Corrigendum: Haplotype Motif-Based Models for KIR-Genotype Informed Selection of Hematopoietic Cell Donors Fail to Predict Outcome of Patients With Myelodysplastic Syndromes or Secondary Acute Myeloid Leukemia
 Johannes Schetelig1,2*
Johannes Schetelig1,2* Henning Baldauf2
Henning Baldauf2 Linda Koster3Michelle Kuxhausen4
Linda Koster3Michelle Kuxhausen4 Falk Heidenreich1,2
Falk Heidenreich1,2 Liesbeth C. de Wreede2,5
Liesbeth C. de Wreede2,5 Stephen Spellman4Michel van Gelder6
Stephen Spellman4Michel van Gelder6 Benedetto Bruno7Francesco Onida8
Benedetto Bruno7Francesco Onida8 Vinzenz Lange9Carolin Massalski9Victoria Potter10
Vinzenz Lange9Carolin Massalski9Victoria Potter10 Per Ljungman11Nicolaas Schaap12Patrick Hayden13Stephanie J. Lee14
Per Ljungman11Nicolaas Schaap12Patrick Hayden13Stephanie J. Lee14 Nicolaus Kröger15
Nicolaus Kröger15 Kathy Hsu16
Kathy Hsu16 Alexander H. Schmidt2,9
Alexander H. Schmidt2,9 Ibrahim Yakoub-Agha17
Ibrahim Yakoub-Agha17 Marie Robin18
Marie Robin18- 1Medizinische Klinik und Poliklinik I, University Hospital Dresden, Dresden, Germany
- 2DKMS Clinical Trials Unit, Dresden, Germany
- 3EBMT Data Office Leiden, Leiden, Netherlands
- 4Center for International Blood and Marrow Transplant Research, Minneapolis, MN, United States
- 5Leiden University Medical Center, Department of Biomedical Data Sciences, Leiden, Netherlands
- 6Maastricht University Medical Center, Department of Internal Medicine, Maastricht, Netherlands
- 7A.O.U. Citta della Salute e della Scienza di Torino, Turin, Italy
- 8Fondazione IRCCS Ca’Granda Ospedale Maggiore Policlinico, University of Milan, Milan, Italy
- 9DKMS Life Science Lab, Dresden, Germany
- 10GKT School of Medicine, London, United Kingdom
- 11Karolinska University Hospital and Karolinska Institutet, Stockholm, Sweden
- 12Radboud University Medical Centre, Nijmegen, Netherlands
- 13St. James’s Hospital, Dublin, Ireland
- 14Fred Hutchinson Cancer Research Center, Seattle, WA, United States
- 15University Hospital Eppendorf, Hamburg, Germany
- 16Memorial Sloan Kettering Cancer Center, New York & Scientific Director, CIBMTR Immunobiology Working Committee, New York City, NY, United States
- 17CHU de Lille, LIRIC, INSERM U995, Université de Lille, Lille, France
- 18Hopital Saint-Louis, APHP, Université de Paris, Paris, France
Results from registry studies suggest that harnessing Natural Killer (NK) cell reactivity mediated through Killer cell Immunoglobulin-like Receptors (KIR) could reduce the risk of relapse after allogeneic Hematopoietic Cell Transplantation (HCT). Several competing models have been developed to classify donors as KIR-advantageous or disadvantageous. Basically, these models differ by grouping donors based on distinct KIR–KIR–ligand combinations or by haplotype motif assignment. This study aimed to validate different models for unrelated donor selection for patients with Myelodysplatic Syndromes (MDS) or secondary Acute Myeloid Leukemia (sAML). In a joint retrospective study of the European Society for Blood and Marrow Transplantation (EBMT) and the Center for International Blood and Marrow Transplant Research (CIBMTR) registry data from 1704 patients with secondary AML or MDS were analysed. The cohort consisted mainly of older patients (median age 61 years) with high risk disease who had received chemotherapy-based reduced intensity conditioning and anti-thymocyte globulin prior to allogeneic HCT from well-matched unrelated stem cell donors. The impact of the predictors on Overall Survival (OS) and relapse incidence was tested in Cox regression models adjusted for patient age, a modified disease risk index, performance status, donor age, HLA-match, sex-match, CMV-match, conditioning intensity, type of T-cell depletion and graft type. KIR genes were typed using high-resolution amplicon-based next generation sequencing. In univariable and multivariable analyses none of the models predicted OS and the risk of relapse consistently. Our results do not support the hypothesis that optimizing NK-mediated alloreactivity is possible by KIR-genotype informed selection of HLA-matched unrelated donors. However, in the context of allogeneic transplantation, NK-cell biology is complex and only partly understood. KIR-genes are highly diverse and current assignment of haplotype motifs based on the presence or absence of selected KIR genes is over-simplistic. As a consequence, further research is highly warranted and should integrate cutting edge knowledge on KIR genetics, and NK-cell biology into future studies focused on homogeneous groups of patients and treatment modalities.
Introduction
To harness natural killer (NK) cells for graft-versus-leukemia reactions by selecting donors based on KIR genotype information could further improve outcome after allogeneic hematopoietic cell transplantation (alloHCT). Evidence for the potential of allogeneic NK cells to attack cancer cells comes from a series of clinical studies demonstrating activity of haploidentical NK cell infusion or transplantation for patients with relapsed or refractory acute myeloid leukemia (AML) or myelodysplastic syndrome (MDS) (1–5). NK cytotoxicity may be triggered by activating Killer Immunoglobulin like Receptors (KIRs), which encounter their cognate ligand on target cells, or by inhibitory KIRs, which do not encounter their cognate ligands on target cells. KIR–KIR–ligand interactions may elicit NK-alloreactivity also in the setting of HLA-compatible related or unrelated donor transplantation. This was supported by a series of retrospective registry studies which reported associations between certain donor-patient KIR genotype patterns and the risk of relapse after alloHCT (6–13).
The human KIR region has a complex architecture and comprises 15 genes and 2 pseudogenes. Six KIR genes (KIR2DL1, KIR2DL2/3, KIR2DL5, KIR3DL1, KIR3DL2, KIR3DL3) encode inhibitory receptors with long (L) cytoplasmic tails, while six genes encode receptors with short (S) activating cytoplasmic tails (KIR2DS1, KIR2DS2, KIR2DS3, KIR2DS4, KIR2DS5, KIR3DS1). Two pseudogenes are not expressed as proteins (KIR2DP1, KIR3DP1). Four genes are present in most of the common haplotypes and have been designated as “framework” genes (KIR3DL3, KIR3DP1, KIR2DL4, KIR3DL2) (14, 15). KIR genes exhibit substantial allelic diversity (16). Furthermore, KIR haplotypes vary with respect to the presence or absence of specific KIR genes and are subject to copy number variation (17, 18).
Additionally, when improved sequencing technology allowed for allele-level resolution of KIR genes it became clear that absence/presence typing was not sufficient to determine the functional status of KIRs (18, 19). For example, the third most common allele of KIR3DL1, KIR3DL1*004, accounts for 17% of all KIR3DL1 genes but is not expressed on the cell surface. KIR3DL1 allotypes differ with respect to their expression patterns (20). In the context of HIV infection, certain subtype combinations of KIR3DL1 allotypes together with their cognate ligand Bw4 were strongly associated with the risk of progression to AIDS (21). Genetic information on presence versus absence alone might thus lead to wrong conclusions about function. However, individual KIR genes show extensive sequence polymorphism with 1110 alleles currently named in total (IPD-KIR Database, Release 2.9.0 as of July 2020) (22). Allelic diversity across genes ranges from 16 alleles for KIR2DS3 and KIR2DS1 to 183 alleles for KIR3DL1. As a consequence, the development of cutting edge typing technology that could generate allelic and copy number results was critical to move the research field forward (16).
Against this background we set out to validate an HLA-matched unrelated donor selection algorithm for patients with AML and MDS which was essentially aimed at reducing the inhibitory potential of donor NK cells and increasing the activating potential by donor selection based on information on KIR2DS1 and KIR3DL1 (12, 13). We recently reported results of a study in AML patients where we failed to replicate the findings of the original report (23). Here, we present the data on patients with MDS or secondary AML. Again, we were unable to confirm the KIR3DL1/KIR2DS1- based donor selection algorithm. In addition, we tested other major models to predict the risk of relapse and death based on donor KIR genotype information.
Methods
Inclusion Criteria
We conducted a joint study of the European Society for Blood and Marrow Transplantation (EBMT) and the Center for International Blood and Marrow Transplant Research (CIBMTR) on the impact of KIR genotype information on patient outcome after alloHCT for MDS or secondary AML. This study used DNA samples from stem cell donors which were stored at the Collaborative Biobank (www.cobi-biobank.de). All stem cell donors had provided written informed consent when they contributed a sample to the biobank. The study was approved by the responsible Ethical Committee at the Technische Universtität Dresden, Germany. Access to medical data was approved by the Review Boards of the Chronic Malignancies Working Party of EBMT, the Immunobiology Working Committee of the CIBMTR and the National Marrow Donor Program Institutional Review Board.
Patient inclusion criteria were first allogeneic HCT from an unrelated donor between January 2008 and December 2017, a diagnosis of Myelodysplastic Syndrome (MDS) or secondary AML at HCT, and age above 18 years with an available donor sample in the Collaborative Biobank.
Sample Identity
Donor information was mapped to the medical data of the patient using the Donor ID as a key. In order to rule out errors during the mapping process, all donor samples were typed for HLA and KIR genes. Information on the HLA-genotype was used to double-check sample identity by comparing the typing result with the original typing results for that donor and by checking HLA-compatibility with the corresponding patient information. The HLA compatibility between donors and recipients was assessed based on two-field information for HLA-A, -B, -C, -DRB1 and -DQB1. Donor-recipient pairs, whose HLA-compatibility could not be confirmed, were excluded.
KIR Genotyping
Genotyping was performed using a high-resolution short-amplicon-based next generation sequencing workflow. KIR typing at the allele-level was based on sequencing of exons 3, 4, 5, 7, 8, and 9 and subsequent bioinformatic analysis as described previously (16).
Classification Models Into KIR Advantageous and Disadvantageous Donors
HLA-C alleles were grouped in C1 and C2 ligands and HLA-B alleles were grouped into Bw4-80I/Bw-80T/Bw6 epitope bearing ligands based on information retrieved from https://www.ebi.ac.uk/ipd/kir/ligand.html. Information on KIR3DL1 and KIR2DS1 and their cognate ligands was grouped according to publications by Venstrom et al. (12) and Boudreau et al. (13). Further, we classified donors according to A versus B haplotype motifs using definitions for haplotype assignment as provided by Cooley et al. (10, 11). Finally, we calculated scores for selected additive models which integrate information on KIR-ligand combinations of donor-recipient pairs. We calculated the functional inhibitory KIR count by assigning a score of 1 for donor KIR2DL1, KIR2DL2, KIR2DL3, and KIR3DL1 when the cognate ligands were exhibited by patient HLA molecules as described in the original paper by Boelen et al. (24). As an extension of this count we also calculated the weighted inhibitory score using the published weights for functional KIR-ligand pairs as follows: Inhibitory score = (1 if functional KIR2DL1) + (1 if strong functional KIR2DL2 or 0.5 if weak functional KIR2DL2) + (0.75 if functional KIR2DL3) + (1 if functional KIR3DL1) (24).
The score developed by Krieger et al. integrates information on inhibitory and activating KIR-ligand interactions (25). Two versions exist, a non-weighted version which incorporates the inhibitory missing KIR-ligand Score (IM-KIR Score) with assigned scores per interaction, and a weighted version (w-KIR Score). Both versions were calculated according to the original publication (25).
Information Used for Risk Adjustment
MDS and sAML were grouped by adopting definitions from the World Health Organization classification of myeloid neoplasms and acute leukemia (26).
Using information on the genetic risk and disease stage at transplantation from EBMT Minimal Essential Data Forms, we calculated a simplified Disease Risk Index (DRI) for MDS and sAML. For this purpose, cytogenetic risk was classified according to the rules for the refined DRI (27) except for chromosome 17p abnormalities which were assigned to the adverse risk group. For patients with missing stage, disease or cytogenetic risk information, DRI group was imputed based on largest frequencies reported in the publication of the refined DRI. The intensity of conditioning regimens was classified according to working definitions of EBMT and CIBMTR (28).
Risk adjustment in the context of multivariable regression models included information on patients’ performance status, age, sex, CMV serostatus, disease risk index, conditioning intensity, T-cell depletion, HLA-matcing, donor age, donor sex, and donor CMV serostatus.
Primary Endpoint and Power Considerations
Event-free Survival (EFS) was selected as primary endpoint and death, relapse or progression (whichever occurred first) were defined as events for EFS. The study was designed to validate the effect of the classification of donor KIR2DS1 and KIR3DL1 information on predicting EFS as published by Boudreau et al. (13). In sample size estimations for the comparison of strongly inhibiting versus weakly inhibiting donor-patient KIR3DL1 HLA-B subtype combinations, we calculated that data from approximately 1,700 patients were required to detect a 17% reduction in EFS events with a two-sided type I error of 5% at a power of 80%. Since we failed to validate this model in patients with AML (23), we extended the scope of this study and also evaluated alternative models for donor KIR genotype classification. No formal adjustment of the type I error was made for multiple testing for these exploratory analyses. The post hoc power for each comparison was calculated with Schoenfeld’s formula based on the given number of events and the reported effect estimates, with a two-sided 5% type I error probability (29).
Statistical Analysis
Relapse or progression was selected as the major secondary endpoint. Additional endpoints were Non-Relapse Mortality and Overall Survival. Death without previous relapse or progression was defined as non-relapse death. EFS and Overall Survival (OS) probabilities were calculated with the Kaplan-Meier estimator and between-group comparisons were performed with the log-rank test. Relapse/progression and Non-Relapse Mortality were considered as competing risks and univariable comparisons built on cumulative incidence curves. Univariable comparisons for these endpoints were performed with the Gray Test. All time-to-event endpoints were evaluated in (cause-specific) multivariable Cox proportional hazards regression models. Effect sizes were reported as hazard ratios together with 95%-confidence intervals. We performed subgroup analyses for the effect of KIR genotype-based classifications in subpopulations defined by variations in the transplant procedure which could have an impact on NK alloreactivity, e.g. myeloablative conditioning versus reduced-intensity conditioning/non-myeloablative conditioning, use of Anti-Thymocyte Globulin (ATG) and of total body irradiation.
The proportionality assumption was checked for each covariable for the main models analyzing Overall Survival and relapse by means of plots of scaled Schoenfeld residuals and the test of Grambsch and Therneau (30).
Results
Patient Characteristics
Mapping of patients and donors resulted in 1,836 donor-recipient pairs. Data from three donor-recipient pairs were excluded because sample identity could not be confirmed. Typing of 108 samples failed because the DNA quantity or quality was too low for the workflow. No data on outcomes was available for 21 patients. The final statistical analysis set thus contained information on 1,704 patients.
The median age at allogeneic HCT was 61 years (range from 18 to 83 years). Indication for allogeneic HCT was secondary AML for 28% of patients, MDS for 63% of patients and MDS/MPN overlap syndromes in 9% of patients. Disease risk was assessed as intermediate, high or very high in 41, 58, and 0.6%, respectively. Patient and donor pairs were 10/10 matched in 79% of pairs, whereas a one locus mismatch was reported for 20% of pairs. Myeloablative, reduced-intensity and non-myeloablative conditioning regimens were used in 31, 56, and 11% of patients, respectively. Anti-Thymocyte globulin was administered as GVHD-prophylaxis in 56% and Alemtuzumab in 9% of patients. Thirty five percent of patients received no T-cell depletion, and 1% received an ex vivo T-cell depleted graft. Peripheral Blood Stem Cells (PBSC) and Bone Marrow (BM) were used as graft source in 93 and 7% of patients, respectively. Further details and the distribution of patient characteristics are given in Table 1.
TABLE 1
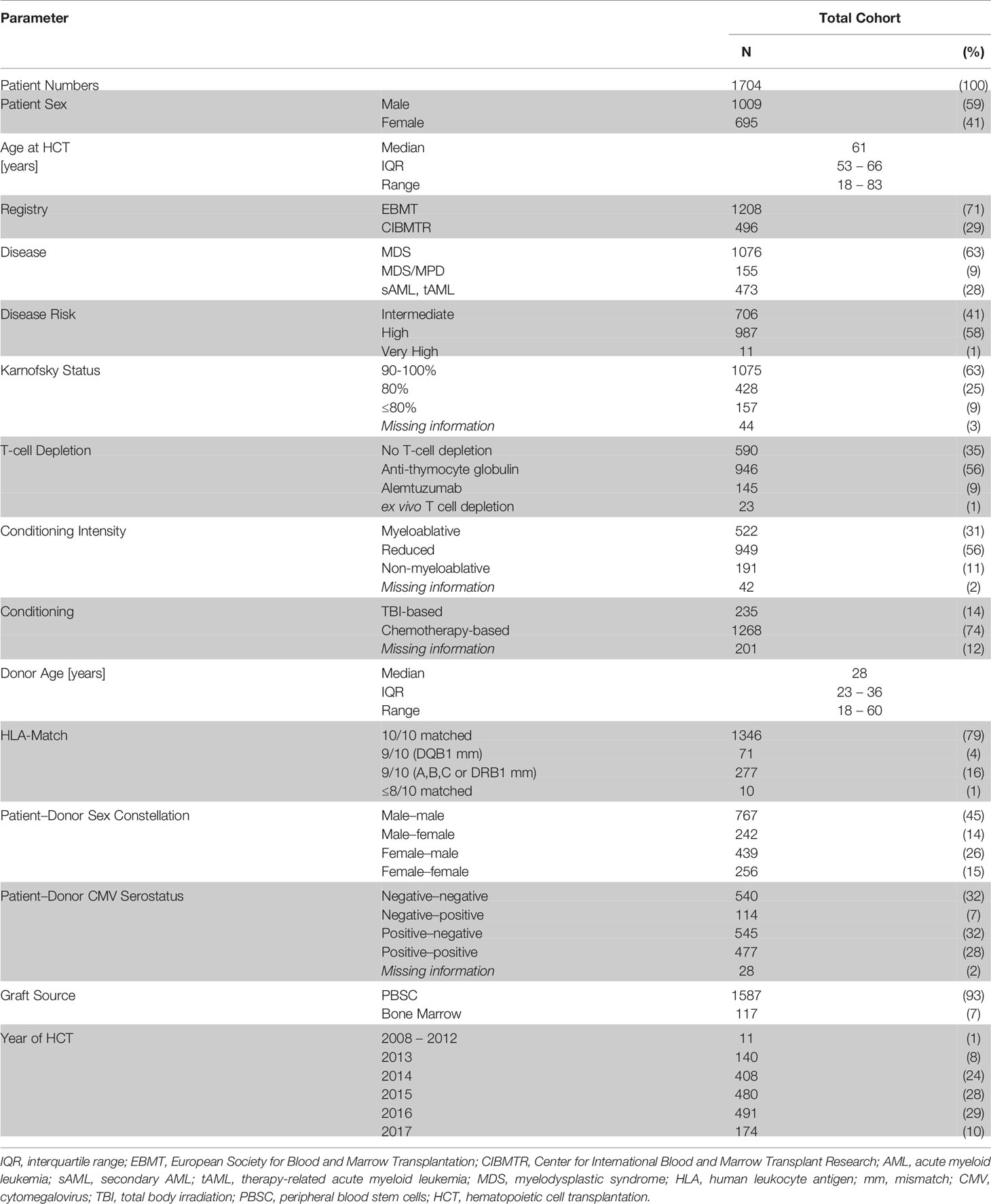
Table 1 Patient and treatment characteristics.
For the whole cohort, 2-year probabilities were 48% (95%-CI: 46–51%) for OS, 42% (95%-CI: 39–44%) for EFS, 29% (95%-CI: 27–32%) for relapse incidence and 29% (95%-CI: 27–32%) for Non-Relapse Mortality. In total, 451 relapses and 450 non-relapse deaths were recorded. Altogether, 780 deaths were reported, including 330 deaths after relapse.
KIR—Ligand Models
First, models integrating information on activating or non-activating KIR2DS1 and various degrees of KIR3DL1-mediated inhibition were tested for their ability to predict the risk of relapse and EFS (12, 13). Basically, these models were built on the idea that strong inhibitory KIR-ligand interactions increase the risk of relapse while activating KIR-ligand interactions reduce it. The results of multivariable modelling are summarized in Table 2. Detailed information on the set of factors used for risk adjustment is provided in Supplemental Table 1. No significant differences for the risk of relapse were found for the respective KIR3DL1/HLA-B subtype combinations and KIR2DS1/C1C2 epitope combinations. Notably, in contrast to the original publications for these models, neither patients whose donors had non-inhibiting KIR3DL1-Ligand interaction (HR 1.08, 95%CI 0.86–1.36; p=.5) nor patients with activating KIR2DS1-Ligand interaction (HR 1.11, 95%CI 0.87–1.16; p=0.9) showed a lower risk of relapse. Cumulative incidence curves for relapse and Kaplan-Meier plots for EFS are shown in Figures 1 and 2 for the respective groups of patients.
TABLE 2
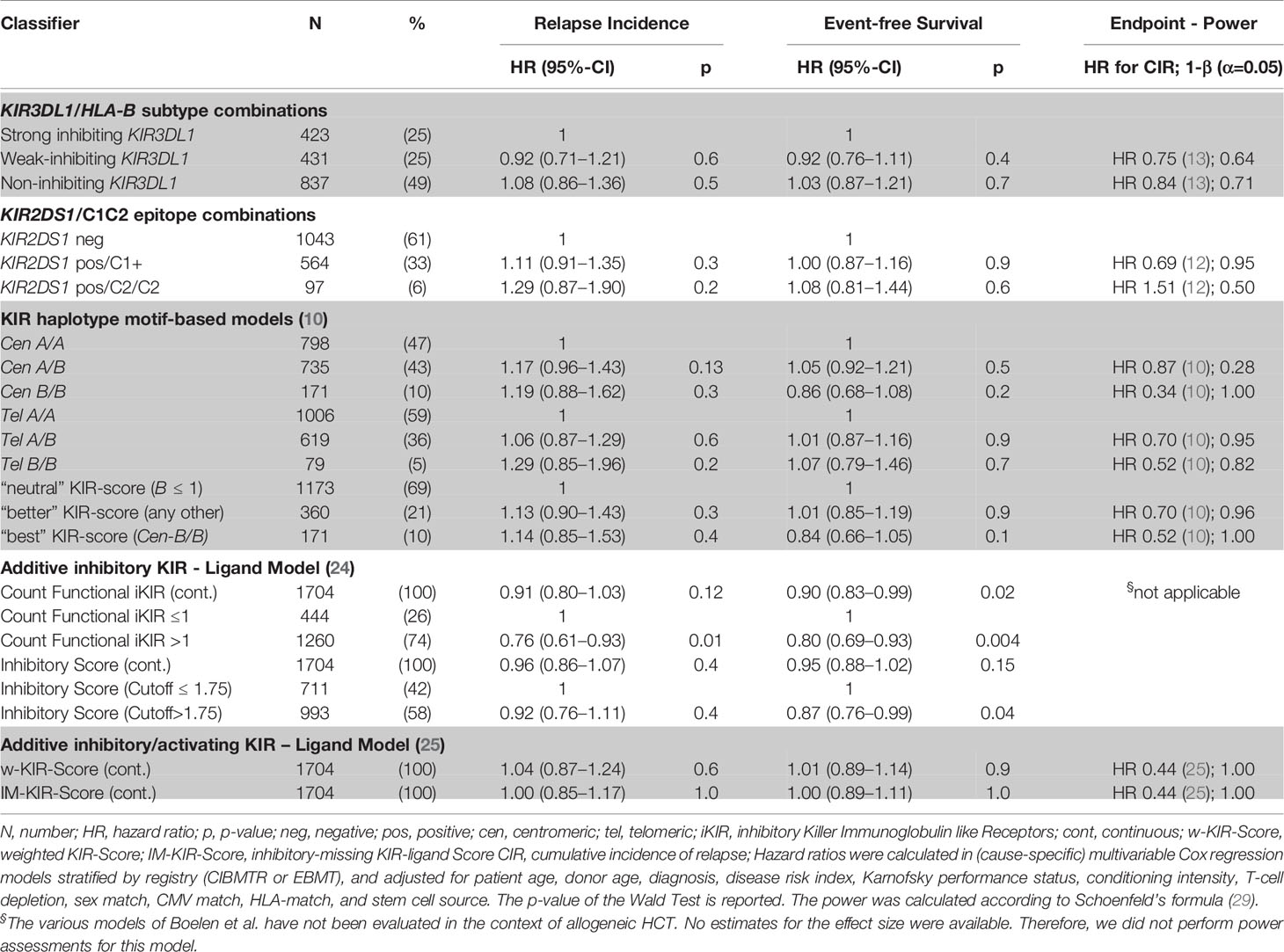
Table 2 Impact of donor KIR genotype models on relapse incidence and event-free survival.
FIGURE 1
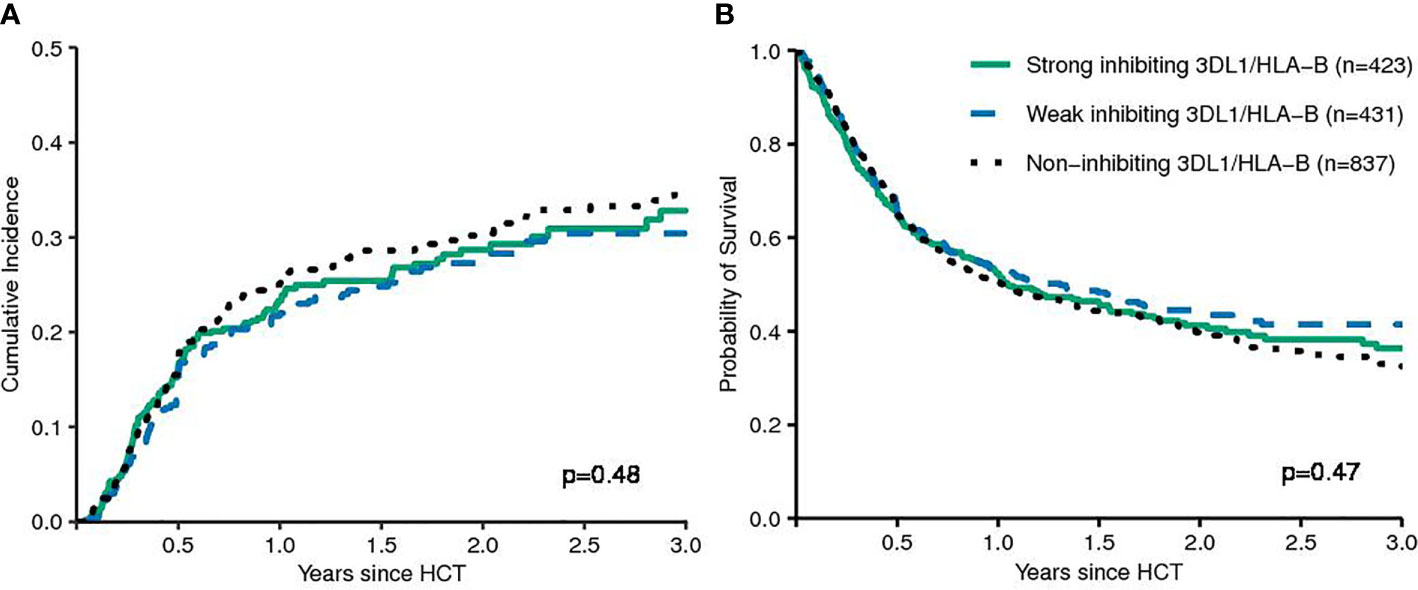
Figure 1 Patients were grouped by their donors’ KIR3DL1/HLA-B subtype combinations (strong inhibiting versus non-inhibiting/weak inhibiting) in terms of relapse incidence (A) and event-free survival (B). The p-values have been calculated by the Gray Test (CIR) and the log-rank test (EFS).
FIGURE 2
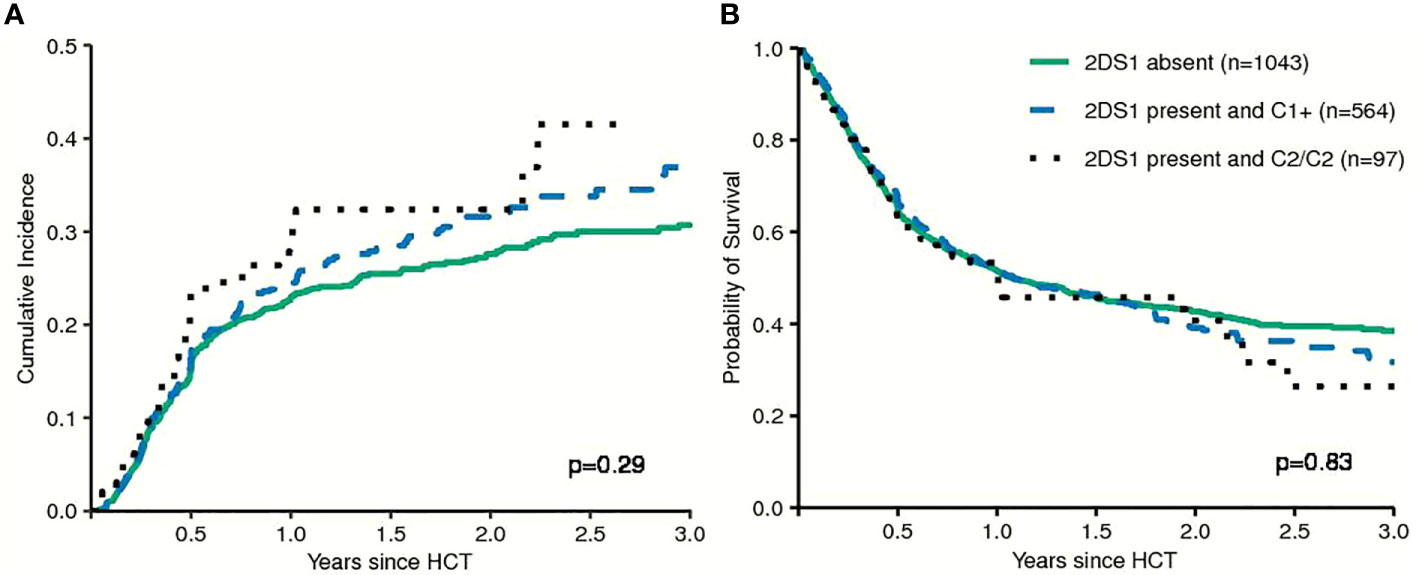
Figure 2 Patients were grouped by their donors' KIR2DS1 status (activating versus non-activating) in terms of Relapse Incidence (A) and Event-free Survival (B). The p-values have been calculated by the Gray Test (Cumulative Incidence of Relapse) and the log-rank test (Event-free Survival).
Haplotype Motif-Based Prediction
Next, we tested models which utilize information on the different composition of KIR haplotypes with respect to activating and inhibitory KIR genes. The KIR A haplotypes are more conserved and contain largely inhibitory KIR genes, whereas the B haplotypes display greater variation in gene content and may include a variety of activating receptors. Some studies suggested that patients whose donors had two B haplotypes had a lower risk of relapse compared to patients whose donors had two A haplotypes (9–11). In our data we did not find a significant impact of telomeric or centromeric KIR haplotype A versus B motifs. Compared to patients, whose donors had centromeric KIR A/A motifs, patients, whose donors had centromeric KIR B/B motifs, even tended to a greater risk of relapse (HR 1.19, 95%CI 0.88–1.62; p=.3). Cumulative incidence curves for relapse and Kaplan-Meier plots for EFS are shown in Figure 3 for the respective grouping of patients according to centromeric donor KIR haplotypes. In contrast to the model, we also found a trend for a greater risk of relapse (HR 1.29, 95%CI 0.85–1.96; p=0.2) for patients with telomeric KIR B/B donors compared to telomeric KIR A/A donors. Table 2 summarizes the results.
FIGURE 3
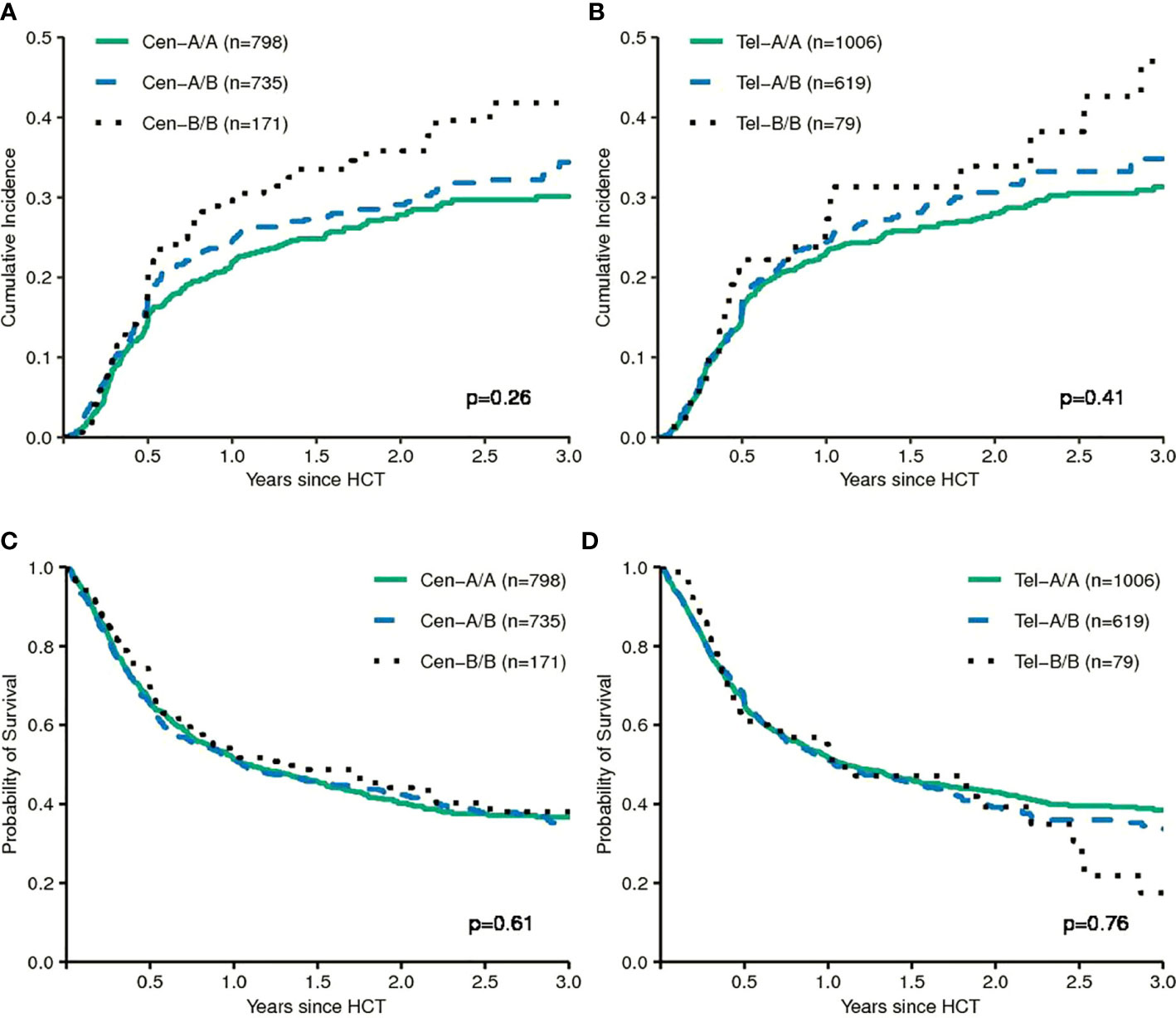
Figure 3 The upper panels show the cumulative incidences of relapse of patients grouped according to centromeric KIR motifs (A) and grouped according to telomeric KIR motifs (B). The lower panels show event-free survival of patients grouped according to centromeric KIR motifs (C) and grouped according to telomeric KIR motifs (D). The p-values have been calculated by the Gray Test (Cumulative Incidence of Relapse) and the log-rank test (Event-free Survival).
Additive Models
Finally, we tested models which integrate information on inhibitory or activating KIR-ligand interactions using a scoring system. Boelen et al. developed an inhibitory score which takes information on functional inhibitory KIR-ligand interactions into account and demonstrated uniform effects of the score on the progression of HIV, HCV, and HTLV infections (24). In the context of these viral infections, high inhibitory scores were mainly associated with protective effects. The weighted inhibitory score did not predict the risk of relapse in our study. However, the functional inhibitory KIR count was significantly associated with the risk of relapse (HR 0.76, 95%CI 0.61–0.93; p=0.01) and EFS (HR 0.8, 95%CI 0.69–0.93; p=0.004), when dichotomized at a cutoff of 1. This comparison indicated that donor genotypes encoding more functionally relevant inhibitory KIR were associated with a lower risk of relapse.
The score proposed by Krieger et al. integrates information on potential inhibitory KIR-ligand interactions and activating KIR-ligand interactions (25). Neither the simple score nor the weighted score predicted the risk of relapse or EFS in our cohort.
Subgroup Analyses
We also calculated KIR haplotype motif-based models for four major subgroups: patients with secondary AML, patients with MDS, patients who had received myeloablative conditioning and patients who had received non-myeloablative or reduced-intensity conditioning. In a series of exploratory analyses we tested the main haplotype motif-based classifications for these subgroups. Results are shown in Supplemental Table 2A. The effects of centromeric and telomeric KIR haplotype B/B motifs compared to haplotype A/A motifs pointed in different directions for patients with secondary AML (HR 0.7, 95%–CI 0.4–1.33) and patients with MDS (HR 1.48, 95%–CI 1.03–2.11). The estimated effects for donor KIR haplotype motifs for patients who had received reduced-intensity or non-myeloablative conditioning also differed slightly from patients who had received myeloablative conditioning. Further, we performed subgroup analyses in patients with at least one C1 epitope, thereby excluding C2/C2 patients, which showed essentially in the same pattern of results (Supplemental Table 2B).
Discussion
We investigated the potential of donor KIR genotype information to predict the risk of relapse after HLA-compatible unrelated allogeneic HCT in a large cohort of patients with secondary acute myeloid leukemia (sAML) or a myelodysplastic syndrome (MDS). To date, few studies have analysed the impact of KIR genotype on the course of the disease for patients with MDS or sAML after HLA-compatible alloHCT (6, 31, 32). Since patient numbers in these studies were small, no meaningful subgroup analyses for patients with MDS or sAML were performed. Therefore, our results should be discussed in the context of studies on patients with AML.
This study was designed as a validation study for the KIR2DS1/KIR3DL1-based prediction model in patients with MDS and sAML. This model claims that the risk of relapse could be reduced by increasing the activating potential through preferentially selecting unrelated donors with activating KIR2DS1 and by minimizing the inhibitory potential mediated through selecting KIR3DL1/HLA-B subtype combinations with minimal or no inhibitory potential (12, 13). We were not able to validate this model for patients with MDS or sAML. Although not statistically significant, unrelated donor-patient pairs with activating KIR2DS1 even had a slightly higher actual risk of relapse indicated by a hazard ratio of 1.11 (Wald test, p=0.3) compared to patients whose donors where KIR2DS1 negative. Contrary to the previously observed effects, donor-patient pairs with non-inhibiting KIR3DL1/HLA-B subtype combinations also had the highest actual risk of relapse with a hazard ratio of 1.08 (Wald test, p=0.5) in multivariable cause-specific Cox regression modelling (see Table 2 and Figure 1). In summary, our study did not align with the pattern of results originally reported. In a recently published study where we attempted to validate the proposed model for patients with AML, the observed data also failed to confirm the proposed prediction model (23). Together, in these two studies data from 3926 patients with AML or MDS were analysed whose HLA-compatible unrelated donors had been typed for KIR genes at the allele-level. However, it should be noted that patient characteristics and transplant procedures of the original cohorts differed substantially compared to the patients whose data were analysed here. In our contemporary validation studies the median patient age was approximately 20 years higher compared to the original cohort and more patients were matched for HLA-A, -B, -C, -DRB1, and DQB1 with their unrelated donors. Patients had predominantly received chemotherapy-based reduced-intensity conditioning instead of Total Body Irradiation (TBI)-based myeloablative conditioning, ATG for the prophylaxis of Graft versus Host Disease (GvHD) instead of ex vivo T-cell depletion, and had been transplanted with Peripheral Blood Stem Cells (PBSC) instead of bone marrow as graft source (12, 13). All factors reflect changes in clinical practice between the past cohorts and this contemporary cohort. Although little is known about the impact of different procedural choices on NK cell reconstitution after alloHCT, these factors could change NK cell mediated alloreactivity.
In order to comprehensively re-assess the work on outcome prediction after alloHCT based on the donor KIR genotype, we also evaluated alternative models. Genotypes can be categorized based on the absence and presence of certain activating and inhibiting KIR genes according to their content of centromeric and telomeric haplotype A and B motifs (33). Haplotype B motifs occur at lower frequencies in the Caucasian population and harbour more activating KIR genes (18). In larger studies, the presence and number of centromeric haplotype B motifs in the donor genome have been associated with a reduced risk of relapse after HLA-compatible unrelated allogeneic transplantation for AML (9–12). An updated and extended analysis was recently published by Weisdorf et al. (34). In a large contemporary cohort of AML patients the authors were able to demonstrate a beneficial impact of KIR haplotype B donors in the subgroup of C1/C1 or C1/C2 patients who had received reduced-intensity conditioning. In a strict sense, the attempt to validate their findings on the impact of haplotype B motifs on the risk of relapse after myeloablative conditioning therefore failed (11). Other groups have reported conflicting results which even indicated a protective effect of KIR haplotype A from smaller registry studies (35–37). In this study, we did not find a protective effect in univariable and multivariable comparisons for centromeric or telomeric KIR haplotype B versus A motifs in a contemporary cohort of patients with MDS of equal size (see Table 2 and Figure 3). Even, when we restricted the analyses to those subgroups, who had shown the greatest effects of KIR haplotype B donors, i.e. C1-positive patients who had received reduced-intensity conditioning, we did not observe significant effects in the predicted direction (Supplemental Table 2B).
Of note, in our exploratory analyses we observed opposite effects of donor KIR haplotype B motifs for patients with MDS versus secondary AML (see Supplemental Table 2A). For example, in MDS donor centromeric KIR haplotype B motifs appeared to be disadvantageous (HR, 1.48, p=0.03 for KIR cen B/B versus KIR cen A/A) whereas in secondary AML it appeared to be advantageous (HR, 0.73, p=0.3 for KIR cen B/B versus KIR cen A/A). Stringaris et al. reported that patients with MDS and KIR haplotype A show a more aggressive course of their disease (38). This would be in line with our results for patients with secondary AML but not with MDS. Nevertheless, we would like to caution against over-interpreting this signal. Reduced expression of HLA class I molecules which represent important KIR-ligands was not different in a recent study analysing samples from MDS and secondary AML for features which might explain escape from immune surveillance (39). Moreover, the group of patients with secondary AML was relatively small (N=473) in our study compared to the group of patients with MDS (N=1231) and it is possible that unknown confounders produced this difference. On the other hand, the different effect directions of KIR haplotypes in different types of diseases require careful consideration for future analyses.
Results from the exploratory subgroup analyses also revealed some differences for the estimated effects of KIR haplotype motifs in the context of reduced-intensity or non-myeloablative conditioning versus myeloablative conditioning, albeit not in a systematic way. Most important, the observed differences were not in line with what has been reported recently by Weisdorf et al. (34) and may be due to random effects.
The current assignment of the putative KIR haplotypes A and B as proposed by Uhrberg et al. in 1997 does not integrate allele-level information. With KIR genotyping at allele-level, KIR gene haplotypes based only on absence/presence information appear over-simplistic and may no longer represent a suitable grouping algorithm (40–42). While we found no impact of the classification into putative KIR haplotypes A and B, improved assignments based on KIR allele groups have the potential to reflect functional information of distinct KIR haplotypes more precisely.
A logical extension to the investigation of single KIR KIR-ligand combinations is creating additive models which integrate information on multiple KIR genes together with the corresponding ligands. One such additive model had been composed to predict NK-cell mediated control of chronic viral infections. The score is based on the number of functional inhibitory KIRs and predicts the progression of HIV, HCV, and HTLV-1 infections (24). The crude score of this model predicted the risk of relapse and EFS also in our data (see Table 2). In multivariable Cox regression analyses unrelated patient-donor pairs with an inhibitory KIR count of greater than 1 had a 24% lower risk of relapse (Wald-test, p=0.01) and a 20% lower risk (Wald-test, p=0.004) for events defining EFS compared to the patients with a single or no functional inhibitory KIR KIR-ligand combination. This association suggested that NK cells which were educated by multiple KIR–KIR–ligand interactions exerted stronger NK-mediated alloreactivity. Downregulation of HLA class I molecules on the malignant target cells would then be a necessary trigger for activation. Although down-regulation of class I molecules to escape T-cell attack is a common feature of cells infected by viruses and cancer cells, this mechanism was not found to be a major immune escape strategy after alloHCT. Recent data suggest that the primary immune escape mechanism after alloHCT starts by down-regulation of HLA-class II molecules (43, 44). The observed beneficial impact of a higher inhibitory KIR count is therefore not supported by the currently-favoured concept of immune escape of malignant cells after allogeneic transplantation (45). On the other hand, due to a lack of a humanized animal model which recapitulates graft versus leukemia effects, the basic biological principles of NK cell mediated allo-reactivity remain uncertain. Thus, the jury is still out, on whether donors with more or less functionally inhibitory KIRs may exert stronger graft-versus leukemia reactions after HLA-compatible unrelated alloHCT. The testing of integrated scores which reflect a conclusive biological concept in large registry studies may further inform our understanding of NK biology and more research in this regard is warranted. However, since the functionally inhibitory KIR count was tested here as part of a series of exploratory analyses without keeping stringent control of the family-wise type I error rate, we do not recommend application of the functionally inhibitory KIR count for donor selection in the context of stem cell transplantation based on the current data.
From a conceptual point of view the integration of information on donor KIRs and patient KIR-ligands in one unifying score is appealing. Very likely, an optimal score for the prediction of relapse after matched unrelated donor alloHCT will be complex and may contain first- and second-order interactions. Weights for single factors must be defined carefully. Mathematically step-functions may be more appropriate to predict threshold-dependent NK cell activation or inhibition than linear functions. However, given the stochastic expression of KIRs on NK cells composing the individual repertoire, the breadth of a potential NK response may also be impacted. This would weigh an argument in favour of linear or monotonous relations. To address these challenges in model-building, machine learning algorithms may become necessary tools. Given recent successes in the use of artificial intelligence in medical science and the complexity of KIR genetics and NK biology in the transplant context, this area of research appears to be especially appealing for the application of these new techniques (46).
Critical resources required to answer the question on whether the donor KIR genotype can be used to predict patient outcome after HLA-compatible unrelated alloHCT, are i) access to donor samples donated for research at biobanks, ii) collaborative efforts to pull together large datasets and sample sizes necessary to apply machine learning tools or other complex statistical models, iii) access to affordable allele-level KIR typing, iv) stringent statistical testing strategies to keep control of the family-wise type I error and to validate findings in independent datasets, and finally v) active research groups committed to advancing the understanding of NK biology in the context of alloHCT (47, 48). The European Society for Blood and Marrow Transplantation (EBMT), the Center for International Blood and Marrow Transplant Research (CIBMTR), the National Marrow Donor Program (NMDP) and DKMS are committed to answering this question and can provide critical resources. A large collaborative effort of these institutions to come to a conclusive answer is currently underway.
In summary, despite availability of KIR genotype information for more than 3 million potential stem cell donors, no KIR-based algorithm for unrelated donor selection has entered clinical practice. After more than 20 years of research, the impact of donor KIR genotype information on the outcome after unrelated donor alloHCT is still not clear. This large study of patients with MDS and secondary AML adds to the growing body of data by showing that the KIR KIR-ligand combinations, KIR2DS1-C2 and KIR3DL1-Bw4(80I/T) and putative haplotype motif based models have no impact on the risk of relapse and mortality after unrelated donor alloHCT. However, with larger studies and intergroup collaborations on the horizon, high-throughput allelic resolution of KIR genes at hand and increasing experience in dealing with high-dimensional data, the chances are good that the question on whether KIR genotype information can be used for donor selection will be resolved in the next couple of years.
Author's Note
This was a joint study of the Chronic Malignancies Working Party of the European Society for Blood and Marrow Transplantation (EBMT) and the Center for International Blood and Marrow Transplant Research (CIBMTR).
Data Availability Statement
The datasets presented in this article are not readily available because the medical data belongs to the EBMT and the CIBMTR. Generated datasets can be requested by the corresponding author conditional on the approval of EBMT and CIBMTR. Requests to access the datasets should be directed to johannes.schetelig@ukdd.de.
Ethics Statement
The studies involving human participants were reviewed and approved by Ethical Committee of the Technische Universität Dresden. Access to medical data was approved by the Review Boards of the Chronic Malignancies Working Party of EBMT and the Immunobiology Working Committee of the CIBMTR and the National Marrow Donor Program Institutional Review Board. All patients and donors gave written informed consent to the use of samples and medical data for medical research.
Author Contributions
JS, HB, FH, AS, and MR designed the study. JS, MK SS, MG, BB, FO, VP, PL, NS, PH, SL, NK, KH, IY-A, and MR contributed medical data. HB, LK, MK, LW, and CM contributed to the different levels of the processing of genetic and medical data. JS, MK, HB, and LW performed the statistical analysis. JS and HB wrote the manuscript. All authors interpreted and discussed the results, and reviewed and approved the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This study was enabled by DKMS gGmbH who financed the donor KIR typing and supported this study with know-how and logistics. No funding from commercial entities was received for this study. The CIBMTR is supported primarily by Public Health Service U24CA076518 from the National Cancer Institute (NCI), the National Heart, Lung and Blood Institute (NHLBI), and the National Institute of Allergy and Infectious Diseases (NIAID); U24HL138660 from NHLBI and NCI; R21HL140314, and U01HL128568 from the NHLBI; HHSH250201700006C, SC1MC31881-01-00, and HHSH250201700007C from the Health Resources and Services Administration (HRSA); and N00014-18-1-2850, N00014-18-1-2888, and N00014-20-1-2705 from the Office of Naval Research. Additional federal support is provided by P01CA111412, R01CA152108, R01CA215134, R01CA218285, R01CA231141, R01HL126589, R01AI128775, R01HL129472, R01HL130388, R01HL131731, U01AI069197, U01AI126612, and BARDA. Support is also provided by Be the Match Foundation, Boston Children’s Hospital, Dana Farber, Japan Hematopoietic Cell Transplantation Data Center, St. Baldrick’s Foundation, the National Marrow Donor Program, the Medical College of Wisconsin and from the following commercial entities: AbbVie; Actinium Pharmaceuticals, Inc.; Adaptive Biotechnologies; Adienne SA; Allovir, Inc.; Amgen, Inc.; Anthem, Inc.; Astellas Pharma US; AstraZeneca; Atara Biotherapeutics, Inc.; bluebird bio, Inc.; Bristol Myers Squibb Co.; Celgene Corp.; Chimerix, Inc.; CSL Behring; CytoSen Therapeutics, Inc.; Daiichi Sankyo Co., Ltd.; Gamida-Cell, Ltd.; Genzyme; GlaxoSmithKline (GSK); HistoGenetics, Inc.; Incyte Corporation; Janssen Biotech, Inc.; Janssen Pharmaceuticals, Inc.; Janssen/Johnson & Johnson; Jazz Pharmaceuticals, Inc.; Kiadis Pharma; Kite Pharma; Kyowa Kirin; Legend Biotech; Magenta Therapeutics; Mallinckrodt LLC; Medac GmbH; Merck & Company, Inc.; Merck Sharp & Dohme Corp.; Mesoblast; Millennium, the Takeda Oncology Co.; Miltenyi Biotec, Inc.; Novartis Oncology; Novartis Pharmaceuticals Corporation; Omeros Corporation; Oncoimmune, Inc.; Orca Biosystems, Inc.; Pfizer, Inc.; Phamacyclics, LLC; Regeneron Pharmaceuticals, Inc.; REGiMMUNE Corp.; Sanofi Genzyme; Seattle Genetics; Sobi, Inc.; Takeda Oncology; Takeda Pharma; Terumo BCT; Viracor Eurofins; and Xenikos BV. The views expressed in this article do not reflect the official policy or position of the National Institute of Health, the Department of Navy, the Department of Defense, Health Resources and Services Administration (HRSA), or any other agency of the U.S. Government.
Conflict of Interest
The DKMS Life Science Laboratory (VL, CM, AS) implemented the KIR genotyping as part of the upfront genotyping profile for volunteers enrolled into the DKMS and offers KIR genotyping also for external customers.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful to the team of the Collaborative Biobank, especially Stephanie Maiwald, who coordinated the sample export and pre-analytic processing, the DKMS Cord Blood bank, and the Scientific Team at DKMS Tübingen, who facilitated this analysis. Further, we would like to acknowledge the excellent contribution of Bose Falk, Jürgen Sauter, and Ute Solloch, who significantly contributed to the complex data management processes.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2020.584520/full#supplementary-material
References
1. Ruggeri L, Capanni M, Urbani E, Perruccio K, Shlomchik WD, Tosti A, et al. Effectiveness of donor natural killer cell alloreactivity in mismatched hematopoietic transplants. Science (2002) 295:2097–100. doi: 10.1126/science.1068440
2. Miller JS, Soignier Y, Panoskaltsis-Mortari A, McNearney SA, Yun GH, Fautsch SK, et al. Successful adoptive transfer and in vivo expansion of human haploidentical NK cells in patients with cancer. Blood (2005) 105:3051–7. doi: 10.1182/blood-2004-07-2974
3. Shaffer BC, Le Luduec JB, Forlenza C, Jakubowski AA, Perales MA, Young JW, et al. Phase II Study of Haploidentical Natural Killer Cell Infusion for Treatment of Relapsed or Persistent Myeloid Malignancies Following Allogeneic Hematopoietic Cell Transplantation. Biol Blood Marrow Transpl (2016) 22:705–9. doi: 10.1016/j.bbmt.2015.12.028
4. Romee R, Rosario M, Berrien-Elliott MM, Wagner JA, Jewell BA, Schappe T, et al. Cytokine-induced memory-like natural killer cells exhibit enhanced responses against myeloid leukemia. Sci Transl Med (2016) 8:357ra123. doi: 10.1126/scitranslmed.aaf2341
5. Bjorklund AT, Carlsten M, Sohlberg E, Liu LL, Clancy T, Karimi M, et al. Complete Remission with Reduction of High-Risk Clones following Haploidentical NK-Cell Therapy against MDS and AML. Clin Cancer Res (2018) 24:1834–44. doi: 10.1158/1078-0432.CCR-17-3196
6. Giebel S, Locatelli F, Lamparelli T, Velardi A, Davies S, Frumento G, et al. Survival advantage with KIR ligand incompatibility in hematopoietic stem cell transplantation from unrelated donors. Blood (2003) 102:814–9. doi: 10.1182/blood-2003-01-0091
7. Hsu KC, Keever-Taylor CA, Wilton A, Pinto C, Heller G, Arkun K, et al. Improved outcome in HLA-identical sibling hematopoietic stem-cell transplantation for acute myelogenous leukemia predicted by KIR and HLA genotypes. Blood (2005) 105:4878–84. doi: 10.1182/blood-2004-12-4825
8. Miller JS, Cooley S, Parham P, Farag SS, Verneris MR, McQueen KL, et al. Missing KIR ligands are associated with less relapse and increased graft-versus-host disease (GVHD) following unrelated donor allogeneic HCT. Blood (2007) 109:5058–61. doi: 10.1182/blood-2007-01-065383
9. Cooley S, Trachtenberg E, Bergemann TL, Saeteurn K, Klein J, Le CT, et al. Donors with group B KIR haplotypes improve relapse-free survival after unrelated hematopoietic cell transplantation for acute myelogenous leukemia. Blood (2009) 113:726–32. doi: 10.1182/blood-2008-07-171926
10. Cooley S, Weisdorf DJ, Guethlein LA, Klein JP, Wang T, Le CT, et al. Donor selection for natural killer cell receptor genes leads to superior survival after unrelated transplantation for acute myelogenous leukemia. Blood (2010) 116:2411–9. doi: 10.1182/blood-2010-05-283051
11. Cooley S, Weisdorf DJ, Guethlein LA, Klein JP, Wang T, Marsh SG, et al. Donor killer cell Ig-like receptor B haplotypes, recipient HLA-C1, and HLA-C mismatch enhance the clinical benefit of unrelated transplantation for acute myelogenous leukemia. J Immunol (2014) 192:4592–600. doi: 10.4049/jimmunol.1302517
12. Venstrom JM, Pittari G, Gooley TA, Chewning JH, Spellman S, Haagenson M, et al. HLA-C-dependent prevention of leukemia relapse by donor activating KIR2DS1. N Engl J Med (2012) 367:805–16. doi: 10.1056/NEJMoa1200503
13. Boudreau JE, Giglio F, Gooley TA, Stevenson PA, Le Luduec JB, Shaffer BC, et al. KIR3DL1/ HL A-B Subtypes Govern Acute Myelogenous Leukemia Relapse After Hematopoietic Cell Transplantation. J Clin Oncol (2017) 35:2268–78. doi: 10.1200/JCO.2016.70.7059
14. Wilson MJ, Torkar M, Haude A, Milne S, Jones T, Sheer D, et al. Plasticity in the organization and sequences of human KIR/ILT gene families. Proc Natl Acad Sci U S A (2000) 97:4778–83. doi: 10.1073/pnas.080588597
15. Norman PJ, Hollenbach JA, Nemat-Gorgani N, Guethlein LA, Hilton HG, Pando MJ, et al. Co-evolution of human leukocyte antigen (HLA) class I ligands with killer-cell immunoglobulin-like receptors (KIR) in a genetically diverse population of sub-Saharan Africans. PLoS Genet (2013) 9:e1003938. doi: 10.1371/journal.pgen.1003938
16. Wagner I, Schefzyk D, Pruschke J, Schofl G, Schone B, Gruber N, et al. Allele-Level KIR Genotyping of More Than a Million Samples: Workflow, Algorithm, and Observations. Front Immunol (2018) 9:2843. doi: 10.3389/fimmu.2018.02843
17. Jiang W, Johnson C, Jayaraman J, Simecek N, Noble J, Moffatt MF, et al. Copy number variation leads to considerable diversity for B but not A haplotypes of the human KIR genes encoding NK cell receptors. Genome Res (2012) 22:1845–54. doi: 10.1101/gr.137976.112
18. Norman PJ, Hollenbach JA, Nemat-Gorgani N, Marin WM, Norberg SJ, Ashouri E, et al. Defining KIR and HLA Class I Genotypes at Highest Resolution via High-Throughput Sequencing. Am J Hum Genet (2016) 99:375–91. doi: 10.1016/j.ajhg.2016.06.023
19. Beziat V, Hilton HG, Norman PJ, Traherne JA. Deciphering the killer-cell immunoglobulin-like receptor system at super-resolution for natural killer and T-cell biology. Immunology (2017) 150:248–64. doi: 10.1111/imm.12684
20. Gardiner CM, Guethlein LA, Shilling HG, Pando M, Carr WH, Rajalingam R, et al. Different NK cell surface phenotypes defined by the DX9 antibody are due to KIR3DL1 gene polymorphism. J Immunol (2001) 166:2992–3001. doi: 10.4049/jimmunol.166.5.2992
21. Martin MP, Qi Y, Gao X, Yamada E, Martin JN, Pereyra F, et al. Innate partnership of HLA-B and KIR3DL1 subtypes against HIV-1. Nat Genet (2007) 39:733–40. doi: 10.1038/ng2035
22. Maccari G, Robinson J, Hammond JA, Marsh SGE. The IPD Project: a centralised resource for the study of polymorphism in genes of the immune system. Immunogenetics (2020) 72:49–55. doi: 10.1007/s00251-019-01133-w
23. Schetelig J, Baldauf H, Heidenreich F, Massalski C, Frank S, Sauter J, et al. External validation of models for KIR2DS1/KIR3DL1-informed selection of hematopoietic cell donors fails. Blood (2020) 135:1386–95. doi: 10.1182/blood.2019002887
24. Boelen L, Debebe B, Silveira M, Salam A, Makinde J, Roberts CH, et al. Inhibitory killer cell immunoglobulin-like receptors strengthen CD8(+) T cell-mediated control of HIV-1, HCV, and HTLV-1. Sci Immunol (2018) 3. doi: 10.1126/sciimmunol.aao2892
25. Krieger E, Sabo R, Moezzi S, Cain C, Roberts C, Kimball P, et al. Killer Immunoglobulin-Like Receptor-Ligand Interactions Predict Clinical Outcomes following Unrelated Donor Transplantations. Biol Blood Marrow Transpl (2020) 26:672–82. doi: 10.1016/j.bbmt.2019.10.016
26. Arber DA, Orazi A, Hasserjian R, Thiele J, Borowitz MJ, Le Beau MM, et al. The 2016 revision to the World Health Organization classification of myeloid neoplasms and acute leukemia. Blood (2016) 127:2391–405. doi: 10.1182/blood-2016-03-643544
27. Armand P, Kim HT, Logan BR, Wang Z, Alyea EP, Kalaycio ME, et al. Validation and refinement of the Disease Risk Index for allogeneic stem cell transplantation. Blood (2014) 123:3664–71. doi: 10.1182/blood-2014-01-552984
28. Bacigalupo A, Ballen K, Rizzo D, Giralt S, Lazarus H, Ho V, et al. Defining the intensity of conditioning regimens: working definitions. Biol Blood Marrow Transpl (2009) 15:1628–33. doi: 10.1016/j.bbmt.2009.07.004
29. Schoenfeld DA. Sample-size formula for the proportional-hazards regression model. Biometrics (1983) 39:499–503. doi: 10.2307/2531021
30. Grambsch P, Therneau T. Proportional hazards tests and diagnostics based on weighted residuals. Biometrika (1994) 81:515–26. doi: 10.1093/biomet/81.3.515
31. Gagne K, Busson M, Bignon JD, Balere-Appert ML, Loiseau P, Dormoy A, et al. Donor KIR3DL1/3DS1 gene and recipient Bw4 KIR ligand as prognostic markers for outcome in unrelated hematopoietic stem cell transplantation. Biol Blood Marrow Transpl (2009) 15:1366–75. doi: 10.1016/j.bbmt.2009.06.015
32. Sobecks RM, Wang T, Askar M, Gallagher MM, Haagenson M, Spellman S, et al. Impact of KIR and HLA Genotypes on Outcomes after Reduced-Intensity Conditioning Hematopoietic Cell Transplantation. Biol Blood Marrow Transpl (2015) 21:1589–96. doi: 10.1016/j.bbmt.2015.05.002
33. Uhrberg M, Valiante NM, Shum BP, Shilling HG, Lienert-Weidenbach K, Corliss B, et al. Human diversity in killer cell inhibitory receptor genes. Immunity (1997) 7:753–63. doi: 10.1016/s1074-7613(00)80394-5
34. Weisdorf D, Cooley S, Wang T, Trachtenberg E, Vierra-Green C, Spellman S, et al. KIR B donors improve the outcome for AML patients given reduced intensity conditioning and unrelated donor transplantation. Blood Adv (2020) 4:740–54. doi: 10.1182/bloodadvances.2019001053
35. Cook MA, Milligan DW, Fegan CD, Darbyshire PJ, Mahendra P, Craddock CF, et al. The impact of donor KIR and patient HLA-C genotypes on outcome following HLA-identical sibling hematopoietic stem cell transplantation for myeloid leukemia. Blood (2004) 103:1521–6. doi: 10.1182/blood-2003-02-0438
36. Kroger N, Binder T, Zabelina T, Wolschke C, Schieder H, Renges H, et al. Low number of donor activating killer immunoglobulin-like receptors (KIR) genes but not KIR-ligand mismatch prevents relapse and improves disease-free survival in leukemia patients after in vivo T-cell depleted unrelated stem cell transplantation. Transplantation (2006) 82:1024–30. doi: 10.1097/01.tp.0000235859.24513.43
37. McQueen KL, Dorighi KM, Guethlein LA, Wong R, Sanjanwala B, Parham P. Donor-recipient combinations of group A and B KIR haplotypes and HLA class I ligand affect the outcome of HLA-matched, sibling donor hematopoietic cell transplantation. Hum Immunol (2007) 68:309–23. doi: 10.1016/j.humimm.2007.01.019
38. Stringaris K, Marin D, Barrett AJ, Hills R, Sobieski C, Cao K, et al. KIR gene haplotype: an independent predictor of clinical outcome in MDS patients. Blood (2016) 128:2819–23. doi: 10.1182/blood-2016-05-713099
39. Montes P, Bernal M, Campo LN, Gonzalez-Ramirez AR, Jimenez P, Garrido P, et al. Tumor genetic alterations and features of the immune microenvironment drive myelodysplastic syndrome escape and progression. Cancer Immunol Immunother (2019) 68:2015–27. doi: 10.1007/s00262-019-02420-x
40. Vierra-Green C, Roe D, Jayaraman J, Trowsdale J, Traherne J, Kuang R, et al. Estimating KIR Haplotype Frequencies on a Cohort of 10,000 Individuals: A Comprehensive Study on Population Variations, Typing Resolutions, and Reference Haplotypes. PLoS One (2016) 11:e0163973. doi: 10.1371/journal.pone.0163973
41. Roe D, Vierra-Green C, Pyo CW, Eng K, Hall R, Kuang R, et al. Revealing complete complex KIR haplotypes phased by long-read sequencing technology. Genes Immun (2017) 18:127–34. doi: 10.1038/gene.2017.10
42. Solloch UV, Schefzyk D, Schafer G, Massalski C, Kohler M, Pruschke J, et al. Estimation of German KIR Allele Group Haplotype Frequencies. Front Immunol (2020) 11:429. doi: 10.3389/fimmu.2020.00429
43. Toffalori C, Zito L, Gambacorta V, Riba M, Oliveira G, Bucci G, et al. Immune signature drives leukemia escape and relapse after hematopoietic cell transplantation. Nat Med (2019) 25:603–11. doi: 10.1038/s41591-019-0400-z
44. Christopher MJ, Petti AA, Rettig MP, Miller CA, Chendamarai E, Duncavage EJ, et al. Immune Escape of Relapsed AML Cells after Allogeneic Transplantation. N Engl J Med (2018) 379:2330–41. doi: 10.1056/NEJMoa1808777
45. Boudreau JE, Hsu KC. Natural Killer Cell Education and the Response to Infection and Cancer Therapy: Stay Tuned. Trends Immunol (2018) 39:222–39. doi: 10.1016/j.it.2017.12.001
46. Ho D. Artificial intelligence in cancer therapy. Science (2020) 367:982–3. doi: 10.1126/science.aaz3023
47. Schmidt AH, Sauter J, Baier DM, Daiss J, Keller A, Klussmeier A, et al. Immunogenetics in stem cell donor registry work: The DKMS example (Part 1). Int J Immunogenet (2020) 47:13–23. doi: 10.1111/iji.12471
Keywords: KIR, KIR2DS1, KIR3DL1, hematopoietic stem cell transplantation, donor selection, unrelated donor
Citation: Schetelig J, Baldauf H, Koster L, Kuxhausen M, Heidenreich F, de Wreede LC, Spellman S, van Gelder M, Bruno B, Onida F, Lange V, Massalski C, Potter V, Ljungman P, Schaap N, Hayden P, Lee SJ, Kröger N, Hsu K, Schmidt AH, Yakoub-Agha I and Robin M (2021) Haplotype Motif-Based Models for KIR-Genotype Informed Selection of Hematopoietic Cell Donors Fail to Predict Outcome of Patients With Myelodysplastic Syndromes or Secondary Acute Myeloid Leukemia. Front. Immunol. 11:584520. doi: 10.3389/fimmu.2020.584520
Received: 01 September 2020; Accepted: 25 November 2020;
Published: 19 January 2021.
Edited by:
Aharon Freud, The Ohio State University, United StatesReviewed by:
Michael R. Verneris, University of Colorado Denver, United StatesRafael Solana, University of Cordoba, Spain
Copyright © 2021 Schetelig, Baldauf, Koster, Kuxhausen, Heidenreich, de Wreede, Spellman, van Gelder, Bruno, Onida, Lange, Massalski, Potter, Ljungman, Schaap, Hayden, Lee, Kröger, Hsu, Schmidt, Yakoub-Agha and Robin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Johannes Schetelig, johannes.schetelig@ukdd.de