 Ana R. V. Pedro1,2,3
Ana R. V. Pedro1,2,3 Tânia Lima1
Tânia Lima1 Ricardo Fróis-Martins1
Ricardo Fróis-Martins1 Bárbara Leal4,5
Bárbara Leal4,5 Isabel C. Ramos6
Isabel C. Ramos6 Elisabete G. Martins7,8,9
Elisabete G. Martins7,8,9 Ana R. J. Cabrita3
Ana R. J. Cabrita3 António J. M. Fonseca3
António J. M. Fonseca3 Margarida R. G. Maia3
Margarida R. G. Maia3 Manuel Vilanova1,2
Manuel Vilanova1,2 Alexandra Correia1,2*
Alexandra Correia1,2*- 1Immunobiology Group, i3S – Instituto de Investigação e Inovação em Saúde, Universidade do Porto, Porto, Portugal
- 2Laboratório de Imunologia, DIMFF, ICBAS – Instituto de Ciências Biomédicas Abel Salazar, Universidade do Porto, Porto, Portugal
- 3LAQV, REQUIMTE, ICBAS – Instituto de Ciências Biomédicas Abel Salazar, Universidade do Porto, Porto, Portugal
- 4Laboratório de Imunogenética, DPIM, ICBAS – Instituto de Ciências Biomédicas Abel Salazar, Universidade do Porto, Porto, Portugal
- 5UMIB, Instituto de Ciências Biomédicas Abel Salazar, Universidade do Porto, Porto, Portugal
- 6Animal Nutrition Division, Cooperativa Agrícola de Vila do Conde, Vila do Conde, Portugal
- 7ADM Portugal, SA, Murtede, Portugal
- 8EPIUnit, Instituto de Saúde Pública, Universidade do Porto, Porto, Portugal
- 9Department of Veterinary Medicine, Escola Universitária Vasco da Gama, Coimbra, Portugal
Yeast-derived products containing β-glucans have long been used as feed supplements in domesticated animals in an attempt to increase immunity. β-glucans are mainly recognized by the cell surface receptor CLEC7A, also designated Dectin-1. Although the immune mechanisms elicited through Dectin-1 activation have been studied in detail in mice and humans, they are poorly understood in other species. Here, we evaluated the response of bovine monocytes to soluble and particulate purified β-glucans, and also to Zymosan. Our results show that particulate, but not soluble β-glucans, can upregulate the surface expression of costimulatory molecules CD80 and CD86 on bovine monocytes. In addition, stimulated cells increased production of IL-8 and of TNF, IL1B, and IL6 mRNA expression, in a dose-dependent manner, which correlated positively with CLEC7A gene expression. Production of IL-8 and TNF expression decreased significantly after CLEC7A knockdown using two different pairs of siRNAs. Overall, we demonstrated here that bovine monocytes respond to particulate β-glucans, through Dectin-1, by increasing the expression of pro-inflammatory cytokines. Our data support further studies in cattle on the induction of trained immunity using dietary β-glucans.
Introduction
Immune modulation by natural compounds has long been studied in domesticated animals such as poultry, fish, and livestock, to enhance immunity and improve animal welfare and wellbeing, ultimately reducing the incidence of disease and the overuse of pharmaceutical compounds, such as antibiotics. Dietary supplementation with yeasts (1–6) and yeast-derived compounds, such as mannan-oligosaccharides (MOS) and β-glucans (1, 4–7), is one of the most used strategies to enhance immunity in domesticated animals. β-glucans are naturally occurring polymers present in the cell wall of fungi, bacteria, algae, and plants. Yeast β-glucans are usually composed of linear molecules of D-glucose units linked by β-1,3 glycosidic bonds with β-1,6 branching (8, 9). β-glucans are recognized by immune cell surface pattern recognition receptors (PRR) such as C-type lectin domain family 7 member A (CLEC7A), also designated as Dectin-1, complement receptor-3 (CR3), scavenger receptors, and lactosylceramide (10). In addition, Toll-like receptor (TLR)-2 and TLR-6 can synergistically contribute to the recognition and elicited biological effects of particulate β-glucans, such as Zymosan (11–14). Biological activities of β-glucans depend on their recognition and downstream cell signalling, which in turn depend largely on the structure, conformation, and physical properties of the different β-glucans (9, 15). Although both particulate and soluble β-glucans bind Dectin-1, only the particulate form can induce Dectin-1 signalling and generate a “phagocytic synapse” (16). Activation of Dectin-1 triggers an intracellular signalling cascade eliciting phagocytosis, production of cytokines, and reactive-oxygen species (ROS) (14–19).
The immune recognition of β-glucans and elicited response has been extensively studied in mice and humans at mechanistic level (15, 20, 21). However, in other species, including cattle, the effects of β-glucans on the immune system are mainly supported by observational reports and in vivo studies (22–25). A homologous transcript for human Dectin-1 has been described in bovines (boDectin-1) (26). However, β-glucan recognition and its effects on bovine leukocytes were not fully elucidated.
Bovine monocytes express CLEC7A (26) and this cell type is the most used in innate immune memory studies in other species (27, 28). Here, the response of bovine monocytes to soluble and particulate β-glucans, and to β-glucan-containing particles (Zymosan) was assessed. Our results show that only the particulate β-glucan forms trigger the production of pro-inflammatory cytokines, and implicate boDectin-1 in this effect.
Materials and Methods
Isolation of Bovine Peripheral Blood Monocytes
Bovine blood from Holstein-Friesian cattle was obtained at a local commercial slaughterhouse (PEC Nordeste – Indústria de Produtos Pecuários do Norte, Penafiel, Portugal) and licensed by National competent authority, Direção Geral de Alimentação e Veterinária, under a by-product handling authorization (N.12.006.UDER). There was no intervention on the animals for research purposes, since blood was collected during bleeding/slaughter of animals for human consumption. Blood was collected from jugular and carotid veins to BD Vacutainer® lithium heparin tubes (BD, Franklin Lakes, NJ, USA) and peripheral blood CD14+ monocytes were obtained as previously described, with minor modifications (29). Briefly, whole blood was diluted 1:2 with Dulbecco’s phosphate-buffered saline (DPBS) and density gradient centrifuged on Histopaque®-1077 (both from Sigma-Aldrich) at 1200 × g for 15 min in SepMate™ PBMC isolation tubes (Stemcell™ Technologies, Vancouver, BC, Canada). Peripheral blood mononuclear cells (PBMC) were then washed with DPBS by centrifugation at 400 × g for 10 min and CD14+ cells were isolated with anti-human CD14 MicroBeads, according to manufacturer’s instructions (Miltenyi Biotec, Bergisch Gladbach, Germany). Peripheral blood CD14+ monocytes were washed with DPBS by centrifugation at 300 × g for 10 min and resuspended at 2 × 106 cells/mL in complete RPMI medium - RPMI-1640 Medium (Sigma-Aldrich) supplemented with 10% FBS (Biowest, Nuaillé, France), 50 µM β-mercaptoethanol (Merck, Darmstadt, Germany), 100 U/mL penicillin, 100 µg/mL streptomycin, 4 mM L-glutamine and 10 mM HEPES (all from Sigma-Aldrich). Purity of CD14+ cells exceeded 90%, as evaluated by flow cytometry using an anti-sheep CD14 mAb (clone VPM65, Bio-Rad, Hercules, CA, USA) that cross reacts with bovine, conjugated with DyLight® 405 Conjugation Kit (Abcam, Cambridge, UK).
Cell Culture and Stimulation With β-Glucans
Bovine CD14+ monocytes were plated at 2 × 105 cells/well in flat-bottom 96-well culture plates. Stimulation was done with a pure soluble β-glucan preparation (WGP®-Soluble), a purified insoluble preparation of Saccharomyces cerevisiae lacking TLR activity, composed mainly of β-1,3-glucans (WGP®-Dispersible), and an insoluble preparation of S. cerevisiae cell wall (Zymosan), described to have Dectin-1- and TLR2/6-stimulatory activity (all from InvivoGen, San Diego, CA, USA). β-glucans, labelled endotoxin-free (endotoxin level below 0.001 EU/μg), were prepared according to manufacturer’s instructions. All assays were performed using sterile, pyrogen-free material. Cells were cultured with 10, 50 and 100 µg/mL of WGP-Soluble, WGP-Dispersible or Zymosan. Cells cultured with 1 µg/mL of Escherichia coli lipopolysaccharide (LPS, strain O111:B4; Sigma-Aldrich) or Pam3CSK4 (P3C; InvivoGen) were used as positive controls. A kinetic cytokine mRNA expression analysis was performed in cells cultured for 8, 16, and 24 h at 37°C and 5% CO2 to define the time point for sequent analyses. Cytokine production, mRNA expression, and lactate dehydrogenase (LDH) release, were assessed in cells cultured for 24 h at 37°C and 5% CO2. Cell surface activation markers were assessed by flow cytometry in 8 and 16 h bovine monocyte cultures. Non-stimulated cells were always used as negative controls.
HEK-Blue™ hDectin-1b Reporter Cell Line Culture and Stimulation with β-Glucans
Hek-Blue hDectin-1b cells (InvivoGen) were grown in DMEM medium with 4.5 g/L glucose (Sigma-Aldrich), 10% heat inactivated Fetal Bovine Serum Premium (FBS) (Biowest), 2 mM L-glutamine, 100 U/mL penicillin, 100 μg/mL streptomycin (all from Sigma-Aldrich), 100 μg/mL Normocin™ and 1 µg/mL puromycin (both from InvivoGen) in vented T75 flasks. When cells reached 80% confluency, they were re-seeded at 5 × 104 cells/well in flat-bottom 96 well-culture plates and stimulated with 10, 50 and 100 µg/mL of WGP®-Soluble, WGP®-Dispersible, or Zymosan in HEK-Blue™ Detection medium for 16 h. Substrate hydrolysis by secreted alkaline phosphatase (SEAP), upon activation of the receptor, was assessed at 620-655 nm according to manufacturer’s instructions in a BioteK™ µQuant Microplate Reader using Biotek™ Gen5™ Data Collection and Analysis Software (Thermo Fisher Scientific, Waltham, MA, USA).
Cell Viability Assays
LDH release was quantified in cell culture supernatants using CyQUANT™ LDH Cytotoxicity Assay kit, according to manufacturer’s instructions (Invitrogen, Waltham, MA, USA).
Cytokine Production
Cytokine levels were assessed in cell culture supernatants by sandwich ELISA. Assessment of bovine Tumor Necrosis Factor Alpha (TNF-α) and bovine Interleukin (IL)-6 was done using Bovine TNF-alpha and Bovine IL-6 DuoSet ELISA kits (R&D Systems, Minneapolis, MN, USA) according to the manufacturer’s protocol with a minor modification: 1% molecular grade bovine serum albumin (BSA, Albumine Bovine Fraction V, NZYTech, Lisbon, Portugal) in DPBS was used as reagent diluent, instead of 5% Tween 20 in DPBS. Bovine IL-8 (Bovine IL-8 [CXCL8] ELISA development kit, Mabtech AB, Nacka Strand, Sweden) was quantified according to manufacturer’s instructions. Detection limits: 8 pg/mL for IL-8 and 125 pg/mL for TNF-α and IL-6. Bovine IL-1β was quantified using the IL-1 beta Bovine Uncoated ELISA Kit (Invitrogen), according to manufacturer’s instructions. Detection limit <31.3 pg/mL. Bovine IL-10 was assessed in cell culture supernatants using an in-house ELISA kit, following the standard procedure of Mabtech ELISA Bovine IL-8 kit. Briefly, Nunc Maxisorp™ plates were coated with 1 µg/mL anti-bovine IL-10 mAb (clone CC318; Bio-Rad) in PBS and incubated overnight at 4°C. A nine-point standard curve from 2000 to 8 pg/mL was done using Recombinant Bovine Interleukin-10 (Bio-Rad). Cell supernatants and standards were incubated at room temperature for 2 h, followed by incubation for 1 h with the detection antibody at 0.5 µg/mL (mouse anti-Bovine Interleukin-10:Biotin; clone CC320; Bio-Rad) and 1 h with Mabtech’s streptavidin-HRP, according to manufacturer’s instructions. Detection limit: 8 pg/mL. Only samples above detection limits were used for comparison.
Bovine Dectin-1 Knockdown Assays (Small Interference RNA)
The following small interfering RNAs (siRNA) were designed by Custom siRNA Design Service (Merck) to target both isoforms of bovine Dectin-1 and achieve the knockdown of this receptor: siRNA #1 sense AUG AAG AUG GAU AUA CUC A dTdT, antisense UGA GUA UAU CCA UCU UCA U dTdT; siRNA #2 sense UGA GGA UAG CUG UUA UCU A dTdT, antisense UAG AUA ACA GCU AUC CUC A dTdT; siRNA #3 sense GAG GAU AGC UGU UAU CUA U dTdT, antisense AUA GAU AAC AGC UAU CCU C dTdT (all from Sigma-Aldrich). Transfection procedure was performed for 4 h in serum-free X-VIVO™ 15 haematopoietic medium (Lonza, Basel, Switzerland) with ScreenFect®siRNA transfection reagent, according to manufacturer’s instructions (ScreenFect GmbH, Eggenstein-Leopoldshafen, Germany), and 300 nM of siRNA duplexes or siRNA negative control (MISSION® siRNA Universal Negative Control #1, Sigma-Aldrich). After transfection, cells were washed with non-supplemented RPMI-1640 medium and incubated for 24 h at 37°C and 5% CO2 with WGP®-Soluble, WGP®-Dispersible and Zymosan® at 50 µg/mL in RPMI medium or with medium alone. Supernatants were collected to assess IL-8 production and cells were preserved in NZYol reagent (NZYTech, Lisboa, Portugal) to assess CLEC7A, TNF, IL1B, IL6, and IL10 mRNA expression.
RNA Extraction and cDNA Synthesis
Total RNA was obtained using NZYol according to manufacturer’s protocol, with minor modifications. Bovine monocytes were lysed with 200 µL NZYol and incubated with 0.1 µg/mL RNA-grade Glycogen (Thermo Fisher Scientific). Each sample was incubated with 80 µL chloroform for phase separation and 200 µL isopropanol was added for RNA precipitation. The precipitated RNA was washed with 70% ethanol and resuspended in 5 µL RNase-free water. Synthesis of first-strand cDNA was done in an Applied Biosystems® 2720 Thermal Cycler (Thermo Fisher Scientific) at 25°C for 10 min, 50°C for 30 min, and 85°C for 5 min using NZY First-Strand cDNA Synthesis Kit, according to manufacturer’s instructions (NZYtech). Samples were kept at -20°C. Negative controls using RNA samples for cDNA synthesis without reverse transcriptase (no RT control), and with no added template (no template control) were also included for all primer pairs.
Real-Time qPCR
Primers for β2 microglobulin (B2M), CLEC7A, IL1B, and IL6 were designed using Primer-BLAST web tool developed by NCBI (30). Primers for TNF and IL10 were previously designed (31, 32). Sequences of each primer and expected amplicon sizes are detailed in Table 1. Primers targeting CLEC7A were designed to both short and long isoforms. Determination of TNF, IL1B, IL6, IL10 and CLEC7A mRNA levels was performed in a CFX96™ Real-Time PCR Detection System (Bio-Rad), using NZYSpeedy qPCR Green Master Mix (2×) ROX plus (NZYTech). B2M and MARVEL domain containing 1 (MARVELD1), already used as reference genes in bovine gene expression studies, were used for mRNA normalization (33–35). Reaction was performed in low profile, non-skirted, 96-well PCR plates (Thermo Fisher Scientific) containing 5 µL Master Mix, 1 µL cDNA, 3.6 µL H2O and 0.2 mM of specific forward and reverse primers (all from Sigma-Aldrich). PCR program was as follows: denaturation for 5 min at 95°C followed by 40 cycles at 95°C for 5 s and 62°C for 20 s for amplification. Gene expression values were analyzed by the comparative threshold cycle method using the formula 2-(CT gene of interest - CT housekeeping gene) (36). CLEC7A PCR products were run in 1.5% (w/v) Tris-acetate-EDTA (TAE) agarose gel electrophoresis to confirm amplicon size. Bands were visualized in a Syngene™ NuGenius Gel Documentation System, excised from the gel and purified using NZYGelpure columns (NZYTech) following manufacturer’s instructions. PCR products and DNA purified from excised gel bands were Sanger sequenced to confirm primer specificity.
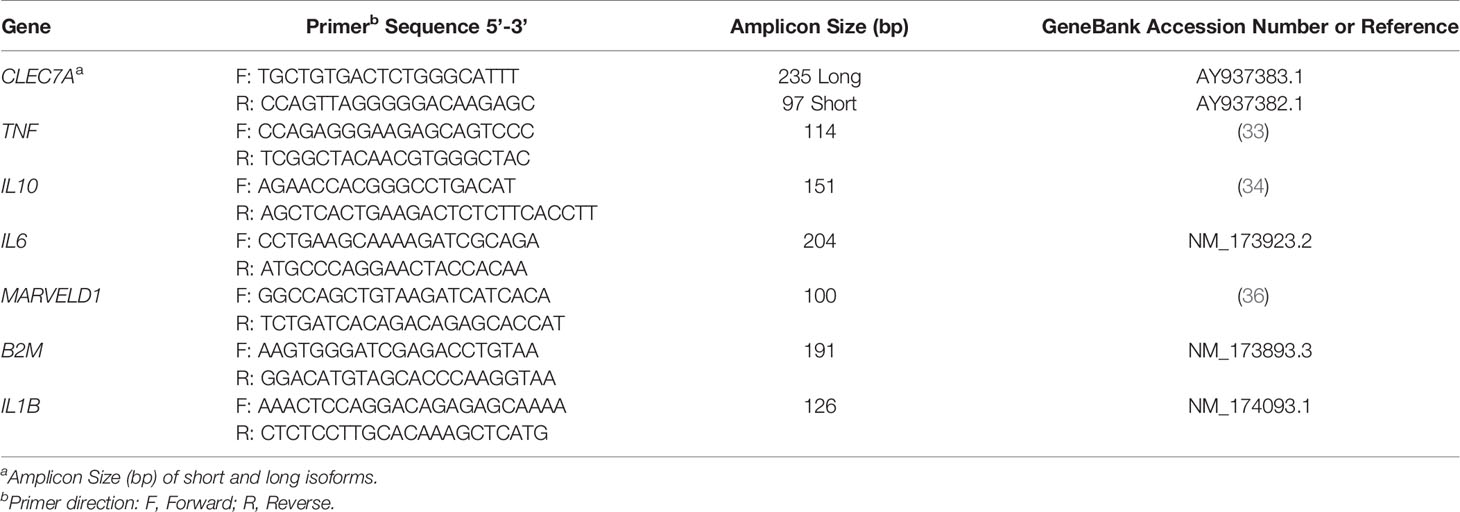
Table 1 List of primers used for quantitative real-time PCR.
Flow Cytometry
Since the commercially existing labelled antibodies for the bovine species are available in limited fluorophore diversity, we conjugated mouse monoclonal anti-bovine MHC class II DR (clone CC108, Bio-Rad) antibody with peridinin-chlorophyll protein-cychrome 5.5 (PerCP-Cy5.5) with LYNX Rapid PerCP-Cy5.5 Antibody Conjugation Kit (Bio-Rad), according to manufacturers’ instructions, and used it at 1:200 to allow multiparametric simultaneous analysis with the monoclonal antibodies mouse anti-bovine CD80 conjugated with R-Phycoerythrin (RPE) (clone IL-A159, Bio-Rad) and mouse anti-bovine CD86 conjugated with Fluorescein isothiocyanate (FITC) (clone IL-A190, Bio-Rad), both used at 1:50. All the antibodies were previously titrated to determine the optimal concentration for bovine monocyte staining. A fixable viability dye (FVD) was included before surface antibody staining to exclude dead cells from the analysis. For that, cells were incubated with eFluor® 506 Fixable Viability Dye (eBioscience, San Diego, CA, USA) diluted at 1:1000 in DPBS for 15 min at 4°C. After washing cells with DPBS, cells were incubated in 2% mouse serum in FACS Buffer (1% BSA in DPBS) for 15 min at 4°C in the dark before antibody staining to minimize nonspecific binding. A mix containing all antibodies was added to samples that were incubated for 25 min at 4°C in the dark. Cells were washed with FACS Buffer and analyzed by flow cytometry. Single stainings using UltraComp beads (eBioscience) or/and cells were used for compensation. Fluorescence minus one controls (FMO) of each antibody and FVD were used for gating purposes. Data were acquired in a BD CantoII™ equipment (BD Biosciences) and analyzed with FlowJo version 10.6.2. (FlowJo LLC, Ashland, OR, USA).
Bovine Dectin-1 Staining
Bovine monocytes and Hek-Blue hDectin-1b cells were incubated with monoclonal mouse anti-Human Dectin-1/CLEC7A antibody (Clone 259931, R&D Systems) at 10 µg/mL for 1 h at 4°C followed by incubation with the anti-mouse IgG (H+L) F(ab’)2 Fragment conjugated with Alexa Fluor® 488 (Cell Signaling Technology, Danvers, MA, USA) at 1:200 for 30 min at 4°C in the dark. Cell staining was evaluated by flow cytometry in a BD CantoII™ cytometer (BD Biosciences) and analyzed with FlowJo version 10.6.2.
Imaging of Dectin-1 on the surface of bovine monocytes and HEK-Blue™ hDectin-1b cells was done in cells stained as before. Nuclei were stained with DAPI and samples were plated in 8 well microscopy chamber plates (Ibidi, Gräfelfing, Germany) and observed in a laser scanning confocal microscope Leica TCS SP5 II system (Leica DMI6000-CS microscope with LAS AF Software, Leica Microsystems, Wetzlar, Germany). Images were obtained with a HC PL APO CS 40x/1.10 CORR Water objective. Cells were observed using 405 nm and 488 nm lasers, in the xy plane.
Statistical Analysis
Log transformations were applied to cytotoxicity, cytokine production, mRNA expression, siRNA assays, and flow cytometry data following a lognormal distribution. All data were analyzed using the MIXED Procedure of the SAS software (Version 9.1, SAS Institute Inc., Carry, NC, USA). The model included the fixed effect of treatment (Medium, WGP-Soluble, WGP-Dispersible, and Zymosan), the random effect of animal blood donor and the random residual error. The Tukey-Kramer’s post-hoc test was used to compare means of cytotoxicity, cytokine production, cytokine mRNA expression, siRNA assays, and flow cytometry data, whereas for hDectin-1b activity in HEK-Blue™ hDectin-1b cells data were used the Dunnett’s multiple comparisons test (SAS software). Cytokine levels or mRNA expression were expressed as Log fold changes to the respective values of control (Medium) samples. The Pearson correlations between bovine Log CLEC7A mRNA expression of non-stimulated samples and Log Fold change cytokine production or cytokine mRNA expression, for each stimulus, were estimated using the CORR procedure of the SAS software. Results were considered statistically significant if P<0.05 and a tendency if 0.05≥P<0.1. Graphs were constructed with the GraphPad software (Version 9.0.2, San Diego, CA, USA).
Results
HEK-Blue™ Cell Stimulation
The stimulatory effect of different commercial β-glucans or β-glucan-containing particles were tested in the HEK-Blue™ hDectin-1b reporter assay as a control prior to stimulation of bovine monocytes (Supplementary Figure 1). Soluble β-glucans did not activate hDectin-1b at any of the concentrations used, while dispersible β-glucans activated hDectin-1b at 50 and 100 µg/mL (P<0.01) comparatively to unstimulated cells. Zymosan significantly stimulated HEK-Blue™ hDectin-1b cells at all concentrations tested, 10 µg/mL (P<0.001), 50 µg/mL (P<0.0001) and 100 µg/mL (P<0.01).
Bovine Monocyte Stimulation
Viability assays were performed to assure none of the stimuli were cytotoxic to bovine monocytes at the working concentrations. None of the β-glucan sources, LPS, or P3C induced statistically significant cell death when compared to unstimulated cells (Supplementary Figure 2). The production/expression of the pro-inflammatory cytokines IL-1β, IL-6, IL-8, and TNF-α, and of the anti-inflammatory cytokine IL-10 by bovine monocytes was evaluated after stimulation with different β-glucans, LPS or P3C. These cytokines are the most commonly assessed in β-glucan-stimulated cell studies (16, 28, 37, 38). Cells were stimulated for 8, 16, and 24 h to evaluate the kinetics of cytokine expression (Supplementary Figure 3). The 24 h time point was selected for further studies since it allows simultaneous cytokine protein and mRNA analysis. Cytokine production and mRNA expression were affected by treatment (P<0.0001 for IL-8, IL-6, TNF-α levels, and IL1B, IL6 and TNF mRNA expression; P=0.0013 for IL-1β and P=0.0002 for IL-10 levels; P=0.0005 for IL10 mRNA expression) and a dose-response effect was observed for IL-8 levels (Figure 1), and IL1B, IL6 and TNF expression (Figure 2). WGP-Soluble treatment did not significantly affect the production or gene expression of any cytokine compared with unstimulated cells (P>0.05). WGP-Dispersible, Zymosan, LPS and P3C induced the production of IL-8 (Figure 1A). IL-6 levels (Figure 1B) were increased in bovine monocyte cell cultures stimulated with Zymosan, LPS and P3C. TNF-α (Figure 1C) and IL-10 (Figure 1E) levels were only increased in bovine monocytes cultured with Zymosan. A tendency for increased IL-1β production was observed in cells stimulated with Zymosan (Figure 1D). Expression of TNF (Figure 2A) was significantly increased in cells stimulated with WGP-Dispersible, while IL6 (Figure 2B) and IL1B (Figure 2C) were overexpressed in cells stimulated with WGP-Dispersible, Zymosan, and LPS. Although there was an effect of treatment on IL10 mRNA transcript levels (Figure 2D), the expression of this cytokine gene was only upregulated in cells stimulated with Zymosan at 100 µg/mL, when compared to unstimulated cells. The cytokine mRNA expression results were similar when normalization was done to B2M mRNA expression (Supplementary Figure 4).
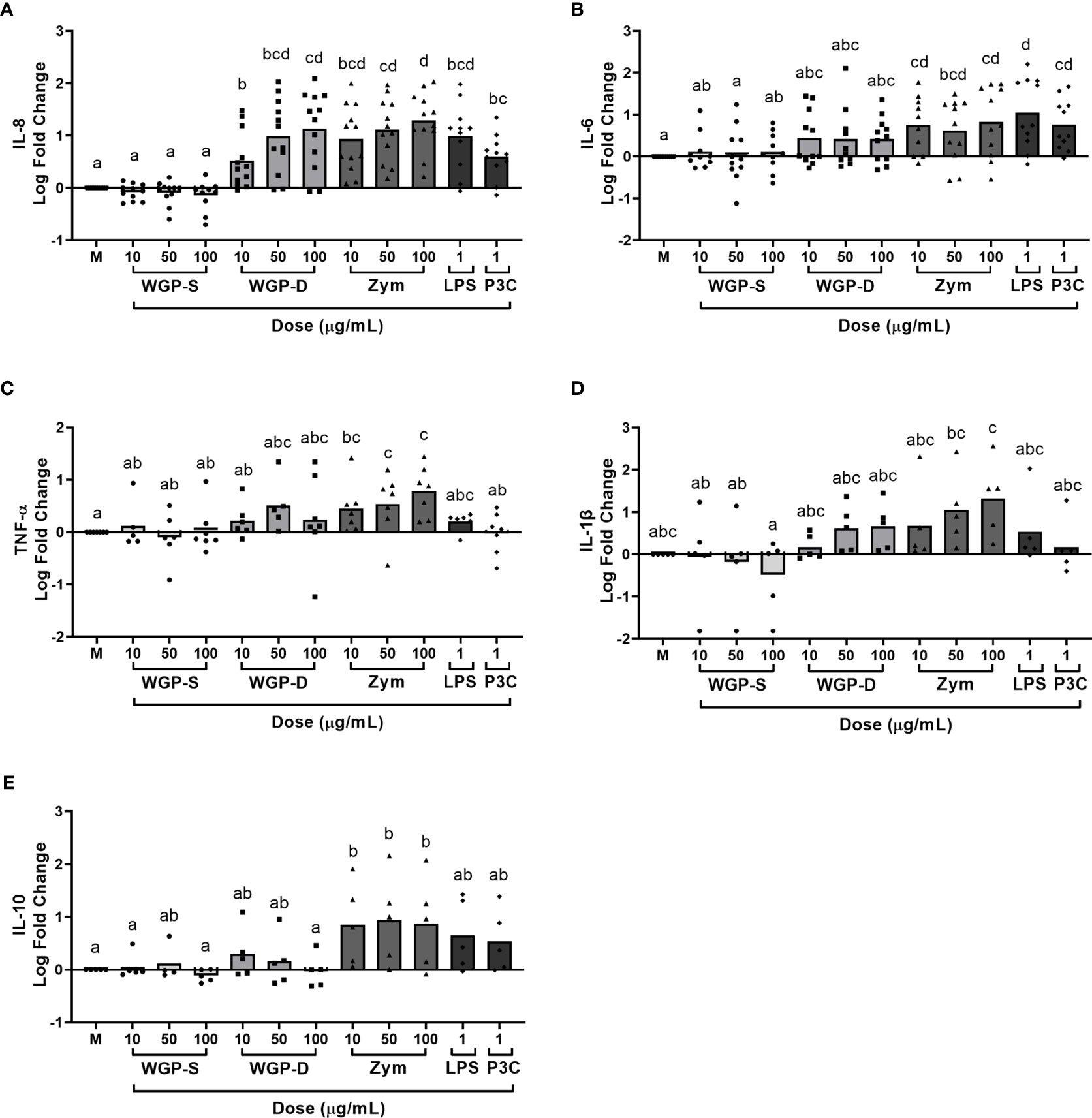
Figure 1 Cytokine production evaluated by ELISA in the supernatants of bovine monocytes cultured for 24 h with WGP Soluble (WGP-S), WGP Dispersible (WGP-D), Zymosan (Zym), LPS, and Pam3csk4 (P3C). Data are presented as Log fold change relative to medium (M) and represent means of 12 animals for IL-8 (A), 11 animals for IL-6 (B), 7 animals for TNF-α (C), and 5 animals for IL-1β (D) and IL-10 (E). Each symbol corresponds to a different animal. a,b,c,d Means with different superscript letters are significantly different (P<0.05).
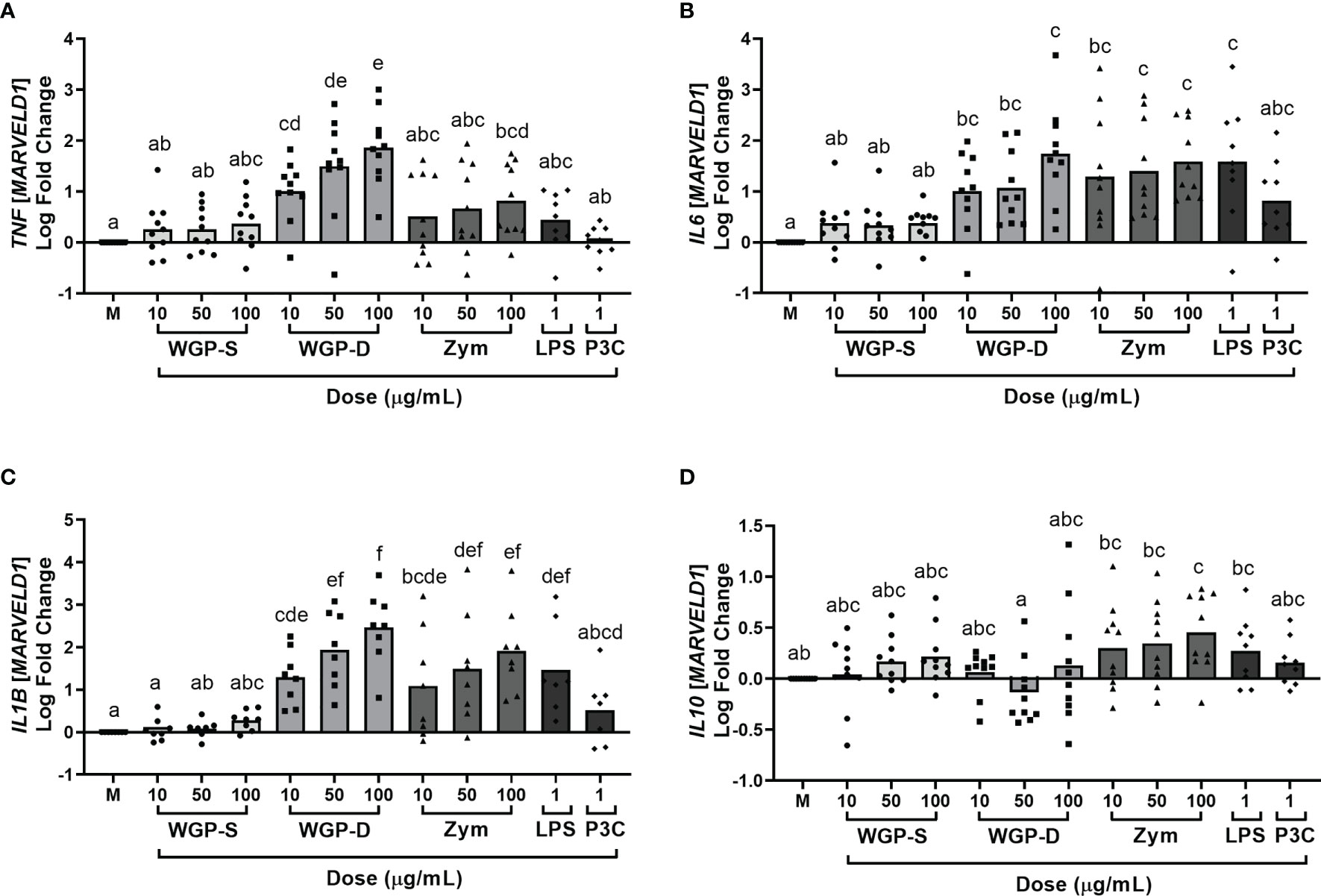
Figure 2 Cytokine relative mRNA expression, evaluated by RT-PCR and normalized to the mRNA expression of the reference gene MARVELD1, in bovine monocytes cultured for 24 h with WGP Soluble (WGP-S), WGP Dispersible (WGP-D), Zymosan (Zym), LPS, and Pam3csk4 (P3C). Data are presented as Log fold change relative to medium (M) and represent means of ten animals for TNF (A), IL6 (B), and IL10 (D), and eight animals for IL1B (C). Each symbol corresponds to a different animal. a,b,c,d,e,f Means with different superscript letters are significantly different (P<0.05).
Since WGP-Dispersible and Zymosan stimulated cytokine production by bovine monocytes, we next examined the correlation between bovine CLEC7A expression of non-stimulated cells and cytokine levels or mRNA expression in response to stimulation with 10, 50 and 100 µg/mL of WGP-Dispersible and Zymosan. A positive correlation was found between CLEC7A mRNA expression and IL-8 concentration in the supernatants of cells stimulated with 10 µg/mL (r = 0.7702) and 50 µg/mL (r = 0.6629) (Figures 3A, B, respectively), but not with 100 µg/mL (Figure 3C) of WGP-Dispersible. TNF mRNA expression was positively correlated with CLEC7A mRNA expression at 10 µg/mL (r = 0.7067) and 50 µg/mL (r = 0.6429) (Figures 3D, E), but not at 100 µg/mL (Figure 3F). IL6 mRNA expression was also correlated with CLEC7A expression in cells stimulated with WGP-Dispersible at 10 µg/mL (r = 0.7093, Figure 3G) and a tendency was observed in cells stimulated with 50 µg/mL (r = 0.5939, Figure 3H). No such correlation was observed when cells were stimulated with 100 µg/mL WGP-Dispersible (Figure 3I). A tendency was observed between CLEC7A mRNA expression and IL1B mRNA expression in cells stimulated with 10 µg/mL (r = 0.6672, Figure 3J), but not when cells were stimulated with 50 µg/mL or 100 µg/mL WGP-Dispersible (Figures 3K, L, respectively). Interestingly, a negative correlation was observed between CLEC7A and IL10 mRNA expression in cells stimulated with 10 µg/mL (r = -0.6978, Figure 3M) and 100 µg/mL (r = -0.7718, Figure 3O) of WGP-Dispersible, and a tendency to a negative correlation in cells stimulated with 50 µg/mL (r = -0.5926, Figure 3N) of this β-glucan form. No correlation was found between CLEC7A mRNA expression and cytokine mRNA expression or production in cells stimulated with Zymosan (Supplementary Figure 5).
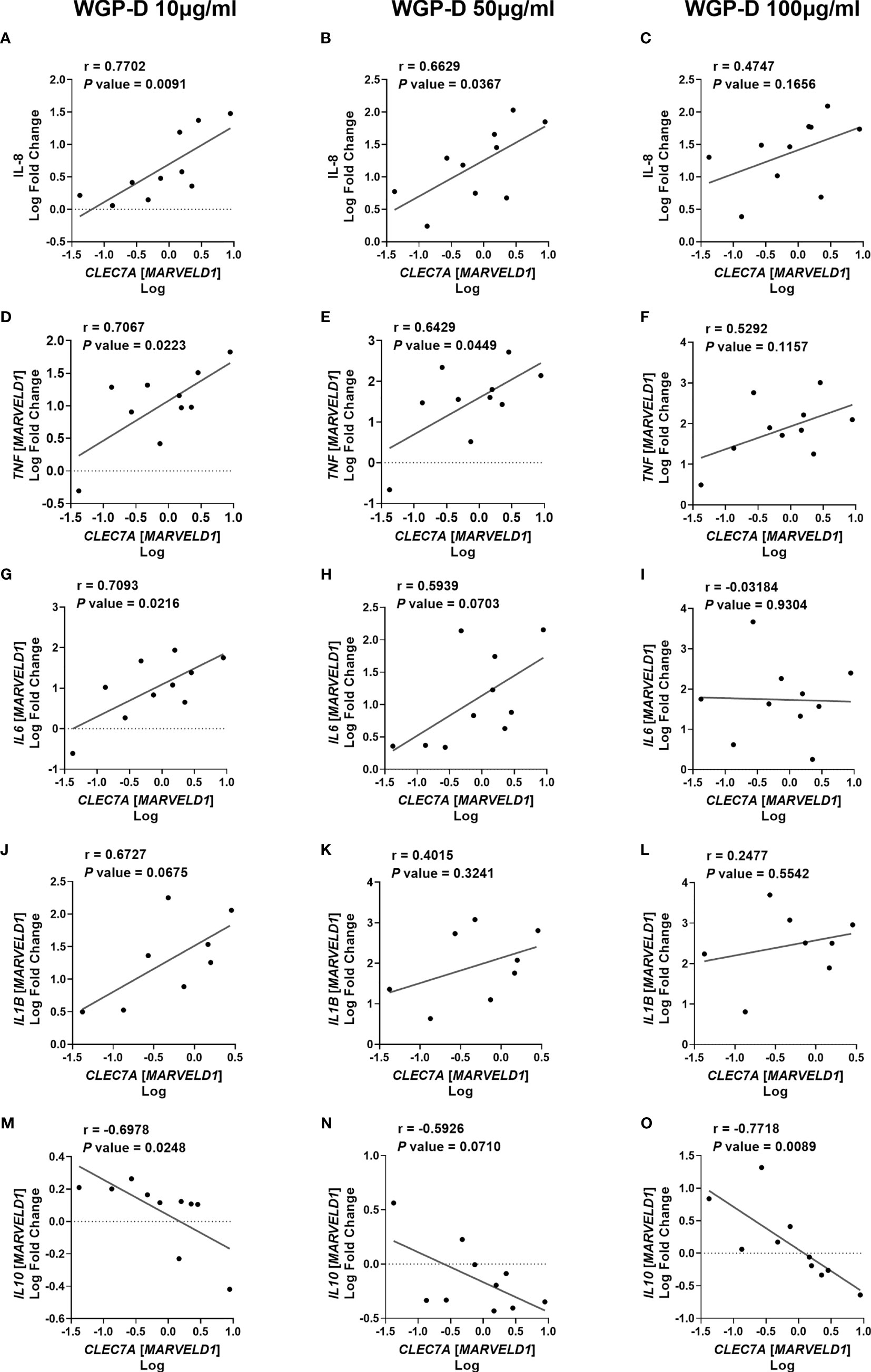
Figure 3 Correlations between CLEC7A mRNA expression and (A–C) IL-8 cytokine production, (D–F) TNF, (G–I) IL6, (J–L) IL1B, and (M–O) IL10 mRNA expression upon stimulation with 10, 50 and 100 µg/mL of WGP Dispersible, as indicated. Results are presented as Log fold changes of each cytokine relative to medium vs Log CLEC7A mRNA. Data represent simple linear regressions, with Pearson correlation coefficients (r) and P values.
Sequencing of PCR products was done to confirm primers’ specificity (Supplementary Figure 6). Since more than one CLEC7A amplicon was amplified in each sample, PCR products were also visualized in agarose gels to confirm the molecular size of the amplicons (Supplementary Figure 6A). According to our data, different CLEC7A isoforms were expressed simultaneously in monocyte samples, since two different bands, matching the expected molecular size distribution for the two different isoform amplicons (97 bp for the short or 235 bp for the long) appeared on the electrophoresis gel. This is in line with what was previously observed and described by Willcocks et al. (26) for short and long isoforms of boDectin-1. The expression of CLEC7A was decreased upon treatment with WGP-Dispersible and Zymosan, but not with WGP-Soluble (Supplementary Figure 7). Previous reports have shown a downregulation of Dectin-1 expression on the surface of human monocyte- and mouse bone marrow-derived dendritic cells and mouse bone marrow-derived macrophages in response to particulate β-glucan stimulation (39–41). However, no mention was made therein whether CLEC7A gene expression was also reduced.
Expression of MHC Class II and Costimulatory Molecules
The expression of costimulatory (CD80 and CD86) and MHC class II molecules on the cell surface of monocytes upon stimulation with the different β-glucans or β-glucan-containing particles was evaluated by flow cytometry, as indicated in Supplementary Figure 8. WGP-Soluble treatment did not alter the expression of MHC class II or costimulatory molecules at any assessed time point (Figure 4). Zymosan treatment upregulated the expression of CD80 and CD86 at 8 h (Figures 4A, C respectively) at 50 and 100 µg/mL, but only the expression of CD80 remained upregulated at 16 h (Figures 4B, D). Cells stimulated for 16 h with 100 µg/mL WGP-Dispersible and P3C increased the expression of CD80 (Figure 4B). No differences in the expression of MHC class II were observed at any of these timepoints (Figures 4E, F), besides a decrease in LPS-treated-cells at 16 h post stimulation.
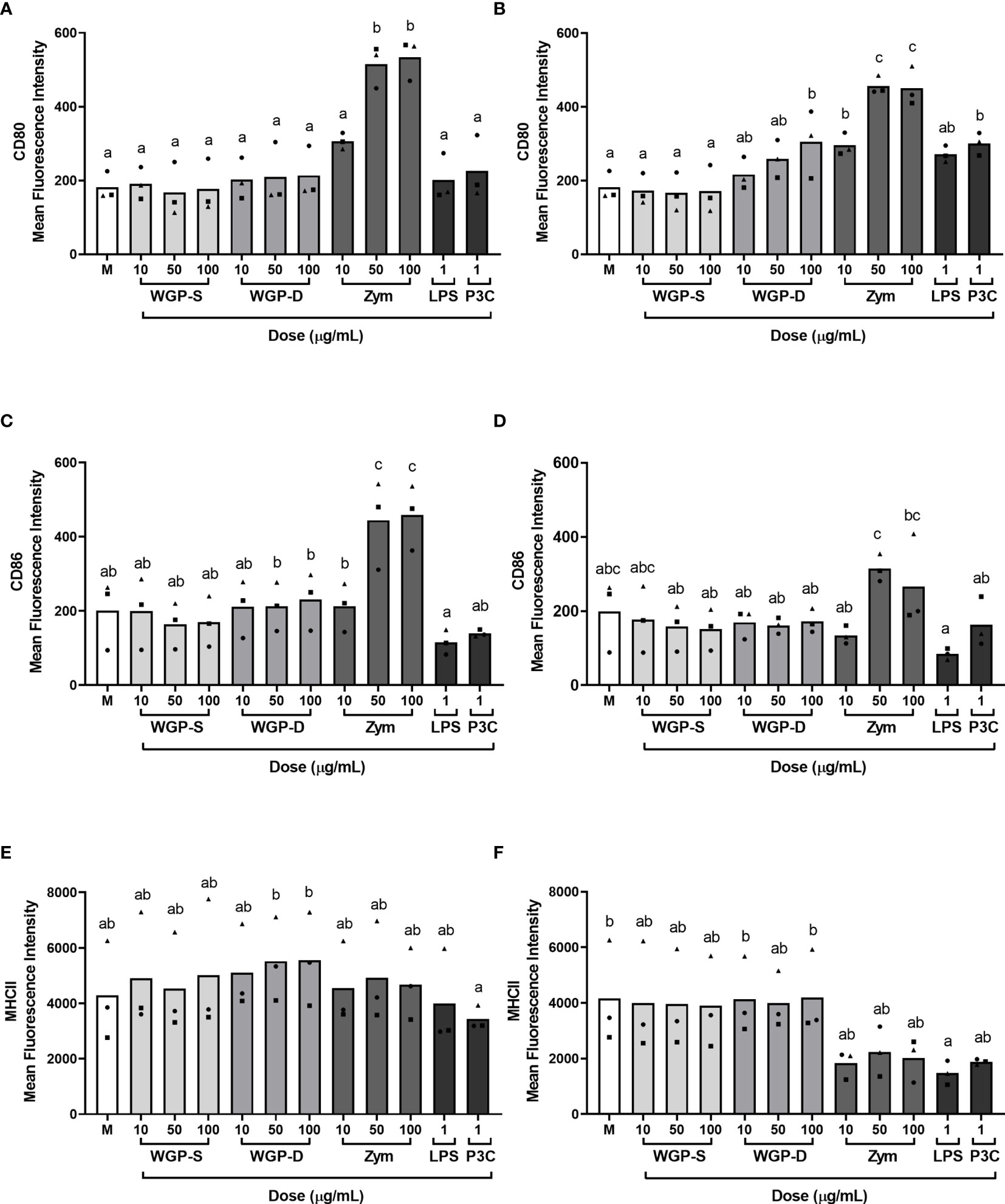
Figure 4 Expression of (A, B) CD80, (C, D) CD86, and (E, F) MHC class II molecule expression on the cell surface of bovine monocytes stimulated with WGP Soluble (WGP-S), WGP Dispersible (WGP-D), Zymosan (Zym), LPS, and Pam3csk4 (P3C) for 8 h (A, C, E) or 16 h (B, D, F), as evaluated by flow cytometry. Results correspond to means of the mean fluorescence intensities for each analyzed molecule of three independent biological samples (each represented by squares, triangles or circles). a,b,c Means with different superscript letters are significantly different (P<0.05).
Small Interference mRNA and Bovine Dectin-1 Knockdown
IL-8 levels and TNF and IL6 expression were significantly increased in monocytes stimulated with either WGP-Dispersible or Zymosan, and their increase was found to be correlated with CLEC7A expression in WGP-Dispersible-treated cells. Therefore, we further investigated the role of boDectin-1 in IL-8 production and TNF and IL6 expression by silencing the receptor using a siRNA approach. Three pairs of siRNA duplexes were designed to target and silence the two bovine Dectin-1 isoforms. Transfection did not affect cell viability (Supplementary Figure 9).
Bovine CLEC7A was successfully knocked down by siRNA duplex #3 (85.5%, P<0.01) and, to a lesser extent, by siRNA #2 (66.6%, P<0.05) at 300 nM (Figure 5A). The knockdown efficiency of this siRNA design was particularly evident in two of the samples tested (Biological samples #2 and #3, Supplementary Table 1). siRNA design #1 was not able to successfully knock down bovine CLEC7A in any of the samples tested (Figure 5A).
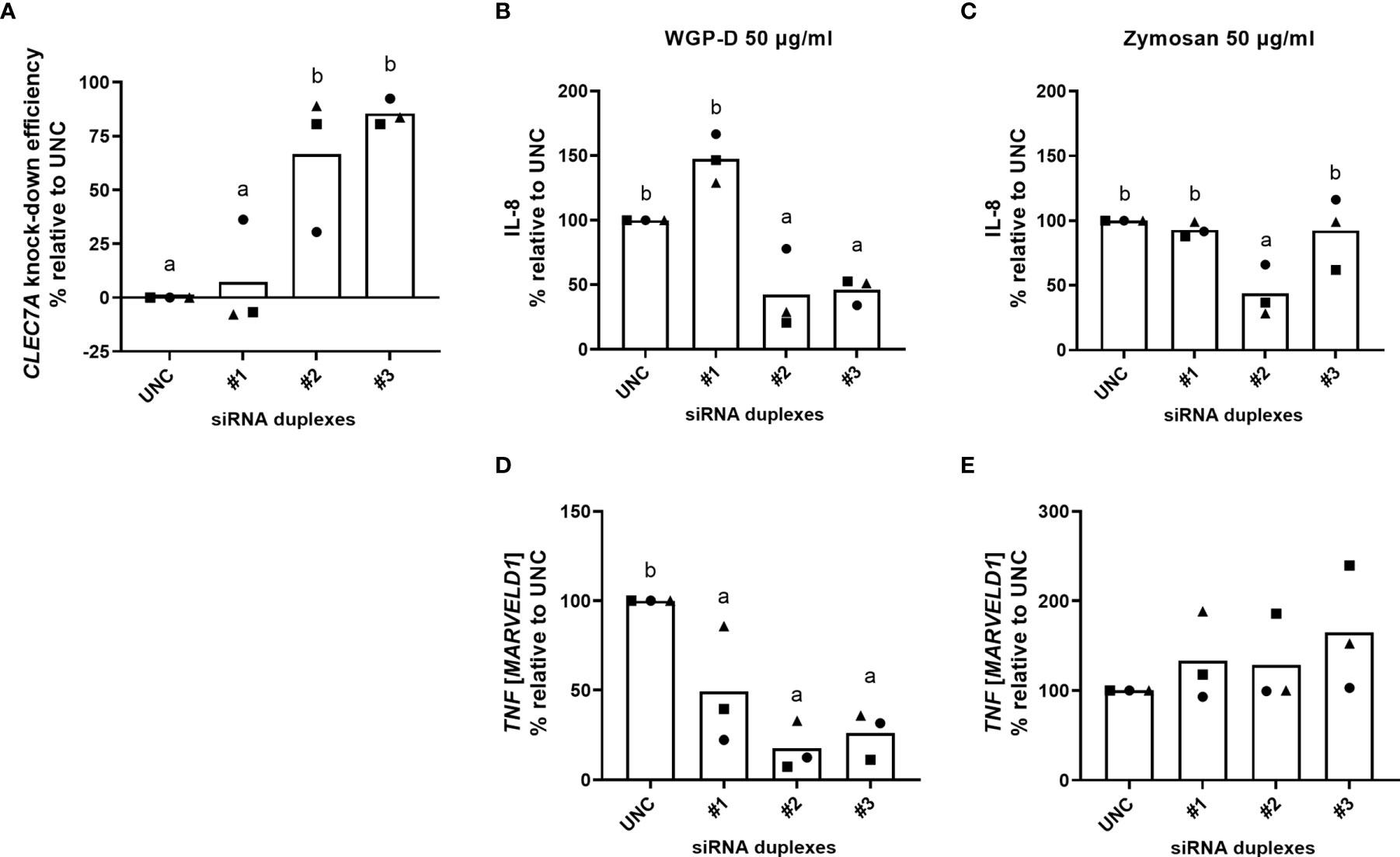
Figure 5 Bovine CLEC7A knockdown efficiency (A), calculated relative to MISSION® siRNA Universal Negative Control #1 (UNC) treated cells. Cells were cultured with RPMI-1640 after transfection procedure with three different siRNA duplexes (#1, #2 and #3). IL-8 production (B, C) and TNF expression (D, E) of cells transfected with duplexes #1, #2 and #3 and MISSION® siRNA Universal Negative Control #1 (UNC) and stimulated with WGP-Dispersible (B, D) or Zymosan (C, E) at 50 µg/mL, calculated in percentual change relative to UNC transfected cells. Results correspond to means from three different animals (each one represented by squares, triangles or circles). a,b Means with different superscript letters are significantly different (P<0.05).
Lower IL-8 levels were found in the supernatants of cells transfected with siRNA pairs #2 and #3 upon stimulation with WGP-Dispersible at 50 µg/mL, comparatively to cells treated with medium alone (X-Vivo), ScreenFect®siRNA transfection reagent (Screenfect), and MISSION® siRNA Universal Negative Control #1 (UNC). The percentual change of cytokine production was then calculated considering IL-8 production of Negative Control transfected cells. IL-8 production was significantly decreased (P<0.05) in WGP-Dispersible stimulated cells (Figure 5B) when transfection was performed with siRNA duplex #2 (57.6% decrease) and siRNA duplex #3 (54.1% decrease). Only siRNA duplex #2 was able to successfully reduce IL-8 production (56.4% decrease, P<0.05) in cells stimulated with Zymosan at 50 µg/mL (Figure 5C). siRNA duplex #2 led to a clear decrease in IL-8 production in two of the samples stimulated with WGP-Dispersible at 50 µg/mL (Figure 5B). A smaller reduction was observed in one of the samples used. The observed effect matches the low CLEC7A knockdown efficiency observed in this particular sample (Figures 5A and Supplementary Table 1). The expression of TNF in cells stimulated with WGP-Dispersible at 50 µg/mL was also significantly affected by CLEC7A knockdown (Figure 5D). No such effect was observed in Zymosan-treated cells (Figure 5E). Although not statistically different, the expression of IL6 and IL1B was decreased upon siRNA#2 treatment in response to WGP-Dispersible (Supplementary Figure 10). Given the high homology of bovine and human Dectin-1, and since no bovine-specific anti-Dectin-1 antibody is available, neutralization of this receptor was attempted using an anti-human Dectin-1 mAb. The used antibody did not recognize boDectin-1 as evaluated by flow cytometry and confocal microscopy (Supplementary Figures 11, 12, respectively).
Discussion
The effects of β-glucans or β-glucan-containing products have been explored in ruminants, either in vitro (42–44) or in vivo, by oral administration (23, 24, 44–47), with the purpose of increasing immunity or response to stressors. However, the immunostimulatory effect of β-glucan-containing products, such as Zymosan, on bovine cells mostly involved the analysis of reactive oxygen and nitrogen species production by neutrophils and monocyte-derived macrophages (48–50). BoDectin-1 has been previously identified and CLEC7A gene expression was detected in several bovine immune cell populations, such as monocytes, monocyte-derived dendritic cells, CD4+ T cells, CD21+ B cells, and NK cells (26). Contrary to human and mouse (10, 51) neutrophils, bovine neutrophils do not seem to express CLEC7A (52). Thus, although bovine neutrophils respond to Zymosan by increasing ROS production, this effect was dependent on Ca+ influx and mediated, at least in part, by CD11b (52), a component of CR3, an important β-glucan receptor in human neutrophils (53). Bovine-derived macrophages also increased the production of ROS in response to Zymosan, although the receptor involved was unraveled (54). In that line, a bovine macrophage cell line (BOMAC) challenged with S. cerevisiae cell wall components consistently expressed higher levels of IL-6, regardless of the yeast strain used, but no confirmation of the receptor responsible for cell activation nor evaluation of putative β-glucan-receptors’ expression were done (55). Nevertheless, a human fibroblast cell line (HEK293) transfected with boDectin-1 responded to Zymosan by increasing the production of IL-8, indicating that this bovine receptor could directly recognize β-glucan-containing particles (37).
Here we demonstrated that bovine monocytes respond to particulate β-glucans, through Dectin-1 triggering, resulting in increased expression of pro-inflammatory cytokines. Incubation of bovine monocytes with soluble β-glucans did not induce the production and mRNA expression of any of the cytokines assessed, nor the expression of MHC class II and costimulatory molecules CD80 and CD86 on the surface of monocytes, suggesting that soluble β-glucans do not activate bovine CLEC7A in vitro. Soluble β-glucans, despite being ligands of human and murine Dectin-1, are not able to cluster and activate in vitro the receptor (16), thus not inducing downstream cell signaling and activation (56). Pro-inflammatory cytokine production and cytokine gene expression were significantly increased in cells stimulated with dispersible β-glucans and Zymosan in a dose-dependent manner. These results are consistent with in vitro data obtained with murine bone marrow-derived macrophages and dendritic cells (57), murine resident macrophages (56), human whole blood (58), and porcine innate immune cells, namely peripheral-blood mononuclear cells and neutrophils (59). Cytokine response was in accordance with the increased costimulatory molecule expression observed on the surface of WGP-Dispersible and Zymosan-treated monocytes. Dispersible β-glucans and Zymosan, induced the upregulation of costimulatory molecules, which might contribute to improve T cell stimulation. Other authors (12, 14) have previously highlighted the importance of a crosstalk between different receptors, such as Dectin-1 and TLR-2. When several PRRs are activated simultaneously by particulate β-glucans, a complex cascade of cell signaling is usually amplified by this collaboration (60). Both Dectin-1 and TLR-2 recognize Zymosan, thus Zymosan is likely able to induce a more sustained and marked cell stimulation with a concomitant higher cytokine production.
The positive correlation found here between CLEC7A expression and TNF and IL6 expression and IL-8 production in cells stimulated with WGP-Dispersible indicates that BoDectin-1 mediates β-glucan recognition in bovine monocytes. We found, however, no correlation between CLEC7A expression and cytokine production or gene expression in Zymosan stimulated cells. Since Zymosan contains other pathogen-associated molecular patterns besides β-glucans, other receptors being triggered by those compounds could be contributing to cytokine production and hamper a direct association. Although a combined recognition of Zymosan by multiple PRRs, was reported in human and mouse cells, namely by Dectin-1 plus TLR-2 (61), Willcocks et al. (37) have reported that HEK293 cell line transfected with both boDectin-1 and boTLR-2 did not increase the production of IL-8 in response to Zymosan comparatively to HEK293 cells expressing boDectin-1 alone. Indeed, in that particular study, HEK293-boTLR-2 did not respond to Zymosan (37).
We found a negative correlation between CLEC7A and IL10 gene expression, reinforcing the ability of this highly pure β-glucan in inducing the production of pro-inflammatory cytokines, rather than anti-inflammatory cytokines. In contrast, Zymosan at 100 µg/mL induced the expression of the anti-inflammatory cytokine IL-10, comparatively to control cells. This is in accordance with previous reports describing an increased production of IL-10 in human and murine dendritic cells in response to these β-glucan-containing particles, which may confer immunological cell tolerance (62). Since WGP-Dispersible, which does not trigger TLR-signaling, did not induce IL10 gene expression, we hypothesize that TLR-2, a receptor already associated with high IL-10 production (63), is being activated upon recognizing other Zymosan components.
These results indicate that boDectin-1 on bovine monocytes is effectively being triggered by particulate β-glucans similarly to mouse and human monocytes. In contrast, in porcine macrophages, Dectin-1 silencing did not affect cytokine production (38), reinforcing the species-specific nature of the PRR response to several agonists (37, 54). We have attempted Dectin-1 blockade with an anti-human Dectin-1 neutralizing mAb without success. The availability of a bovine anti-Dectin-1 mAb, with neutralizing functions, would be of most importance to both assess cell surface Dectin-1 expression and perform further functional assays. This would more directly allow to uncover the signaling pathways elicited by particulate β-glucans. Since Dectin-1 knockdown using siRNAs did not completely resume IL-8 production and TNF, IL6, and IL1B expression to negative control levels, it is possible that other receptors could additionally be involved in the recognition of WGP-Dispersible. It thus remains to be elucidated whether bovine CR3 is also playing a role on the recognition of particulate β-glucans by bovine monocytes, as in human monocytes (64) and in mouse (65) and swine macrophages (38).
Recognition of β-glucans by Dectin-1 has been shown to induce epigenetic modifications in immune cells, that render them more efficient in responding to infection (66), a phenomenon referred to as trained immunity (66, 67). It is therefore conceivable that bovine monocytes, such as those of mice (68), humans (28), dogs (69), and chicken (70) may be prone to induction of trained immunity. That would provide a plausible explanation for the beneficial effects of β-glucan-containing dietary supplements observed in vivo in cattle (23, 45–47). The in vitro results obtained in this study may help clarify β-glucan recognition by bovine monocytes and lend support to further studies addressing trained immunity events in this species.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
AP conducted the experiments, performed data acquisition and analysis, and wrote the manuscript. AC, TL, RF-M, and BL participated in the experiments. EM and IR assisted in the design of the experiments. ARJC and AF assisted in the interpretation of data. AC, MM, and MV conceived and designed the experiments, supervised the experimental work, assisted in data acquisition and analysis, and assisted in manuscript writing. All authors contributed to the article and approved the submitted version.
Funding
This work received financial support from PT national funds (FCT/MCTES, Fundação para a Ciência e Tecnologia and Ministério da Ciência, Tecnologia e Ensino Superior) through the project UIDB/50006/2020. AP was supported by FCT phD grant PD/BDE/135540/2018. AC was supported by FCT Individual CEEC 2017 Assistant Researcher Grant CEECIND/01514/2017. MM was supported by FCT through program DL 57/2016 – Norma transitória (SFRH/BPD/70716/2010).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors are thankful to PEC-NORDESTE - Indústria de produtos pecuários do Norte, S.A. slaughterhouse and Eng. Ana Raquel Cândido for kindly providing the blood samples used in this study. The authors wish to express their gratitude to Eng. Ana Gomes and André Lopes from Animal Nutrition Division of Cooperativa Agrícola de Vila do Conde for expert discussion and to Encarnação Rebelo from ICBAS-UP for excellent technical assistance. We acknowledge the support of the i3S Scientific Platforms, namely Translational Cytometry, Cell Culture and Genotyping, and the technical assistance of Paula Magalhães and Tânia Meireles, Advanced Light Microscopy, member of the national infrastructure PPBI-Portuguese Platform of BioImaging (supported by POCI-01-0145-FEDER-022122), and the technical assistance of Maria Azevedo, and Genomics, and the technical assistance of Ana Mafalda Rocha and Rob Mensink. Sequencing data are a result of the GenomePT project (POCI-01-0145-FEDER-022184).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2021.689879/full#supplementary-material
Abbreviations
B2M, β2 Microglobulin; CD, Cluster of Differentiation; cDNA, Complementary DNA; CLEC7A, C-Type Lectin Domain Containing 7A; CR3, Complement Receptor 3; DAPI, 4′,6-diamidino-2-phenylindole; ELISA, Enzyme-Linked Immunosorbent Assay; FACS, Fluorescence-activated Cell Sorting; FITC, Fluorescein isothiocyanate; FMO, Fluorescence Minus One; FVD, Fixable Viability Dye; IgG, Immunoglobulin G; IL, Interleukine; LDH, Lactate dehydrogenase; LPS, Lipopolysaccharide; mAb, monoclonal Antibody; MARVELD1, Marvel Domain Containing 1; MHC, Major Histocompatibility Complex; NCBI, National Center for Biotechnology Information; P3C, Pam3CSK4 [Pam3Cys-Ser-(Lys)4]; PerCP-Cy5.5, Peridinin-chlorophyll protein-cychrome 5.5; PRR, Pattern Recognition Receptors; RNA, Ribonucleic Acid; ROS, Reactive Oxygen Species; RPE, R-Phycoerythrin; SEAP, Secreted Embryonic Alkaline Phosphatase; siRNA, Small Interference Ribonucleic Acid; PCR - Polymerase Chain Reaction; TAE, Tris acetate EDTA; TLR, Toll-like Receptor; TNF-α, Tumour Necrosis Factor Alpha; UNC - MISSION® siRNA Universal Negative Control #1; WGP-D, Whole Glucan Particles – Dispersible; WGP-S - Whole Glucan Particles – Soluble; Zym, Zymosan.
References
1. Broadway PR, Carroll JA, Sanchez NC. Live Yeast and Yeast Cell Wall Supplements Enhance Immune Function and Performance in Food-Producing Livestock: A Review (Dagger,)(Double Dagger). Microorganisms (2015) 3(3):417–27. doi: 10.3390/microorganisms3030417
2. Villot C, Chen Y, Pedgerachny K, Chaucheyras-Durand F, Chevaux E, Skidmore A, et al. Early Supplementation of Saccharomyces Cerevisiae Boulardii Cncm I-1079 in Newborn Dairy Calves Increases IgA Production in the Intestine At 1 Week of Age. J Dairy Sci (2020) 103(9):8615–28. doi: 10.3168/jds.2020-18274
3. Elghandour MMY, Tan ZL, Abu Hafsa SH, Adegbeye MJ, Greiner R, Ugbogu EA, et al. Saccharomyces Cerevisiae as a Probiotic Feed Additive to non and Pseudo-Ruminant Feeding: A Review. J Appl Microbiol (2020) 128(3):658–74. doi: 10.1111/jam.14416
4. Garcia Diaz T, Ferriani Branco A, Jacovaci FA, Cabreira Jobim C, Pratti Daniel JL, Iank Bueno AV, et al. Use of Live Yeast and Mannan-Oligosaccharides in Grain-Based Diets for Cattle: Ruminal Parameters, Nutrient Digestibility, and Inflammatory Response. PloS One (2018) 13(11):e0207127. doi: 10.1371/journal.pone.0207127
5. Lao EJ, Dimoso N, Raymond J, Mbega ER. The Prebiotic Potential of Brewers’ Spent Grain on Livestock’s Health: A Review. Trop Anim Health Prod (2020) 52(2):461–72. doi: 10.1007/s11250-019-02120-9
6. Nocek JE, Holt MG, Oppy J. Effects of Supplementation With Yeast Culture and Enzymatically Hydrolyzed Yeast on Performance of Early Lactation Dairy Cattle. J Dairy Sci (2011) 94(8):4046–56. doi: 10.3168/jds.2011-4277
7. White LA, Newman MC, Cromwell GL, Lindemann MD. Brewers Dried Yeast as a Source of Mannan Oligosaccharides for Weanling Pigs. J Anim Sci (2002) 80(10):2619–28. doi: 10.2527/2002.80102619x
8. Manners DJ, Masson AJ, Patterson JC. The Structure of a Beta-(1 Leads to 3)-D-Glucan From Yeast Cell Walls. Biochem J (1973) 135(1):19–30. doi: 10.1042/bj1350019
9. Camilli G, Tabouret G, Quintin J. The Complexity of Fungal β-Glucan in Health and Disease: Effects on the Mononuclear Phagocyte System. Front Immunol (2018) 9:673. doi: 10.3389/fimmu.2018.00673
10. Goodridge HS, Wolf AJ, Underhill DM. Beta-Glucan Recognition by the Innate Immune System. Immunol Rev (2009) 230(1):38–50. doi: 10.1111/j.1600-065X.2009.00793.x
11. Soltanian S, Stuyven E, Cox E, Sorgeloos P, Bossier P. Beta-Glucans as Immunostimulant in Vertebrates and Invertebrates. Crit Rev Microbiol (2009) 35(2):109–38. doi: 10.1080/10408410902753746
12. Gantner BN, Simmons RM, Canavera SJ, Akira S, Underhill DM. Collaborative Induction of Inflammatory Responses by Dectin-1 and Toll-Like Receptor 2. J Exp Med (2003) 197(9):1107–17. doi: 10.1084/jem.20021787
13. Ozinsky A, Underhill DM, Fontenot JD, Hajjar AM, Smith KD, Wilson CB, et al. The Repertoire for Pattern Recognition of Pathogens by the Innate Immune System Is Defined by Cooperation Between Toll-like Receptors. Proc Natl Acad Sci (2000) 97(25):13766. doi: 10.1073/pnas.250476497
14. Brown GD, Herre J, Williams DL, Willment JA, Marshall AS, Gordon S. Dectin-1 Mediates the Biological Effects of Beta-Glucans. J Exp Med (2003) 197(9):1119–24. doi: 10.1084/jem.20021890
15. Han B, Baruah K, Cox E, Vanrompay D, Bossier P. Structure-Functional Activity Relationship of Beta-Glucans From the Perspective of Immunomodulation: A Mini-Review. Front Immunol (2020) 11:658. doi: 10.3389/fimmu.2020.00658
16. Goodridge HS, Reyes CN, Becker CA, Katsumoto TR, Ma J, Wolf AJ, et al. Activation of the Innate Immune Receptor Dectin-1 Upon Formation of a ‘Phagocytic Synapse’. Nature (2011) 472(7344):471–5. doi: 10.1038/nature10071
17. Herre J, Marshall ASJ, Caron E, Edwards AD, Williams DL, Schweighoffer E, et al. Dectin-1 Uses Novel Mechanisms for Yeast Phagocytosis in Macrophages. Blood (2004) 104(13):4038–45. doi: 10.1182/blood-2004-03-1140
18. Underhill DM, Rossnagle E, Lowell CA, Simmons RM. Dectin-1 Activates Syk Tyrosine Kinase in a Dynamic Subset of Macrophages for Reactive Oxygen Production. Blood (2005) 106(7):2543–50. doi: 10.1182/blood-2005-03-1239
19. Goodridge HS, Simmons RM, Underhill DM. Dectin-1 Stimulation by Candida Albicans Yeast or Zymosan Triggers NFAT Activation in Macrophages and Dendritic Cells. J Immunol (2007) 178(5):3107–15. doi: 10.4049/jimmunol.178.5.3107
20. Qi C, Cai Y, Gunn L, Ding C, Li B, Kloecker G, et al. Differential Pathways Regulating Innate and Adaptive Antitumor Immune Responses by Particulate and Soluble Yeast-Derived Beta-Glucans. Blood (2011) 117(25):6825–36. doi: 10.1182/blood-2011-02-339812
21. De Marco Castro E, Calder PC, Roche HM. Beta-1,3/1,6-Glucans and Immunity: State of the Art and Future Directions. Mol Nutr Food Res (2020) 65(1):e1901071. doi: 10.1002/mnfr.201901071
22. Byrne KA, Loving CL, McGill JL. Innate Immunomodulation in Food Animals: Evidence for Trained Immunity? Front Immunol (2020) 11:1099. doi: 10.3389/fimmu.2020.01099
23. Eicher SD, Patterson JA, Rostagno MH. Beta-Glucan Plus Ascorbic Acid in Neonatal Calves Modulates Immune Functions With and Without Salmonella Enterica Serovar Dublin. Vet Immunol Immunopathol (2011) 142(3-4):258–64. doi: 10.1016/j.vetimm.2011.05.014
24. Angulo M, Reyes-Becerril M, Cepeda-Palacios R, Angulo C. Oral Administration of Debaryomyces Hansenii CBS8339-beta-glucan Induces Trained Immunity in Newborn Goats. Dev Comp Immunol (2020) 105:103597. doi: 10.1016/j.dci.2019.103597
25. de Oliveira CAF, Vetvicka V, Zanuzzo FS. Beta-Glucan Successfully Stimulated the Immune System in Different Jawed Vertebrate Species. Comp Immunol Microbiol Infect Dis (2019) 62:1–6. doi: 10.1016/j.cimid.2018.11.006
26. Willcocks S, Yamakawa Y, Stalker A, Coffey TJ, Goldammer T, Werling D. Identification and Gene Expression of the Bovine C-Type Lectin Dectin-1. Vet Immunol Immunopathol (2006) 113(1-2):234–42. doi: 10.1016/j.vetimm.2006.04.007
27. Byrne KA, Tuggle CK, Loving CL. Differential Induction of Innate Memory in Porcine Monocytes by Beta-Glucan or Bacillus Calmette-Guerin. Innate Immun (2020) 30:1753425920951607. doi: 10.1177/1753425920951607
28. Bekkering S, Blok BA, Joosten LA, Riksen NP, van Crevel R, Netea MG. In Vitro Experimental Model of Trained Innate Immunity in Human Primary Monocytes. Clin Vaccine Immunol (2016) 23(12):926–33. doi: 10.1128/CVI.00349-16
29. Corripio-Miyar Y, Hope J, McInnes CJ, Wattegedera SR, Jensen K, Pang Y, et al. Phenotypic and Functional Analysis of Monocyte Populations in Cattle Peripheral Blood Identifies a Subset With High Endocytic and Allogeneic T-Cell Stimulatory Capacity. Vet Res (2015) 46:112. doi: 10.1186/s13567-015-0246-4
30. Ye J, Coulouris G, Zaretskaya I, Cutcutache I, Rozen S, Madden TL. Primer-BLAST: A Tool to Design Target-Specific Primers for Polymerase Chain Reaction. BMC Bioinf (2012) 13:134. doi: 10.1186/1471-2105-13-134
31. Moyes KM, Drackley JK, Morin DE, Bionaz M, Rodriguez-Zas SL, Everts RE, et al. Gene Network and Pathway Analysis of Bovine Mammary Tissue Challenged With Streptococcus Uberis Reveals Induction of Cell Proliferation and Inhibition of PPARgamma Signaling as Potential Mechanism for the Negative Relationships Between Immune Response and Lipid Metabolism. BMC Genomics (2009) 10:542. doi: 10.1186/1471-2164-10-542
32. Galvao KN, Felippe MJ, Brittin SB, Sper R, Fraga M, Galvao JS, et al. Evaluation of Cytokine Expression by Blood Monocytes of Lactating Holstein Cows With or Without Postpartum Uterine Disease. Theriogenology (2012) 77(2):356–72. doi: 10.1016/j.theriogenology.2011.08.008
33. Mallikarjunappa S, Shandilya UK, Sharma A, Lamers K, Bissonnette N, Karrow NA, et al. Functional Analysis of Bovine Interleukin-10 Receptor Alpha in Response to Mycobacterium Avium Subsp. Paratuberculosis Lysate Using Crispr/Cas9. BMC Genet (2020) 21(1):121. doi: 10.1186/s12863-020-00925-4
34. Kadegowda AK, Bionaz M, Thering B, Piperova LS, Erdman RA, Loor JJ. Identification of Internal Control Genes for Quantitative Polymerase Chain Reaction in Mammary Tissue of Lactating Cows Receiving Lipid Supplements. J Dairy Sci (2009) 92(5):2007–19. doi: 10.3168/jds.2008-1655
35. Oliveira BM, Pinto A, Correia A, Ferreira PG, Vilanova M, Teixeira L. Characterization of Myeloid Cellular Populations in Mesenteric and Subcutaneous Adipose Tissue of Holstein-Friesian Cows. Sci Rep (2020) 10(1):1771. doi: 10.1038/s41598-020-58678-0
36. Schmittgen TD, Livak KJ. Analyzing Real-Time Pcr Data by the Comparative C(T) Method. Nat Protoc (2008) 3(6):1101–8. doi: 10.1038/nprot.2008.73
37. Willcocks S, Offord V, Seyfert HM, Coffey TJ, Werling D. Species-Specific PAMP Recognition by TLR2 and Evidence for Species-Restricted Interaction With Dectin-1. J Leukoc Biol (2013) 94(3):449–58. doi: 10.1189/jlb.0812390
38. Baert K, Sonck E, Goddeeris BM, Devriendt B, Cox E. Cell Type-Specific Differences in Beta-Glucan Recognition and Signalling in Porcine Innate Immune Cells. Dev Comp Immunol (2015) 48(1):192–203. doi: 10.1016/j.dci.2014.10.005
39. Elder MJ, Webster SJ, Chee R, Williams DL, Hill Gaston JS, Goodall JC. Beta-Glucan Size Controls Dectin-1-Mediated Immune Responses in Human Dendritic Cells by Regulating Il-1beta Production. Front Immunol (2017) 8:791. doi: 10.3389/fimmu.2017.00791
40. Dinh TTH, Tummamunkong P, Padungros P, Ponpakdee P, Boonprakong L, Saisorn W, et al. Interaction Between Dendritic Cells and Candida Krusei Beta-Glucan Partially Depends on Dectin-1 and It Promotes High Il-10 Production by T Cells. Front Cell Infect Microbiol (2020) 10:566661. doi: 10.3389/fcimb.2020.566661
41. Esteban A, Popp MW, Vyas VK, Strijbis K, Ploegh HL, Fink GR. Fungal Recognition Is Mediated by the Association of Dectin-1 and Galectin-3 in Macrophages. Proc Natl Acad Sci USA (2011) 108(34):14270–5. doi: 10.1073/pnas.1111415108
42. Angulo M, Reyes-Becerril M, Tovar-Ramirez D, Ascencio F, Angulo C. Debaryomyces Hansenii Cbs 8339 Beta-Glucan Enhances Immune Responses and Down-Stream Gene Signaling Pathways in Goat Peripheral Blood Leukocytes. Dev Comp Immunol (2018) 88:173–82. doi: 10.1016/j.dci.2018.07.017
43. Medina-Cordova N, Reyes-Becerril M, Ascencio F, Castellanos T, Campa-Cordova AI, Angulo C. Immunostimulant Effects and Potential Application of Beta-Glucans Derived From Marine Yeast Debaryomyces Hansenii in Goat Peripheral Blood Leucocytes. Int J Biol Macromol (2018) 116:599–606. doi: 10.1016/j.ijbiomac.2018.05.061
44. Sun Y, Liu J, Ye G, Gan F, Hamid M, Liao S, et al. Protective Effects of Zymosan on Heat Stress-Induced Immunosuppression and Apoptosis in Dairy Cows and Peripheral Blood Mononuclear Cells. Cell Stress Chaperones (2018) 23(5):1069–78. doi: 10.1007/s12192-018-0916-z
45. Ma T, Tu Y, Zhang N-f, Guo J-P, Deng K-D, Zhou Y, et al. Effects of Dietary Yeast β-Glucan on Nutrient Digestibility and Serum Profiles in Pre-Ruminant Holstein Calves. J Integr Agric (2015) 14(4):749–57. doi: 10.1016/s2095-3119(14)60843-1
46. Cherdthong A, Seankamsorn A, Suriyapha C, Chanjula P, Wanapat M. Effect of Beta-Glucan Supplementation on Feed Intake, Digestibility of Nutrients and Ruminal Fermentation in Thai Native Beef Cattle. J Anim Physiol Anim Nutr (Berl) (2018) 102(6):1509–14. doi: 10.1111/jpn.12989
47. Uchiyama H, Iwai A, Asada Y, Muramatsu D, Aoki S, Kawata K, et al. A Small Scale Study on the Effects of Oral Administration of the Beta-Glucan Produced by Aureobasidium Pullulans on Milk Quality and Cytokine Expressions of Holstein Cows, and on Bacterial Flora in the Intestines of Japanese Black Calves. BMC Res Notes (2012) 5:189. doi: 10.1186/1756-0500-5-189
48. Bounous DI, Enright FM, Gossett KA, Berry CM, Kearney MT. Comparison of Oxidant Production by Bovine Neutrophils and Monocyte-Derived Macrophages Stimulated With Brucella Abortus Strain 2308. Inflammation (1992) 16(3):215–25. doi: 10.1007/BF00918811
49. Gibson AJ, Woodman S, Pennelegion C, Patterson R, Stuart E, Hosker N, et al. Differential Macrophage Function in Brown Swiss and Holstein Friesian Cattle. Vet Immunol Immunopathol (2016) 181:15–23. doi: 10.1016/j.vetimm.2016.02.018
50. Bielefeldt Ohmann H, Babiuk LA. In Vitro Generation of Hydrogen Peroxide and of Superoxide Anion by Bovine Polymorphonuclear Neutrophilic Granulocytes, Blood Monocytes, and Alveolar Macrophages. Inflammation (1984) 8(3):251–75. doi: 10.1007/BF00916415
51. Kerrigan AM, Brown GD. Syk-Coupled C-type Lectin Receptors That Mediate Cellular Activation Via Single Tyrosine Based Activation Motifs. Immunol Rev (2010) 234(1):335–52. doi: 10.1111/j.0105-2896.2009.00882.x
52. Conejeros I, Patterson R, Burgos RA, Hermosilla C, Werling D. Induction of Reactive Oxygen Species in Bovine Neutrophils Is CD11b, But Not Dectin-1-Dependent. Vet Immunol Immunopathol (2011) 139(2-4):308–12. doi: 10.1016/j.vetimm.2010.10.021
53. van Bruggen R, Drewniak A, Jansen M, van Houdt M, Roos D, Chapel H, et al. Complement Receptor 3, Not Dectin-1, Is the Major Receptor on Human Neutrophils for Beta-Glucan-Bearing Particles. Mol Immunol (2009) 47(2-3):575–81. doi: 10.1016/j.molimm.2009.09.018
54. Werling D, Hope JC, Howard CJ, Jungi TW. Differential Production of Cytokines, Reactive Oxygen and Nitrogen by Bovine Macrophages and Dendritic Cells Stimulated With Toll-Like Receptor Agonists. Immunology (2004) 111(1):41–52. doi: 10.1111/j.1365-2567.2004.01781.x
55. Li Z, Kang H, You Q, Ossa F, Mead P, Quinton M, et al. In Vitro Bioassessment of the Immunomodulatory Activity of Saccharomyces Cerevisiae Components Using Bovine Macrophages and Mycobacterium Avium Ssp. Paratuberculosis. J Dairy Sci (2018) 101(7):6271–86. doi: 10.3168/jds.2017-13805
56. Li B, Cramer D, Wagner S, Hansen R, King C, Kakar S, et al. Yeast Glucan Particles Activate Murine Resident Macrophages to Secrete Proinflammatory Cytokines Via MyD88- and Syk Kinase-Dependent Pathways. Clin Immunol (2007) 124(2):170–81. doi: 10.1016/j.clim.2007.05.002
57. Javmen A, Nemeikaitė-Čėnienė A, Grigiškis S, Lysovienė J, Jonauskienė I, Šiaurys A, et al. The Effect of Saccharomyces Cerevisiae β-Glucan on Proliferation, Phagocytosis and Cytokine Production of Murine Macrophages and Dendritic Cells. Biologia (2017) 72(5):561–8. doi: 10.1515/biolog-2017-0063
58. Noss I, Doekes G, Thorne PS, Heederik DJ, Wouters IM. Comparison of the Potency of a Variety of β-Glucans to Induce Cytokine Production in Human Whole Blood. Innate Immun (2013) 19(1):10–9. doi: 10.1177/1753425912447129
59. Sonck E, Stuyven E, Goddeeris B, Cox E. The Effect of Beta-Glucans on Porcine Leukocytes. Vet Immunol Immunopathol (2010) 135(3-4):199–207. doi: 10.1016/j.vetimm.2009.11.014
60. Hontelez S, Sanecka A, Netea MG, van Spriel AB, Adema GJ. Molecular View on PRR Cross-Talk in Antifungal Immunity. Cell Microbiol (2012) 14(4):467–74. doi: 10.1111/j.1462-5822.2012.01748.x
61. Takahara K, Tokieda S, Nagaoka K, Inaba K. Efficient Capture of Candida Albicans and Zymosan by SIGNR1 Augments TLR2-Dependent Tnf-Alpha Production. Int Immunol (2012) 24(2):89–96. doi: 10.1093/intimm/dxr103
62. Dillon S, Agrawal S, Banerjee K, Letterio J, Denning TL, Oswald-Richter K, et al. Yeast Zymosan, a Stimulus for TLR2 and Dectin-1, Induces Regulatory Antigen-Presenting Cells and Immunological Tolerance. J Clin Invest (2006) 116(4):916–28. doi: 10.1172/JCI27203
63. Yanagawa Y, Onoe K. Enhanced IL-10 Production by TLR4- and TLR2-Primed Dendritic Cells Upon TLR Restimulation. J Immunol (2007) 178(10):6173–80. doi: 10.4049/jimmunol.178.10.6173
64. Bose N, Chan AS, Guerrero F, Maristany CM, Qiu X, Walsh RM, et al. Binding of Soluble Yeast Beta-Glucan to Human Neutrophils and Monocytes Is Complement-Dependent. Front Immunol (2013) 4:230. doi: 10.3389/fimmu.2013.00230
65. Huang H, Ostroff GR, Lee CK, Agarwal S, Ram S, Rice PA, et al. Relative Contributions of Dectin-1 and Complement to Immune Responses to Particulate Beta-Glucans. J Immunol (2012) 189(1):312–7. doi: 10.4049/jimmunol.1200603
66. van der Meer JW, Joosten LA, Riksen N, Netea MG. Trained Immunity: A Smart Way to Enhance Innate Immune Defence. Mol Immunol (2015) 68(1):40–4. doi: 10.1016/j.molimm.2015.06.019
67. Netea MG, Dominguez-Andres J, Barreiro LB, Chavakis T, Divangahi M, Fuchs E, et al. Defining Trained Immunity and Its Role in Health and Disease. Nat Rev Immunol (2020) 20(6):375–88. doi: 10.1038/s41577-020-0285-6
68. Garcia-Valtanen P, Guzman-Genuino RM, Williams DL, Hayball JD, Diener KR. Evaluation of Trained Immunity by Beta-1, 3 (D)-Glucan on Murine Monocytes In Vitro and Duration of Response In Vivo. Immunol Cell Biol (2017) 95(7):601–10. doi: 10.1038/icb.2017.13
69. Paris S, Chapat L, Pasin M, Lambiel M, Sharrock TE, Shukla R, et al. Beta-Glucan-Induced Trained Immunity in Dogs. Front Immunol (2020) 11:566893. doi: 10.3389/fimmu.2020.566893
Keywords: dectin-1, β-glucans, bovine, monocytes, cytokines, siRNA, CLEC7A
Citation: Pedro ARV, Lima T, Fróis-Martins R, Leal B, Ramos IC, Martins EG, Cabrita ARJ, Fonseca AJM, Maia MRG, Vilanova M and Correia A (2021) Dectin-1-Mediated Production of Pro-Inflammatory Cytokines Induced by Yeast β-Glucans in Bovine Monocytes. Front. Immunol. 12:689879. doi: 10.3389/fimmu.2021.689879
Received: 01 April 2021; Accepted: 13 May 2021;
Published: 28 May 2021.
Edited by:
Nicolas Bertho, INRA Biologie, Épidémiologie et Analyse de Risque en santé animale (BIOEPAR), FranceReviewed by:
Karelle De Luca, Boehringer Ingelheim, FranceKristen Alicia Byrne, National Animal Disease Center (USDA ARS), United States
Copyright © 2021 Pedro, Lima, Fróis-Martins, Leal, Ramos, Martins, Cabrita, Fonseca, Maia, Vilanova and Correia. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alexandra Correia, alexandra.correia@ibmc.up.pt