Synbiotics Supplements Lower the Risk of Hand, Foot, and Mouth Disease in Children, Potentially by Providing Resistance to Gut Microbiota Dysbiosis
 Xiaoying Guo1,2†
Xiaoying Guo1,2†  Zixin Lan3† Yaling Wen4 Chanjiao Zheng5 Zuhua Rong1
Zixin Lan3† Yaling Wen4 Chanjiao Zheng5 Zuhua Rong1  Tao Liu1 Siyi Chen1
Tao Liu1 Siyi Chen1  Xingfen Yang2* Huimin Zheng1*
Xingfen Yang2* Huimin Zheng1*  Wei Wu1,2*
Wei Wu1,2*- 1Guangdong Provincial Institute of Public Health, Guangdong Provincial Center for Disease Control and Prevention, Guangzhou, China
- 2School of Public Health, Southern Medical University, Guangzhou, China
- 3The Second Clinical Medical College, Southern Medical University, Guangzhou, China
- 4School of Mathematics and Computational Science, Guilin University of Electronic Technology, Guangxi, China
- 5Modern Service Industry Department, Guangzhou Technician College, Guangzhou, China
Background: Hand, foot and mouth disease (HFMD) is an acute enterovirus-induced disease. Gut microbiota dysbiosis has been identified as a factor that plays an important role in enteral virus infection, but the gut microbiota profile in hand, foot and mouth disease has rarely been studied in a large population.
Methods: A total of 749 children (HFMD: n = 262, healthy control: n = 487) aged 2 to 7 years were recruited from hospitals and communities in the period from May to July, 2017. Clinical and demographical information was collected by trained personnel, and fecal samples were collected and processed for 16S ribosomal RNA(rRNA) gene sequencing.
Results: We observed a significant alteration in the microbiota profile of children with HFMD compared with that of control children. Patients with enteroviruses A71(EV71) positive had more dysbiotic gut microbiota than those with coxsackievirus A16 (CAV16) positive. We found that Prevotella and Streptococcus were enriched in children with HFMD, whereas beneficial bacteria, including Bifidobacterium and Faecalibacterium, were depleted. Children with synbiotics supplements had lower risk of HFMD and we observed that the gut microbiota of HFMD patients who were administered synbiotics exhibited potential resistance to the dysbiosis detected in HFMD.
Conclusions: This study suggested that the gut microbiota of patients with hand, foot and mouth disease exhibits dysbiosis and that synbiotics supplements potentially helps maintain the homeostasis of the gut flora.
Introduction
Hand, foot, and mouth disease (HFMD), a gastrointestinal contagious disease that mostly affects individuals under the age of 10 years, is one of the top three class C notifiable infectious diseases in China (Ying et al., 2019). Hence, HFMD poses a great threat to pediatric and neonatal populations in Asian countries.
The HFMD pathogens, mainly enteroviruses A71(EV71) and coxsackievirus A16 (CVA16), target the gastrointestinal (GI) epithelium and utilize intestinal bacteria to enhance multiplication, pathogenesis, and transmission and thereby lead to development of the disease. Additionally, the gut microbiota has been reported to work in conjunction with the intestinal barrier to orchestrate a defense network that impedes the invasion of pathogens and maintains the homeostasis and functionality of the gut (Sanz and De, 2009; Odenwald and Turner, 2017; Takiishi et al., 2017; Robinson, 2019). Dysbiosis of the gut microbiota could also lead to intestinal barrier disruption by influencing the self-renewal of epithelial cell and the secretion of the mucus layer and tight junctions of intestinal epithelial cells, which could increase the susceptibility of the host to immune or inflammatory disorders, including enterovirus infection (Wu et al., 2013; Winglee et al., 2014; Jakobsson et al., 2015; Schuijt et al., 2015; Sun et al., 2016; Heidrich et al., 2018; Alam and Neish, 2018). While majority studies had primarily focused on patients in epidemiological and virological studies, the role of the fecal bacterial microbiome in HFMD progression remains barely undocumented. Thus far, only limited studies with very small sample sizes have suggested the existence of gut microbiota dysbiosis in children with HFMD, and larger population studies are urgently needed to reveal the relationship and the underlying mechanisms. Probiotics and prebiotics are widely utilized as complements to nutrition strategies designed to reinforce intestinal immunity. Several studies have provided scientific evidence of the benefits resulting from the use of probiotics and prebiotics for intestinal diseases. Currently, researchers are considering the use of probiotics and prebiotics for the prevention of HFMD, but no related studies have been conducted to date. It is necessary to assess whether the use of probiotics and prebiotics exhibits potential preventive efficacy on intestinal flora in children with HFMD.
Here, we conducted a two-site recruitment and investigated 749 participants (including 262 HFMD patients and 487 healthy children) from two regions in Guangdong, China. By collecting their information and fecal samples, we aimed to explore the differences in gut microbiota communities between children with HFMD and healthy children and the potential effects of probiotics and prebiotics on HFMD infection.
Materials and Methods
Study Design and Subjects
All study participants were recruited from an investigation that aimed to investigate the distribution of intestinal bacteria and its influencing factors in children with HFMD that was conducted in Guangdong Province, from May to July, 2017. The investigation sites were Xinhui District of Jiangmen City and Longchuan County of Heyuan City, Guangdong Province. Two local hospitals, including a children’s hospital and a county-level hospital, were selected in each investigation site. HFMD patients were recruited from local hospitals, and healthy children were recruited from the same hospitals or nearby communities where the patients live. The inclusion criteria were as follows: a) 2- to 7-year-old children; b) living in the county or district for at least 6 months. The exclusion criteria: a) having a critical illness, herpetic pharyngitis and severe organic lesions; b) having a disease related to the risk factors for the study, including diarrhea, gastroenteritis, previous diagnosis of HFMD, and herpetic angina, among others; and c) taking antibiotics within one month. The HFMD cases were diagnosed by medical institutions and clinics in the county or district according to the Chinese Guidelines for HFMD diagnosis and treatment (2010 Edition) issued by the Ministry of Health1 Ministry of Health of the People’s Republic of China.(2010). Diagnosis and treatment guideline on hand-foot-mouth disease. http://yzs.satcm.gov.cn/gongzuodongtai/2018-03-24/3073.html. [Accessed March 1, 2017].
Samples and Sequencing
Fecal samples were collected from all 749 subjects who completed the questionnaire, and within 2 h, these samples were placed in a refrigerator at 4°C for temporary preservation. Within 24 h, the samples were stored at -80°C before analysis.
According to the instructions provided with the QIAamp DNA Stool Mini kit (QIAGEN, Hilden, Germany), total DNA were extracted from fecal samples. The V4 region of 16S rRNA gene was amplified by polymerase chain reaction (PCR) from the total isolated DNA samples. The amplification reaction conditions were as follows: initial activation step of 94°C for 2 min followed by 30 cycles of 94°C for 30 s, 52°C for 30 s and 72°C for 30 s and a final incubation at 72°C for 5 min. The V4 region of 16S rRNA was sequenced by second-generation sequencing (Illumina HiSeq 2000) in combination with third-generation sequencing (PacBio SMRT).
Bioinformatic Analysis
Using R V.4.0.3 (under RStudio V. 1.4.1106), the 16S rRNA gene reads were merged, subjected to quality control and clustered into amplicon sequence variants (ASVs). The R package DADA2 was used to process the Illumina demultiplexed paired-end sequenced dataset to correct for amplicon errors, to identify chimeras and to merge paired-end reads. Low-quality regions at the tails in the sequences were first trimmed for removal. Based on the estimated error rate, filtered forward and reverse sequences were then dereplicated. Unique sequences were identified using pseudo-pooling, and paired reads were merged to generate fully denoised sequences. The denoised sequences were then assigned to ASVs. Taxonomically, the samples were classified according to the Greengenes reference database (McDonald et al., 2012).
The values for alpha diversity (Chao1 index, Shannon’s index, PD whole tree index and observed ASVs), beta diversity (Bray Curtis, weighted and unweighted UniFrac distance metrics) and principal coordinate analysis (PCoA) were generated with R. Permutational multivariate analysis of variance was performed to determine whether the compositions of the microbiota differed between groups. To identify the enriched and depleted bacteria between groups, differential taxa in the cases and the controls were discovered by LEfSe using the web-based program Galaxy (Segata et al., 2011).
Statistical Analysis
The categorical variables are expressed as means and standard deviation and the numeric variables are reported as percentages. The characteristics of the individuals in the cohort were compared by Student’s t-test and Chi-squared test. The Wilcoxon rank-sum test was used to compare the Chao1, Shannon index, observed ASVs, and PD whole-tree index between the cases and controls. The relative abundances of genera were compared among the groups using the Kruskal-Wallis rank sum test. Benjamini-Hochberg method was used to adjust for multiple comparation. We used logistic regression models to calculate the odds ratios between the HFMD and control groups, and the Wald’s test and likelihood ratio test were then used to analyze the relationship of the groups. Propensity Score Matching (PSM) was applied to minimize the effect of potentially confounding factors, such as matching two cohorts to have the same distribution of age and sex. Then, a propensity score for the relative abundances of selected genera of each individual was calculated after using results of this model. Finally, matching with a 1:1 ratio was performed using a nearest-neighbor approach with caliper restriction between case group and controls. In addition, the association between genus and HFMD was assessed by multivariate association with linear models 2 (MaAsLin2) as described by Morgan et al. (Himel, 2021). Age and sex were used as confounders, and the false discovery rate was limited to 0.05. All analyses were performed using R software, version 4.0.2 (R Foundation for Statistical Computing, Vienna, Austria). Statistical significance was accepted at P < 0.05.
Results
Study Population
In the present study, a total of 749 children (487 healthy childrenand 262 children with HFMD) from Heyuan and Jiangmen were included. The mean age of the patients with HFMD was 3.6 years, whereas the average age of the control group was 4.6 years. Males made up 53 percent of the subjects in the control group, whereas 62 percent of the group of children with HFMD were males. The age, sex, probiotics, prebiotics and synbiotics supplements, hygienic habits, city information and statistics and of the study participants are detailed in Table 1.
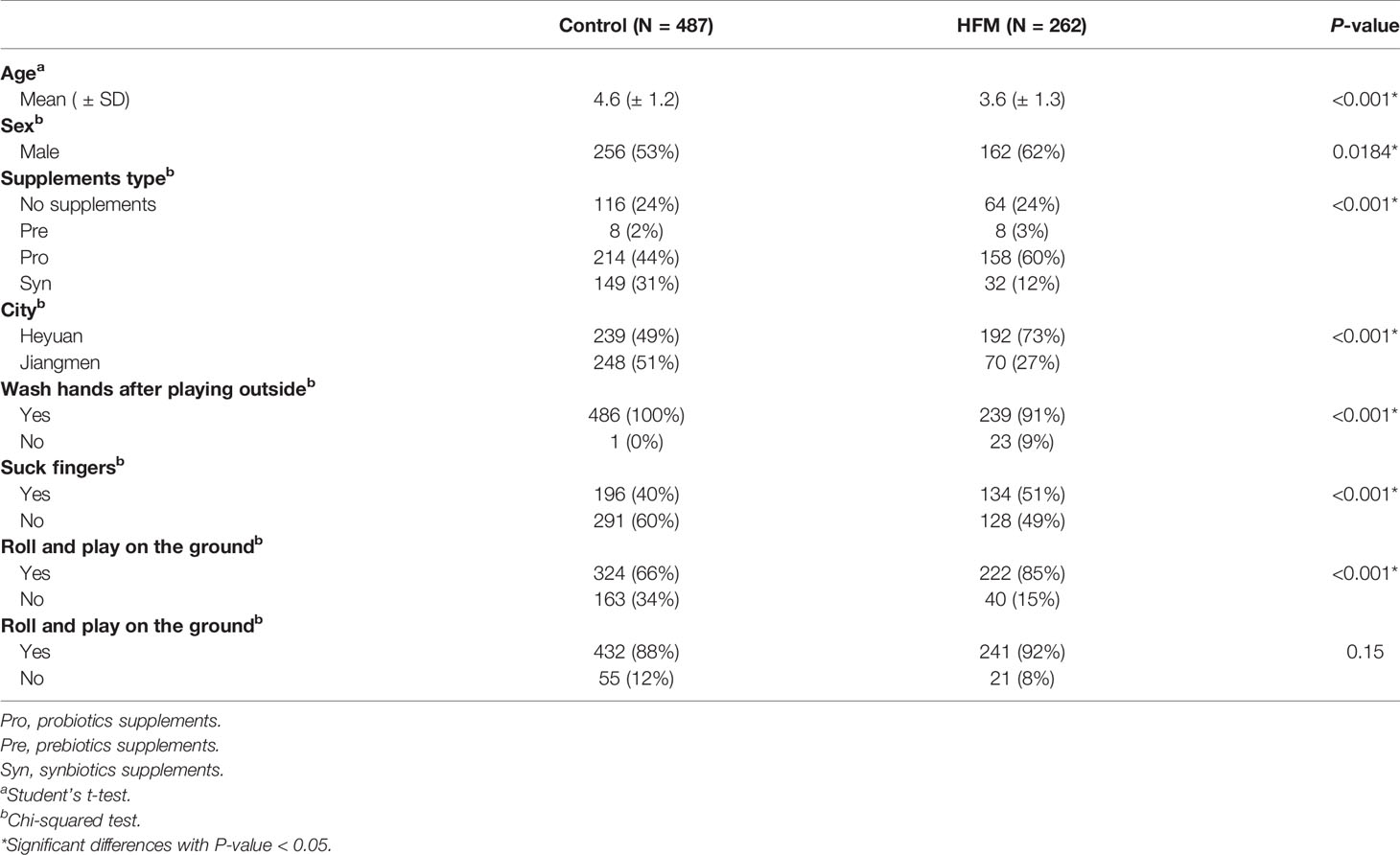
Table 1 Demographic characteristics and probiotics, prebiotics, and synbiotics supplement of the participants.
Altered Gut Microbiota Profile of Children With HFMD
The gut microbiota analysis revealed significant differences in the alpha and beta diversity. The HFMD group exhibited significantly lower alpha diversity indices, including the Shannon index, observed ASVs, PD whole-tree index, and Chao1 index, than the control group. This finding suggested that the gut microbiome of the HFMD group exhibited lower richness and reduced evenness than that of the control group (Figures 1A–D). According to the PCoA plot based on the unweighted and weighted UniFrac distances, the gut microbiota of the HFMD group differed significantly from that of the control group, which indicated that the composition of the gut microbiota of the HFMD group was significantly different from that of the control group (Figures 1E, F).
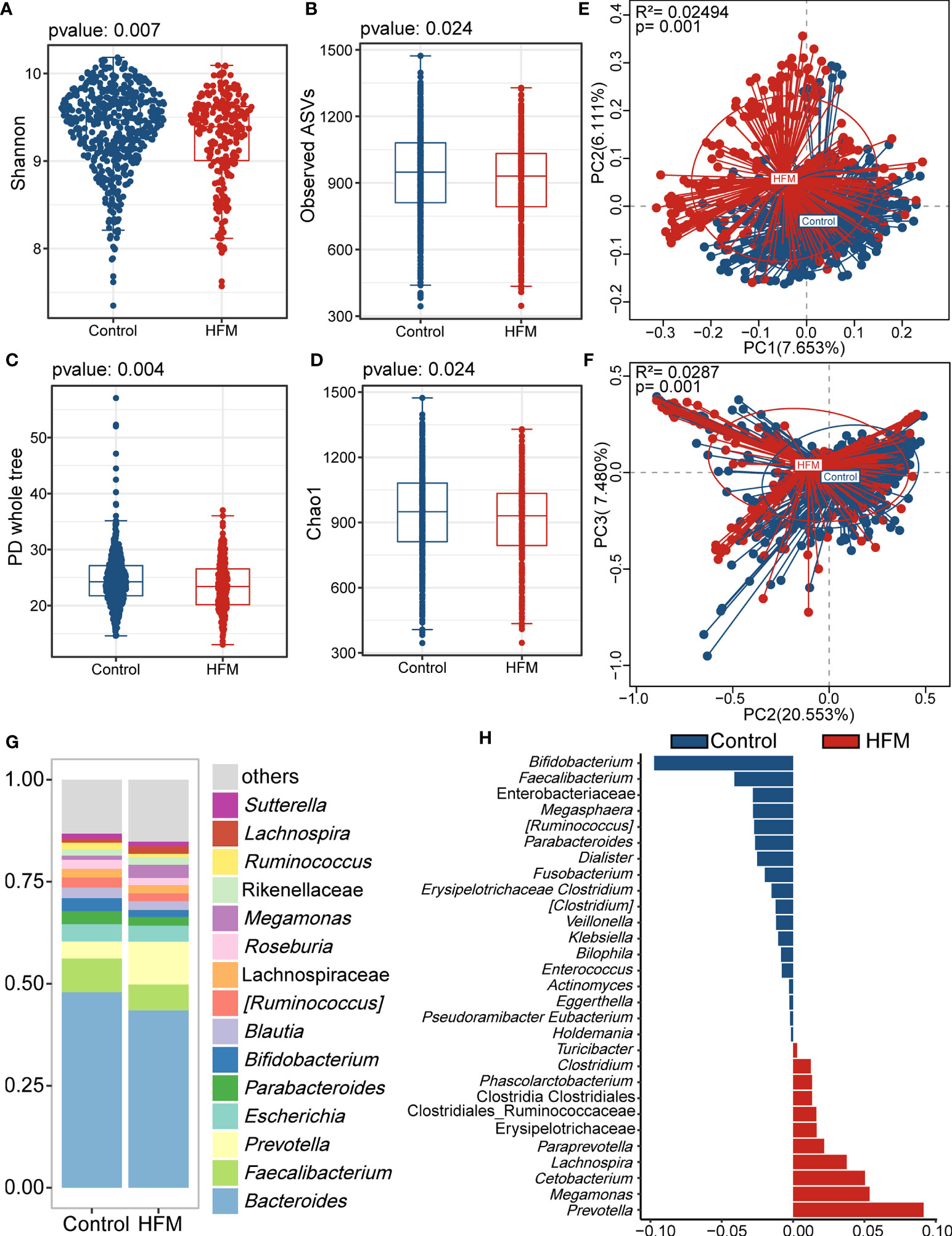
Figure 1 Altered bacterial microbiota biodiversity and composition in HFMD based on whole cohort. (A–D) Alpha diversity. Comparison of alpha-diversity indices (Shannon, observed ASVs, PD whole tree and Chao 1 index) between the male HFMD and male control groups (Wilcoxon rank-sum tests). (E, F) Principal coordinate analysis based on unweighted and unweighted UniFrac distances revealed that the bacterial communities of patients with HFMD clustered separately from the bacterial communities of healthy control children. Each point represents a single sample, which is colored based on the group. PC1, PC2 and PC3 represent the top three principal coordinates that captured most of the diversity. The explanation of diversity captured by the coordinate is given as a percentage. (G) Relative abundance of the microbiota at genus level. (H) Differences in the bacterial taxon between patients with HMFD and control children. The beta coefficient was calculated with the multivariate association with linear models 2 (MaAsLin2) after adjusting for age and sex.
Analysis of the bacterial genera stack plots of the HFMD and control groups revealed that Faecalibacterium, Prevotella, Bacteroidetes, Escherichia, Parabacteroides and Bifidobacterium were the major genera in both groups (Figure 1G). To further identify the bacteria that differed between the HFMD and control groups, we performed a MaAsLin2 analysis using age and sex as confounders and identified genera that exhibited significant differences (Figure 1H). Prevotella, Cetobacterium, and Megamonas were enriched in the feces of patients with HFMD compared with those of the control group, whereas Bifidobacterium and Faecalibacterium were depleted in patients with HFMD.
Previous studies have revealed that age and sex should be considered co-factors in analysis of the gut microbiota (Xiu et al., 2021). Thus, we employed Propensity Score Matching (PSM) to adjust for age and sex in our study population. After PSM, the mean age of two cohorts was 4.0 ± 1.2 years. The majority of subjects (63%) were male. There were significant differences in various supplement types between matched groups. In the hygienic habits analysis, washing hands after playing outside, sucking fingers and rolling and playing on the ground presented significantly difference between two groups (Supplementary Table 1). In the adjusted cohort, the alpha-diversity indices of the HFMD group were significantly lower than those of the control group (Figures 2A–D). The PCoA plot also showed that the gut flora of the HFMD group was different from that of the control group (Figures 2E, F). The majority of genera in this adjusted cohort were in consistence with the previous results (Figure 2G). We then performed the linear discriminant analysis effect size (LEfSe) analysis and identified genera that exhibited significant differences, including Prevotella, Cetobacterium, Megamonas, Bifidobacterium and Faecalibacterium (Figure 2H).
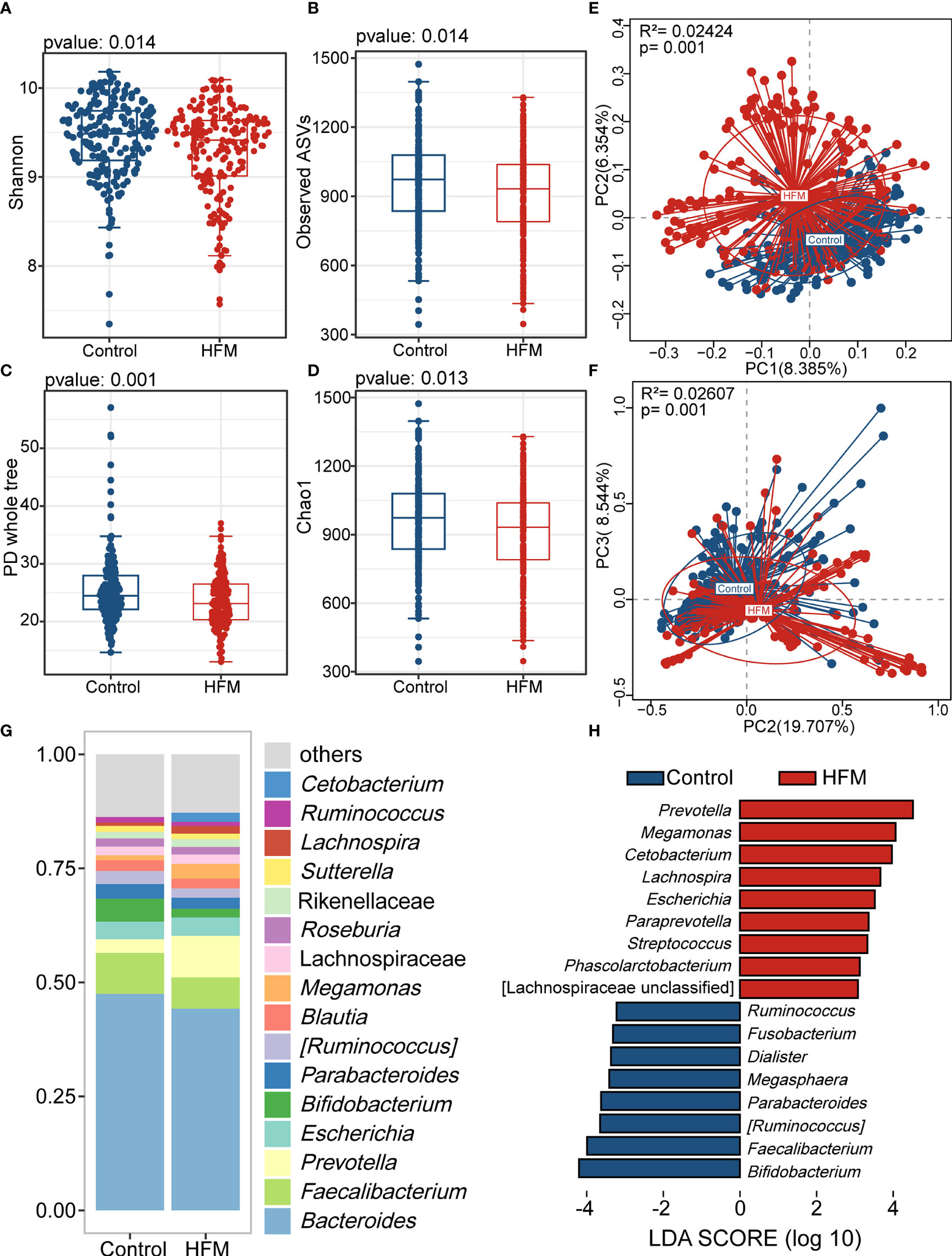
Figure 2 Altered bacterial microbiota biodiversity and composition in HFMD based on cohort matched age and sex. (A–D) Alpha diversity. Comparison of alpha-diversity indices (Shannon, observed ASVs, PD whole tree and Chao 1 index) between the HFMD and control groups (Wilcoxon rank-sum tests). (E, F) Principal coordinate analysis based on unweighted and weighted UniFrac distances revealed that the bacterial communities of patients with HFMD clustered separately from the bacterial communities of healthy control children. Each point represents a single sample, which is colored based on the group. PC1, PC2 and PC3 represent the top three principal coordinates that captured most of the diversity. The explanation of diversity captured by the coordinate is given as a percentage. (G) Relative abundance of the bacterial microbiota at the genus level. (H) Differences in the bacterial taxon abundance between patients with HMFD and control children by using Linear discriminant analysis effect size (LEfSe) analysis. HFMD-enriched taxa are indicated with a positive LDA score, and taxa enriched in healthy controls have a negative score. Only taxa meeting an LDA significance threshold of >3 are shown. LDA, linear discriminant analysis.
In the present study, there were 144 patients infected EV71 while 21 patients infected CVA16. Shannon index of EV71 group was lower than control group and PCoA analysis showed a longer distance of EV71 to control group comparing with CVA16 (Figure 3). In the analysis within each sex group, the results echoed our results mentioned above. The PD whole tree index was lower in HFMD group comparing with control group and both showed significant gut microbiome compositional shift. Prevotella, Cetobacterium and Megamonas were enriched while Faecalibacterium and Bifidobacterium were depleted in HFMD groups in both male and female group (Supplementary Figures 2, 3).
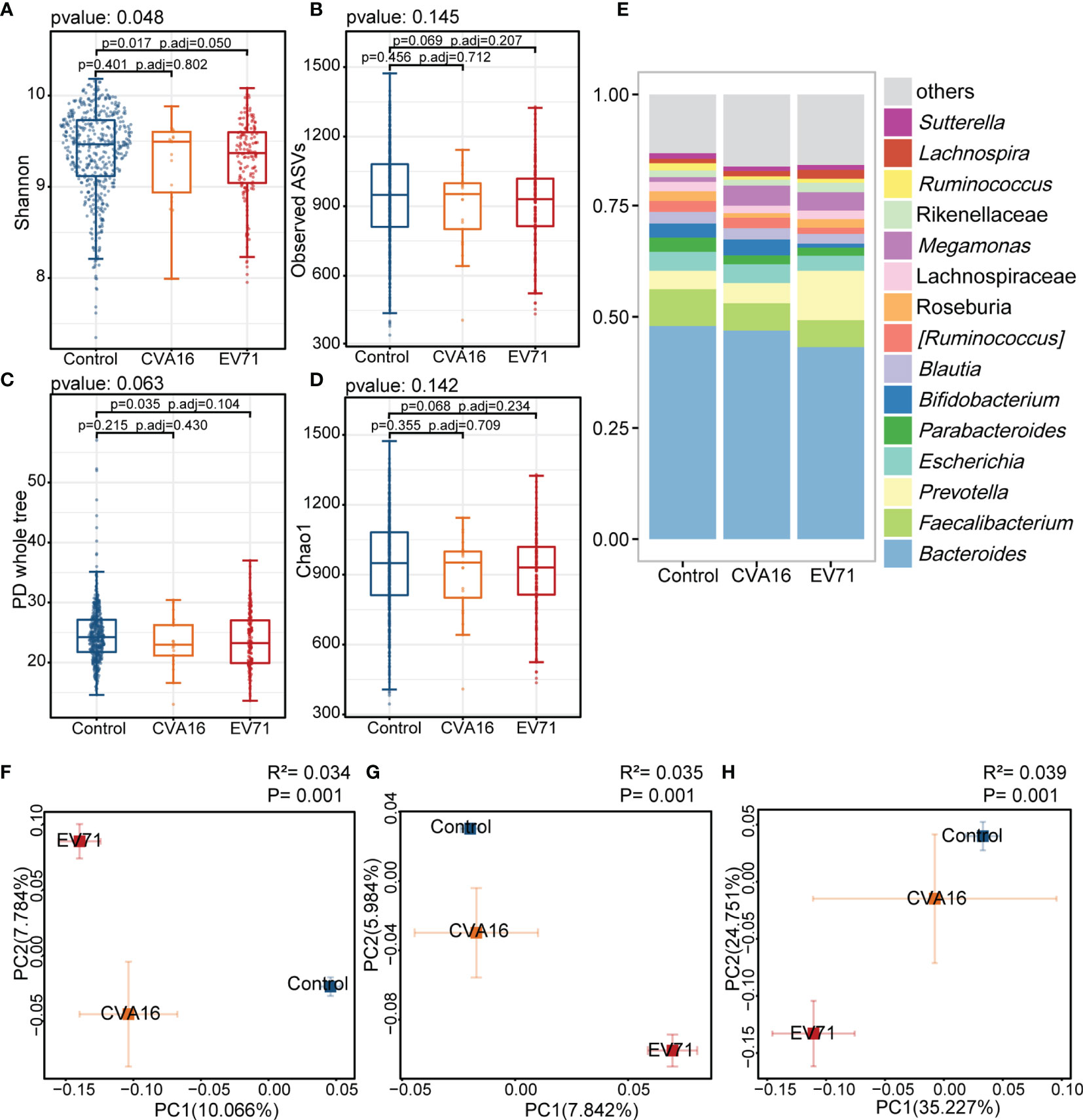
Figure 3 Comparison of the gut microbiota between EV71, CVA16, and control group. (A–D) Alpha diversity. Comparison of alpha-diversity indices (Shannon, observed ASVs, PD whole tree and Chao 1 index) among EV71-infected HFMD, CVA16-infected HFMD and control groups (Kruskal-Wallis tests). (E) Relative abundance of the bacterial microbiota at the genus level among EV71, CVA16, and control group. (F–H) Beta diversity. Principal coordinate analysis (PCoA) based on the Bray-Curtis distance (F), unweighted UniFrac distances (G) and weighted UniFrac distances (H) revealed that the relationship of bacterial communities among EV71-infected HFMD, CVA16-infected HFMD and control groups. The explanation of diversity captured by the coordinate is given as a percentage.
These results showed that the gut microbiota was altered in children with HFMD, which indicated that the gut microbiota were associated with hand, foot, and mouth disease.
Synbiotics Supplements Exerted a Protective Effect in HFMD
Probiotics and prebiotics supplements have been demonstrated to play positive roles in the modulation of the gut microbiota (Ki et al., 2012; Liu et al., 2015). To examine whether probiotics, prebiotics and synbiotics supplements exerted a protective effect on HFMD, we performed logistic regression models to calculate the odds ratios of HFMD for probiotics, prebiotics and synbiotics supplement use. After adjusting for age and sex, the odds ratios (ORs) (95% CIs) for HFMD according to the application of probiotics, prebiotics and synbiotics were 1.65 (0.55,4.92), 1.59 (1.07,2.37) and 0.49 (0.29,0.82), respectively (Supplementary Table 2). In PSM-adjusted cohort, the ORs of all groups (95% CIs) were reduced to 1.34 (0.39, 4.65), 1.8 (1.12, 2.9) and 0.45 (0.25, 0.81), respectively (Supplementary Table 3). We further adjusted for age, sex and hygiene, and consistent findings were observed in Table 2. The results indicated that children who utilized synbiotics were at lower risk of infection than those who used probiotics alone or prebiotics alone.
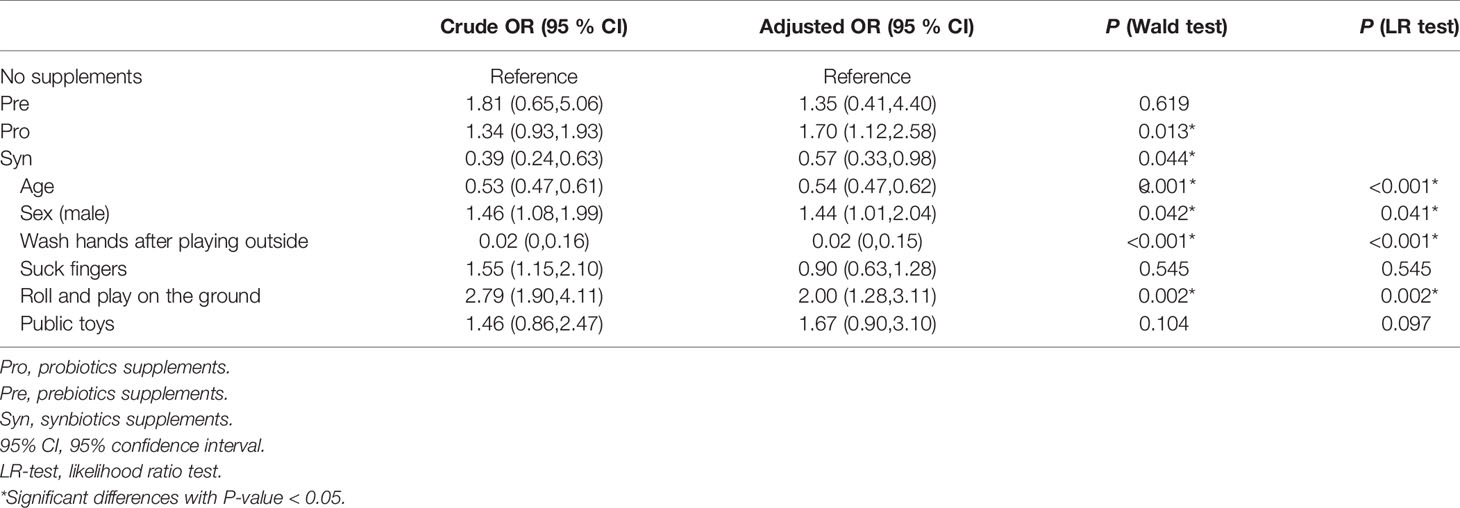
Table 2 Effects of the intake of probiotics, prebiotics, or synbiotics on gut microbiota of patients with HFMD after adjusting for age, sex and hygiene.
Effect of Probiotics, Prebiotics and Synbiotics on the Gut Microbiota of Children With HFMD
In the studied cohort, the number of patients who received prebiotics was relatively small (n=8), and we thus had to perform an analysis of the gut microbiota without considering the population administered prebiotics alone. The PCoA plot showed that the gut microbiota structure of the patients who used synbiotics supplements was closer to that of the control group than that of patients with probiotics alone supplements and patients without supplement groups, as could be observed with different distance algorithms (Figures 4A–C). Most of the genera found in each group were similar, and the main genera were Faecalibacterium, Prevotella, Bacteroidetes, Escherichia, Parabacteroides and Bifidobacterium (Figure 4D). The abundance of genera identified as significantly altered in HFMD showed less extensive changes in the patients who were administered synbiotics. Faecalibacterium was depleted in patients who were not administered any supplement and was restored in patients applied synbiotics supplements (Figure 4E). Cetobacterium, Megamonas and Prevotella were increased in patients with probiotic supplements but tended to decrease in the patients applied synbiotics supplements (Figures 4F–H). These data suggested that the gut microbiota of the children who were administered synbiotics supplements showed potential resistance to HFMD-related dysbiosis.

Figure 4 Comparison of the gut microbiota among the probiotics (Pro), prebiotics (Pre) and Synbiotics (Syn) supplements. (A–C) Beta diversity. Principal coordinate analysis (PCoA) based on the Bray-Curtis distance (A), unweighted UniFrac distances (B) and weighted UniFrac distances (C) revealed the relationship of bacterial communities among the Pro, Pre and Syn groups. The explanation of diversity captured by the coordinate is given as a percentage. (D) Relative abundance at the genus level of the different groups. (E–H) Relative abundances of Faecalibacterium, Cetobacterium, Megamonas and Prevotella between various groups (Kruskal-Wallis rank sum test and multiple comparison between groups adjusted by Benjamini-Hochberg method). * indicates P < 0.05, ** indicates P < 0.01, *** indicates P < 0.001, # indicates P < 0.1.
Discussion
HFMD has become an emerging public health problem in Asia, but there is currently no vaccine to protect against the responsible viruses. The present study aimed to seek answers to unanswered questions related to the gut microbiota. The differences in the gut microbiota patterns of patients with HFMD compared with those of the healthy group and the effect of synbiotics supplements on the intestinal flora of patients with HFMD were explored. We observed that the gut microbiota were altered in children with HFMD. Furthermore, children with HFMD who were administered synbiotics supplements exhibited reduced morbidity, and their gut microbiota showed potential resistance to HFMD-related gut flora dysbiosis.
First, the distribution characteristics of the gut microbiota of patients with HFMD differed from those of the gut microbiota of the healthy group. A reduction in bacterial diversity, which is considered to indicate an unhealthy state, has been observed in children with HFMD (Rivera-Chávez et al., 2016; Sassone-Corsi et al., 2016; Kriss et al., 2018). This result was consistent with previous research, which demonstrated that changes in the diversity of butyrate-producing bacteria and inflammation-inducing bacteria are associated with HFMD. In the present study, several bacteria were found to differ between the HFMD and control groups. Our results showed that Prevotella and Streptococcus were found at higher relative abundances in patients with HFMD than in the healthy group. Prevotella is widely considered a commensal genus, and a series of controversial reports on its pathogenicity have been published (Brook, 1998; Brook, 2004; Nagy, 2010). For example, Prevotella copri, a widely distributed species, has been linked to inflammatory diseases and intestinal barrier dysfunction by directly promoting the upregulation of inflammatory cytokines. Additionally, Prevotella is a lipopolysaccharide-producing anaerobic gram-negative bacterium that can cause inflammatory reactions and damage the gut barrier (Hofstad et al., 1993; Choi et al., 2011). Streptococcus, which is one of the most classic opportunistic pathogens, comprises a wide variety of pathogenic bacterial strains. Streptococcus is one of the most prevalent inhabitants of the respiratory tract (Engholm et al., 2017), skin surface (Ho et al., 2021) and gut (Sokol et al., 2017; Ooi et al., 2018) of adults and children. Studies have shown that influenza virus can interact with S. pneumoniae and induce susceptibility to viruses by activating cytokines and dendritic cells (Short et al., 2012). The enrichment of Streptococcus has also been observed in the gut microbiota of patients with viral hepatitis cirrhosis (Wang et al., 2019). The cause of HFMD is enteral virus infection, e.g., infection with EV71 and CVA16. A study found that EV71 is transported from the intestine to peripheral tissue and other organs, which suggests that the small intestine is a gateway for EV71 infection in vivo (Feng et al., 2020). EV71 infection could reduce the expression of goblet cell-derived mucins (Good et al., 2019), which indicates that EV71 infection could alter the luminal environment of the gut microbiota. Previous data have confirmed that EV71 infect and replicate in intestinal epithelial cells by activating the p38 mitogen-activated protein kinase (MAPK) and protein kinase signaling pathways. This inflammatory pathway overlaps with the inflammatory reactions caused by bacterial lipopolysaccharide or translocation. In addition, EV71 is most frequently associated with severe diseases (Shu et al., 2015). In our study, the gut microbiota of patients infected with EV71 were more dysbiotic than that of patients with CVA16. Hence, in HFMD pathogenesis, the observed gut dysbiosis might interact with enteroviruses.
In contrast, the relative abundances of Faecalibacterium and Bifidobacterium were decreased in the feces of the HFMD group. These two bacterial genera, which are generally considered producers of short-chain fatty acids, are reportedly capable of regulating intestinal epithelial nutrition, stabilizing immune homeostasis and reinforcing intestinal barrier functions (Hiippala et al., 2018; Routy et al., 2018). Bifidobacteria protect gut mucus against diet-induced microbiota-mediated deterioration, and gut barrier defects (Schroeder et al., 2018). Bifidobacteria have also been identified as health-promoting genera that increases butyrate production and thus contributes to improvements in the gut barrier and metabolic outcomes (Seganfredo et al., 2017). Similarly, Faecalibacterium exhibits an enriched polyphenol-rich dietary pattern, which could improve gut barrier function (Del Bo' et al., 2020), and the prophylactic use of Faecalibacterium could also prevent the acute breakdown of the colonic epithelial barrier (Lapiere et al., 2020). Thus, in both the research and healthcare product markets, Bifidobacterium is a very common commercialized probiotics, and Faecalibacterium shows strong promise as effective and safe probiotics.
In clinical practice, probiotics and prebiotics are widely used as supplements for healthy children or are included in therapeutic approaches for many diseases (Krumbeck et al., 2016). In this study, we found that children who were administered supplements exhibited a lower risk of HFMD-related infection, whereas the application of probiotics or prebiotics alone was not found to be significantly associated with HFMD risk. This very interesting finding indicates that synbiotics was related to an improved protective effect. A series of studies conducted by Professor D. A. Mills from the University of California Davis revealed that Bifidobacterium can process the prebiotics in human milk oligosaccharides and that the colonization of the probiotics Bifidobacterium breve could also be influenced by prebiotics (Underwood et al., 2017). These studies suggested a potential close association between probiotics and prebiotics, and this association is also being investigated by Professor Mills’s group. The intake of prebiotics may modulate the composition of the microbiota, be fermented by the beneficial gut microbiota and increase the abundance of probiotics (Gibson and Roberfroid, 1995; Pandey et al., 2015). Synbiotics, defined as combinations of probiotics and prebiotics, could benefit the host by modulating the gut microbiota (Gibson and Roberfroid, 1995). Previous studies have reported that synbiotics exert protective or preventive effects against gastrointestinal or other diseases by maintaining the gut flora structure (Shimizu et al., 2018; Tian et al., 2018). Similarly, in our study, the beta diversity analysis showed that the composition of the gut flora of patients applied synbiotics was closer to that of the control group than to those of the other patient groups. The analysis of the abundances of identified biomarkers indicated their relative restoration and depletion in patients applied synbiotics compared with those found in the other patient groups. These gut microbiota alterations suggested that the gut microbiota of the children who received synbiotics supplements could acquire resistance to HFMD-related gut flora changes, which might manifest as a reduction in HFMD susceptibility.
Our study has some limitations. In the present study, we only collected information regarding whether the children were administered supplements, including probiotics and prebiotics. However, the specific types and dose of probiotics or prebiotics are unknown. Besides, prebiotics are also found in food (Roberta, et al., 2020), especially those that contain complex carbohydrates, such as fiber and resistant starch. Diet information wasn’t collected and dietary prebiotics assessment were required in further study. Another major limitation is that all the samples from the HFMD group were collected after infection. Thus, the analysis could only suggest a potential bacterial resistance in the children who were administered synbiotics. However, the existence of this resistance in the gut microbiota should be validated in future follow-up studies with repeated and larger populations sampled before and after infection.
To our knowledge, the present study involves the largest HFMD cohort used for gut microbiota analyses thus far. We found that the gut microbiota of children with HFMD exhibits dysbiosis, and synbiotics supplements might provide a potential protective effect against HFMD. However, further studies with a larger population and detailed information are needed to validate our findings and explore the underlying mechanisms of HFMD. In conclusion, our study indicated the changes in the composition of the gut flora of patients with HFMD and identified potential bacterial biomarkers of HFMD. Additionally, we provided new evidence showing that synbiotics might lower the susceptibility to HFMD by modulating the gut microbiota.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ebi.ac.uk/ena/browser/view/PRJEB45719.
Ethics Statement
The studies involving human participants were reviewed and approved by Ethical Review Committee of Chinese Center for Disease Control and Prevention. Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin.
Author Contributions
WW, HZ, and XY designed the study. ZR, CZ, TL, YW, and SC processed the samples. XG and ZL searched literature, analyzed the data, and drafted the manuscript. WW, HZ, and XY reviewed and revised the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was funded by the Science and Technology Planning Project Foundation of Guangzhou city (202102080584).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The study was supported by the Science and Technology Planning Project of Guangzhou city, China. We thank the staff from Heyuan and Jiangmen center for disease control organized the collection points and trained local hospital and community investigators. We thank the staff from Guangdong Provincial Center for Disease Control and Prevention for excellent support during the field work.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2021.729756/full#supplementary-material
References
Alam, A., Neish, A. (2018). Role of Gut Microbiota in Intestinal Wound Healing and Barrier Function. Tissue Barriers. 6, 1539595. doi: 10.1080/21688370.2018.1539595
Brook, I. (1998). Microbiology of Common Infections in the Upper Respiratory Tract. Prim Care 25, 633–648. doi: 10.1016/s0095-4543(15)30006-3
Brook, I. (2004). Anaerobic Pulmonary Infections in Children. Pediatr. Emerg. Care 20, 636–640. doi: 10.1097/01.pec.0000139751.63624.0b
Choi, E. Y., Jin, J. Y., Lee, J. Y., Choi, J. I., Choi, I. S., Kim, S. J. (2011). Melatonin Inhibits Prevotella Intermedia Lipopolysaccharide-Induced Production of Nitric Oxide and Interleukin-6 in Murine Macrophages by Suppressing NF-κb and STAT1 Activity. J. Pineal Res. 50, 197–206. doi: 10.1111/j.1600-079X.2010.00829.x
Del Bo', C., Bernardi, S., Cherubini, A., Porrini, M., Gargari, G., Hidalgo-Liberona, N., et al. (2020). A Polyphenol-Rich Dietary Pattern Improves Intestinal Permeability, Evaluated as Serum Zonulin Levels, in Older Subjects: The MaPLE Randomised Controlled Trial. Clin. Nutr. 40, 3006–3018. doi: 10.1016/j.clnu.2020.12.014
Engholm, D. H., Kilian, M., Goodsell, D. S., Andersen, E. S., Kjærgaard, R. S. (2017). A Visual Review of the Human Pathogen Streptococcus Pneumoniae. FEMS Microbiol. Rev. 41, 854–879. doi: 10.1093/femsre/fux037
Feng, Q., Zhou, H., Zhang, X., Liu, X., Wang, J., Zhang, C., et al. (2020). Acarbose, as a Potential Drug, Effectively Blocked the Dynamic Metastasis of EV71 From the Intestine to the Whole Body. Infect. Genet. Evol. 81, 104210. doi: 10.1016/j.meegid.2020.104210
Gibson, G. R., Roberfroid, M. B. (1995). Dietary Modulation of the Human Colonic Microbiota: Introducing the Concept of Prebiotics. J. Nutr. 125, 1401–1412. doi: 10.1093/jn/125.6.1401
Good, C., Wells, A. I., Coyne, C. B. (2019). Type III Interferon Signaling Restricts Enterovirus 71 Infection of Goblet Cells. Sci. Adv. 5, eaau4255. doi: 10.1126/sciadv.aau4255
Heidrich, B., Vital, M., Plumeier, I., Döscher, N., Kahl, S., Kirschner, J., et al. (2018). Intestinal Microbiota in Patients With Chronic Hepatitis C With and Without Cirrhosis Compared With Healthy Controls. Liver Int. 38, 50–58. doi: 10.1111/liv.13485
Hiippala, K., Jouhten, H., Ronkainen, A., Hartikainen, A., Kainulainen, V., Jalanka, J., et al. (2018). The Potential of Gut Commensals in Reinforcing Intestinal Barrier Function and Alleviating Inflammation. Nutrients 10, 988. doi: 10.3390/nu10080988
Himel, M. (2021). Multivariable Association Discovery in Population-Scale Meta-Omics Studies. In: Biorxiv. Available at: https://huttenhower.sph.harvard.edu/maaslin (Accessed January 20, 2021).
Hofstad, T., Skaug, N., Sveen, K. (1993). Stimulation of B Lymphocytes by Lipopolysaccharides From Anaerobic Bacteria. Clin. Infect. Dis. 16, 200–202. doi: 10.1093/clinids/16.Supplement_4.S200
Ho, S. X., Min, N., Wong, E. P. Y., Chong, C. Y., Chu, J. J. H. (2021). Characterization of Oral Virome and Microbiome Revealed Distinctive Microbiome Disruptions in Paediatric Patients With Hand, Foot and Mouth Disease. NPJ Biofilms Microbiomes. 7, 19. doi: 10.1038/s41522-021-00190-y
Jakobsson, H. E., Rodríguez-Piñeiro, A. M., Schütte, A., Ermund, A., Boysen, P., Bemark, M., et al. (2015). The Composition of the Gut Microbiota Shapes the Colon Mucus Barrier. EMBO Rep. 16, 164–177. doi: 10.15252/embr.201439263
Ki, C. B., Mun, J. S., Hwan, C. C., Song, I. D., Woong, L. H., Joon, K. H., et al. (2012). The Effect of a Multispecies Probiotic Mixture on the Symptoms and Fecal Microbiota in Diarrhea-Dominant Irritable Bowel Syndrome: A Randomized, Double-Blind, Placebo-Controlled Trial. J. Clin. Gastroenterol. 46, 220–227. doi: 10.1097/MCG.0b013e31823712b1
Kriss, M., Hazleton, K. Z., Nusbacher, N. M., Martin, C. G., Lozupone, C. A. (2018). Low Diversity Gut Microbiota Dysbiosis: Drivers, Functional Implications and Recovery. Curr. Opin. Microbiol. 44, 34–40. doi: 10.1016/j.mib.2018.07.003
Krumbeck, J. A., Maldonado-Gomez, M. X., Ramer-Tait, A. E., Hutkins, R. W. (2016). Prebiotics and Synbiotics: Dietary Strategies for Improving Gut Health. Curr. Opin. Gastroenterol. 32, 110–119. doi: 10.1097/MOG.0000000000000249
Lapiere, A., Geiger, M., Robert, V., Demarquay, C., Auger, S., Chadi, S., et al. (2020). Prophylactic Faecalibacterium Prausnitzii Treatment Prevents the Acute Breakdown of Colonic Epithelial Barrier in a Preclinical Model of Pelvic Radiation Disease. Gut Microbes 12, 1–15. doi: 10.1080/19490976.2020.1812867
Liu, X., Cao, S., Zhang, X. (2015). Modulation of Gut Microbiota-Brain Axis by Probiotics, Prebiotics, and Diet. J. Agric. Food Chem. 63, 7885–7895. doi: 10.1021/acs.jafc.5b02404
McDonald, D., Price, M., Goodrich, J., Nawrocki, E., DeSantis, T., Probst, A., et al. (2012). An Improved Greengenes Taxonomy With Explicit Ranks for Ecological and Evolutionary Analyses of Bacteria and Archaea. ISME J 6, 610–618. doi: 10.1038/ismej.2011.139
Ministry of Health of the People’s Republic of China. (2010). Diagnosis and Treatment Guideline on Hand-Foot-Mouth Disease. Available at: http://yzs.satcm.gov.cn/gongzuodongtai/2018-03-24/3073.html (Accessed March 1, 2017).
Nagy, E. (2010). Anaerobic Infections: Update on Treatment Considerations. Drugs 70, 841–858. doi: 10.2165/11534490-000000000-00000
Odenwald, M. A., Turner, J. R. (2017). The Intestinal Epithelial Barrier: A Therapeutic Target? Nat. Rev. Gastroenterol. Hepatol. 14, 9–21. doi: 10.1038/nrgastro.2016.169
Ooi, C. Y., Syed, S. A., Rossi, L., Garg, M., Needham, B., Avolio, J., et al. (2018). Impact of CFTR Modulation With Ivacaftor on Gut Microbiota and Intestinal Inflammation. Sci. Rep. 8, 17834. doi: 10.1038/s41598-018-36364-6
Pandey, K. R., Naik, S. R., Vakil, B. V. (2015). Probiotics, Prebiotics and Synbiotics- a Review. J. Food Sci. Technol. 52, 7577–7587. doi: 10.1007/s13197-015-1921-1
Rivera-Chávez, F., Zhang, L. F., Faber, F., Lopez, C. A., Byndloss, M. X., Olsan, E. E., et al. (2016). Depletion of Butyrate-Producing Clostridia From the Gut Microbiota Drives an Aerobic Luminal Expansion of Salmonella. Cell Host Microbe 19, 443–454. doi: 10.1016/j.chom.2016.03.004
Roberta, M. S., de, A., Sara, S., Célia M. da, S. F. C., Mariana, V., Eduardo, C., et al. (2020). Potential Prebiotic Effect of Fruit and Vegetable Byproducts Flour Using In Vitro Gastrointestinal Digestion. Food Res. Int. 137, 109354. doi: 10.1016/j.foodres.2020.109354
Robinson, C. M. (2019). Enteric Viruses Exploit the Microbiota to Promote Infection. Curr. Opin. Virol. 37, 58–62. doi: 10.1016/j.coviro.2019.06.002
Routy, B., Gopalakrishnan, V., Daillère, R., Zitvogel, L., Wargo, J. A., Kroemer, G. (2018). The Gut Microbiota Influences Anticancer Immunosurveillance and General Health. Nat. Rev. Clin. Oncol. 15, 382–396. doi: 10.1038/s41571-018-0006-2
Sanz, Y., De, P. G. (2009). Gut Microbiota Probiotics in Modulation of Epithelium and Gut-Associated Lymphoid Tissue Function. Int. Rev. Immunol. 28, 397–413. doi: 10.3109/08830180903215613
Sassone-Corsi, M., Nuccio, S. P., Liu, H., Hernandez, D., Vu, C. T., Takahashi, A. A., et al. (2016). Microcins Mediate Competition Among Enterobacteriaceae in the Inflamed Gut. Nature 540, 280–283. doi: 10.1038/nature20557
Schroeder, B. O., Birchenough, G. M. H., Ståhlman, M., Arike, L., Johansson, M. E. V., Hansson, G. C., et al. (2018). Bifidobacteria or Fiber Protects Against Diet-Induced Microbiota-Mediated Colonic Mucus Deterioration. Cell Host Microbe 23, 27–40.e7. doi: 10.1016/j.chom.2017.11.004
Schuijt, T. J., Lankelma, J. M., Scicluna, B. P., de Sousa e Melo, F., Roelofs, J. J., de Boer, J. D., et al. (2015). The Gut Microbiota Plays a Protective Role in the Host Defence Against Pneumococcal Pneumonia. Gut 65, 575–583. doi: 10.1136/gutjnl-2015-309728
Seganfredo, F. B., Blume, C. A., Moehlecke, M., Giongo, A., Casagrande, D. S., Spolidoro, J. V. N., et al. (2017). Weight-Loss Interventions and Gut Microbiota Changes in Overweight and Obese Patients: A Systematic Review. Obes. Rev. 18, 832–851. doi: 10.1111/obr.12541
Segata, N., Izard, J., Waldron, L., Gevers, D., Miropolsky, L., Garrett, W. S., et al. (2011). Nomic Biomarker Discovery and Explanation. Genome Biol. 12, 60. doi: 10.1186/gb-2011-12-6-r60
Shimizu, K., Yamada, T., Ogura, H., Mohri, T., Kiguchi, T., Fujimi, S., et al. (2018). Synbiotics Modulate Gut Microbiota and Reduce Enteritis and Ventilator-Associated Pneumonia in Patients With Sepsis: A Randomized Controlled Trial. Crit. Care 22, 239. doi: 10.1186/s13054-018-2167-x
Short, K. R., Habets, M. N., Hermans, P. W., Diavatopoulos, D. A. (2012). Interactions Between Streptococcus Pneumoniae and Influenza Virus: A Mutually Beneficial Relationship? Future Microbiol. 7, 609–624. doi: 10.2217/fmb.12.29
Shu, Y. Z., Mei, Y. X., Hong, M. X., Xiu, J. L., Shu, J. D., Xian, J. W., et al. (2015). Immunologic Characterization of Cytokine Responses to Enterovirus 71 and Coxsackievirus A16 Infection in Children. Med. (Baltimore). 94, e1137. doi: 10.1038/s41571-018-0006-2
Sokol, H., Leducq, V., Aschard, H., Pham, H. P., Jegou, S., Landman, C., et al. (2017). Fungal Microbiota Dysbiosis in IBD. Gut 66, 1039–1048. doi: 10.1136/gutjnl-2015-310746
Sun, Y., Ma, Y., Lin, P., Tang, Y. W., Yang, L., Zhong, S. Y., et al. (2016). Fecal Bacterial Microbiome Diversity in Chronic HIV-Infected Patients in China. Emerg. Microbes Infect. 5, e31. doi: 10.1038/emi.2016.25
Takiishi, T., Morales Fenero, C. I., Saraiva Câmara, N. O. (2017). Intestinal Barrier and Gut Microbiota: Shaping Our Immune Responses Throughout Life. Tissue Barriers. 5, e1373208. doi: 10.1080/21688370.2017.1373208
Tian, X., Pi, Y. P., Liu, X. L., Chen, H., Chen, W. Q. (2018). Supplemented Use of Pre-, Pro-, and Synbiotics in Severe Acute Pancreatitis: An Updated Systematic Review and Meta-Analysis of 13 Randomized Controlled Trials. Front. Pharmacol. 9, 690. doi: 10.3389/fphar.2018.00690
Underwood, M. A., Davis, J. C. C., Kalanetra, K. M., Gehlot, S., Patole, S., Tancredi, D. J., et al. (2017). Digestion of Human Milk Oligosaccharides by Bifidobacterium Breve in the Premature Infant. J. Pediatr. Gastroenterol. Nutr. 65, 449–455. doi: 10.1097/MPG.0000000000001590
Wang, Y., Pan, C. Q., Xing, H. (2019). Advances in Gut Microbiota of Viral Hepatitis Cirrhosis. BioMed. Res. Int. 2019, 9726786. doi: 10.1155/2019/9726786
Winglee, K., Eloe-Fadrosh, E., Gupta, S., Dan, G. H., Fraser, C., Bishai, W., et al. (2014). Aerosol Mycobacterium tuberculosis Infection Causes Rapid Loss of Diversity in Gut Microbiota. PloS One 9, e97048. doi: 10.1371/journal.pone.0097048
Wu, S., Jiang, Z. Y., Sun, Y. F., Yu, B., Chen, J., Dai, C. Q., et al. (2013). Microbiota Regulates the TLR7 Signaling Pathway Against Respiratory Tract Influenza A Virus Infection. Curr. Microbiol. 67, 414–422. doi: 10.1007/s00284-013-0380-z
Xiu, Y. Z., Huan, Z. Z., Yu, F. L., Shi, Z., Hua, H. R., Zhe, Z., et al. (2021). Sex- and Age-Related Trajectories of the Adult Human Gut Microbiota Shared Across Populations of Different Ethnicities. Nat. Aging 1, 87–100. doi: 10.1038/s43587-020-00014-2
Keywords: hand, foot, and mouth disease, gut microbiota, synbiotics, probiotics, prebiotics
Citation: Guo X, Lan Z, Wen Y, Zheng C, Rong Z, Liu T, Chen S, Yang X, Zheng H and Wu W (2021) Synbiotics Supplements Lower the Risk of Hand, Foot, and Mouth Disease in Children, Potentially by Providing Resistance to Gut Microbiota Dysbiosis. Front. Cell. Infect. Microbiol. 11:729756. doi: 10.3389/fcimb.2021.729756
Received: 23 June 2021; Accepted: 27 August 2021;
Published: 30 September 2021.
Edited by:
George Seghal Kiran, Pondicherry University, IndiaReviewed by:
Ravichandra Vemuri, Wake Forest Baptist Medical Center, United StatesHai-Feng Pan, Anhui Medical University, China
Copyright © 2021 Guo, Lan, Wen, Zheng, Rong, Liu, Chen, Yang, Zheng and Wu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Wei Wu, cnnjwuwei@163.com; Huimin Zheng, zhenghuimin91@126.com; Xingfen Yang, xfyang@vip.163.com
†These authors have contributed equally to this work and share first authorship