 Caiyun Yang1
Caiyun Yang1 Qi Wang1Paulina N. Simon1Jinyu Liu1Lincong Liu1Xianzhu Dai1Xiaohui Zhang1
Qi Wang1Paulina N. Simon1Jinyu Liu1Lincong Liu1Xianzhu Dai1Xiaohui Zhang1 Jialiang Kuang2Yasuo Igarashi1Xuejun Pan3*
Jialiang Kuang2Yasuo Igarashi1Xuejun Pan3* Feng Luo1*
Feng Luo1*- 1Research Center of Bioenergy and Bioremediation, Southwest University, Chongqing, China
- 2State Key Laboratory of Biocontrol, Guangdong Key Laboratory of Plant Resources and Conservation of Guangdong Higher Education Institutes, College of Ecology and Evolution, Sun Yat-sen University, Guangzhou, China
- 3Faculty of Environmental Science and Engineering, Kunming University of Science and Technology, Kunming, China
Particle-associated bacteria (PAB) and free-living bacteria (FLB) from aquatic environments during phytoplankton blooms differ in their physical distance from algae. Both the interactions within PAB and FLB community fractions and their relationship with the surrounding environmental properties are largely unknown. Here, by using high-throughput sequencing and network-based analyses, we compared the community and network characteristics of PAB and FLB from a plateau lake during a Microcystis aeruginosa bloom. Results showed that PAB and FLB differed significantly in diversity, structure and microbial connecting network. PAB communities were characterized by highly similar bacterial community structure in different sites, tighter network connections, important topological roles for the bloom-causing M. aeruginosa and Alphaproteobacteria, especially for the potentially nitrogen-fixing (Pleomorphomonas) and algicidal bacteria (Brevundimonas sp.). FLB communities were sensitive to the detected environmental factors and were characterized by significantly higher bacterial diversity, less connectivity, larger network size and marginal role of M. aeruginosa. In both networks, covariation among bacterial taxa was extensive (>88% positive connections), and bacteria potentially affiliated with biogeochemical cycling of nitrogen (i.e., denitrification, nitrogen-fixation and nitrite-oxidization) were important in occupying module hubs, such as Meganema, Pleomorphomonas, and Nitrospira. These findings highlight the importance of considering microbial network interactions for the understanding of blooms.
Introduction
Algal blooms could cause severe ecological problems in both marine and fresh water and have both a high frequency and large distribution (Yang C. et al., 2012; Woodhouse et al., 2016). Blooms reduce the water quality (Cai et al., 2013a,b) (e.g., restrict light penetration and increase water viscosity), increase the economic costs of water treatment, change food web dynamics (Turner and Chislock, 2010) and threaten human health (Paerl and Otten, 2013). The relationship between bloom-forming algae and other water microbes has received great attention because they are reported to closely interact; the diversity and community composition of water microbes can be significantly influenced by blooms (Yang C. et al., 2012, 2015; Woodhouse et al., 2016) but can also affect bloom development (Grossart et al., 2005, 2006; Yang C. et al., 2012; Cai et al., 2013a).
Water microbes consist of numerous diverse species (Cai et al., 2013a) and play key roles in biospheric biogeochemical cycling (Woese, 1994). In water ecosystem, bacteria can live either freely or attached to particle (e.g., algae) surfaces (Jasti et al., 2005), with obvious differences in community composition, phylogeny and metabolism (Riemann and Winding, 2001; Grossart et al., 2005; Cao et al., 2015). Free-living bacteria (FLB) are the primary degraders of dissolved organic matter (DOM) in aquatic ecosystems (Karner and Herndl, 1992), and particle-associated bacteria (PAB) are important for the remineralization of both dissolved and particulate organic matter (Tang et al., 2009). PAB are usually associated with colloids and algae (Shao et al., 2014) and are considered algae-associated microbes, particularly during algal blooms (Riemann and Winding, 2001; Cai et al., 2013b), that were shown to have positive or negative effects on algal growth (e.g., algicidal bacteria) in a previous study (Berg et al., 2009). Microbes do not live separately but rather interact with each other to form ecological networks and accomplish ecosystem functions (e.g., material and energy flows, ecosystem stability) (Deng et al., 2015; Shi et al., 2016). Therefore, explaining and predicting the variation in network structure are essential parts of microbial ecology (Zhou et al., 2010) and are useful for guiding practical applications, including biodiversity preservation and ecosystem management. Although the importance of FLB and PAB during algal blooms has been widely addressed (Tuomainen et al., 2006; Cai et al., 2013b; Shao et al., 2014; Cao et al., 2015; Yang C. et al., 2015), the bacterial interactions within the two fractions are still unknown.
Our understanding of microbial interactions in natural environments is limited because most of these microbes are unculturable (Zhou et al., 2010). Culture-independent molecular technology has developed rapidly in recent years and has been acknowledged as a feasible approach to study complex microbial communities. Emerging high-throughput sequencing technologies can provide a comprehensive understanding of the specific microbial species that live and thrive in various environments (Logares et al., 2014), and network-based approaches have been successfully used to evaluate the influence of environmental disturbances on the whole community (Raes and Bork, 2008; Zhou et al., 2010) and analyze microbial interactions in diverse habitats, including marine (Wang Y. et al., 2015), soil (Tu et al., 2015), acid mine drainage (Kuang et al., 2016) and groundwater environments (Deng et al., 2015). However, relevant research on microbial interactions during algal blooms is still limited, especially for the relationship between algae and associated bacteria. Woodhouse et al. (2016) found that microbial communities were closely associated with cyanobacterial composition by using datasets of cyanobacterial cell densities, bacterial community composition and abiotic parameters to demonstrate the interactions of abiotic and biotic factors during cyanobacterial blooms in a shallow ephemeral lake. Nevertheless, the network interaction of PAB or FLB with algae during blooms is still largely unknown.
To fill this gap, in this study, we focused on the following questions during a cyanobacterial bloom in a freshwater lake in the Yunnan-Guizhou plateau: (1) the differences in community composition between PAB and FLB during cyanobacterial blooms; (2) how they respond to environmental fluctuation; and most of all, (3) how the microbes interact in PAB and FLB, particularly considering the role of cyanobacteria in the network interactions.
Materials and Methods
Site Description and Sample Collection
The largest plateau lake in China-Dianchi Lake is found in Yunnan Province (Southwest China), with an altitude of 1887 m and a water area of ∼330 km2. It is separated into two areas (Caohai and Waihai) by the Haigeng Dam (Figure 1A). This lake has become a heavily eutrophied body of water since the 1990s, mainly due to sewage import and phosphorite deposits from the Early Cambrian (Maixing et al., 2003) in this area. Excessive nutrients, strong sunlight and warm temperatures result in the considerable growth of cyanobacteria in this lake. Blooms occur nearly year round and are much more severe in summer and autumn (Li et al., 2003). Specifically, blooms occur regionally in the northwestern side of Waihai with obvious free-floating cyanobacteria and often last more than 8 months (from April to November) annually (Wang et al., 2012).
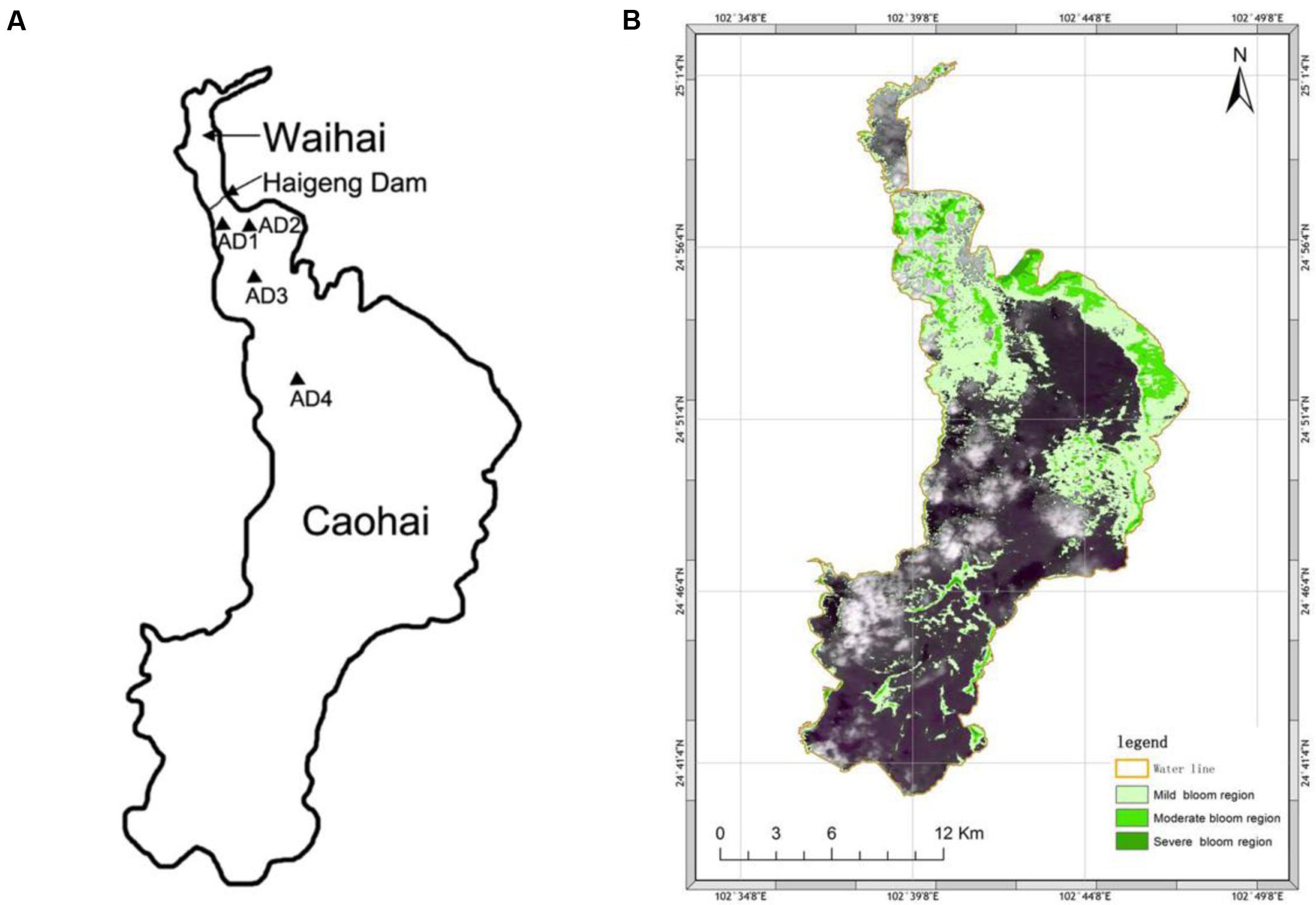
FIGURE 1. Sampling location (A) and bloom area distribution (B) of Dianchi Lake during sampling. Sites are marked with triangular icons in Caohai.
The dynamics of the bacterial community associated with a cyanobacterial bloom in Dianchi Lake were investigated in this study. Samples were collected in the Waihai of Dianchi Lake at four sites [AD1 (N24.571, E102.394), AD2 (N24.571, E102.385), AD3 (N 24.555, E 102.394), and AD4 (N 24.526, E 102.405)] during a cyanobacterial bloom on October 16th, 2015 (Figure 1A). AD1 and AD2 were taken from an area where blooms occur frequently, while AD3 and AD4 were close to and away from this bloom area, respectively. Water samples (2 L for each sample) were collected in three replicates from each site within 20 m2 (depth of 0.5 m) using autoclaved polypropylene sampling vials. Immediately after collection, samples for phytoplankton species identification and counts were preserved using 1.5% Lugol’s iodine solution (Lack, 1971), and a separate sample was fixed with PBS-buffered formaldehyde (2% final concentration) for bacteria enumeration. The 5-μm filters combined with a filtration volume strategy were used to separate the PAB and FLB. Specifically, water was filtered through 5-μm filters (Millipore, United States) to collect PAB, and the water volume (60–85 ml) was chosen to avoid retention of FLB (the filtration process should be smooth and rapid). Although the single bloom-causing algae (M aeruginosa, cell size ∼1–2 μm) could pass through, they usually aggregate together with other cells in natural water and are retained on the 5-μm filter. Thus, the PAB community contained both algae and bacteria that were attached to algae. The FLB and small amounts of single M. aeruginosa in the filtrate were collected with 0.22-μm filters (Millipore, United States). In total, 24 samples (12 for PAB, 12 for FLB) were collected, and all filters were stored at -70°C until nucleic acid extractions.
Water Characterization
The pH was measured using a handheld pH meter (Model IQ150, IQ Scientific Instruments, United States). Dissolved oxygen (DO) and temperature were measured using a handheld DO meter (AM-39-Sensotechnik Meinsberg, Germany). Turbidity and suspended particles were measured using a MultiDirect spectrophotometer (Lovibond, Germany).
Bacteria enumeration was performed using epifluorescence microscopy using DAPI stain. Samples fixed with PBS-buffered formaldehyde were stained with 4′,6-diamidino-2-phenylindole fluorochrome (DAPI; final concentration of 5 μg ml-1) (Porter and Feig, 1980); bacterial cells were counted using epifluorescence microscopy and mean bacterial volumes were determined using measurements of 30–100 cells using photographic slides; 15 random visual fields were analyzed for each sample. Phytoplankton cells preserved with Lugol’s iodine solution were counted under an inverted microscope with a hemocytometer (Lund et al., 1958).
The concentrations of NH4-N, NO2-N, NO3-N, total N and soluble phosphate were measured using Nessler’s reagent spectrophotometry (Chen and Hu, 2011), spectrophotometry, phenol disulfonic acid spectrophotometry, alkaline potassium persulfate digestion ultraviolet spectrophotometry and ammonium molybdate spectrophotometry using a UV-2600 UV-VIS spectrophotometer (State Environmental Protection Administration of China, 1989). Chemical oxygen demand (COD) was measured using a Lovibond RD 125 COD Reactor (Germany) and an ET99732 multi-parameter water quality analyzer (Lovibond, Germany) with an ET99955 kit (determination range: 0–150 mg/L O2) according to the operation manual. The eutrophication index (EI) was calculated as EI = DIN × DIP × COD × 106/4500 (Yang C. et al., 2015), where DIN is the dissolved inorganic nitrogen (sum of NO3-N, NO2-N, and NH4-N) content in mg/L; DIP is the dissolved inorganic phosphorus content (soluble phosphorus PO4-P) in mg/L; and COD is the chemical oxygen demand in mg/L.
Microbial Community Analysis
DNA Extraction and PCR Amplification
The microbial cells were scraped from the collected filters using sodium phosphate buffer. Total DNA was extracted using a FastDNA Spin Kit for Soil (MP Biomedicals, United States) and detected using 1% agarose gel electrophoresis. The V3–V4 region of the microbial 16S rRNA gene was amplified with primers 338F (5′-barcode-ACT CCT ACG GGA GGC AGC AG-3′) and 806R (5′-GGA CTA CHV GGG TWT CTA AT-3′). Amplicons were purified using an AxyPrepTM DNA Gel Extraction Kit (AXYGEN, United States) and quantified using a QuantiFluor-ST Fluorometer (Promega, United States), where the barcode is an eight-base sequence unique to each sample. PCR reactions were performed in triplicate in a 20-μL mixture containing 4 μL of 5 × FastPfu Buffer, 2 μL of 2.5 mM dNTPs, 0.8 μL of each primer (5 μM), 0.4 μL of FastPfu Polymerase, and 10 ng of template DNA.
Illumina MiSeq Sequencing
Equimolar amounts of purified amplicons were pooled and paired-end sequences (2 × 250bp) were retrieved from an Illumina MiSeq platform at Majorbio Bio-Pharm Technology Co., Ltd. (Shanghai, China) using a TruSeqTM DNA Sample Prep Kit (Illumina, United States). Raw reads of this study have been deposited into the NCBI Sequence Read Archive using accession No. PRJNA341818.
Processing of High-Throughput Sequencing Data
Fastq files of raw reads were demultiplexed and quality-filtered using QIIME (version 1.9.1) using the following criteria: (i) the 300 bp reads were truncated at any site with an average quality score < 20 over a 50-bp sliding window, discarding the truncated reads that were shorter than 50 bp; (ii) exact barcode matching, 2-nucleotide mismatch in primer matching and reads containing ambiguous characters were removed; (iii) only sequences with overlap longer than 10 bp were assembled according to their overlap sequence. Reads that could not be assembled were discarded.
After deleting unqualified sequences, the identification of operational taxonomic units (OTUs) at a 97% similarity level were performed using UPARSE (version 7.1)1; chimeric sequences were identified and removed using UCHIME. Taxonomic assignment was performed using RDP Classifier2 against the SILVA (SSU123)16S rRNA database using a confidence threshold of 70% (Amato et al., 2013). The alpha-diversities, including Shannon index (Hill, 1973), Chao richness estimator (Chao, 1984) and Good’s coverage (Esty, 1986) were calculated using Mothur (version: 1.30.1) (Schloss et al., 2011). The number of reads ranged from 31,736 to 99,733 in our samples, and 31,736 sequences were randomly resampled for subsequent statistical analyses. Additionally, some analyses were conducted based on the 4512 resampled sequences that were not affiliated with cyanophyta.
Network Construction and Analysis
The network analyses were performed using the online Molecular Ecological Network Approach Pipeline3, which implements a random matrix theory (RMT) for threshold identification (Deng et al., 2012). The RMT-based approach is a widely used and robust tool for network construction and has been successfully used to construct various networks, including gene regulatory networks (Zhou et al., 2010; Lin et al., 2011, 2013), functional molecular ecological networks (Zhou et al., 2010) and phylogenetic molecular ecological networks (Zhou et al., 2011; Tu et al., 2016). To construct reliable co-occurrence ecological networks, the RMT-based network method was used (Deng et al., 2015), and only OTUs observed in at least 8 of the 12 samples were used for subsequent network analyses. Specifically, the validated pairwise correlations between OTUs were determined based on a Pearson correlation coefficient cutoff of 0.9 and 0.89 that was automatically identified based on RMT for PAB and FLB, respectively, and then was used for network construction. Modules were detected by fast greedy modularity optimization (Clauset et al., 2010), and the reconstructed ecological networks were then visualized using Cytoscape (Shannon et al., 2003). The node connectivity was determined based on both within-module connectivity (Zi) and among-module connectivity (Pi) (Guimerà and Amaral, 2005), which were then used to classify nodes into four topological roles: module hubs (highly connected nodes within modules with Zi > 2.5), network hubs (highly connected nodes within the entire network with Zi > 2.5 and Pi > 0.62), connectors (nodes that connect modules with Pi > 0.62), and peripherals (nodes connected in modules with few outside connections with Zi < 2.5 and Pi < 0.62) (Zhou et al., 2010; Deng et al., 2012).
Statistical Analyses
Most statistical analyses were performed using the vegan (Oksanen et al., 2013) and agricolae packages (Mendiburu, 2014) in R (R Core Team, 2015). Dissimilarity tests were conducted based on the Bray-Curtis dissimilarity index and two non-parametric tests [i.e., permutational multivariate analysis of variance (Adonis) and multiple-response permutation procedure (MRPP)]. Principal component analysis (PCA) was used to display and compare the patterns of microbial communities among different samples, and canonical correspondence analysis (CCA) was used to link variations in microbial communities to environmental properties. Variation partitioning analysis (VPA) was conducted to examine the contribution of environmental factors in influencing microbial communities according to the CCA analysis. All significant differences were defined as a p-value of < 0.05.
Results
Water Chemical, Physical, and Biotic Characteristics
A large bloom dominated the northern portion of Dianchi Lake during the sampling period (Figure 1). It was a Microcystis sp. bloom, according to microscopic examination (Microcystis cells represented 98.27–100% of the total algae, Supplementary Table S1). Table 1 shows the measured environmental properties of all samples. The water pH was much higher than 7 and even reached 10.19 because of the algal bloom (Yang C. et al., 2015). The lake exhibited eutrophication (EI > 1), which was much more severe in AD1 and AD2 (Table 1). The soluble phosphorus (PO4-P) concentrations were very low; most were below the detectable limit (0.01 mg/L) of our method. Furthermore, the bacterial density was much higher in AD1 and AD2 and was positively correlated at all sites with NO3-N (spearman correlation, p < 0.01), NO2-N (p = 0.03), EI (p = 0.04) and pH (p = 0.035). Additionally, algal density was positively correlated with NO2-N (p = 0.02), NO3-N (p = 0.04), COD (p < 0.01), and EI (p < 0.01) at all sites.
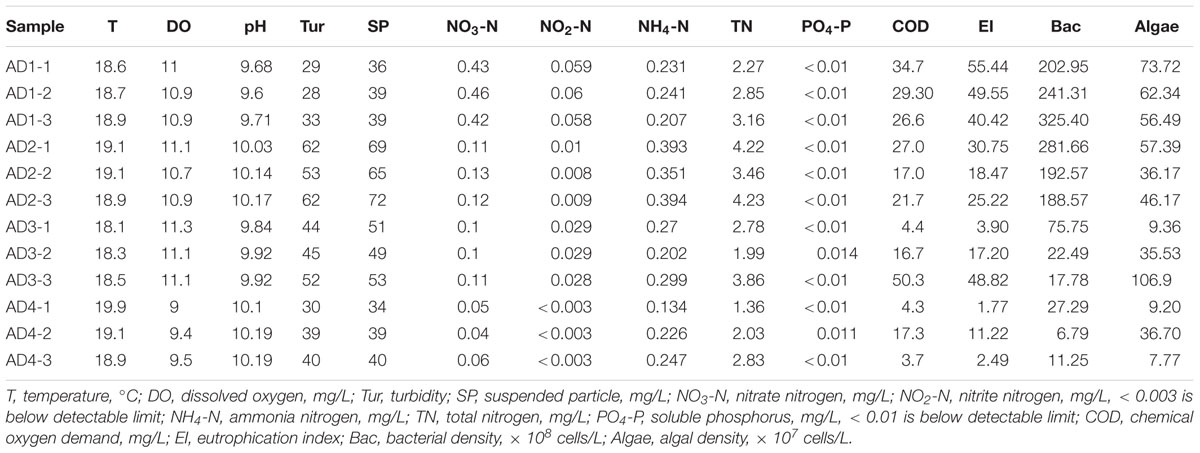
TABLE 1. Environmental and biological properties of samples.
Microbial Diversity and Community Composition
In this study, a total of 1,638,726 high-quality sequences remained after processing and singleton removal (total 118,108 sequences), and 1633 OTUs (PAB: 1224 OTUs and FLB: 1574 OTUs) were identified at a 97% similarity level. The sequencing depth was sufficient to represent entire whole microbial community because the sequence Good’s coverage was higher than 99.5%. These OTUs belonged to 37 phyla, 54 classes, 104 orders, 182 families and 275 genera (Supplementary Figure S1a). Most taxa were shared between the microbial communities of PAB and FLB, and more unique taxa were found in the community of FLB at all five taxonomic levels (Supplementary Figure S1b).
A randomly selected subset of 31,736 sequences per sample was obtained based on total qualified sequences (without singletons) using the subsample function of Mothur (version v.1.30.1) (Schloss et al., 2011). The predominant phyla (Supplementary Figure S2 and Table S3) of the PAB communities were affiliated with Cyanobacteria (73.4 ± 12.2%), Proteobacteria (17.7 ± 8.8%) and Bacteroidetes (5.3 ± 2.6 %), whereas Actinobacteria (36.1 ± 5.9%), Proteobacteria (26 ± 3.9%), and Bacteroidetes (21.3 ± 6.4%) were the most abundant phyla in the FLB communities. M. aeruginosa was the dominant species in PAB (Supplementary Table S3) and accounted for 92.97–99.04% of Cyanobacteria, indicating that M. aeruginosa was the bloom-causing species.
A comparison of PAB and FLB composition based on algae-free data (Supplementary Table S2-Resampling 2) was performed to investigate the difference in bacterial community without cyanobacteria. As shown in Figure 2A, the predominant PAB phyla were affiliated with Proteobacteria (65.88 ± 6.3%) and Bacteroidetes (21.24 ± 6.9%), whereas Proteobacteria (26.77 ± 4.3%), Actinobacteria (37.43 ± 5.7%) and Bacteroidetes (21.8 ± 6.5%) were the most abundant FLB phyla. The relative abundance of 14 phyla (43.75%) significantly differed in the two bacterial communities, as shown in Supplementary Table S4. Down to the genus level, three uncultured genera from the phylogenetic cluster I-10 (9.28 ± 2.4%, belongs to Rhodospirillales), Alcaligenaceae (8.68 ± 2.0%) and Cytophagaceae (6.47 ± 2.2%) were the most dominant genera in the PAB community, whereas hgcI clade, CL500-29 marine group and uncultured OPB56 from Chlorobiales dominated the FLB community (Figure 2B). The physiological characteristics were unclear for these most dominant taxa in PAB and FLB; e.g., the HgcI cluster is only known to be a widespread and typical bacteria group in freshwater (Frank et al., 2000).
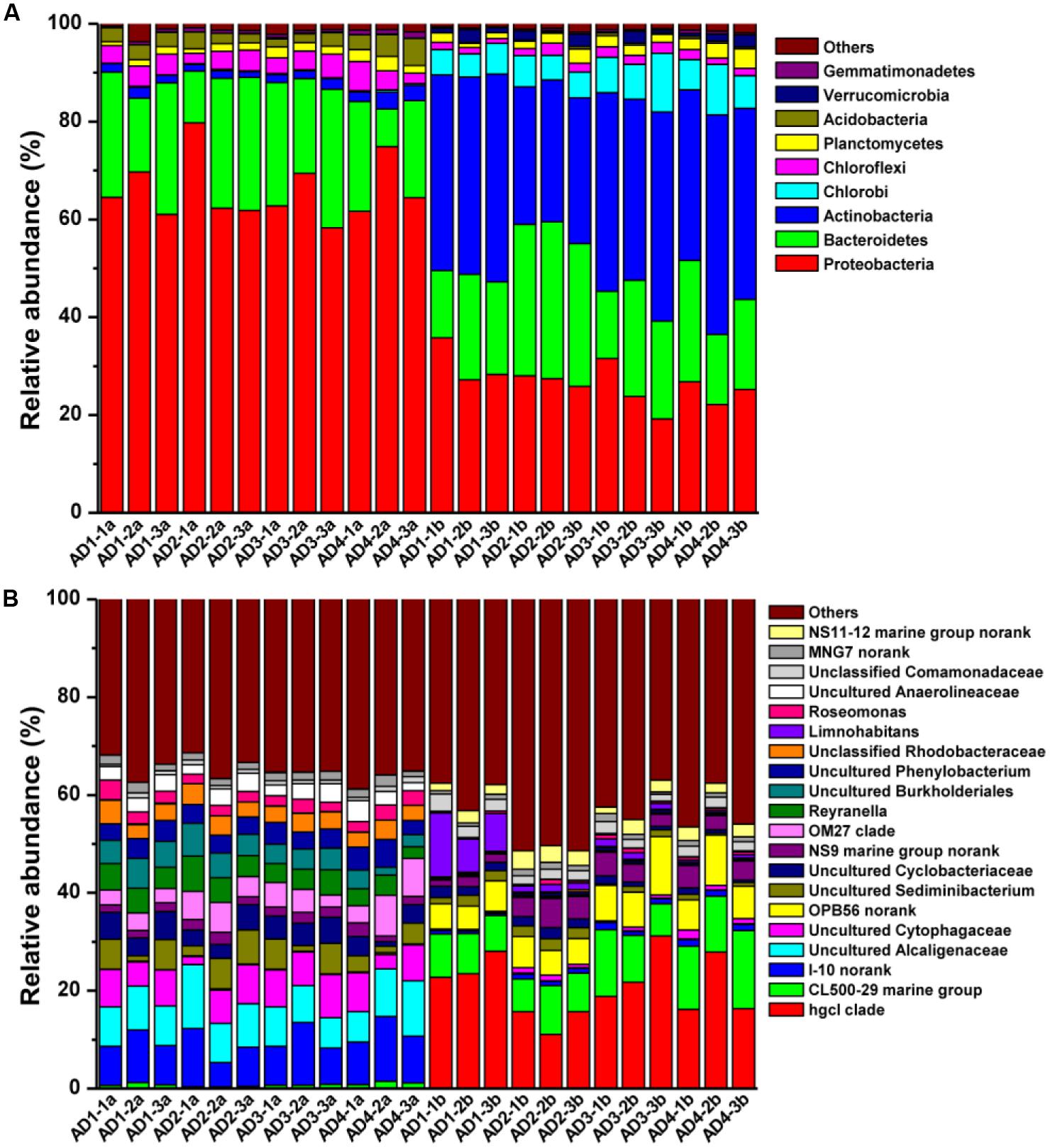
FIGURE 2. Distribution of bacterial taxa at phylum (A) and genus (B) levels based on sequence data without cyanobacteria (Supplementary Table S2-resample 2). Sample names with a and b represent PAB and FLB, respectively.
Compared to the FLB community, 76 (19.54%) and 103 (26.48%) taxa at the genus level had a significantly higher and lower (p < 0.05) relative abundance in PAB, respectively. Most of these taxa could not be assigned to known genera, other than the 86 genera shown in Supplementary Table S5. Additionally, the Shannon diversities of FLB communities were significantly higher (T-test, p < 0.01) than those of PAB, according to the datasets with or without Cyanobacteria (FLB: 4.87 ± 0.24/4.72 ± 0.23; PAB: 1.85 ± 0.62/4.25 ± 0.16; Shannon diversity with / without Cyanobacteria).
Differences in Whole Community Structure between PAB and FLB
Dissimilarity tests based on both MRPP (δ = 0.23, P < 0.01) and Adonis (R2 = 0.869, P < 0.01) showed significant differences between the PAB and FLB communities. The PCA plots suggested that the whole community structure of PAB was markedly different from that of FLB (Figure 3A), and this pattern was consistent even when the cyanobacterial sequences, which were much more abundant in PAB communities, were removed (Figure 3B).
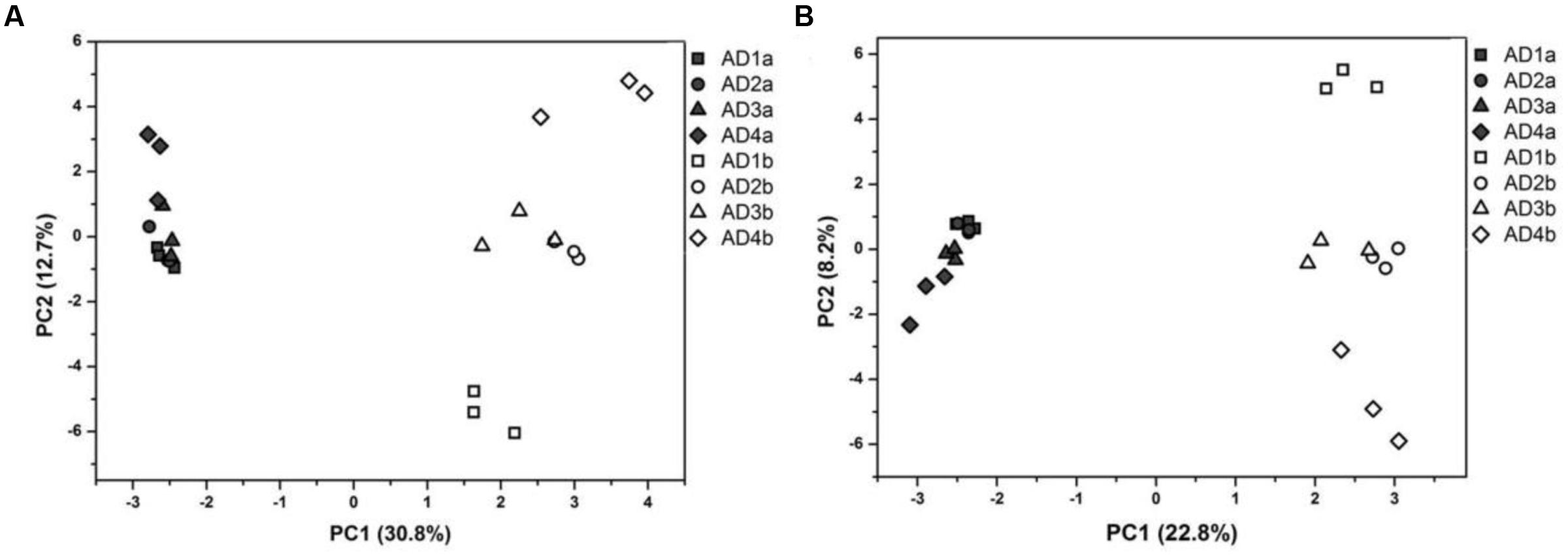
FIGURE 3. Principal component analyses of total bacteria (A) and bacteria without cyanobacteria (B). Filled symbols represent PAB, and hollow symbols represent FLB, respectively.
Relationship between Bacterial Communities and Environmental Properties
In this study, 13 environmental variables were used to test their correlation with the two communities (PAB and FLB) (Table 2). The results showed that no detected variable had significant (P < 0.05) effects on the microbial PAB communities, whereas six environmental variables, including pH, NO3-N, NO2-N, EI, bacterial density and algae density, significantly correlated with the FLB communities (P-values were less than 0.05 based on both Bray-Curtis and Euclidean distances). Because FLB communities were greatly correlated with these environmental properties, CCA was further performed to link the variation of FLB communities to these seven selected variables. The CCA ordination plot revealed that the first two canonical axes explained 36.9 and 17.5% of the constrained variations, respectively (Figure 4A). Samples in AD1 were characterized by higher algal density, NO3-N, NO2-N, EI and bacterial density, and these variables showed positive correlations with each other. AD3 and AD4 were characterized by higher pH. Additionally, samples in AD2 had weak relevance with these environmental properties but had higher Tur, SP, NH4-N, and TN that were not counted in CCA. Because of the importance of nutrient properties (i.e., NO3-N, NO2-N, EI), algae and pH in this CCA result, the VPA based on partial CCA was subsequently conducted using these selected variables. VPA revealed that nutrient properties, algae and pH and their interactions explained 26.2, 7.2, 4.2, and 29.2% of the variation in the FLB communities, respectively, leaving 33.1% unexplained (Figure 4B).
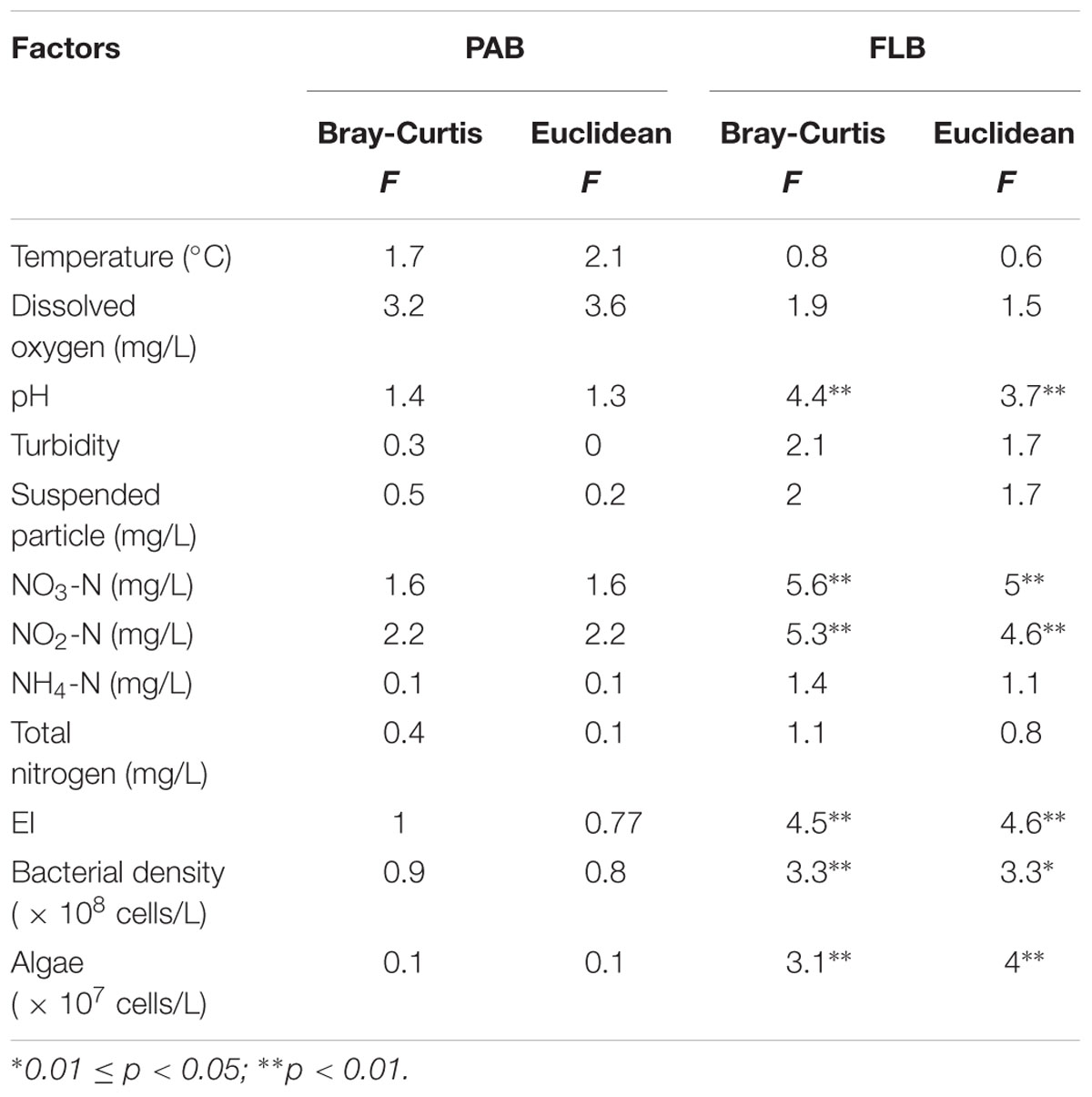
TABLE 2. Correlation between factors and bacterial communities based on Adonis (permutational multivariate analysis of variance using distance matrices).
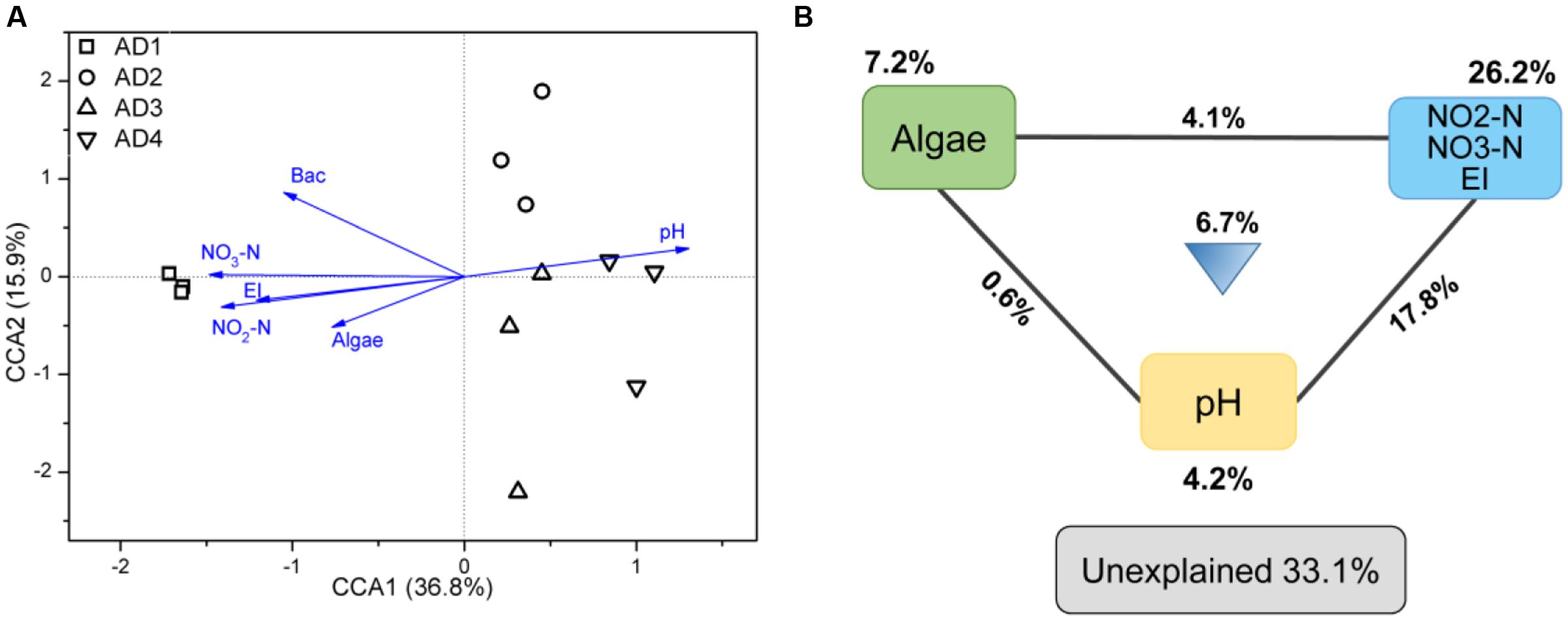
FIGURE 4. Canonical correspondence analysis (CCA) of FLB with selected environmental variables (A) and partial CCA-based variation partitioning analysis (VPA) (B). Selected environmental variables include pH, nitrate nitrogen (NO3-N), nitrite nitrogen (NO2-N), bacterial density (Bac), algae, eutrophication index (EI). Each diagram represents biological variation partitioned into the relative effects of each factor or a combination of factors.
Distinct Characteristics and of PAB and FLB Networks
In this study, a total of 913 and 1201 OTUs remained for PAB and FLB network construction, respectively (Figure 5 and Table 3), after removing OTUs that did not contain sequence reads in either the PAB or the FLB communities. The sizes (i.e., number of node) of these two networks differed (PAB: 222 and FLB: 480), with only shared 130 nodes (22.7%). Overall, nodes tended to be positively correlated (red lines) rather than negatively correlated (green lines), and positive correlations accounted for 88.2 and 88.8% in the PAB and FLB networks, respectively. The overall topologies (Table 3) revealed a more complex PAB network, with a higher average connectivity (avgK) and significantly tighter connections between nodes with a smaller average path length (GD, p < 0.01). Similar hierarchical properties in PAB and FLB networks were observed, with a close average clustering coefficients (avgCC).
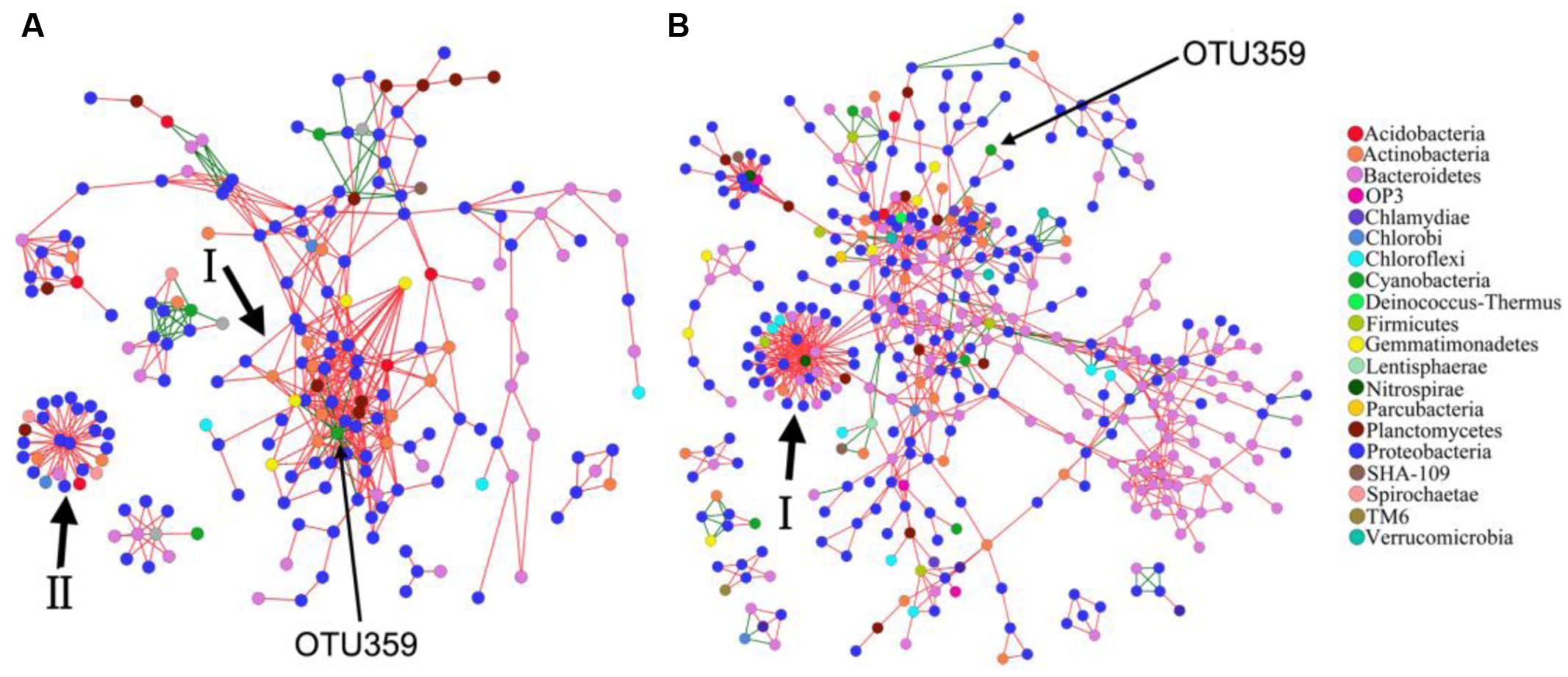
FIGURE 5. Network interaction of OTUs in PAB (A) and FLB (B). Node of bloom-causing M. aeruginosa OTU 359 is marked with long arrows, and major modules are marked with short arrows and Roman numerals. Red lines indicate positive interactions, and green lines indicate negative interactions.
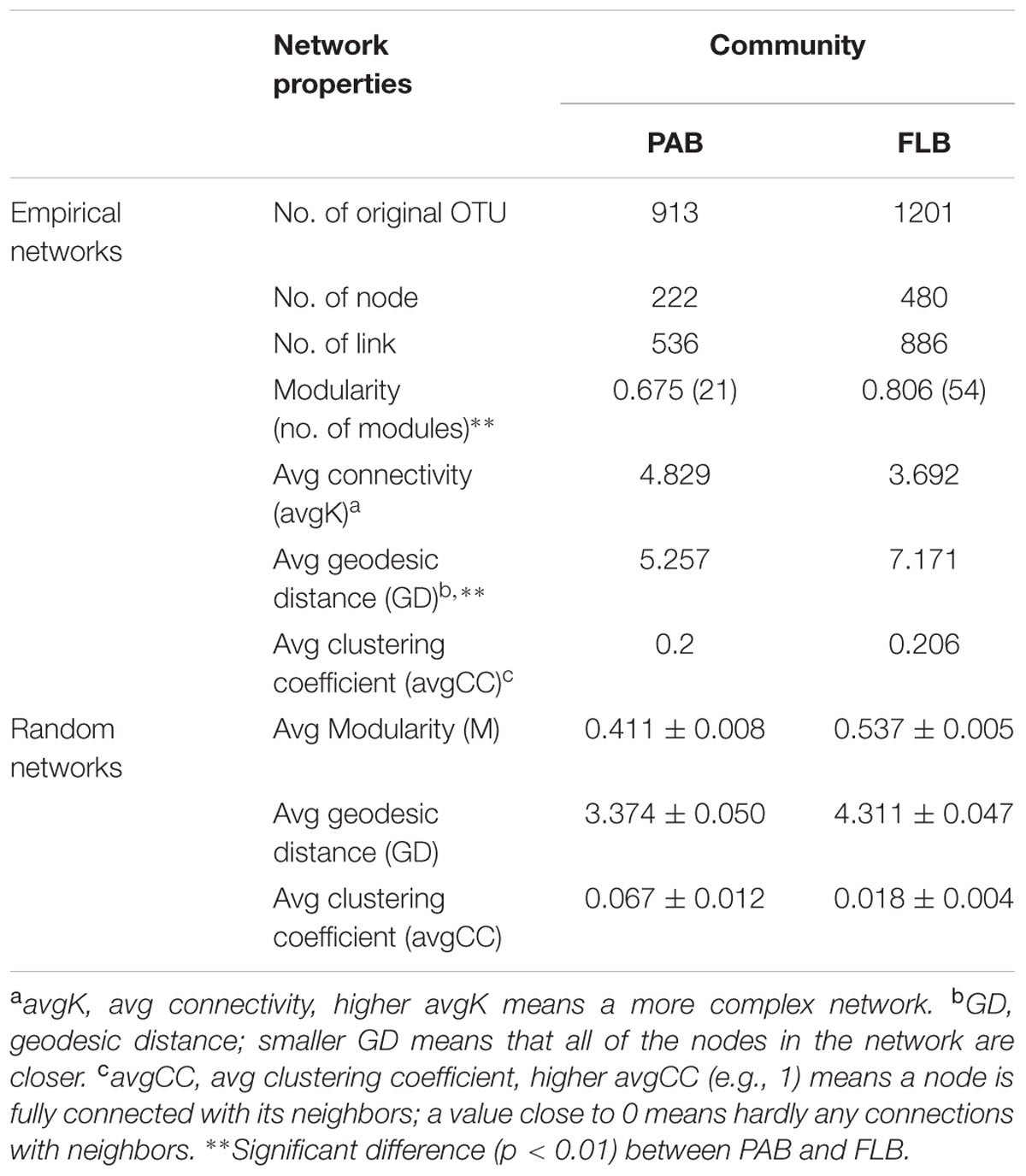
TABLE 3. Major topological properties of empirical and random MENs of PAB and FLB.
For the PAB network (Figure 5A), a total of 21 modules were generated, and the nodes were affiliated with 12 phyla, mainly from Proteobacteria (60.4%), Bacteroidetes (14.9%) and Actinobacteria (7.7%). The 222 nodes were connected by 536 links (i.e., 473 positive and 63 negative interactions). The Alphaproteobacteria was the largest group (21.6%) in the PAB network, and three module hubs (OTUs 1106- Meganema, 1109- Pleomorphomonas and 1156- Brevundimonas sp., Figures 6A,B) of the PAB network were all affiliated with Alphaproteobacteria (Table 4), with a Zi value larger than 2.5. Node OTU1156 had the highest connectivity in the PAB network, with 28 positively correlated nodes and 1 negatively correlated node (Figure 6A). There was no connector in the PAB network. Compared to the PAB network, the FLB network was much larger, with 54 modules (Figure 5B) and 480 nodes connected by 886 links (i.e., 787 positive and 99 negative interactions). The nodes were from 21 phyla and were dominated by Proteobacteria (46%), Bacteroidetes (30%) and Actinobacteria (6.9%). There were three connectors (Pi > 0.62) and seven module hubs (Zi > 2.5), as shown in Table 4. Module hub OTU38 (Bacteroidetes) had highest connectivity in the FLB network, with 36 positively correlated nodes (Figures 5B-I, 6D). For these module hubs and connectors, only five genera were identified (Table 4): Nitrospira, Fluviicola, CL500-29 marine group, Legionella, Crocinitomix.
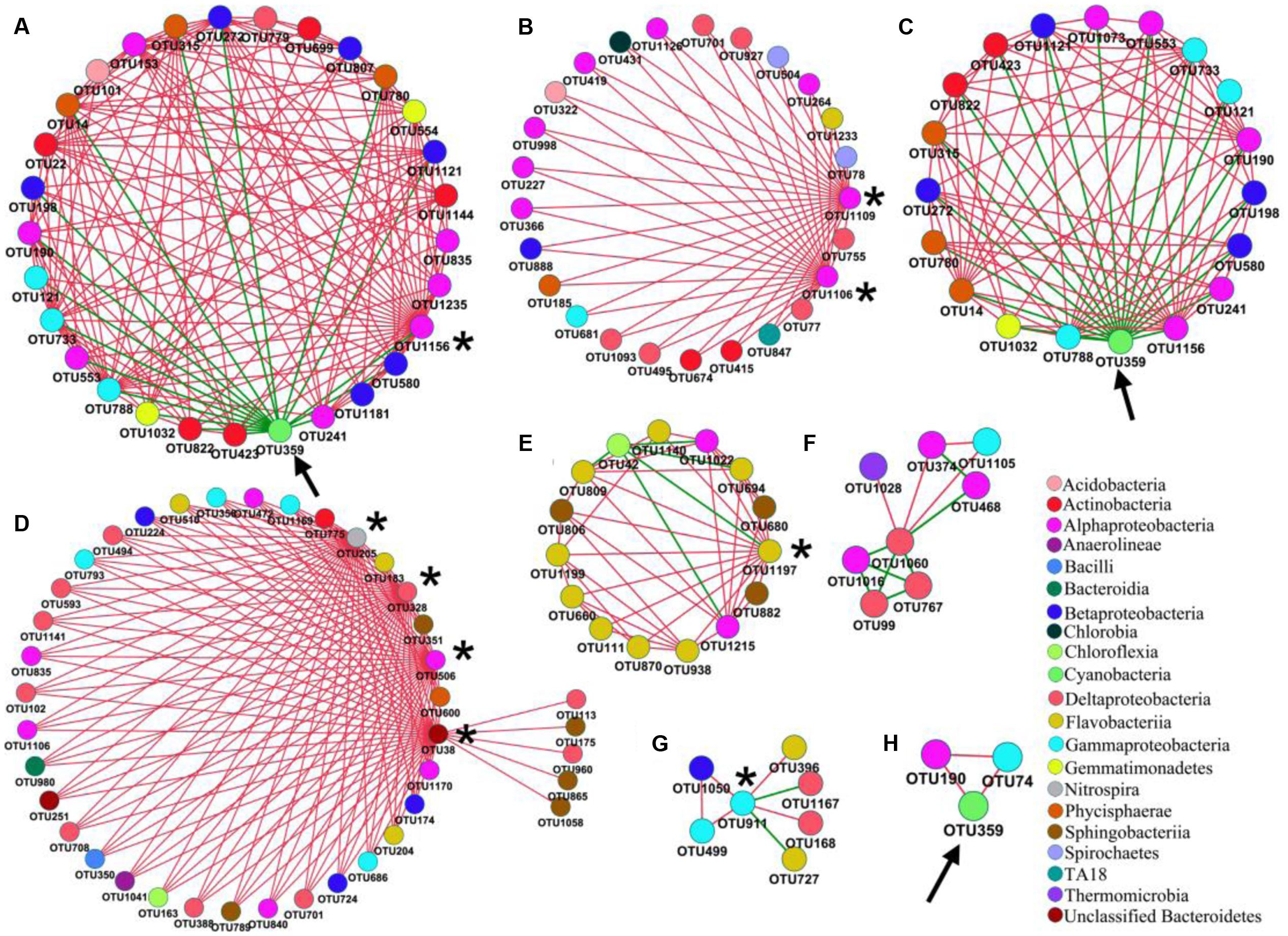
FIGURE 6. Network interactions between nodes of interest and their nearest neighbors in PAB (A–C) and FLB (D–H). Interactions of (A,B,D–G) are centered with module hubs (marked with asterisk). Interactions of (C,H) are centered with M. aeruginosa OTU 359 (marked with arrow).
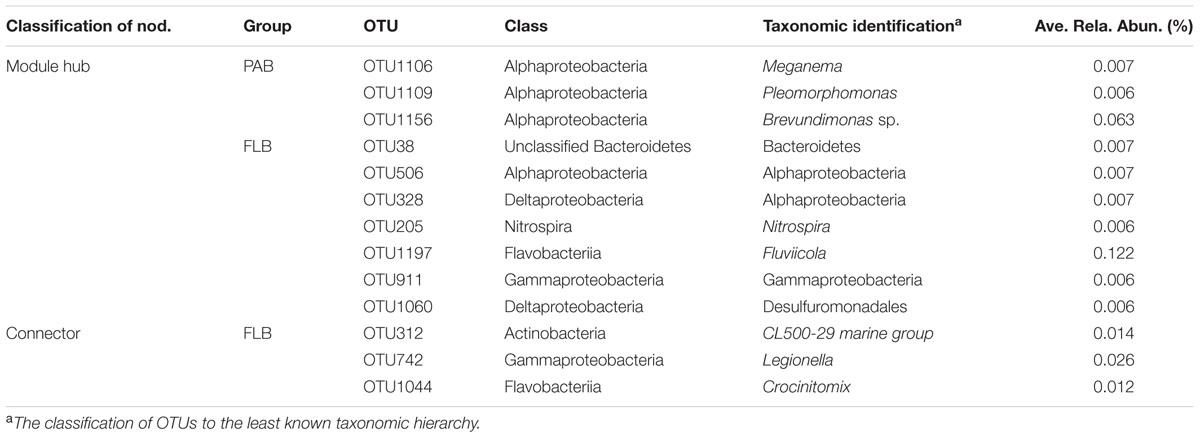
TABLE 4. Classification and relative abundance of module hub and connector nodes in PAB and FLB networks.
Network-Based Associations between Bacterial Species and M. aeruginosa
The network topology was used to explore the correlations among bloom-causing algae and algae-associated microorganisms. In our dataset, OTU359 was the bloom-causing M. aeruginosa species, with a relative abundance ranging from 48.9 to 86.1% in PAB (Supplementary Table S3) and a distinct, characteristic connection in the 2 networks, as shown in Figure 5 (marked with long arrows) and Figures 6E,H (marked with arrows). In the PAB network, OTU359 had only negative connections and among its 18 neighbor nodes (Table 5), only 8 genera were identified: SM1A02 (Phycisphaerae), Brevundimonas, Silanimonas, CL500-29 marine group (Acidimicrobiales), Gemmatimonas, Phenylobacterium, Reyranella, and Tepidicella. In contrast, the OTU 359 connected with only 2 nodes (OTU190 and OTU74) with positive links (Figure 6 and Table 5) in FLB network, and one was identified as a genus of Rheinheimera.
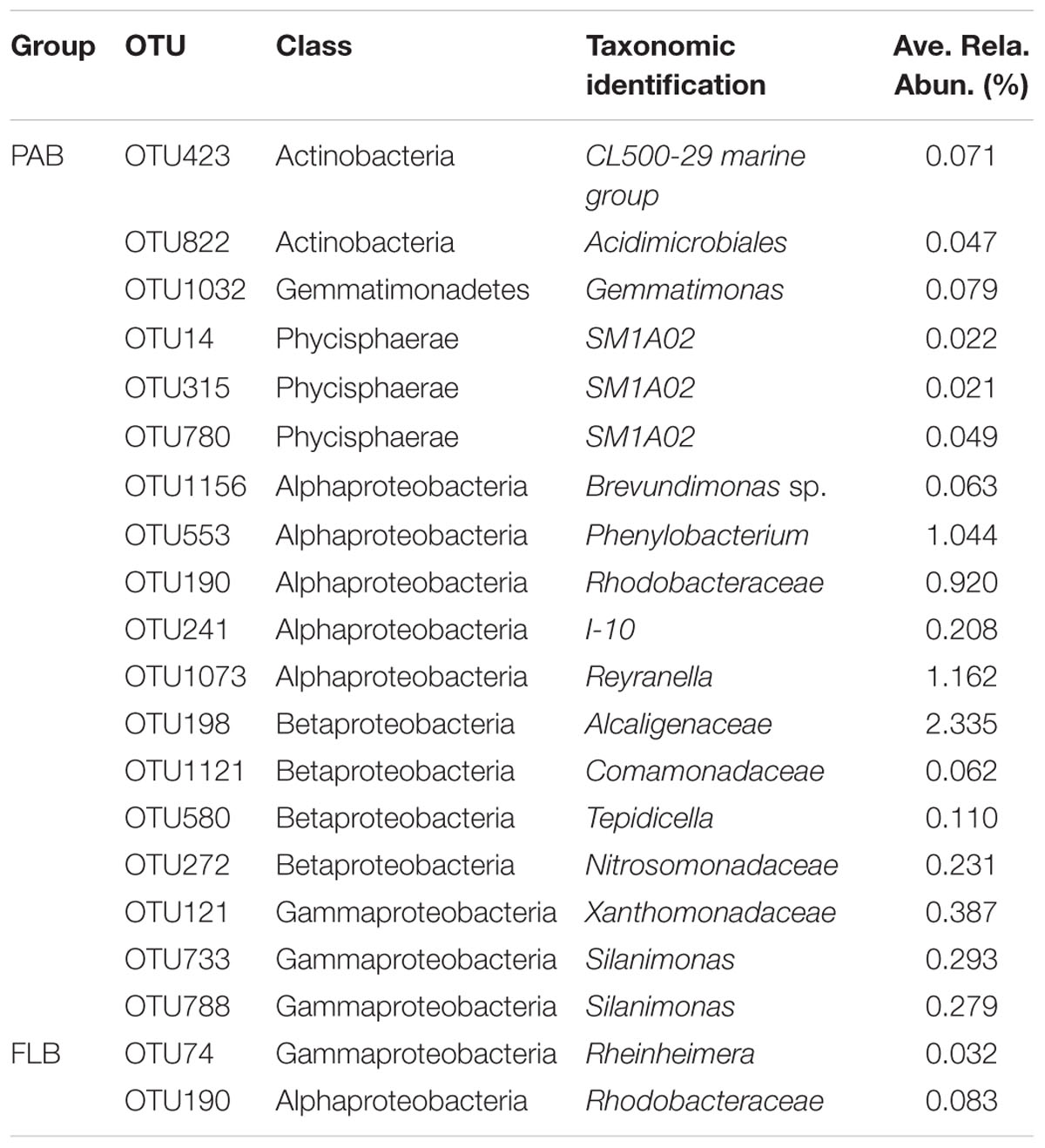
TABLE 5. Classification and relative abundance of nodes connected to OTU359 (bloom-causing M. aeruginosa).
Discussion
In this study, we found significant differences between PAB and FLB communities in terms of diversity, taxonomic composition and microbial network interactions. The greater difference observed between fractions (i.e., PAB vs. FLB) rather than sites was consistent with previous bacterial community studies during a diatom bloom (Riemann and Winding, 2001; Rieck et al., 2015). Although PAB community structure was quite conserved across the environmental changes in surrounding water, the FLB community was significantly correlated with the measured environmental variables (Table 2).
Cyanobacteria release considerable amounts of organic matters to surroundings and these organic substrates can attract a lot of heterotrophic bacteria into the phycosphere (Cai et al., 2013a). Because the compositions of these exudates from different algal species are distinct (Sapp et al., 2007; Paver et al., 2013), the divergent availability and physicochemical characteristics of these substrates result in different bacterial communities in the specific algal phycosphere (Louati et al., 2015). Only bacteria from the aquatic system adapting to this microenvironment live and persist in the phycosphere. When M. aeruginosa dominated the phytoplankton of Dianchi Lake, phycosphere selection theory could well explain the stability and lower diversity of PAB in this conserved phycosphere (Louati et al., 2015). The regional water environment fluctuated in different sites, including organic and inorganic matter (Table 1). This could lead to significant differences in FLB communities from the four sites.
For those genera whose relative abundance significantly changed between PAB and FLB communities, their most likely ecological physiological functions were investigated based on taxonomy according to the literature. The relative abundance of bacteria affiliating with nitrogen-fixing taxa Candidatus Competibacter (Mcilroy et al., 2014), Pleomorphomonas (Xie and Yokota, 2005; Im et al., 2006) and Bradyrhizobium (Kaneko et al., 2002) significantly increased in the PAB community, while the abundance of a potential nitrogen fixer affiliating with Paenibacillus (von der Weid et al., 2002) significantly increased in FLB community. Inorganic N is essential and beneficial for cyanobacterial growth; also algal density was found significantly positive correlations with the concentrations of NO2-N and NO3-N in this study. In our previous study, the nifH gene (nitrogen fixation) intensity was also found increased when algal bloom of Akashiwo sanguinea occurred (Yang et al., 2016). Nitrogen-fixing microorganisms and algae can form symbiotic relationships, e.g., azotobacteria provided cyanobacteria with inorganic N in exchange for organic exudates from algae (Lili et al., 2014) and Xu et al. (2016) found the cell density of Bradyrhizobium japonicum significantly increased when co-cultured with transgenic Chlamydomonas reinhardtii hemHc-lbac. This mutually beneficial relationship may have resulted in a significantly higher relative abundance of nitrogen-fixing bacteria in the PAB community than in FLB.
For the bacteria known for methane-utilizing, methanotrophy genera, the relative abundance of bacteria affiliating with methanotrophy genera Methylomonas (Koh et al., 1993), LD28 freshwater group (Salcher et al., 2015) and Candidatus Methylacidiphilum (Khadem et al., 2012), and methanotrophic genus Methylocaldum (Murrell, 2010) significantly increased in the FLB community, while the abundance of one filamentous methane oxidizer affiliating with Crenothrix (Stoecker et al., 2006) significantly increased in the PAB community. The relative abundance of some potentially algicidal bacteria of M. aeruginosa also significantly changed, e.g., Aquimonas (Cui et al., 2012) was more abundant in PAB community, and Pedobacter (Yang et al., 2012) and Aeromonas (Liu et al., 2012) were more so in FLB community. For potentially organic matter-degrading bacteria, Flavobacterium (O’Reilly and Crawford, 1989), Novosphingobium (Tiirola et al., 2002), Sphingopyxis (Xu et al., 2015), their relative abundances significantly increased in FLB community, while Labrys (Carvalho et al., 2005) was more abundant in PAB community. Many strains from Sphingopyxis have microcystin-degrading ability (Shimizu et al., 2012; Xu et al., 2015). As a water-soluble compound, microcystin released by Microcystis can easily diffuse into surrounding water and could be utilized by microcystin-degraders, e.g., Sphingopyxis. In PAB community, the relative abundance of some potentially methanol-utilizing [i.e., Phreatobacter (Tóth et al., 2014)] and ferrous iron-oxidizing genera [i.e., Ferroviario (Dubinina and Sorokina, 2014)] also significantly increased. In FLB community, the relative abundance of genera known for algal-derived DOC-utilizing (e.g., Limnohabitans and Limnobacter), eutrophic water-affiliating [e.g., CL500-29 marine group and hgcI clade (Lindh et al., 2015)] and pathogenic microorganism [e.g., Flavobacterium (Nicolas et al., 2008), Legionella (Hägele et al., 2000), Mycobacterium (Cosma et al., 2003)] significantly increased. The physiology of many other genera remains unclear (e.g., Hirschia, Candidatus Trichorickettsia and so on).
Ecosystems are very complex, with complicated networks that form between considerably interacting organisms (Montoya et al., 2006). In this study, a network-based approach using RMT was used to show the co-occurrence pattern and delineated interactions of bacterial communities during cyanobacterial blooms. The specific microbial interaction networks of PAB and FLB were established, and the role of cyanobacteria (especially for M. aeruginosa) in the network was proposed.
The stability of an ecosystem function lies in species interactions (Zhou et al., 2010). Lower connectivity in a community leads to higher functional stability of the system (e.g., scale-free network) because the whole network module will be less affected by the loss of nodes. (Zhou et al., 2010). For PAB, the higher connectively (avgK), highly concentrated modularity and fewer nodes could result in greater vulnerability than that of FLB. For FLB, the lower connectivity (avgK), higher network size and higher bacterial diversity could result in greater resilience to disturbances than that of PAB. The differences in topological features (e.g., connectivity and geodesic distance) of PAB and FLB networks could be associated with the spatial distances among bacteria, i.e., bacteria on particles are overall closer to each other than FLB. Accordingly, direct interactions, such as mutualism, competition or predation are more likely to occur in the PAB community. Taxa tended to covary in both PAB and FLB networks with predominantly positive correlations (>88%), indicting similarly preferred conditions or cooperative behaviors (e.g., cross feeding, syntrophic interactions and mutualistic interactions) of most bacteria during M. aeruginosa bloom (Raes and Bork, 2008).
In an ecological network, module hubs and connectors play crucial roles in the maintenance of community stability and are considered keystone species in the system (Zhou et al., 2010; Shi et al., 2016). The putative keystone nodes in PAB and FLB networks were lowly abundant, which indicates that low abundance taxa may play important roles in maintaining network structures in freshwater microbial communities. This finding is consistent with studies of soil microbial communities (Lupatini et al., 2014; Shi et al., 2016). Alphaproteobacteria was the most important group in the PAB network and occupied the three module centers: Meganema, Pleomorphomonas and Brevundimonas sp.. For the Meganema genus, M. perideroedes is the sole described species, which was found to be a versatile facultative anaerobe in activated sludge with ability to utilize various organic substrates in aerobic conditions and use nitrate and nitrite as electron acceptors (Kragelund et al., 2005), and McIlroy et al. (2015) thought it is a potentially facultative methylotrophy based on the genome of stain Gr1; for Pleomorphomonas, almost all the species of this genus are N-fixing bacteria, i.e., P. diazotrophica, P. diazotrophica, and P. oryzae (Xie and Yokota, 2005; Im et al., 2006; Madhaiyan et al., 2013), and Pleomorphomonas sp. were found can methylate Hg (II) (Achá et al., 2012); Brevundimonas sp. have been found the abilities of algicidal effect on M. aeruginosa, growth promotion on Chlorella ellipsoide, cadmium biosorption, lactofen-hydrolyzing, quinoline-degrading and antimicrobial silver nanoparticles producing (Park et al., 2008; Masoudzadeh et al., 2011; Rajamanickam et al., 2013; Wang C. et al., 2015).
For the five identified genera of module hubs and connectors in FLB network, Nitrospira are known as the most diverse and widespread nitrifiers in various ecosystems, e.g., natural environments and biological wastewater treatment plants (Lücker et al., 2010; Feng et al., 2016; Pinto et al., 2016). Ecological function or relationships with algal blooms of the remaining four genera are unclear except for the following information: for Fluviicola, only novel species descriptions have been reported and they were mainly isolated from fresh water and chemical industrial wastewater (O’Sullivan et al., 2005; Yang et al., 2014); CL500-29 marine group is widespread in waters, particularly in copiotrophic environments (Lindh et al., 2015); Legionella are wide-spread pathogens of human, animals, protozoa and aquatic eukaryotes (Golovlev, 2000; Hägele et al., 2000), and most species are aquatic origin; C. catalasitica is the sole species of Crocinitomix, which was isolated from algal-rich littoral zone of Auke Bay, AK, United States (Bowman et al., 2003). In general, module hubs of both networks were potentially involved in biogeochemical cycling of nitrogen taxa. This finding indicates that bacteria with ecological physiological functions of N metabolism might be quite important in the two communities. Moreover, module I of PAB network was centered with a potentially algicidal bacteria of M. aeruginosa (OTU1156, Brevundimonas), which indicates that algicidal bacteria might have important roles in PAB communities.
As the dominant phytoplankton phylum in Dianchi Lake, cyanobacteria (green nodes in Figure 5) were mostly connected with negative links (especially for the node OTU359), representing antagonistic relationships such as predator-prey, competition or variable fitness (Chow et al., 2014). As the photosynthetic organism, phytoplankton is often the most important biotic importer of organic substrates (e.g., DOM) in water systems. Cyanobacteria could act as a nutrient contributor by supplying heterotrophic bacteria with organic substrates from photosynthesis or by acting as prey (Paver et al., 2013).
The cyanobacteria node OTU359 represents the bloom-causing species M. aeruginosa, which is the most common colonial cyanobacteria in fresh water. As an oxygenic and photoautotrophic species, M. aeruginosa often form blooms in eutrophic waters and produce toxic microcystins. OTU359 was the only node with purely negative connections in module I, indicating its uniquely antagonistic relationships with neighboring nodes. It had 18 neighboring nodes in the PAB network, including the potentially algicial bacteria OTU1156 (Brevundimonas), which was the hub of module I. Among other neighbors of OTU359, OTU423-CL500-29 marine group are widespreadly aquatic bacteria; OTU1032-Gemmatimonas strains have photosynthetic pigments [i.e., bacteriochlorophyll a (Zeng et al., 2015) and carotenoids (Takaichi et al., 2009)] and polyphosphate-accumulating functions (Zhang et al., 2003). It is difficult to understand the connection of other neighbors with OTU359 because their ecophysiology is unclear (e.g., SM1A02, I-10) or appears irrelevant to cyanobacteria [e.g., Phenylobacterium; the physiology of reported strains are mainly involved in organic contaminant biodegradation (Tiago et al., 2005; Eberspächer and Lingens, 2006)]. The relationships between OTU359 and the related two neighbors (OTU74 and OTU 190) in the FLB network were also unclear.
The bloom-causing species M. aeruginosa (OTU359) was assumed to play different roles in PAB and FLB communities according to its network topological characteristics. Although node OTU 359 has few connections (two links) within the major module of the FLB network, M. aeruginosa seems more important because it connected to the hub of the largest module (module I) and occupied a higher hierarchical position with the 18 links in the PAB network. Therefore, M. aeruginosa possibly played a more important role in PAB communities than that of FLB communities. Additionally, the tight connections between nodes in the PAB network could indicate quick and effective metabolic interactions (Zhou et al., 2010) among species by transferring considerable organic matter from algae quickly in this phycosphere, thus indicating their potentially important roles in material and energy flows in this aquatic ecosystem.
In general, the communities of PAB and FLB during a M. aeruginosa bloom were significantly different in their structure, diversity, microbial interaction network and the topological role of M. aeruginosa. For PAB network, bacterial connections were tighter, potentially indicating more efficient metabolism interactions than that of FLB network; the higher importance of M. aeruginosa (OTU359) was indicated by high connectivity in the core region of the PAB network; Alphaproteobacteria were predominant, among which potentially nitrogen-fixing and algicidal bacteria occupied the module centers. FLB communities had significantly higher diversity and were sensitive to the detected environmental factors; genera related to methane-utilizing and pathogenic bacteria had significantly higher relative abundance; M. aeruginosa was marginal in the network, with less connectivity and more modules and nodes, possibly leading to the higher resilience of the ecological function than that of PAB. In both PAB and FLB interaction networks, a co-occurrence pattern was predominant, with positive connections. Bacteria affiliated with N metabolism were important by occupying the module hubs. Here, we have made an important attempt to explore the differential response of PAB and FLB communities to a cyanobacterial bloom by applying network-based analyses. Our findings provide ecological insights into the different interactions among PAB or FLB communities during algal blooms.
Author Contributions
YI and FL designed the research and revised the manuscript. CY, QW, PS, JL, and JK analyzed the data and drafted the manuscript. CY, QW, PS, JL, LL, XD, XZ, and XP contributed to the sampling and experiments. LL, XD, XZ, and XP revised the manuscript. All authors approved the final manuscript and agreed to be responsible for all aspects of the work.
Funding
This work was financed by the China Post-doctoral Science Foundation (grant No: 2015M582498) to CY; Fundamental Research Funds for the Central Universities (grant No: XDJK2016C084 and XDJK2011B009) to CY and FL, respectively; National Natural Science Foundation of China (grant No: 31600095 and 41301263) to CY and XD, respectively; Chongqing Post-doctoral Science Foundation (grant No: Xm2016082), Chongqing Research Program of Advanced Technology and Application Foundation (grant No: cstc2016jcyjA0525) to CY and Research Funds of Scientific Platform and Base Construction (grant No: cstc2014pt-sy0017) to FL. These funders supported the related work, including study design, sampling, experiments, and writing.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
We greatly thank Dr. Bo Chen, Xiaodong Ma, and Huan He of Kunming University of Science and Technology, for their helping during sampling.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2017.01202/full#supplementary-material
FIGURE S1 | The number of identified taxa at different levels based on total sequences (a), the Venn diagrams (b) for taxa number of PAB (gray-tinted cycles with dashed) and FLB (white cycles with solid line).
FIGURE S2 | Distribution of bacterial taxa at phylum level based on sequence data of Resample 1. Sample names with a and b represent the PAB and FLB, respectively.
Footnotes
References
Achá, D., Pabón, C. A., and Hintelmann, H. (2012). Mercury methylation and hydrogen sulfide production among unexpected strains isolated from periphyton of two macrophytes of the Amazon. FEMS Microbiol. Ecol. 80, 637–645. doi: 10.1111/j.1574-6941.2012.01333.x
Amato, K. R., Yeoman, C. J., Kent, A., Righini, N., Carbonero, F., Estrada, A., et al. (2013). Habitat degradation impacts black howler monkey (Alouatta pigra) gastrointestinal microbiomes. ISME J. 7, 1344–1353. doi: 10.1038/ismej.2013.16
Berg, K. A., Lyra, C., Sivonen, K., Paulin, L., Suomalainen, S., Tuomi, P., et al. (2009). High diversity of cultivable heterotrophic bacteria in association with cyanobacterial water blooms. ISME J. 3, 314–325. doi: 10.1038/ismej.2008.110
Bowman, J. P., Nichols, C. M., and Gibson, J. A. (2003). Algoriphagus ratkowskyi gen. nov., sp. nov., Brumimicrobium glaciale gen. nov., sp. nov., Cryomorpha ignava gen. nov., sp. nov. and Crocinitomix catalasitica gen. nov., sp. nov., novel flavobacteria isolated from various polar habitats. Int. J. Syst. Evol. Microbiol. 53, 1343–1355. doi: 10.1099/ijs.0.02553-0
Cai, H., Jiang, H., Krumholz, L. R., and Yang, Z. (2013a). Bacterial community composition of size-fractioned aggregates within the phycosphere of cyanobacterial blooms in a eutrophic freshwater lake. PLoS ONE 9:e102879. doi: 10.1371/journal.pone.0102879
Cai, H., Yan, Z., Wang, A., Krumholz, L. R., and Jiang, H. (2013b). Analysis of the attached microbial community on mucilaginous cyanobacterial aggregates in the eutrophic lake Taihu reveals the importance of Planctomycetes. Microb. Ecol. 66, 73–83. doi: 10.1007/s00248-013-0224-1
Cao, X., Zhou, Y., Wang, Z., and Song, C. (2015). The contribution of attached bacteria to microcystis bloom: evidence from field investigation and microcosm experiment. Geomicrobiology 33, doi: 10.1080/01490451.2015.1074319
Carvalho, M. F., Ferreira, J. R., Pacheco, C. C., De, M. P., and Castro, P. M. (2005). Isolation and properties of a pure bacterial strain capable of fluorobenzene degradation as sole carbon and energy source. Environ. Microbiol. 7, 294–298. doi: 10.1111/j.1462-2920.2004.00714.x
Chao, A. (1984). Non-parametric estimation of the classes in a population. Scand. J. Stat. 11, 265–270.
Chen, L. Q., and Hu, Y. (2011). “Influencing factors on determination of ammonia nitrogen in water by nessler’s reagent spectrophotometry,” in Proceedings of the 2011 International Symposium on Water Resource and Environmental Protection (ISWREP) (Piscataway, NJ: IEEE), 1173–1176. doi: 10.1109/ISWREP.2011.5893223
Chow, C.-E. T., Kim, D. Y., Sachdeva, R., Caron, D. A., and Fuhrman, J. A. (2014). Top-down controls on bacterial community structure: microbial network analysis of bacteria, T4-like viruses and protists. ISME J. 8, 816–829. doi: 10.1038/ismej.2013.199
Clauset, A., Newman, M. E., and Moore, C. (2010). Finding community structure in very large networks. Phys. Rev. E 70(6 Pt 2), 264–277. doi: 10.1103/PhysRevE.70.066111
Cosma, C. L., Sherman, D. R., and Ramakrishnan, L. (2003). The secret lives of the pathogenic mycobacteria. Microbiology 57, 641–676. doi: 10.1146/annurev.micro.57.030502.091033
Cui, Y., Yong, X., Zhang, F., Yang, X., and Shen, Q. (2012). Isolation and identification of an algae-lysing bacterium and its algicidal effect. Chin. J. Appl. Environ. Biol. 5, 752–760. doi: 10.3724/SP.J.1145.2012.00752
Deng, Y., Jiang, Y. H., Yang, Y., He, Z., Luo, F., and Zhou, J. (2012). Molecular ecological network analyses. BMC Bioinformatics 13:113. doi: 10.1186/1471-2105-13-113
Deng, Y., Zhang, P., Qin, Y., Tu, Q., Yang, Y., He, Z., et al. (2015). Network succession reveals the importance of competition in response to emulsified vegetable oil amendment for uranium bioremediation. Environ. Microbiol. 18, 205–218. doi: 10.1111/1462-2920.12981
Dubinina, G. A., and Sorokina, A. Y. (2014). Neutrophilic lithotrophic iron-oxidizing prokaryotes and their role in the biogeochemical processes of the iron cycle. Microbiology 83, 1–14. doi: 10.1134/S0026261714020052
Eberspächer, J., and Lingens, F. (2006). The genus Phenylobacterium. Prokaryotes 5, 250–256. doi: 10.1007/0-387-30745-1_13
Esty, W. W. (1986). The efficiency of good’s nonparametric coverage estimator. Annal. Stat. 14, 1257–1260. doi: 10.1214/aos/1176350066
Feng, G., Sun, W., Zhang, F., Loganathan, K., and Li, Z. (2016). Inhabitancy of active Nitrosopumilus-like ammonia-oxidizing archaea and Nitrospiranitrite-oxidizing bacteria in the sponge Theonella swinhoei. Sci. Rep. 6:24966. doi: 10.1038/srep24966
Frank, O. G., Zaichikov, E., Belkova, N., Denissova, L., Pernthaler, J., Pernthaler, A., et al. (2000). Comparative 16S rRNA analysis of lake Bacterioplankton reveals globally distributed phylogenetic custers Including an abundant group of Actinobacteria. Appl. Environ. Microbiol. 66, 5053–5065. doi: 10.1128/AEM.66.11.5053-5065.2000
Golovlev, E. L. (2000). General and molecular ecology of Legionella. Microbiology 69, 1–6. doi: 10.1007/BF02757247
Grossart, H. P., Czub, G., and Simon, M. (2006). Algae–bacteria interactions and their effects on aggregation and organic matter flux in the sea. Environ. Microbiol. 8, 1074–1084. doi: 10.1111/j.1462-2920.2006.00999.x
Grossart, H. P., Levold, F., Allgaier, M., Simon, M., and Brinkhoff, T. (2005). Marine diatom species harbor distinct bacterial communities. Environ. Microbiol. 7, 860–873. doi: 10.1111/j.1462-2920.2005.00759.x
Guimerà, R., and Amaral, L. A. N. (2005). Functional cartography of complex metabolic networks. Nature 433, 895–900. doi: 10.1038/nature03288
Hägele, S., Köhler, R., Merkert, H., Schleicher, M., Hacker, J., and Steinert, M. (2000). Dictyostelium discoideum: a new host model system for intracellular pathogens of the genus Legionella. Cell Microbiol. 2, 165–171. doi: 10.1046/j.1462-5822.2000.00044.x
Hill, M. O. (1973). Diversity and evenness: a unifying notation and its consequences. Ecology 54, 427–432.f doi: 10.2307/1934352
Im, W. T., Kim, S. H., Kim, M. K., Ten, L. N., and Lee, S. T. (2006). Pleomorphomonas koreensis sp. nov., a nitrogen-fixing species in the order rhizobiales. Int. J. Syst. Evol. Microbiol. 56, 1663–1666. doi: 10.1099/ijs.0.63499-0
Jasti, S., Sieracki, M. E., Poulton, N. J., Giewat, M. W., and Rooneyvarga, J. N. (2005). Phylogenetic diversity and specificity of bacteria closely associated with Alexandrium spp. and other phytoplankton. Appl. Environ. Microbiol. 71, 3483–3494. doi: 10.1128/AEM.71.7.3483-3494.2005
Kaneko, T., Nakamura, Y., Sato, S., Minamisawa, K., Uchiumi, T., Sasamoto, S., et al. (2002). Complete genomic sequence of nitrogen-fixing symbiotic bacterium Bradyrhizobium japonicum USDA110. DNA Res. 9, 189–197. doi: 10.1093/dnares/9.6.189
Karner, M., and Herndl, G. J. (1992). Extracellular enzymatic activity and secondary production in free-living and marine-snow-associated bacteria. Mar. Biol. 113, 341–347. doi: 10.1007/BF00347289
Khadem, A. F., Pol, A., Wieczorek, A. S., Jetten, M. S. M., and Camp, H. J. M. O. D. (2012). Metabolic regulation of “Ca. Methylacidiphilum fumariolicum” SolV cells grown under different nitrogen and oxygen limitations. Front. Microbiol. 3:266. doi: 10.3389/fmicb.2012.00266
Koh, S. -C., Bowman, J. P., and Sayler, G. S. (1993). Soluble methane monooxygenase production and trichloroethylene degradation by a type I methanotroph, Methylomonas methanica 68-1. Appl. Environ. Microbiol. 59, 960–967.
Kragelund, C., Nielsen, J. L., Thomsen, T. R., and Nielsen, P. H. (2005). Ecophysiology of the filamentous Alphaproteobacterium Meganema perideroedes in activated sludge. FEMS Microbiol. Ecol. 54, 111–122. doi: 10.1016/j.femsec.2005.03.002
Kuang, J., Huang, L., He, Z., Chen, L., Hua, Z., Pu, J., et al. (2016). Predicting taxonomic and functional structure of microbial communities in acid mine drainage. ISME J. 10, 1527–1539. doi: 10.1038/ismej.2015.201
Lack, T. J. (1971). Quantitative studies on the phytoplankton of the rivers thames and kennet at reading. Freshw. Biol. 1, 213–224. doi: 10.1111/j.1365-2427.1971.tb01558.x
Li, X., Liu, Y., Song, L., and Liu, J. (2003). Responses of antioxidant systems in the hepatocytes of common carp (Cyprinus carpio L.) to the toxicity of microcystin-LR. Toxicon 42, 85–89. doi: 10.1016/S0041-0101(03)00104-1
Lili, X. U., Wang, Q., Shuangxiu, W. U., and Dezhi, L. I. (2014). Optimization of co-cultivation conditions of transgenic alga hemHc-lbac and Bradyrhizobium japonicum for hydrogen production. Ecol. Sci. 33, 106–112.
Lin, L., Ji, Y., Tu, Q., Huang, R., Lin, T., Zeng, X., et al. (2013). Microevolution from shock to adaptation revealed strategies improving ethanol tolerance and production in Thermoanaerobacter. Biotechnol. Biofuels 6:103. doi: 10.1186/1754-6834-6-103
Lin, L., Song, H., Tu, Q., Qin, Y., Zhou, A., Liu, W., et al. (2011). The Thermoanaerobacter glycobiome reveals mechanisms of pentose and hexose co-utilization in bacteria. PLoS Genet. 7:e1002318. doi: 10.1371/journal.pgen.1002318
Lindh, M. V., Lefébure, R., Degerman, R., Lundin, D., Andersson, A., and Pinhassi, J. (2015). Consequences of increased terrestrial dissolved organic matter and temperature on bacterioplankton community composition during a Baltic Sea mesocosm experiment. Ambio 44(Suppl. 3), 402–412. doi: 10.1007/s13280-015-0659-3
Liu, Y., Wang, M., Jia, R., and Li, L. (2012). Removal of cyanobacteria by an Aeromonas sp. Desalination Water Treatment 47, 205–210. doi: 10.3390/toxins8030055
Logares, R., Sunagawa, S., Salazar, G., Cornejo-Castillo, F. M., Ferrera, I., Sarmento, H., et al. (2014). Metagenomic 16S rDNA Illumina tags are a powerful alternative to amplicon sequencing to explore diversity and structure of microbial communities. Environ. Microbiol. 16, 2659–2671. doi: 10.1111/1462-2920.12250
Louati, I., Pascault, N., Debroas, D., Bernard, C., Humbert, J. F., and Leloup, J. (2015). Structural diversity of bacterial communities associated with bloom-forming freshwater cyanobacteria differs according to the cyanobacterial genus. PLoS ONE 10:e0140614. doi: 10.1371/journal.pone.0140614
Lücker, S., Wagner, M., Maixner, F., Pelletier, E., Koch, H., Vacherie, B., et al. (2010). A Nitrospira metagenome illuminates the physiology and evolution of globally important nitrite-oxidizing bacteria. Proc. Natl. Acad. Sci. U.S.A. 107, 13479–13484. doi: 10.1073/pnas.1003860107
Lund, J. W. G., Kipling, C., and Cren, E. D. L. (1958). The inverted microscope method of estimating algal numbers and the statistical basis for counting. Hydrobiologia 11, 143–170. doi: 10.1007/BF00007865
Lupatini, M., Suleiman, A. K. A., Jacques, R. J. S., Antoniolli, Z. I., Ferreira, A. O. D. S., Kuramae, E. E., et al. (2014). Network topology reveals high connectance levels and few key microbial genera within soils. Soil Process. 2, 1–11. doi: 10.3389/fenvs.2014.00010
Madhaiyan, M., Jin, T. Y., Roy, J. J., Kim, S. J., Weon, H. Y., Kwon, S. W., et al. (2013). Pleomorphomonas diazotrophica sp. nov., an endophytic N-fixing bacterium isolated from root tissue of Jatropha curcas L. Int. J. Syst. Evol. Microbiol. 63(Pt 7), 2477. doi: 10.1099/ijs.0.044461-0
Maixing, D., Xuehui, X., Shengping, T., Jianmin, Z., Zhuqing, F., and Zhou, Z. (2003). On a role of microorganism in precipitation of phosphorus Geology of Chemical Minerals (in Chinese). 25, 65–75.
Masoudzadeh, N., Zakeri, F., Bagheri Lotfabad, T., Sharafi, H., Masoomi, F., Zahiri, H. S., et al. (2011). Biosorption of cadmium by Brevundimonas sp. ZF12 strain, a novel biosorbent isolated from hot-spring waters in high background radiation areas. J. Hazard. Mater. 197, 190–198. doi: 10.1016/j.jhazmat.2011.09.075
Mcilroy, S. J., Albertsen, M., Andresen, E. K., Saunders, A. M., Kristiansen, R., Bjerregaard, M. S., et al. (2014). “Candidatus Competibacter”-lineage genomes retrieved from metagenomes reveal functional metabolic diversity. ISME J. 8, 613–624. doi: 10.1038/ismej.2013.162
McIlroy, S. J., Lapidus, A., Thomsen, T. R., Han, J., Haynes, M., Lobos, E., et al. (2015). High quality draft genome sequence of Meganema perideroedes str. Gr1T and a proposal for its reclassification to the family Meganemaceae fam. nov. Stand. Genomic Sci. 10:23. doi: 10.1186/s40793-015-0013-1
Mendiburu, F. D. (2014). Agricolae: Statistical Procedures for Agricultural Research. R Package Version 1.2-1. Available at: http://CRAN.R-project.org/package=agricolae
Montoya, J. M., Pimm, S. L., and Solé, R. V. (2006). Ecological networks and their fragility. Nature 442, 259–264. doi: 10.1038/nature04927
Murrell, J. C. (2010). “The aerobic methane oxidizing bacteria (Methanotrophs),” in Handbook of Hydrocarbon & Lipid Microbiology, ed. K. N. Timmis (Berlin: Springer Science & Business Media), 1953–1966. doi: 10.1007/978-3-540-77587-4_143
Nicolas, P., Mondot, S., Achaz, G., Bouchenot, C., Bernardet, J. F., and Duchaud, E. (2008). Population structure of the fish-pathogenic bacterium Flavobacterium psychrophilum. Appl. Environ. Microbiol. 74, 3702–3709. doi: 10.1128/AEM.00244-08
Oksanen, J., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., O’Hara, R. B., et al. (2013). Vegan: community ecology package. R package version 2.0-10. http://CRAN.R-project.org/package=vegan. J. Stat. Softw. 48, 1–21. doi: 10.1186/s12866-016-0748-x
O’Reilly, K. T., and Crawford, R. L. (1989). Degradation of pentachlorophenol by polyurethane-immobilized Flavobacterium cells. Appl. Environ. Microbiol. 55, 2113–2118.
O’Sullivan, L. A., Rinna, J., Humphreys, G., Weightman, A. J., and Fry, J. C. (2005). Fluviicola taffensis gen. nov., sp. nov., a novel freshwater bacterium of the family Cryomorphaceae in the phylum ’Bacteroidetes’. Int. J. Syst. Evol. Microbiol. 55, 2189–2194. doi: 10.1099/ijs.0.63736-0
Paerl, H. W., and Otten, T. G. (2013). Harmful cyanobacterial blooms: causes, consequences, and controls. Microb. Ecol. 65, 995–1010. doi: 10.1007/s00248-012-0159-y
Park, K. W., Lee, K., Jung, S. E., and Choi, T. J. (2008). Growth promotion of Chlorella ellipsoidea by co-inoculation with Brevundimonas sp. isolated from the microalga. Hydrobiologia 598, 219–228. doi: 10.1007/s10750-007-9152-8
Paver, S. F., Hayek, K. R., Gano, K. A., Fagen, J. R., Brown, C. T., Davis-Richardson, A. G., et al. (2013). Interactions between specific phytoplankton and bacteria affect lake bacterial community succession. Environ. Microbiol. 15, 2489–2504. doi: 10.1111/1462-2920.12131
Pinto, A. J., Marcus, D. N., Ijaz, U. Z., Bautista-de lose Santos, Q. M., Dick, G. J., and Raskin, L. (2016). Metagenomic evidence for the presence of comammox Nitrospira-like bacteria in a drinking water system. mSphere 1, e54-15. doi: 10.1128/mSphere.00054-15
Porter, K. G., and Feig, Y. S. (1980). The use of DAPI for identifying and counting aquatic microflora1. Limnol. Oceanogr. 25, 943–948. doi: 10.4319/lo.1980.25.5.0943
R Core Team (2015). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Available at: http://www.R-project.org/
Raes, J., and Bork, P. (2008). Molecular eco-systems biology: towards an understanding of community function. Nat. Rev. Microbiol. 6, 693–699. doi: 10.1038/nrmicro1935
Rajamanickam, K., Sudha, S., Francis, M., Sowmya, T., Rengaramanujam, J., Sivalingam, P., et al. (2013). Microalgae associated Brevundimonas sp. MSK 4 as the nano particle synthesizing unit to produce antimicrobial silver nanoparticles. Spect. Acta Part A: Mol. Biomol. Spectros. 113, 10–14. doi: 10.1016/j.saa.2013.04.083
Rieck, A., Herlemann, D. P. R., Klaus, J., and Grossart, H.-P. (2015). Particle-associated differ from free-Living bacteria in surface waters of the Baltic Sea. Front. Microbiol. 6:1297. doi: 10.3389/fmicb.2015.01297
Riemann, L., and Winding, A. (2001). Community dynamics of free-living and particle-associated bacterial assemblages during a freshwater phytoplankton bloom. Microb. Ecol. 42, 274–285. doi: 10.1007/s00248-001-0018-8
Salcher, M. M., Neuenschwander, S. M., Posch, T., and Pernthaler, J. (2015). The ecology of pelagic freshwater methylotrophs assessed by a high-resolution monitoring and isolation campaign. ISME J. 9, 2442–2453. doi: 10.1038/ismej.2015.55
Sapp, M., Schwaderer, A. S., Wiltshire, K. H., Hoppe, H.-G., Gerdts, G., and Wichels, A. (2007). Species-specific bacterial communities in the phycosphere of microalgae? Microb. Ecol. 53, 683–699. doi: 10.1007/s00248-006-9162-5
Schloss, P. D., Gevers, D., and Westcott, S. L. (2011). Reducing the effects of PCR amplification and sequencing artifacts on 16S rRNA-based studies. PLoS ONE 6:e27310. doi: 10.1371/journal.pone.0027310
Shannon, P., Markiel, A., Ozier, O., Baliga, N. S., Wang, J. T., Ramage, D., et al. (2003). Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504. doi: 10.1101/gr.1239303
Shao, K., Lei, Z., Wang, Y., Xin, Y., Tang, X., Qin, B., et al. (2014). The responses of the taxa composition of particle-attached bacterial community to the decomposition of Microcystis blooms. Sci. Total Environ. 488-489(3 Suppl.), 236–242. doi: 10.1016/j.scitotenv.2014.04.101
Shi, S., Nuccio, E. E., Shi, Z. J., He, Z., Zhou, J., and Firestone, M. K. (2016). The interconnected rhizosphere: high network complexity dominates rhizosphere assemblages. Ecol. Lett. 19, 926–936. doi: 10.1111/ele.12630
Shimizu, K., Maseda, H., Okano, K., Kurashima, T., Kawauchi, Y., Qiang, X., et al. (2012). Enzymatic pathway for biodegrading microcystin LR in Sphingopyxis sp. C-1. J. Biosci. Bioeng. 114, 630–634. doi: 10.1016/j.jbiosc.2012.07.004
State Environmental Protection Administration of China. (1989). Monitoring and Analysis Method of Water and Wastewater, 3rd Edn. Beijing: China Environmental Science Press.
Stoecker, K., Bendinger, B., Schöning, B., Nielsen, P. H., Nielsen, J. L., Baranyi, C., et al. (2006). Cohn’s Crenothrix is a filamentous methane oxidizer with an unusual methane monooxygenase. Proc. Natl. Acad. Sci. U.S.A. 103, 2363–2367. doi: 10.1073/pnas.0506361103
Takaichi, S., Maoka, T., Takasaki, K., and Hanada, S. (2009). Carotenoids of Gemmatimonas aurantiaca (Gemmatimonadetes): identification of a novel carotenoid, deoxyoscillol 2-rhamnoside, and proposed biosynthetic pathway of oscillol 2,2’-dirhamnoside. Microbiology 156(Pt 3), 757–763. doi: 10.1099/mic.0.034249-0
Tang, X., Gao, G., Qin, B., Zhu, L., Chao, J., Wang, J., et al. (2009). Characterization of bacterial communities associated with organic aggregates in a large, shallow, eutrophic freshwater lake (Lake Taihu, China). Microb. Ecol. 58, 307–322. doi: 10.1007/s00248-008-9482-8
Tiago, I., Mendes, V., Pires, C., Morais, P. V., and Veríssimo, A. (2005). Phenylobacterium falsum sp. nov., an Alphaproteobacterium isolated from a nonsaline alkaline groundwater, and emended description of the genus Phenylobacterium. Syst. Appl. Microbiol. 28, 295–302. doi: 10.1016/j.syapm.2005.02.005
Tiirola, M. A., Männistö, M. K., Puhakka, J. A., and Kulomaa, M. S. (2002). Isolation and characterization of Novosphingobium sp. strain MT1, a dominant polychlorophenol-degrading strain in a groundwater bioremediation sysytem. Appl. Environ. Microbiol. 68, 173–180. doi: 10.1128/AEM.68.1.173-180.2002
Tóth, E. M., Vengring, A., Homonnay, Z. G., Kéki, Z., Spröer, C., Borsodi, A. K., et al. (2014). Phreatobacter oligotrophus gen. nov., sp. nov., an alphaproteobacterium isolated from ultrapure water of the water purification system of a power plant. Int. J. Syst. Evol. Microbiol. 64(Pt 3), 839–845. doi: 10.1099/ijs.0.053843-0
Tu, Q., Zhou, X., He, Z., Kai, X., Wu, L., Reich, P., et al. (2015). The diversity and co-occurrence patterns of N2 -fixing communities in a CO2 -enriched grassland ecosystem. Microb. Ecol. 71, 1–12. doi: 10.1007/s00248-015-0659-7
Tu, Q., Zhou, X., He, Z., Kai, X., Wu, L., Reich, P., et al. (2016). The diversity and co-occurrence patterns of N2-fixing communities in a CO2-enriched grassland ecosystem. Microb. Ecol. 71, 604–615. doi: 10.1007/s00248-015-0659-7
Tuomainen, J., Hietanen, S., Kuparinen, J., Martikainen, P. J., and Servomaa, K. (2006). Community structure of the bacteria associated with Nodularia sp. (Cyanobacteria) aggregates in the Baltic Sea. Microb. Ecol. 52, 513–522. doi: 10.1007/s00248-006-9130-0
Turner, A. M., and Chislock, M. F. (2010). Blinded by the stink: nutrient enrichment impairs the perception of predation risk by freshwater snails. Ecol. Appl. 20, 2089–2095. doi: 10.1890/10-0208.1
von der Weid, I., Duarte, G. F., van Elsas, J. D., and Seldin, L. (2002). Paenibacillus brasilensis sp. nov., a novel nitrogen-fixing species isolated from the maize rhizosphere in Brazil. Int. J. Syst. Evol. Microbiol. 52(Pt 6), 2147–2153.
Wang, C., Zhang, M., Cheng, F., and Geng, Q. (2015). Biodegradation characterization and immobilized strains’ potential for quinoline degradation by Brevundimonas sp. K4 isolated from activated sludge of coking wastewater. Biosci. Biotechnol. Biochem. 79, 164–170. doi: 10.1080/09168451.2014.952615
Wang, Y., Zhang, R., Zheng, Q., Deng, Y., Nostrand, J. D. V., Zhou, J., et al. (2015). Bacterioplankton community resilience to ocean acidification: evidence from microbial network analysis. ICES J. Mar. Sci. 73, 865–875. doi: 10.1093/icesjms/fsv187
Wang, S., Zhang, M., Li, B., Xing, D., Xin, W., Wei, C., et al. (2012). Comparison of mercury speciation and distribution in the water column and sediments between the algal type zone and the macrophytic type zone in a hypereutrophic lake (Dianchi Lake) in Southwestern China. Sci. Total Environ. 41, 204–213. doi: 10.1016/j.scitotenv.2011.12.036
Woese, C. R. (1994). There must be a prokaryote somewhere: microbiology’s search for itself. Microbiol. Rev. 58, 1–9.
Woodhouse, J. N., Kinsela, A. S., Collins, R. N., Bowling, L. C., Honeyman, G. L., Holliday, J. K., et al. (2016). Microbial communities reflect temporal changes in cyanobacterial composition in a shallow ephemeral freshwater lake. ISME J. 10, 1337–1351. doi: 10.1038/ismej.2015.218
Xie, C.-H., and Yokota, A. (2005). Pleomorphomonas oryzae gen. nov., sp. nov., a nitrogen-fixing bacterium isolated from paddy soil of Oryza sativa. Int. J. Syst. Evol. Microbiol. 55, 1233–1237. doi: 10.1099/ijs.0.63406-0
Xu, H., Wang, H., Xu, Q., Lv, L., Yin, C., Liu, X., et al. (2015). Pathway for biodegrading microcystin-YR by Sphingopyxis sp. USTB-05. PLoS ONE 10:e0124425. doi: 10.1371/journal.pone.0124425
Xu, L., Li, D., Wang, Q., and Wu, S. (2016). Improved hydrogen production and biomass through the co-cultivation of Chlamydomonas reinhardtii and Bradyrhizobium japonicum. Int. J. Hydrogen Ener. 41, 9276–9283. doi: 10.1016/j.ijhydene.2016.04.009
Yang, C., Li, Y., Zhou, B., Zhou, Y., Zheng, W., Tian, Y., et al. (2015). Illumina sequencing-based analysis of free-living bacterial community dynamics during an Akashiwo sanguine bloom in Xiamen sea, China. Sci. Rep. 5:8476. doi: 10.1038/srep08476
Yang, C., Li, Y., Zhou, Y., Lei, X., Zheng, W., Tian, Y., et al. (2016). A comprehensive insight into functional profiles of free-living microbial community responses to a toxic Akashiwo sanguinea bloom. Sci. Rep. 6:34645. doi: 10.1038/srep34645
Yang, C., Li, Y., Zhou, Y., Zheng, W., Tian, Y., and Zheng, T. (2012). Bacterial community dynamics during a bloom caused by Akashiwo sanguinea in the Xiamen sea area, China. Harmful Algae 20, 132–141. doi: 10.1016/j.hal.2012.09.002
Yang, L., Maeda, H., Yoshikawa, T., and Zhou, G. Q. (2012). Algicidal effect of bacterial isolates of Pedobacter sp. against cyanobacterium Microcystis aeruginosa. Water Sci. Eng. (Chinese journal) 5, 375–382.
Yang, H. X., Wang, X., Liu, X. W., Zhang, J., Yang, G. Q., Lau, K. W., et al. (2014). Fluviicola hefeinensis sp. nov., isolated from the wastewater of a chemical factory. Int. J. Syst. Evol. Microbiol. 64, 700–704. doi: 10.1099/ijs.0.056705-0
Zeng, Y., Selyanin, V., Lukeš, M., Dean, J., Kaftan, D., Feng, F., et al. (2015). Characterization of microaerophilic bacteriochlorophyll a - containing bacterium Gemmatimonas phototrophica sp. nov., and emended descriptions of the genus Gemmatimonas and Gemmatimonas aurantiaca. Int. J. Syst. Evol. Microbiol. 65, 2410–2419. doi: 10.1099/ijs.0.000272
Zhang, H., Sekiguchi, Y., Hanada, S., Hugenholtz, P., Kim, H., Kamagata, Y., et al. (2003). Gemmatimonas aurantiaca gen. nov., sp. nov., a gram-negative, aerobic, polyphosphate-accumulating micro-organism, the first cultured representative of the new bacterial phylum Gemmatimonadetes phyl. nov. Int. J. Syst. Evol. Microbiol. 53, 1155–1163. doi: 10.1099/ijs.0.02520-0
Zhou, J., Deng, Y., Luo, F., He, Z., Tu, Q., and Zhi, X. (2010). Functional molecular ecological networks. Mbio 1, 1592–1601. doi: 10.1128/mBio.00169-10
Keywords: bloom, Microcystis aeruginosa, particle-associated bacteria (PAB), free-living bacteria (FLB), microbial interaction network
Citation: Yang C, Wang Q, Simon PN, Liu J, Liu L, Dai X, Zhang X, Kuang J, Igarashi Y, Pan X and Luo F (2017) Distinct Network Interactions in Particle-Associated and Free-Living Bacterial Communities during a Microcystis aeruginosa Bloom in a Plateau Lake. Front. Microbiol. 8:1202. doi: 10.3389/fmicb.2017.01202
Received: 23 March 2017; Accepted: 13 June 2017;
Published: 30 June 2017.
Edited by:
Télesphore Sime-Ngando, Centre National de la Recherche Scientifique (CNRS), FranceReviewed by:
Ye Deng, Chinese Academy of Sciences, ChinaSara Beier, Leibniz Institute for Baltic Sea Research (LG), Germany
Copyright © 2017 Yang, Wang, Simon, Liu, Liu, Dai, Zhang, Kuang, Igarashi, Pan and Luo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Feng Luo, van77@xmu.edu.cn Xuejun Pan, xjpan@kmust.edu.cn