76, 4684–4690. doi: 10.1128/AEM.03036-09
Tayabali, A. F., Coleman, G., Crosthwait, J., Nguyen, K. C., Zhang, Y., and Shwed, P. (2017). Composition and pathogenic potential of a microbial bioremediation product used for crude oil degradation. PLOS One 12:e0171911. doi: 10.1371/journal.pone.0171911
Ulson de Souza, S., Forgiarini, E., and Ulson de Souza, A. A. (2007). Toxicity of textile dyes and their degradation by the enzyme horseradish peroxidase (HRP). J. Hazard. Mater. 147, 1073–1078. doi: 10.1016/j.jhazmat.2007.06.003
Venil, C. K., and Lakshmanaperumalsamy, P. (2010). Dye decolorizing potential of a novel fungus Coriolus versicolor ML04 in the medium optimized by response surface methodology. Braz. Arch. Biol. Technol. 53, 1503–1510. doi: 10.1590/S1516-89132010000600028
Wahman, D. G., Katz, L. E., and Speitel, G. E. Jr. (2005). Cometabolism of trihalomethanes by Nitrosomonas europaea. Appl. Environ. Microbiol. 71, 7980–7986. doi: 10.1128/AEM.71.12.7980-7986.2005
Wang, J., Gao, F., Liu, Z., Qiao, M., Niu, X., Zhang, K. Q., et al. (2012). Pathway and molecular mechanisms for malachite green biodegradation in Exiguobacterium sp. MG2. PLOS One 7:e51808. doi: 10.1371/journal.pone.0051808
Xiao, X., Xu, C. C., Wu, Y. M., Cai, P. J., Li, W. W., Du, D. L., et al. (2012). Biodecolorization of naphthol green B dye by Shewanella oneidensis MR-1 under anaerobic conditions. Bioresour. Technol. 110, 86–90. doi: 10.1016/j.biortech.2012.01.099
Yang, C. W., Lee, C. C., Ku, H., and Chang, B. V. (2017). Bacterial communities associated with anaerobic debromination of decabromodiphenyl ether from mangrove sediment. Environ. Sci. Pollut. Res. 24, 5391–5403. doi: 10.1007/s11356-016-8259-8
Yang, J., Yang, X., Lin, Y., Ng, T. B., Lin, J., and Ye, X. (2015). Laccase-catalyzed decolorization of malachite green: performance optimization and degradation mechanism. PLOS One 10:e0127714. doi: 10.1371/journal.pone.0127714
Yang, X., Zheng, J., Lu, Y., and Jia, R. (2016). Degradation and detoxification of the triphenylmethane dye malachite green catalyzed by crude manganese peroxidase from Irpex lacteus F17. Environ. Sci. Pollut. Res. Int. 23, 9585–9597. doi: 10.1007/s11356-016-6164-9
Yong, L., Zhanqi, G., Yuefei, J., Xiaobin, H., Cheng, S., Shaogui, Y., et al. (2015). Photodegradation of malachite green under simulated and natural irradiation: kinetics, products, and pathways. J. Hazard. Mater. 285, 127–136. doi: 10.1016/j.jhazmat.2014.11.041
Zhang, C., Zhang, S., Diao, H., Zhao, H., Zhu, X., Lu, F., et al. (2013). Purification and characterization of a temperature- and pH-Stable laccase from the spores of Bacillus vallismortis fmb-103 and Its application in the degradation of malachite green. J. Agric. Food Chem. 61, 5468–5473. doi: 10.1021/jf4010498
Keywords: mangrove sediment, metagenome, MG biodegradation, gene expression, biochemical analysis, degradation pathway
Citation: Qu W, Liu T, Wang D, Hong G and Zhao J (2018) Metagenomics-Based Discovery of Malachite Green-Degradation Gene Families and Enzymes From Mangrove Sediment. Front. Microbiol. 9:2187. doi: 10.3389/fmicb.2018.02187
Received: 28 April 2018; Accepted: 27 August 2018;
Published: 11 September 2018.
Edited by:
Shaohua Chen, South China Agricultural University, ChinaReviewed by:
Dong Li, University of California, Santa Barbara, United StatesQing Hong, Nanjing Agricultural University, China
Hongzhi Tang, Shanghai Jiao Tong University, China
Copyright © 2018 Qu, Liu, Wang, Hong and Zhao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jing Zhao, sunnyzhaoj@xmu.edu.cn orcid.org/0000-0003-1895-7620