 Sandrine Bessette1,2,3
Sandrine Bessette1,2,3 Yann Moalic1,2,3
Yann Moalic1,2,3 Sébastien Gautey1,2,3
Sébastien Gautey1,2,3 Françoise Lesongeur1,2,3Anne Godfroy1,2,3
Françoise Lesongeur1,2,3Anne Godfroy1,2,3 Laurent Toffin1,2,3*
Laurent Toffin1,2,3*- 1Institut Carnot Ifremer EDROME, Centre de Bretagne, REM/EEP, Laboratoire de Microbiologie des Environnements Extrêmes, UMR 6197, Plouzané, France
- 2Laboratoire de Microbiologie des Environnements Extrêmes, Institut Universitaire Européen de la Mer, UMR 6197, Université de Bretagne Occidentale, Plouzané, France
- 3CNRS, Laboratoire de Microbiologie des Environnements Extrêmes, Technopôle Brest Iroise, UMR 6197, Plouzané, France
Sitting at ∼5,000 m water depth on the Congo-Angola margin and ∼760 km offshore of the West African coast, the recent lobe complex of the Congo deep-sea fan receives large amounts of fluvial sediments (3–5% organic carbon). This organic-rich sedimentation area harbors habitats with chemosynthetic communities similar to those of cold seeps. In this study, we investigated relative abundance, diversity and distribution of aerobic methane-oxidizing bacteria (MOB) communities at the oxic–anoxic interface of sedimentary habitats by using fluorescence in situ hybridization and comparative sequence analysis of particulate mono-oxygenase (pmoA) genes. Our findings revealed that sedimentary habitats of the recent lobe complex hosted type I and type II MOB cells and comparisons of pmoA community compositions showed variations among the different organic-rich habitats. Furthermore, the pmoA lineages were taxonomically more diverse compared to methane seep environments and were related to those found at cold seeps. Surprisingly, MOB phylogenetic lineages typical of terrestrial environments were observed at such water depth. In contrast, MOB cells or pmoA sequences were not detected at the previous lobe complex that is disconnected from the Congo River inputs.
Introduction
Deep-sea fans, also known as deep-sea turbidite systems, trap enormous amounts of terrestrial organic matter (OM) inputs delivered from continental shelves to deep ocean basins by rivers. Therefore, deep-sea fans are among the largest sedimentary systems on earth and contain large reservoirs of organic carbon buried in marine sediments (Shanmugam and Moiola, 1988). The Quaternary Congo deep-sea fan located on the Congo-Angola passive margin, is one of the largest mud-rich fans (300,000 km2) in the world and constitutes the major carbon burial region in the southeastern Atlantic Ocean (Rabouille et al., 2009). Due to its direct connection to the Congo River, the second largest river in the world in terms of fluvial discharge, continuous massive sedimentary inputs are transferred via turbidity currents following the path of the canyon deeply incised in the shelf (Khripounoff et al., 2003), regardless of sea level fluctuations (Droz et al., 2003). These sedimentary inputs flow 760 km off the Congo-Angola margin along the present-day active channel-levee system, which ends with lobe-shaped sedimentary deposits called the recent lobe complex (Savoye et al., 2009). The accumulated sediments (1.25 × 106 t of organic carbon yr-1) of the recent lobe complex contain high organic carbon content (up to 5 wt% TOC; Baudin et al., 2017b) of terrestrial origin (70–80%; Stetten et al., 2015; Baudin et al., 2017b, which were deposited at extremely high sedimentation rates (>2–20 mm yr-1) (Stetten et al., 2015; Rabouille et al., 2016). As a consequence, substantial localized degradation of recent OM forms diffuse seepages enriched in methane (CH4) with hydrogen sulfide rising upward (Khripounoff et al., 2015) and a steep oxygen (O2) gradient at the sediment-water interface (Rabouille et al., 2009). These CH4-enriched sedimentary areas support remarkable chemosynthetic fauna on the seafloor that resemble the chemosynthetic communities associated with cold seeps (Rabouille et al., 2016).
The major biological sink of CH4 produced in sulfate-depleted anoxic sediments is anaerobic CH4 oxidation (Boetius and Wenzhofer, 2013). However, an unknown fraction of the CH4 rising upward through oxygenated sediments bypasses this benthic CH4 filter and is consumed by aerobic methane-oxidizing bacteria (MOB). MOB utilizes CH4 as their sole carbon and energy source at the sediment-water interface when oxygen from bottom waters is available as electron acceptor (Boetius and Wenzhofer, 2013). In disturbed seeps, such as the center of active Haakon Mosby Mud Volcano (HMMV) where anaerobic CH4 oxidation is repressed, or at Hikurangi Margin where the bioirrigation of the sediment-dwelling organisms enhances O2 advection in surface sediments (e.g., Hikurangi Margin), aerobic CH4 oxidation has been suggested to act as an efficient benthic filter regulating CH4 efflux at the surface sediments (Niemann et al., 2006; Thurber et al., 2013).
The first step of aerobic CH4 oxidation is performed by the particulate methane mono-oxygenase enzyme, encoded by the pmoCAB operon. All MOB described so far possess the canonical pmoA gene, with the exception of Methylocella spp. and Methyloferula stellata (Chen et al., 2010; Dedysh et al., 2015). The pmoA gene has been shown to be a relevant group-specific biomarker as its phylogeny is congruent to tree topology based on the 16S rRNA gene and it has been widely used in molecular studies of methanotrophs (McDonald et al., 2008).
The presence of MOB in marine methane-rich sediments, such as active mud volcano (Niemann et al., 2006), gas hydrates (Yan et al., 2006), carbonate mounds (Marlow et al., 2014) and in bottom waters of methane vent and seeps (Tavormina et al., 2008, 2010) has been demonstrated, as well as their abundance and activity (Lösekann et al., 2007; Steinle et al., 2015). These studies have revealed extensive undocumented and diverse phylogenetic lineages of MOB belonging mainly to Gammaproteobacteria (type I), which are present in these ecosystems, although Alphaproteobacteria (type II) have also occasionally been reported in shallow estuary sediments (McDonald et al., 2005).
The Congo deep-sea fan represents a new marine CH4-rich environment, which originates from the recycling of rich terrigenous organic carbon deposits in turbidite sediments and harbors peculiar habitats as well as biogeochemical processes similar to active cold seeps systems. The aims of the present study were (1) to identify and to quantify potential MOB at the oxic–anoxic interface of chemosynthetic habitats in five sites in Congo lobe complexes, (2) to assess whether the MOB communities vary between habitats, over the distal-proximal transect and across various ages of lobe complexes, (3) to compare phylogenetic diversity of MOB in the Congo deep-sea turbidite with a characteristic cold seep, and (4) to identify potential chemical variables shaping the MOB diversity and distribution.
Materials and Methods
Study Sites and Samples
The Congolobe cruise (Rabouille, 2011) investigated the recent lobe complex at the distal of the present-day active channel-levee system (Figure 1). The recent lobe complex is thus an active system and is made of five partly stacked lobes that have a grape-like prograding downstream organization. Therefore, each lobe is characterized by a chronosequence of decreasing age (4 ka to present) in the upstream to downstream orientation and lobes were labeled 1–5 along this sequence (Figure 1).

FIGURE 1. Sampling sites and observed organic-rich habitats investigated in this study; adapted from (Rabouille et al., 2016). (A) Bathymetric map of the recent lobe complex of the Congo turbidite system with the location of the five sites studied during the Congolobe cruise. The recent lobe complex of the Congo deep sea fan covers an estimated area of 3,000 km2 and is composed of a series of lobes (labeled 1–5 from the oldest to the youngest) having a grape-like prograding downstream organization. (B–H) Sampling sites into chemosynthetic habitats observed using ROV Victor 6000 and core-tube sampler. Refer to Table 1 for label significance. (B) Reduced sediment at site A with vesicomyids clusters (Col_A_V). (C) Sparse vesicomyids at site B (Col_B_V). Ecosystems at site C : (D) Black patch of reduced sediment (Col_C_BP), (E) Microbial mat (Col_C_MM), (F) Vesicomyids clusters and adjacent brown sediment (Col_C_V). (G) Microbial mat at site F (Col_F_MM). (H) Brown sediment at site E (Col_E_BS).
The sites studied were chosen according to (i) a distal-proximal transect from the active channel on the recent lobe complex (sites A, B, F, C) and (ii) lobe complexes ages: the recent lobe complex (sites A, B, F, C) and the previous lobe complex (site E) aged between 6 and 4 ka BP (Picot et al., 2016) (Figure 1). Sediments were recovered within various types of dense chemosynthetic habitats at different sites between 4644 and 4947 m water-depth using the ROV Victor 6000 (Ifremer) core-tube sampler aboard the R/V Pourquoi Pas? (Figures 1B–H).
The description of the studied sites is summarized below (Table 1) and further information is provided in Rabouille et al. (2016):
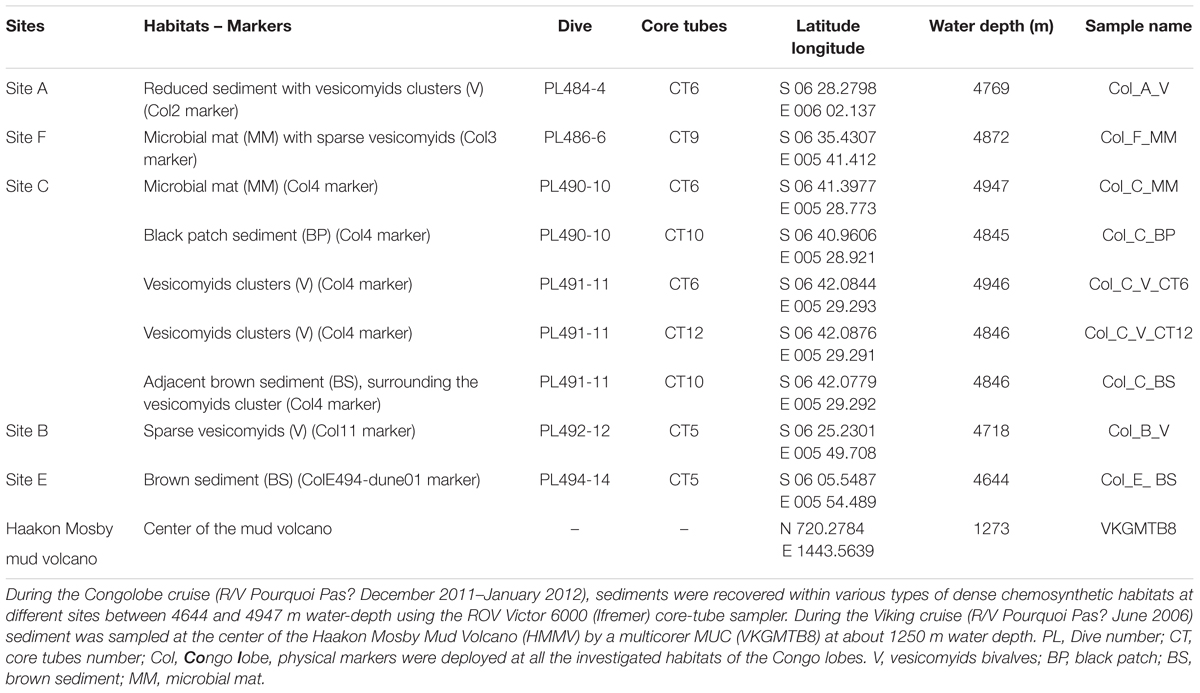
TABLE 1. Origin of the sites and samples investigated in this study.
• Site A is located at the entrance of the recent lobe complex, which is also the entrance of the oldest lobe (#1) and was colonized by vesicomyid bivalve patches edging black reduced sediments (Col_A_V).
Site B is located 22 km northwestward from site A in lobe (#3). It is abandoned from the active channel but still receives material from the turbiditic current overflow during important episodic turbiditic events (Bonnel, 2005). Black reduced sediments patches dominated by sparse vesicomyid bivalve’s shells and disarticulated valves were observed (Col_B_V).
• Site F is in the active-channel located 40 km downstream of site A, at the entrance of the youngest lobe (#5). It was characterized by thin grayish-whitish mats surrounding black reduced sediments (Col_F_MM).
• Site C is located 25 km from site F in the distal part of the youngest lobe (#5). It was characterized by the occurrence of diverse habitats: reduced sediments, defined by black patches with a few empty shells of vesicomyid bivalves (Col_C_BP), white microbial mat area surrounding black reduced sediment (Col_C_MM), dense vesicomyid bivalve’s colonies at the periphery of black reduced sediment patches (Col_C_V_CT6 and Col_C_V_CT12) characterized by dense populations of ampharetids and adjacent brown and bare sediments (Col_C_BS).
• Site E is located approximately 45 km to the north of site A and explored the feeding channel of the previous lobe complex, which is disconnected from the active channel and is interpreted as having been abandoned for several 1000 years. No seafloor biological manifestation was observed. One core-tube (Col_E_BS) sampled at this site, provides a reference for the onset to the hemipelagic sedimentation after the interruption of turbiditic deposition.
At each location, separate sediment core-tubes were collected for microbiological and geochemical analyses.
After retrieval, the cores were transferred to the cold room at 4°C and were sliced at intervals in 2-cm-thick layers. In this study, the first centimeters (0–2 cm) were analyzed and corresponded to the oxic–anoxic interface at shallow sediments. Sediment subsamples for nucleic acid extractions were frozen at -80°C. One cm3 of sediment subsample was fixed in PBS (phosphate-buffered saline)/formaldehyde (3% final), washed twice with PBS, and finally stored in PBS/Ethanol (1:1, vol:vol) at -20°C for fluorescence in situ hybridization (FISH) (Pernthaler et al., 2001).
In order to partially answer one of the questions, MOB communities in deep-sea turbidite system were compared with an additional sediment sample collected in 2006 from the active center of the HMMV, where freshly expelled fluid and mud support dense populations of MOB communities (Niemann et al., 2006; Lösekann et al., 2007). Briefly, this sediment was sampled by a multicorer MUC (VKGMTB8) at about 1250 m water depth on the slope of the Barents Sea continental margin (VICKING Cruise, R/V Pourquoi Pas? June 2006) (Nouze, 2006). The core was stored at 4°C and sliced at intervals in 2-cm-thick layers. In this study, similarly the first centimeters (0–2 cm), corresponding to the oxic–anoxic interface was analyzed and sediment subsamples were frozen at -80°C for nucleic acid extraction.
Preparation of Cell Staining and Fluorescence In Situ Hybridization
Cell staining and FISH reactions were performed for four selected samples at site C in the recent lobe complex (Col_C_MM, Col_C_BP, Col_C_V_CT6) and one reference sample at site E from the previous lobe complex (Col_E_BS).
Ten μL of a 100-fold dilution of fixed sediment subsamples, were gently mixed with a Vortex-Genie® 2 Mixers (Scientific Industries) to ensure that cells were in suspension and decanted and kept on ice for 10–15 min prior to transferring 40 μL of the supernatant into 8 mL of PBS. Samples were filtered through a 0.22 μm GTTP polycarbonate filter (Merck Millipore, Darmstadt, Germany) and air-dried.
In situ hybridization conditions were optimized to maximize efficiency and specificity of fluorescence signal. An optimal signal was obtained with 40% formamide. The specificity of the probes was further checked on a pure culture of Methylomarinum vadi strain IT-4T (Hirayama et al., 2013). Dual hybridizations with the Alexa-labeled EUB338 probe (16S rRNA; positions 338–355) for members of the domain Bacteria (Amann et al., 1990) and either a mix of Cy3-labeled 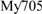 and
and 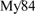 (16S rRNA; positions 84–101 and positions 705–722) probes or Cy3-labeled Mα450 (16S rRNA; positions 450–469) for the detection of Type I and Type II MOB cells, respectively (Eller et al., 2001) as well as Cy3-labeled MTMC-701 (16S rRNA; positions 701–718) for most members of the order Methylococcales (Boetius, 2000) were used (Supplementary Table 2).
(16S rRNA; positions 84–101 and positions 705–722) probes or Cy3-labeled Mα450 (16S rRNA; positions 450–469) for the detection of Type I and Type II MOB cells, respectively (Eller et al., 2001) as well as Cy3-labeled MTMC-701 (16S rRNA; positions 701–718) for most members of the order Methylococcales (Boetius, 2000) were used (Supplementary Table 2).
Filters were hybridized in a reaction mix containing 0.5 μM of each probe in a formamide hybridization buffer [0.9 M NaCl, 0.02 M Tris-HCl, 0.01% sodium dodecyl sulfate (SDS), 40% formamide] for 3 h at 46°C. Filters were then washed for 20 min at 48°C in a washing buffer [NaCl, 0.02 M Tris-HCl, 0.004 M EDTA, 0.01% SDS], rinsed briefly with sterile MilliQ water and air-dried (Pernthaler et al., 2001). Cells were counterstained with 10 μL of 4′, 6′-diamidino-2-phenylindole (DAPI; 1 μg/mL in MilliQ; 10 μL) for 10 min, briefly rinsed with sterile MilliQ water and left to air-dry. Finally, filters were fixed on microscopic slides and embedded in Citifluor AF1 (Citifluor, Ltd, UK) and stored at -20°C.
Procedure for Single Cells and Aggregated Cells
Single cells were counted after in situ hybridization by epifluorescence microscopy according to the method described by Goulder (1977). Triplicate filters for all samples were prepared for each dual hybridization conditions to minimize count variability and were subsequently counted in 100 independent microscopic fields. Blank samples were processed by treating 8 mL of 0.2 μm filtered PBS solution. Aggregate cells were counted as described by Lösekann et al. (2007). Observations, enumerations, and images were performed with a Zeiss Axio Imager 2 epifluorescence microscope (Carl Zeiss) equipped with the slider module ApoTome® (Zeiss) at 100X magnification, the illumination system HXP-X and Colibri light technology (Zeiss) with appropriate filter sets for Cy3, Alexa Fluor 488, GFP, and DAPI fluorescence using an AxioCam MRm camera (Zeiss). Epifluorescence acquisitions were analyzed using the ZEN Pro 2012 software (Zeiss).
DNA Extraction
Total nucleic acids were directly extracted from 3 to 5 g of wet sediments using the PowerMax® Soil DNA Isolation Kit (MO BIO Laboratories, Inc., Carlsbad, CA, USA) according to the manufacturers’ instructions. DNA was purified and concentrated by adding 0.2 mL of NaCl 5 M (SIGMA, St. Louis, MO, USA) and 10.4 mL of cold absolute ethanol (CARLOS EBRA, Val de Reuil, France). Samples were incubated 10 min at room temperature following by 5 min on ice, and then centrifuged for 30 min at 2,500 g at room temperature. The DNA pellets were washed with 5 mL of 70% ethanol and left to air-dry. The pellets were suspended in 200 μL of sterilized MilliQ water overnight at 4°C. Purified DNA extracts were stored at -20°C.
PCR, Clone Libraries, and Sequencing
Bacterial 16S rRNA genes were amplified by polymerase chain reaction (PCR) using the primer set E8F (5′-AGAGTTTGATCATGGCTCAG-3′) and U907R (5′-CCGTCAATTCMTTTRAGTTT-3′) (Lane et al., 1985). PmoA genes were amplified using the primer set A189f (5′-GGNGACTGGGACTTCTGG-3′) and mb661r (5′-CCGGMGCAACGT CYTTACC-3′) (Holmes et al., 1995; Costello and Lidstrom, 1999). These primers provide the widest coverage of the known pmoA diversity (McDonald et al., 2008) and are known to discriminate against sequences of the amoA genes (Dumont and Murrell, 2005); a pmoA homolog encoding the α-subunit of ammonia mono-oxygenase.
Polymerase chain reactions were performed in independent triplicates and PCR master mix consisted of 1X Green GoTaq®Flexi Buffer (Promega, Madison, WI, USA), 2 mM MgCl2 (Promega, Madison, WI, USA), 0.4 mM dNTP mix, 0.2 μM of each primer (Eurogentec, Liège, Belgique), 0.6 U of GoTaq®Flexi DNA polymerase (Promega, San Luis Obispo, CA, USA) and 1 μL of purified DNA template and sterile water to a final volume of 25 μL.
The PCR protocol consisted of an initial 95°C denaturing step for 5 min, followed by 30 cycles of 95°C for 45 s, 56°C for 1 min and 72°C for 45 s for pmoA genes or 94°C for 1 min, 57°C for 1 min 30 and 72°C for 2 min for bacterial 16S rRNA genes, and then a final 72°C elongation step for 6 min. Absence of contaminations was checked by negative controls. All PCR reactions were carried out using a GeneAmp® PCR system 9700 (Applied Biosystems, Foster City, CA, USA) thermal cycler, and PCR products were visualized using gel electrophoresis.
Amplified 16S rRNA and pmoA gene fragments from pooled triplicate reactions were purified on TAE agarose gels (0.8%) using NucleoSpin® Gel and PCR® Clean-up kits (Macherey Nagel, Düren, Germany) according to the manufacturer’s instructions. The purified amplicons were ligated into pCR2.1-TOPO vector (Invitrogen, Carlsbad, CA, USA) and transformed into Escherichia coli X-Gold® chemically competent cells (Stratagene) according to the manufacturer’s recommendations. Cloned pmoA fragments were sequenced using M13 universal primer with BigDye terminator chemistry and determined on an ABI3730xl Genetic Analyser (Applied Biosystems) (GATC Biotech, Konstanz, Germany).
The quality of retrieved 16S rRNA and pmoA sequences chromatograms and base calls were manually verified using Geneious© R6 version 6.1.7 (Kearse et al., 2012) and the vector and priming sites were excluded. Sequences were also screened for the closest relatives using blastn against the NCBI NR database. A customized pmoA database was created from sequences available in the NCBI database. Sequences were imported into a seed alignment of the ARB software package (Ludwig et al., 2004) and pmoA sequences were translated into amino acid sequences.
Assignment of OTUs, Diversity Metrics, and Statistics
Lower triangular distance matrices were calculated based on a SAI filter including 150 deduced amino acid positions using the neighbor-joining method (Saitou and Nei, 1987) and corrected with the PAM model (Wilbur, 1985) in ARB. The pmoA matrices were exported to Mothur (version 1.36.0) (Schloss et al., 2009) and sequences were clustered into operational taxonomic units (OTUs) using the average linkage algorithm with a cutoff distance of 7%. The PmoA distance of 7% was demonstrated to correspond to the 3% 16S rRNA distance, thus representing an indication for a representative methanotroph species (Lüke and Frenzel, 2011). Representative sequences of calculated OTUs (OTU-rep) were used for further in-depth phylogenetic analyses. OTU-rep is a sequence that had the minimum distance to the other closest sequences of the same OTU as calculated in Mothur (Schloss et al., 2009). Richness (Chao), Shannon -alpha diversity indices, as well as rarefaction curves, were calculated using Mothur (Schloss et al., 2009).
Analysis of variance (ANOVA) was performed with PAST software (Hammer et al., 2001) to test for significant contributions of MOB abundances between habitats.
To investigate patterns in the MOB community structure of the Congo lobe sediments and the link with selected chemical parameters (Supplementary Data and Table 1), we performed a canonical correspondence analysis (CCA) to determine parameters that could best explain the MOB community structure. The significance of the models was tested by analysis of variance with 1000 permutations. Co-inertia analysis (CoIA) was performed as well to identify trends or co-relationships in multiple datasets. A Monte Carlo permutation test (999 times) was performed to assess the significance of the co-structure of the data tables. Chemical data (CH4 concentration and efflux, diffusive oxygen uptake, oxygen penetration depth) were standardized to zero mean and unit variance before analysis. CoIA and CCA were done with the Ade4 and vegan packages (Thioulouse et al., 1997; Oksanen et al., 2013).
Phylogenetic Analysis
The 16S rRNA nucleic acid and PmoA amino acid gene sequences were aligned with Clustal W (Thompson et al., 1994) implemented in ARB and the alignments were refined manually. Phylogenetic trees were generated from an observed distance matrix using the neighbor-joining method and the partial adjustment model (PAM) correction (Wilbur, 1985) using ARB. The robustness of inferred topologies was calculated by bootstrap analyses with 100 resamplings of trees. Phylogenetic analyses were used to assign taxonomic for all pmoA sequences and to plot phylogenetic trees of OTU-rep.
Bipartite Network
A bipartite network was constructed to investigate co-occurrence patterns of MOB between samples. The two categories of node coexisting in this network correspond to the OTUs (circles) and the samples (squares). The network is based on Jaccard’s distance at the OTU level, which means that links denote the OTU memberships presents within samples. We visualized and computed the eigenvector centrality (ranging from 0 to 1) of the network with Gephi (Bastian et al., 2009). This measure assesses how influential a node is in the network topology through the number of connections weighted by the degree of its neighbors (Bonacich, 2007).
Nucleotide Sequence Accession Numbers
The nucleotide sequence data reported in this paper were submitted to GenBank/EMBL/DDBJ sequence databases under accession numbers KY350874–KY351540.
Results
Relative Abundances of Bacteria and MOB in Sedimentary Habitats of the Congo Lobe Complexes
Relative bacterial abundances detected by FISH were higher in chemosynthetic habitats (ranged from 1.3 to 4.2 × 107 cell/ml) of the recent lobe complex than in adjacent sediment (Col_C_BS) of the same lobe complex (0.1 ± 0.0 × 107 cell/ml) or than in sediment from the previous lobe complex (Col_E_BS; 0.1 ± 0.0 × 107 cell/ml) (Table 2). Black patch sediment (Col_C_BP) contained the highest relative number of bacterial cells, which mainly occurred as aggregates (Table 2).
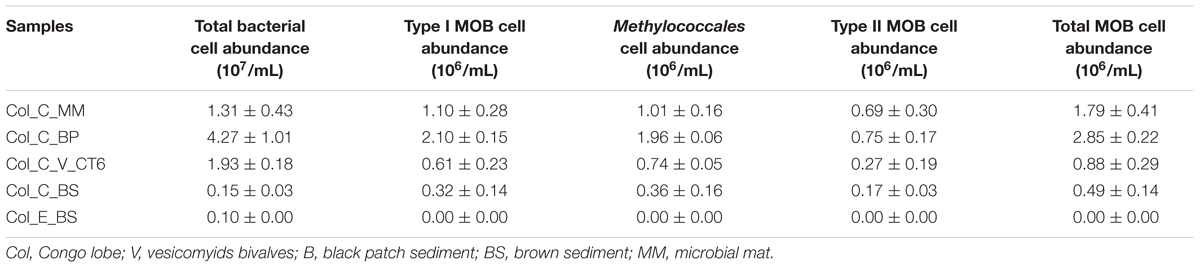
TABLE 2. Fluorescence in situ hybridization (FISH) quantification of bacteria and methane-oxidizing bacteria (MOB) at the oxic–anoxic interface of sedimentary habitats at site C from the recent lobe complex (active system) and at site E from the previous lobe complex (abandoned system) of the Congo deep-sea fan.
In sediments from the previous lobe complex (Col_E_BS), neither MOB cells were observed (Table 2) nor CH4 efflux detected (Supplementary Table 1). In contrast, MOB cells were detected in all sediments from habitats at site C from the recent lobe complex (Table 2). The highest relative number of MOB was observed in black patch sediment (Col_C_BP; 2.8 ± 0.2 × 106 cell/ml) while the lowest MOB abundance was observed in adjacent sample (Col_C_BS; 0.49 ± 0.1 × 106 cell/ml). The ratio between relative MOB and bacteria abundances ranged from 4 to 33% depending on the habitats. Type I abundances were higher in black patch sediment and microbial mat habitat while type II abundances were in the same order of magnitude in all samples (Table 2). The contribution of types I and II suggested a trend (one-way ANOVA; F = 3.9, p = 0.07) of higher abundances of type I cells (up to 2 × 106 cells mL-1, 61–74% of MOB mainly represented by members of the order Methylococcales) compared with type II (up to 0.7 × 106 cells mL-1, 26–38% of MOB).
Type II-MOB cells occurred as single/double cells or multicellular colonies and were rod- or coccoid-shaped of ∼0.5–2 μm in size and free-living (Figures 2K,L). In contrast, type I-MOB cells occurred as double cells or paired- multicellular colonies and differed in morphology as large coccoid- or pear-shaped cells of ∼1–2 μm diameter (Figures 2A–J).
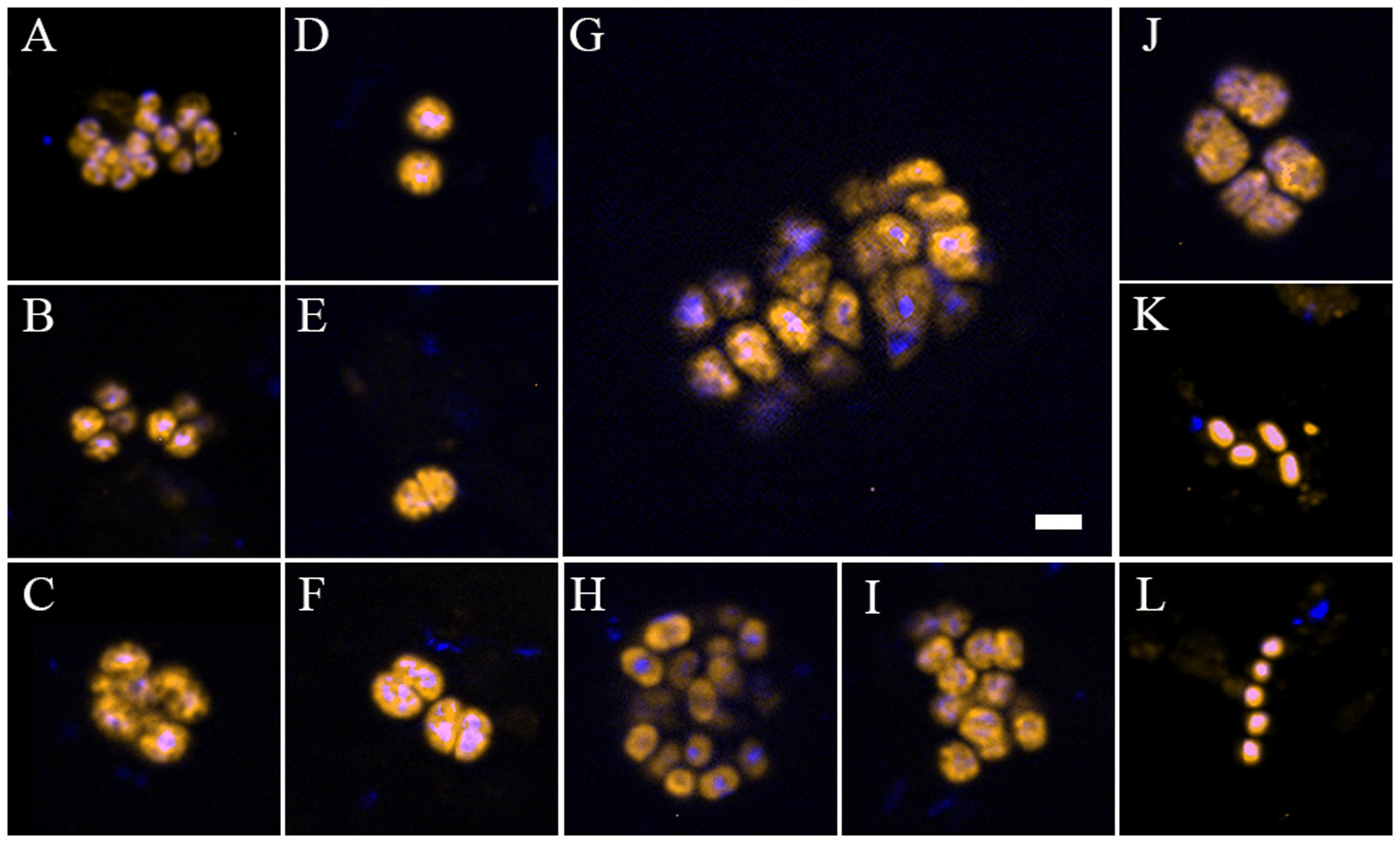
FIGURE 2. Individual cells and cell aggregates of methane-oxidizing bacteria (MOB) at the oxic–anoxic interface of organic-rich sedimentary habitats at the recent lobe complex (site C) of the Congo deep-sea fan, visualized with specific fluorescent-labeled oligonucleotides probes. (A–C) Clusters of type I MOB detected with the specific Cy3-labeled probe Mγ84/705 in parallel with DAPI staining. (D–J) Shape, spatial organization, and size of Methylococcales (Cy3-labeled probe MTMC-701) in parallel with DAPI staining. (K,L) Type II MOB cells hybridized with Cy3-labeled probe Mα450 in parallel with DAPI staining. Image A was taken from a sample collected at microbial mat habitats (Col_C_MM); (B,C,D–L) from black patch habitat (Col_C_BP). The scale bar represents 2 μm length.
Richness and Diversity of MOB in Congo Lobe Sedimentary Habitats
To get further insights into the diversity of MOB communities, we analyzed 16S rRNA and pmoA genes between sedimentary habitats of the recent lobe complexes (active system) and the previous lobe complex (abandoned system) (Figure 1). None of the presumed MOB representative sequences were detected in 16S rRNA gene clone libraries. However, genes encoding pmoA were detected by PCR in all DNA samples from the recent lobe complex, yielding a total of 667 accurate sequences that were classified into 28 OTUs (Table 3). In all attempts, no pmoA gene fragment could be amplified by PCR from DNA of the sediment from the previous lobe complex (Col_E_BS) (Figure 3), which is consistent with FISH observations (Table 2).
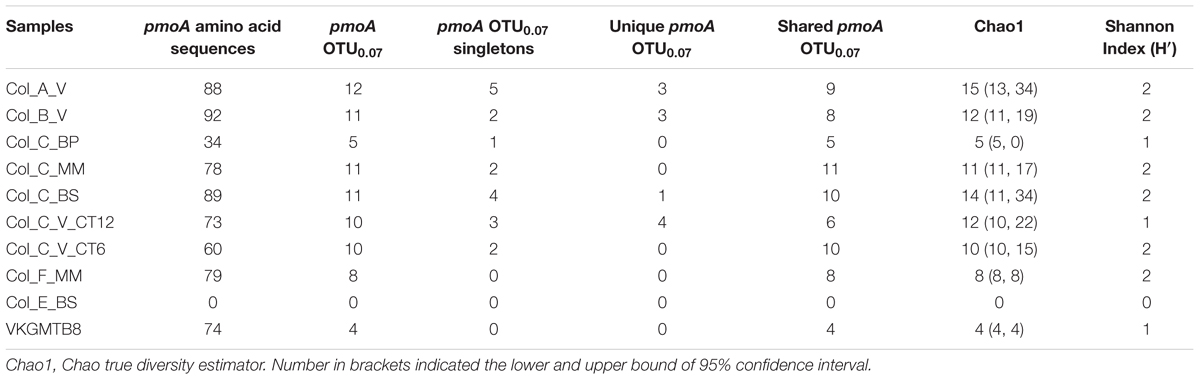
TABLE 3. Number of pmoA amino acid sequences, richness, diversity, and characteristics of operational taxonomic units (OTUs) of pmoA amino acid sequences at a cutoff value of 0.07.
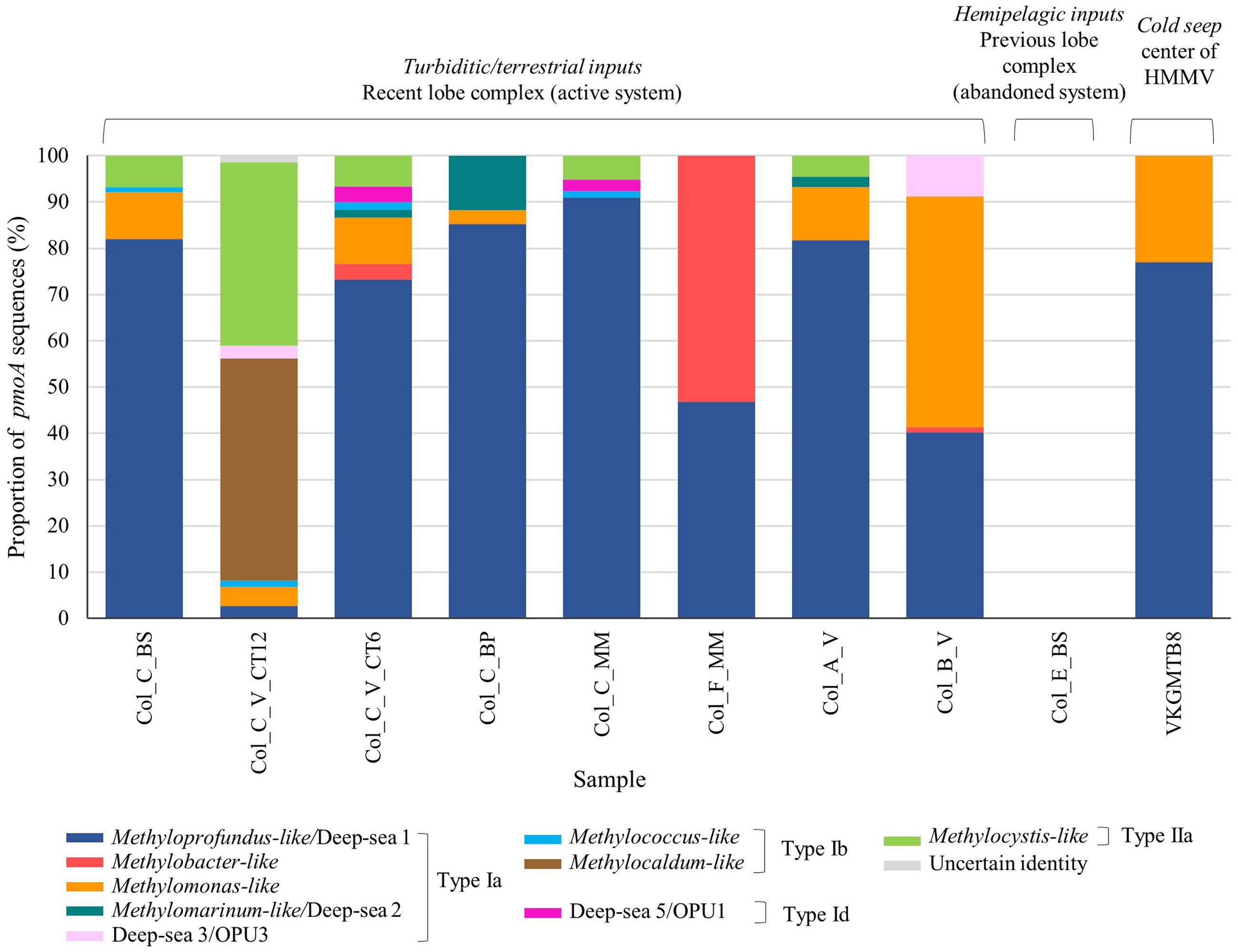
FIGURE 3. Phylogenetic affiliations and proportion of the pmoA amino acid sequences retrieved at surface organic-rich and seep sediments of habitats in the Congo lobe deep-sea fan (Col) and Haakon Mosby volcano (VKG) respectively based on neighbor-joining tree constructed using the ARB package and inferred with a PAM correction and 100 bootstrap replicates (158 curated amino acid positions).
Comparisons using the rarefaction curves showed that our sequencing effort did not cover the full pmoA diversity, with exceptions of the microbial mat habitat (Col_F_MM) and the black patch sediment (Col_C_BP) from the youngest lobe (#5), as well as the cold seep sample, HMMV (VKGMTB8) (Supplementary Figure 1). The richness estimates (Chao1) and rarefaction curves indicated that the highest richness was found in the vesicomyid habitat at site A (Col_A_V) (Table 3 and Supplementary Figure 1). In contrast, both analyses showed that the lowest richness was found in the black patch sediment of site C (Col_C_BP) and in the cold seep sediment, HMMV (VKGMTB8) (Table 3 and Supplementary Figure 1). These two samples with one of the two sediment samples from vesicomyids habitat at site C (Col_C_V_CT12) exhibited the lowest Shannon diversity index (H′) (Table 3).
Phylogenetic Diversity of MOB Communities in Congo Lobe Sedimentary Habitats
Phylogenetic analysis of the pmoA sequences revealed a great diversity of four subdivided phylotypes Ia, Ib, Id, and type IIa in the sedimentary habitats of the recent lobe complex (Figure 3). With the exception of one sediment sample of the vesicomyid habitat (Col_C_V_CT12), all sediments were composed of sequences affiliated to Methyloprofundus-like clade (type Ia), a marine strain of the deep-sea 1 clade (Tavormina et al., 2015). Other deep-sea clades (2, 3/OPU3, and 5/OPU1) were found in very low relative proportion (less than ∼10%) in some samples (Figure 3). In contrast, the sediment sample from the vesicomyid habitat (Col_C_V_CT12) was dominated by sequences affiliated to Methylocaldum-like (type Ib) and to Methylocystis-like (type IIa) clades. Interestingly, the microbial mat habitat at site F (Col_F_MM) and vesicomyid habitat at site B (Col_B_V) were respectively dominated by sequences closely related to Methylobacter-like and to Methylomonas-like clades respectively.
In comparison to sediments of the recent lobe complex from the Congo turbidite system, sediment of the typical cold seep HMMV exhibited lower pmoA diversity and was represented only by two clades of type Ia: Methyloprofundus-like and Methylomonas-like (Figure 3).
Phylogenetic analysis indicated that OTUs were similar to pmoA sequences retrieved from both marine and terrestrial environments (Figures 4, 5). The majority of the OTUs affiliated to type Ia fell into environmental clusters composed only of pmoA marine sequences isolated from methane seeps (Reed et al., 2009; Redmond et al., 2010; Kessler et al., 2011). In particular, most of the abundant OTUs were highly similar (90–100%) and closely affiliated to pmoA sequences recovered from a New Zealand cold seep at Hikurangi margin (Ruff et al., 2013) and to the cultivated genus Methyloprofundus sedimenti WF1T (Figure 4). Within type Ib (Figure 4), sequences showed a close relationship to strain Methylogaea oryzae E10T and grouped within a second environmental cluster of terrestrial sequences collected from soil and freshwater lake sediments (Ho et al., 2011a; Chauhan et al., 2012; Sow et al., 2014). Unique pmoA sequence affiliated to type Id (Figure 4) was related to a pmoA sequence retrieved from methane seeps from the North American margin and grouped within the deep-sea 5/OPU1 clade, supported with strong bootstrap values (Tavormina et al., 2008, 2010). Finally, within type IIa, five OTUs were closely related to the Methylocystis-like clade and pmoA sequences collected from rice paddies (Qiu et al., 2008; Bao et al., 2014) (Figure 5).
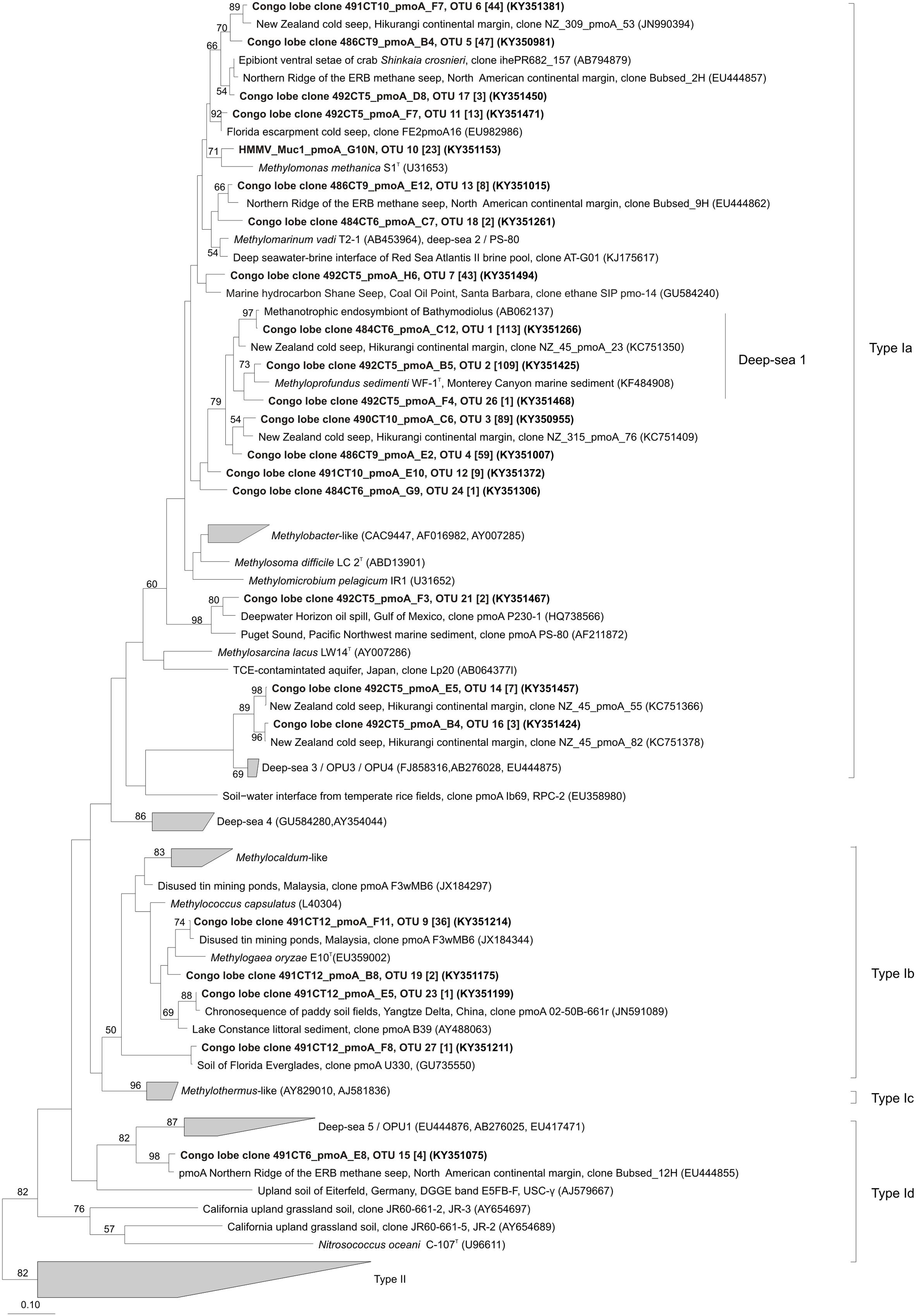
FIGURE 4. Phylogenetic analysis of the inferred amino acid sequences encoded by the pmoA gene of type I methane-oxidizing bacteria. The neighbor-joining tree was constructed using the ARB package and inferred with a PAM correction and 100 bootstrap replicates (158 curated amino acid positions). Bootstrap values higher than 50% are indicated. Scale bar represents 10% estimated substitutions per amino acid position. Representative sequences for each OTUs from this study are depicted in black bold and the corresponding number of sequences in OTUs are indicated in brackets.

FIGURE 5. Phylogenetic analysis of the inferred amino acid sequences encoded by the pmoA gene of type II methane-oxidizing bacteria. The neighbor-joining tree was constructed using the ARB package and inferred with a PAM correction and 100 bootstrap replicates (158 curated amino acid positions). Bootstrap values higher than 50% are indicated. Scale bar represents 10% estimated substitutions per amino acid position. Representative sequences for each OTUs from this study are depicted in black bold and the corresponding number of sequences in OTUs are indicated in brackets.
Co-occurrence of MOB Communities and Factors Shaping Their Structure
To study the associations between OTUs based on pmoA genes and their habitats, a network was constructed by computing the eigenvector centrality (Figure 6). Overall, none of the OTUs occurred at all sites, revealing heterogeneity in their distribution among the habitats. The center of the network is composed of a group of six influential OTUs: OTU 1, OTU 2, OTU 3, OTU 4, OTU 6, OTU 8 (having eigenvector centrality values higher than 0.5). All these taxa belong to type Ia (87.5%) except one of type IIa (OTU 8; 12.5%) (Figure 6), and were shared differentially between all samples as illustrated by the short paths. Among the major type Ia, the most abundant OTUs were OTU 1 (21% of pmoA sequences), OTU 2 (20% of pmoA sequences), OTU 3 (16% of pmoA sequences) and they were related to the deep-sea 1 clade (Lüke and Frenzel, 2011) (Figures 4, 6).
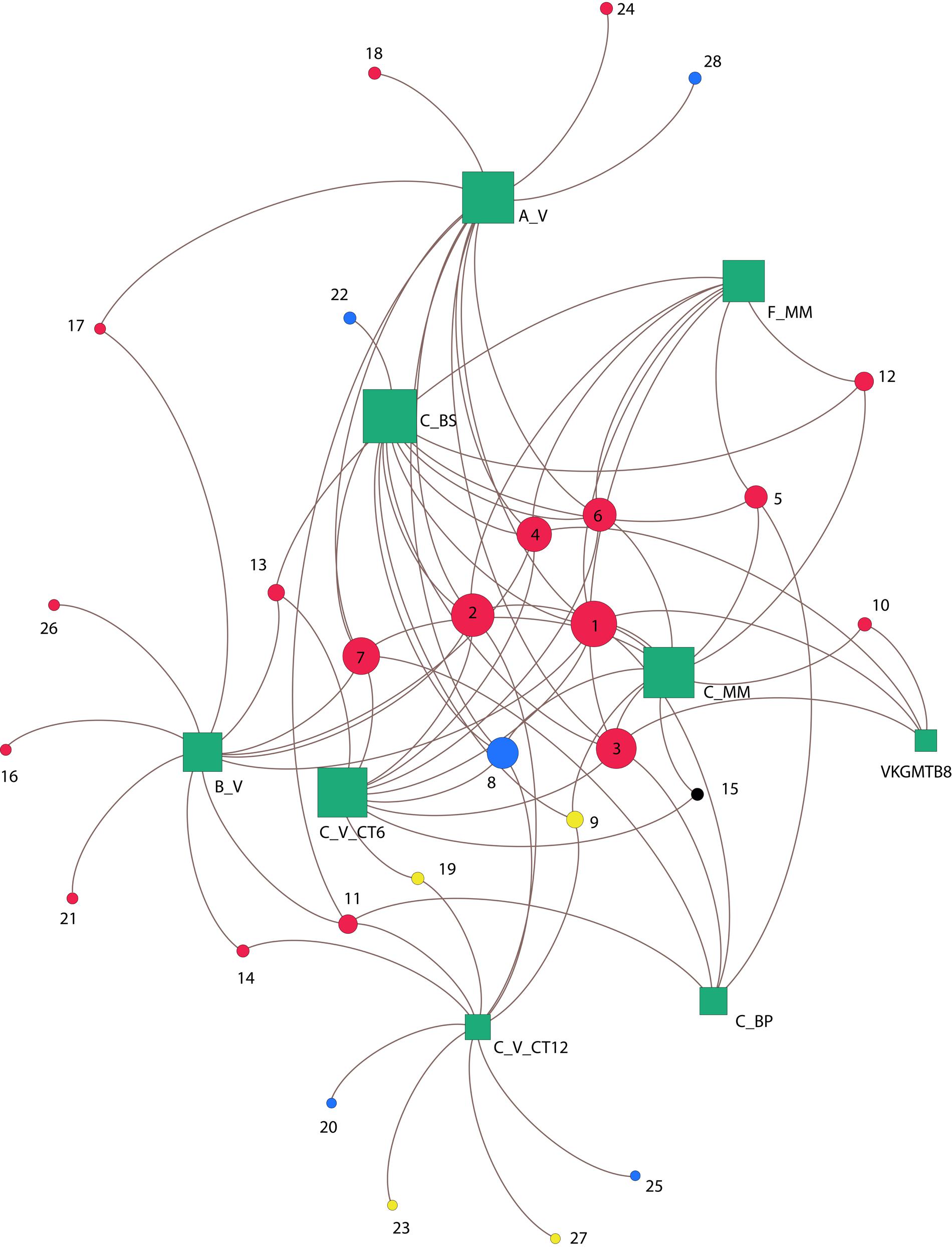
FIGURE 6. Co-occurrence of pmoA OTUs between sites. Green squares represent sedimentary habitats and circles correspond to OTUs memberships detected within habitats, based on the Betweenness-Centrality property. Node sizes are proportional to the number of connections. Colors circles indicate OTUs methane-oxidizing bacteria types (type Ia red; type Ib yellow; type Id black; and type IIa blue). Numbers indicates OTU names. Congo lobe habitats are labeled as follow: Col_C_MM (C_MM); Col_C_BP (C_BP); Col_C_V_CT6 (C_V_CT6); Col_C_V_CT12 (C_V_CT12); Col_C_BS (C_BS); Col_F_MM (F_MM); Col_A_V (A_V); Col_B_V (B_V). Haakon Mosby mud volcano is represented by sample VKGMTB8.
The sediments from the cold seep HMMV (VKGMTB8) shared distantly (long paths) three of the most abundant type Ia OTUs (OTU 1, OTU 3, OTU 4) with samples of the recent lobe complex from the Congo turbidite system.
Likewise, with exception of one sediment sample of the vesicomyid habitat (Col_C_V_CT6), all the vesicomyid habitats showed the occurrence of unique OTUs (39%) (Figure 6). These unique OTUs belonged to the type Ia (18%), type IIa (14%) and few were associated to type Ib (7%) as well. It should be noted that only one OTU of type Id was found (OTU 15) among all the samples. Nevertheless, this OTU occurred in two habitats of the site C (Col_C_MM and Col_C_V_CT6) (Figure 6).
We attempted to identify the relationships between chemical parameters and MOB communities of the Congo sedimentary habitats using CCA and CoIA analysis. Both CCA and CoIA indicated no significant correlation (p > 0.05) with the selected chemical parameters (CH4 efflux, oxygen uptake, oxygen penetration depth) (Supplementary Table 1) and the MOB community.
Discussion
Comparison of Relative MOB Abundances between Recent and Abandoned Congo Lobe Complex
Relative MOB abundances assessed by FISH indicated the presence of intact type I and type II cells for all site C samples (recent lobe complex) and they accounted for up to 32% of the bacterial cells. The recent lobe complex is receiving turbiditic inputs and fresh terrestrial OM in the lobe sediments (Baudin et al., 2017b). Moreover, OM mineralization in the recent lobe sediments produces considerable CH4 efflux (Supplementary Table 1) (Khripounoff et al., 2015), comparable to cold seeps (Boetius and Wenzhofer, 2013), which could support MOB communities. In contrast, MOB cells were not detected in site E sediment (previous lobe complex), either with FISH or pmoA amplification attempts. Formerly connected to the present-day active channel, the previous lobe complex (site E) was disconnected after the avulsion of the active channel to the south that is currently feeding the recent lobe complex. It is believed that the previous lobe complex has not been affected by turbiditic sediment events for several 1000 years and is now fed with hemi-pelagic sedimentation mostly dominated by strongly oxidized marine OM with residual overflow of turbiditic inputs (Baudin et al., 2017b). The absence of recent turbiditic deposition might explain why CH4-depleted sediments are observed at site E (Supplementary Table 1) and thus, the absence of MOB cells in the previous lobe complex.
In all sedimentary habitats from the recent lobe complex, type I MOB cells were always around two- to three-fold more abundant than type II MOB cells, which is in agreement with previous studies reporting that type I cells are largely dominant in methane seeps (Niemann et al., 2006; Lösekann et al., 2007; Steinle et al., 2015). Moreover, the abundance of type I and type II MOB cells in Congo lobe sediments were two orders of magnitude higher than reported from the water column above methane seeps at the Svalbard continental margin (Steinle et al., 2015) and two orders of magnitude less abundant than those found in surface sediment at the center of HMMV, which is characterized as a very active seep poor in OM (Lösekann et al., 2007). This suggests that Congo deep-sea fan harbored relatively high abundance of MOB compared to cold seeps.
Among the habitats at site C (recent lobe complex), black patch sediments hosted the highest number of MOB cells (Table 2). MOB cells may be stimulated by high CH4 efflux (Supplementary Table 1) and by high TOC contents (>4.5%), characteristic of black patch habitats (Baudin et al., 2017b). Black patches were widespread in the recent lobe complex (Rabouille et al., 2016) and they likely correspond to areas where CH4 escapes from the sediment due to rapid diagenesis of OM. The black uniform color of the recovered sediments indicates an O2 depletion due to CH4 oxidation (Baudin et al., 2017b). In contrast, a lower abundance of type I cells and smaller aggregates formation in vesicomyid habitats (Col_C_V_CT6) might be due to high grazing pressure by feeding heterotrophic ampharetid polychaetes as previously shown in Hikurangi Margin seep sediments (Thurber et al., 2013).
MOB Community Composition and Structure in the Congo Recent Lobe Complex
The representative sequences of MOB were not detected in the bacterial 16S rRNA gene libraries, indicating that the low sequencing effort (n = 100) using the cloning and Sanger sequencing based-method might not cover all the bacterial diversity. Previous studies were also unable to detect 16S rRNA gene sequences related to MOB with clone libraries but succeeded using high-throughput sequencing and pmoA functional gene clone libraries (Marlow et al., 2014).
Similarly, pmoA clone libraries showed that sedimentary habitats of the recent lobe complex hosted a great diversity of MOB lineages. However, general primer pairs used may not cover all the diversity of pmoA genes present in the Congo sediments. The design of revised general and new group specific primer pairs would improve the detection of pmoA genes, even amplify novel phylotypes. Most of the samples were dominated by type Ia sequences closely affiliated to the deep-sea 1 clade represented by the only cultivated member Methyloprofundus sedimenti WF1T. This obligate methanotroph was recently isolated from surface sediments (0–1 cm) in Monterey submarine canyon off the coast of California (1828 m below sea-level) close to a whale fall lying 500 m from a methane cold seep and 23 km from shore (Tavormina et al., 2015). This is in agreement with studies on sediments of hydrocarbon seeps on the continental margin of California (Tavormina et al., 2008), in the Gulf of Mexico (Yan et al., 2006) and off the coast of Japan (Inagaki et al., 2004) that have also revealed the dominance of diverse and novel type I methanotrophs.
Microbial mat habitat at site F was dominated by sequences related to the Methylobacter-like clade. This habitat is characterized by the highest CH4 efflux (208.8 mmol m-2 d-1) measured and relatively high diffusive oxygen-uptake (9.9 mmol m-2 d-1) concomitant to low oxygen penetration (4 mm) (Supplementary Table 1). Several studies have shown that Methylobacter spp. are particularly active at low O2/high CH4 ratios at the oxic–anoxic interface of soils and freshwater sediments (Reim et al., 2012; Hernandez et al., 2015).
Interestingly, vesicomyid habitats harbored unique and the most diversified OTUs, and their sediment-water interfaces were colonized by diverse polychaetes (Rabouille et al., 2016). These bottom dwellers create burrows networks that provide conduits for the transport of O2 deeply into the anoxic zone of the sediment, stimulating the recycling of OM (Aller and Aller, 1998). Hence ventilated animal burrows and bioturbation by vesicomyid bivalves may help to prevent local O2 depletion and may provide diverse CH4 or O2 micro-niches for diverse MOB lineages. Furthermore, vesicomyid habitats exhibited most of the type IIa sequences observed, which were related exclusively to the Methylocystis-like clades. Type II MOB are prevalent in terrestrial environments (Knief, 2015) and often persist in inactive and robust resting stages under environmental stress (Whittenbury et al., 1970) that may provide a potential seed bank of high diversity (Ho et al., 2011b). Some pmoA sequences were closely related to Methylogaea oryzae E10T (type Ib), a mesophilic methanotroph isolated from a rice paddy field growing on CH4 and methanol as sole carbon and energy sources (Geymonat et al., 2011). It has been shown that in a 2000 year-old paddy field, some species of type Ib and type II MOB emerged from the potential seed bank to become active when CH4 and O2 were available (Ho et al., 2011a). These findings suggest that some MOB might have been transported by alluvial sediment to the recent lobe complex where most turbidity currents die and lose the material held in suspension by turbulence (Khripounoff et al., 2003).
Comparison of MOB Communities between Turbiditic and Seep Sediments
Molecular identification of pmoA sequences showed that turbiditic sediments of the recent Congo lobe complex harbored a greater diversity and richness of MOB than seep sediment from the center of HMMV (Figures 4, 6). HMMV exhibited only two clades of type Ia closely related to the marine genera Methylomonas methanica S1T (Whittenbury and Krieg, 1984) and Methyloprofundus sedimenti WF1T (Tavormina et al., 2015) without type II members. Variations in diversity could be explained by the difference in the methane sources as previously reported. The central area of HMMV is connected to a deep gas reservoir and is thus impacted by high velocities of ascending methane-rich fluid flows, which episodically (1 or 2 per year) expels deep sediments enriched in CH4 and poor in OM (Niemann et al., 2006). As a result, the high upward flow increases CH4 concentrations in the uppermost oxic sediment horizon stimulating the growth of MOB and preventing the diffusion of sulfate into the sediment, hence inhibiting sulfate dependent anaerobic CH4 oxidation (Niemann et al., 2006). In contrast, the presence of deep reservoirs in the recent Congo lobe complex has not been demonstrated and the fluvial source is the main known contributor of the OM and CH4 efflux in the recent Congo lobe complex (Baudin et al., 2017b). This region is affected by strong episodic turbidity events (∼2 years, Heezen et al., 1964) with high terrestrial organic carbon content (3–5%) (Stetten et al., 2015) that creates disturbances and OM-rich areas (Rabouille et al., 2016; Baudin et al., 2017a). Localized intense remineralization of the OM during early diagenesis produces high CH4 efflux that may provide broad ecological niches and the potential for higher diversity of MOB communities. The high similarity between MOB lineages found at turbiditic sediments and cold seeps could support the concept of stepping stones (Smith et al., 1989) existing between diverse deep-sea ecosystems including deep-sea fans.
Turbiditic sediments from the recent Congo lobe complex harbored diverse MOB lineages within the type I-a, -b, and -d and type II-a subgroups that were affiliated mainly with pmoA sequences from marine environments (unclassified MOB sequences from methane seeps; deep-sea clades 1–3, 5) but also with pmoA sequences from aquatic (freshwater) and terrestrial (soils, rice paddies) environments. Elemental and isotopic analysis have shown that the nature of OM in the recent lobe complex is dominated (>80%) by a terrestrial source containing a mixture of higher plant debris and soil derived OM and that signatures of sediments from the recent lobe complex and from the Congo River were similar (Stetten et al., 2015; Baudin et al., 2017b). Furthermore, uncommon elevated values of a specific bacteriohopanepolyols (BHPs) produced by type I MOB were reported down to 115 mbsf in marine sediments located on the lower Congo fan (ODP Site 1075) (Talbot et al., 2014). It was further shown that the source of BHPs were tropical wetland systems and soils in the Congo river catchment (Spencer-Jones et al., 2015). The presence of these compounds have also been observed in high relative abundance in marine sediments of the Amazon shelf and deep-sea fan (Wagner et al., 2014). These observations support that turbiditic deposits of the Congo deep-sea fan host MOB communities of possible terrestrial origin.
Conclusion
Both pmoA and relative cell abundances surveys indicate the presence of typical marine and terrestrial MOB phylogenetic lineages within the oxic–anoxic interface sediments of diverse organic-rich habitats in the recent Congo lobe complex. The predominant terrigenous origin of the OM pool and the detection of typical terrestrial MOB phylogenetic lineages at deep water depth (5000 m) is one of the most surprising results of this study. The comparison with collection of sediments from the Congo River and canyon as well as other fan systems would provide additional information regarding the origin of terrestrial guilds in the marine environment. Moreover, the assessment of active communities using microcosms enrichments, methane oxidation rates and metagenomic-isotope labeling experiments could widen our understanding concerning the origin and role of MOB in the functioning of the peculiar ecosystems of the Congo deep-sea fan.
Author Contributions
SB designed experiments, acquired, analyzed and interpreted data, wrote manuscript. YM analyzed and interpreted data, revised manuscript. SG collected and analyzed sequence data. FL collected samples and sequence data, revised manuscript. AG designed experiments, collected samples and revised manuscript. LT designed experiments, analyzed data, revised manuscript and supervised SB project’s.
Funding
This study was funded by the French National Research Agency project, ANR Congolobe leaded by C. Rabouille (ANR-11-BS56-0030, 2011–2016: organic carbon transfer and ecosystem functioning in the terminal lobes of the Congo deep-sea fan). SB was supported by a doctoral fellowship from the French Research Institute for Exploitation of the Sea (Ifremer).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer SER and handling Editor declared their shared affiliation, and the handling Editor states that the process nevertheless met the standards of a fair and objective review.
Acknowledgments
We express our gratitude to Captain G. Ferrand, the chief scientist C. Rabouille and crew for the sampling campaign Congolobe onboard the R/V Pourquoi Pas? We acknowledge P. Cruaud for sample collection and conditioning for microbial studies during the Congolobe campaign. We thank H. Hirayama for kindly providing a cultivation of Methylomarinum vadi strain IT-4T for FISH optimization. We thank V. Cueff-Gauchard and M. A. Cambon-Bonavita and E. Roussel for advices regarding FISH analyses and cell counting methods. We acknowledge L. Pastor for the revision and M. G. Dumont for proofreading of the manuscript, and thank K. Olu for providing insight about vesicomyids. We thank A. Khripounoff, J.-C. Caprais, J. Le Bruchec, L. Pozzato, and C. Rabouille for providing chemical data.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2017.00715/full#supplementary-material
References
Aller, R. C., and Aller, J. Y. (1998). The effect of biogenic irrigation intensity and solute exchange on diagenetic reaction rates in marine sediments. J. Mar. Res. 56, 905–936. doi: 10.1357/002224098321667413
Amann, R. I., Krumholz, L., and Stahl, D. A. (1990). Fluorescent-oligonucleotide probing of whole cells for determinative phylogenetic and environmental studies in microbiology. J. Bacteriol. 172, 762–770. doi: 10.1128/jb.172.2.762-770.1990
Bao, Z., Watanabe, A., Sasaki, K., Okubo, T., Tokida, T., Liu, D., et al. (2014). A rice gene for microbial symbiosis, Oryza sativa CCaMK, reduces CH4 flux in a paddy field with low nitrogen input. Appl. Environ. Microbiol. 80, 1995–2003. doi: 10.1128/aem.03646-13
Bastian, M., Heymann, S., and Jacomy, M. (2009). “Gephi: an open source software for exploring and manipulating networks,” in Proceedings of the Third International AAAI Conference on Weblogs and Social Media, San Jose, CA.
Baudin, F., Martinez, P., Dennielou, B., Charlier, K., Marsset, T., Droz, L., et al. (2017a). Organic carbon accumulation in modern sediments of the Angola basin influenced by the Congo deep sea fan. Deep Sea Res. Part II Top. Stud. Oceanogr. (in press). doi: 10.1016/j.dsr2.2017.01.009
Baudin, F., Stetten, E., Schnyder, J., Charlier, K., Martinez, P., Dennielou, B., et al. (2017b). Origin and distribution of the organic matter in the distal lobe of the Congo deep-sea fan–A Rock-Eval survey. Deep Sea Res. Part II Top. Stud. Oceanogr. (in press). doi: 10.1016/j.dsr2.2017.01.008
Boetius, A. (2000). A marine microbial consortium apparently mediating anaerobic oxidation of methane. Nature 407, 623–626. doi: 10.1038/35036572
Boetius, A., and Wenzhofer, F. (2013). Seafloor oxygen consumption fuelled by methane from cold seeps. Nat. Geosci. 6, 725–734. doi: 10.1038/ngeo1926
Bonacich, P. (2007). Some unique properties of eigenvector centrality. Soc. Netw. 29, 555–564. doi: 10.1016/j.socnet.2007.04.002
Bonnel, C., (2005). Mise en Place des Lobes Distaux dans les Systèmes Turbiditiques Actuels: Analyse Comparée des Systèmes du Zaïre, Var et Rhône. Thése de Doctorat, Université de Bordeaux I, Talence, 314.
Chauhan, A., Pathak, A., and Ogram, A. (2012). Composition of methane-oxidizing bacterial communities as a function of nutrient loading in the florida everglades. Microb. Ecol. 64, 750–759. doi: 10.1007/s00248-012-0058-2
Chen, Y., Crombie, A., Rahman, M. T., Dedysh, S. N., Liesack, W., Stott, M. B., et al. (2010). Complete genome sequence of the aerobic facultative methanotroph Methylocella silvestris BL2. J. Bacteriol. 192, 3840–3841. doi: 10.1128/jb.00506-10
Costello, A. M., and Lidstrom, M. E. (1999). Molecular characterization of functional and phylogenetic genes from natural populations of methanotrophs in lake sediments. Appl. Environ. Microbiol. 65, 5066–5074.
Dedysh, S. N., Naumoff, D. G., Vorobev, A. V., Kyrpides, N., Woyke, T., Shapiro, N., et al. (2015). Draft genome sequence of Methyloferula stellata AR4, an obligate methanotroph possessing only a soluble methane monooxygenase. Genome Announc. 3:e01555–14. doi: 10.1128/genomeA.01555-14
Droz, L., Marsset, T., Ondréas, H., Lopez, M., Savoye, B., and Spy-Anderson, F.-L. (2003). Architecture of an active mud-rich turbidite system: the zaire fan (congo–angola margin southeast atlantic):Results from ZaïAngo 1 and 2 cruises. Am. Assoc. Pet. Geol. Bull. 87, 1145–1168. doi: 10.1306/03070300013
Dumont, M. G., and Murrell, J. C. (2005). “Community-level analysis: key genes of aerobic methane oxidation,” in Methods in Enzymology, eds J. Abelson, M. Simon, G. Verdine, and A. Pyle (Cambridge, MA: Academic Press), 413–427.
Eller, G., Stubner, S., and Frenzel, P. (2001). Group-specific 16S rRNA targeted probes for the detection of type I and type II methanotrophs by fluorescence in situ hybridisation. FEMS Microbiol. Lett. 198, 91–97. doi: 10.1111/j.1574-6968.2001.tb10624.x
Geymonat, E., Ferrando, L., and Tarlera, S. E. (2011). Methylogaea oryzae gen. nov., sp. nov., a mesophilic methanotroph isolated from a rice paddy field. Int. J. Syst. Evol. Microbiol. 61, 2568–2572. doi: 10.1099/ijs.0.028274-0
Goulder, R. (1977). Attached and free bacteria in an estuary with abundant suspended solids. J. Appl. Bacteriol. 43, 399–405. doi: 10.1111/j.1365-2672.1977.tb00766.x
Hammer,Ø., Harper, D. A. T., and Ryan, P. D. (2001). PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 4, 1–9.
Heezen, B. C., Menzies, R. J., Schneider, E. D., Ewing, W. M., and Granelli, N. C. L. (1964). Congo submarine Canyon. Am. Assoc. Pet. Geol. Bull. 48, 1126–1149.
Hernandez, M. E., Beck, D. A. C., Lidstrom, M. E., and Chistoserdova, L. (2015). Oxygen availability is a major factor in determining the composition of microbial communities involved in methane oxidation. PeerJ 3:e801. doi: 10.7717/peerj.801
Hirayama, H., Fuse, H., Abe, M., Miyazaki, M., Nakamura, T., Nunoura, T., et al. (2013). Methylomarinum vadi gen. nov., sp. nov., a methanotroph isolated from two distinct marine environments. Int. J. Syst. Evol. Microbiol. 63, 1073–1082. doi: 10.1099/ijs.0.040568-0
Ho, A., Lüke, C., Cao, Z., and Frenzel, P. (2011a). Ageing well: methane oxidation and methane oxidizing bacteria along a chronosequence of 2000 years. Environ. Microbiol. Rep. 3, 738–743. doi: 10.1111/j.1758-2229.2011.00292.x
Ho, A., Luke, C., and Frenzel, P. (2011b). Recovery of methanotrophs from disturbance: population dynamics, evenness and functioning. ISME J. 5, 750–758. doi: 10.1038/ismej.2010.163
Holmes, A. J., Costello, A., Lidstrom, M. E., and Murrell, J. C. (1995). Evidence that participate methane monooxygenase and ammonia monooxygenase may be evolutionarily related. FEMS Microbiol. Lett. 132, 203–208. doi: 10.1111/j.1574-6968.1995.tb07834.x
Inagaki, F., Tsunogai, U., Suzuki, M., Kosaka, A., Machiyama, H., Takai, K., et al. (2004). Characterization of C1-metabolizing prokaryotic communities in methane seep habitats at the kuroshima knoll, southern ryukyu arc, by analyzing pmoA, mmoX, mxaF, mcrA, and 16S rRNA Genes. Appl. Environ. Microbiol. 70, 7445–7455. doi: 10.1128/aem.70.12.7445-7455.2004
Kearse, M., Moir, R., Wilson, A., Stones-Havas, S., Cheung, M., and Sturrock, S. (2012). Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28, 1647–1649. doi: 10.1093/bioinformatics/bts199
Kessler, J. D., Valentine, D. L., Redmond, M. C., Du, M., Chan, E. W., Mendes, S. D., et al. (2011). A persistent oxygen anomaly reveals the fate of spilled methane in the deep gulf of mexico. Science 331, 312–315. doi: 10.1126/science.1199697
Khripounoff, A., Caprais, J. C., Decker, C., Essirard, M., Le Bruchec, J., Noel, P., et al. (2015). Variability in gas and solute fluxes through deep-sea chemosynthetic ecosystems inhabited by vesicomyid bivalves in the Gulf of Guinea. Deep Sea Res. Part I Oceanogr. Res. Pap. 95, 122–130. doi: 10.1016/j.dsr.2014.10.013
Khripounoff, A., Vangriesheim, A., Babonneau, N., Crassous, P., Dennielou, B., and Savoye, B. (2003). Direct observation of intense turbidity current activity in the Zaire submarine valley at 4000 m water depth. Mar. Geol. 194, 151–158. doi: 10.1016/S0025-3227(02)00677-1
Knief, C. (2015). Diversity and habitat preferences of cultivated and uncultivated aerobic methanotrophic bacteria evaluated based on pmoA as molecular marker. Front. Microbiol. 6:1346. doi: 10.3389/fmicb.2015.01346
Lane, D. J., Pace, B., Olsen, G. J., Stahl, D. A., Sogin, M. L., and Pace, N. R. (1985). Rapid determination of 16S ribosomal RNA sequences for phylogenetic analyses. Proc. Natl. Acad. Sci. U.S.A. 82, 6955–6959. doi: 10.1073/pnas.82.20.6955
Lösekann, T., Knittel, K., Nadalig, T., Fuchs, B., Niemann, H., Boetius, A., et al. (2007). Diversity and abundance of aerobic and anaerobic methane oxidizers at the Haakon Mosby Mud Volcano, Barents Sea. Appl. Environ. Microbiol. 73, 3348–3362. doi: 10.1128/AEM.00016-07
Ludwig, W., Strunk, O., Westram, R., Richter, L., Meier, H., Yadhukumar, et al. (2004). ARB: a software environment for sequence data. Nucleic Acids Res. 32, 1363–1371. doi: 10.1093/nar/gkh293
Lüke, C., and Frenzel, P. (2011). Potential of pmoA amplicon pyrosequencing for methanotroph diversity studies. Appl. Environ. Microbiol. 77, 6305–6309. doi: 10.1128/AEM.05355-11
Marlow, J. J., Steele, J. A., Ziebis, W., Thurber, A. R., Levin, L. A., and Orphan, V. J. (2014). Carbonate-hosted methanotrophy represents an unrecognized methane sink in the deep sea. Nat. Commun. 5, 5094. doi: 10.1038/ncomms6094
McDonald, I. R., Bodrossy, L., Chen, Y., and Murrell, J. C. (2008). Molecular ecology techniques for the study of aerobic methanotrophs. Appl. Environ. Microbiol. 74, 1305–1315. doi: 10.1128/AEM.02233-07
McDonald, I. R., Smith, K., and Lidstrom, M. E. (2005). Methanotrophic populations in estuarine sediment from Newport Bay, California. FEMS Microbiol. Lett. 250, 287–293. doi: 10.1016/j.femsle.2005.07.016
Niemann, H., Lösekann, T., de Beer, D., Elvert, M., Nadalig, T., Knittel, K., et al. (2006). Novel microbial communities of the Haakon Mosby mud volcano and their role as a methane sink. Nature 443, 854–858. doi: 10.1038/nature05227
Oksanen, J., Blanchet, F., Kindt, R., Legendre, P., Minchin, P., O’Hara, R., et al. (2013). vegan: Community Ecology Package. 2013. R Package Version 2.0–7. Available at: http://CRAN.R-project.org/package=vegan
Pernthaler, J., Glöckner, F.-O., Schönhuber, W., and Amann, R. (2001). “Fluorescence in situ hybridization (FISH) with rRNA-targeted oligonucleotide probes,” in Methods in Microbiology, ed. J. Paul (London: Academic Press), 207–226.
Picot, M., Droz, L., Marsset, T., Dennielou, B., and Bez, M. (2016). Controls on turbidite sedimentation: insights from a quantitative approach of submarine channel and lobe architecture (Late Quaternary Congo Fan). Mar. Pet. Geol. 72, 423–446. doi: 10.1016/j.marpetgeo.2016.02.004
Qiu, Q., Noll, M., Abraham, W.-R., Lu, Y., and Conrad, R. (2008). Applying stable isotope probing of phospholipid fatty acids and rRNA in a Chinese rice field to study activity and composition of the methanotrophic bacterial communities in situ. ISME J. 2, 602–614. doi: 10.1038/ismej.2008.34
Rabouille, C., Caprais, J. C., Lansard, B., Crassous, P., Dedieu, K., Reyss, J. L., et al. (2009). Organic matter budget in the Southeast Atlantic continental margin close to the Congo Canyon: in situ measurements of sediment oxygen consumption. Deep Sea Res. Part II Top. Stud. Oceanogr. 56, 2223–2238. doi: 10.1016/j.dsr2.2009.04.005
Rabouille, C., Olu, K., Baudin, F., Khripounoff, A., Dennielou, B., Arnaud-Haond, S., et al. (2016). The Congolobe project, a multidisciplinary study of Congo deep-sea fan lobe complex: overview of methods, strategies, observations and sampling. Deep Sea Res. Part II Top. Stud. Oceanogr. doi: 10.1016/j.dsr2.2016.05.006
Redmond, M. C., Valentine, D. L., and Sessions, A. L. (2010). Identification of novel methane-, ethane-, and propane-oxidizing bacteria at marine hydrocarbon seeps by stable isotope probing. Appl. Environ. Microbiol. 76, 6412–6422. doi: 10.1128/AEM.00271-10
Reed, A. J., Dorn, R., Van Dover, C. L., Lutz, R. A., and Vetriani, C. (2009). Phylogenetic diversity of methanogenic, sulfate-reducing and methanotrophic prokaryotes from deep-sea hydrothermal vents and cold seeps. Deep Sea Res. Part II Top. Stud. Oceanogr. 56, 1665–1674. doi: 10.1016/j.dsr2.2009.05.012
Reim, A., Luke, C., Krause, S., Pratscher, J., and Frenzel, P. (2012). One millimetre makes the difference: high-resolution analysis of methane-oxidizing bacteria and their specific activity at the oxic-anoxic interface in a flooded paddy soil. ISME J. 6, 2128–2139. doi: 10.1038/ismej.2012.57
Ruff, S. E., Arnds, J., Knittel, K., Amann, R., Wegener, G., Ramette, A., et al. (2013). Microbial communities of deep-sea methane seeps at Hikurangi continental margin (New Zealand). PLoS ONE 8:e72627. doi: 10.1371/journal.pone.0072627
Saitou, N., and Nei, M. (1987). The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4, 406–425.
Savoye, B., Babonneau, N., Dennielou, B., and Bez, M. (2009). Geological overview of the Angola–Congo margin, the Congo deep-sea fan and its submarine valleys. Deep Sea Res. Part II Top. Stud. Oceanogr. 56, 2169–2182. doi: 10.1016/j.dsr2.2009.04.001
Schloss, P. D., Westcott, S. L., Ryabin, T., Hall, J. R., Hartmann, M., Hollister, E. B., et al. (2009). Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 75, 7537–7541. doi: 10.1128/AEM.01541-09
Shanmugam, G., and Moiola, R. J. (1988). Submarine fans: characteristics, models, classification, and reservoir potential. Earth Sci. Rev. 24, 383–428. doi: 10.1016/0012-8252(88)90064-5
Smith, C. R., Kukert, H., Wheatcroft, R. A., Jumars, P. A., and Deming, J. W. (1989). Vent fauna on whale remains. Nature 341, 27–28. doi: 10.1038/341027a0
Sow, S. L. S., Khoo, G., Chong, L. K., Smith, T. J., Harrison, P. L., and Ong, H. K. A. (2014). Molecular diversity of the methanotrophic bacteria communities associated with disused tin-mining ponds in Kampar, Perak, Malaysia. World J. Microbiol. Biotechnol. 30, 2645–2653. doi: 10.1007/s11274-014-1687-z
Spencer-Jones, C. L., Wagner, T., Dinga, B. J., Schefuß, E., Mann, P. J., Poulsen, J. R., et al. (2015). Bacteriohopanepolyols in tropical soils and sediments from the Congo River catchment area. Org. Geochem. 8, 1–13. doi: 10.1016/j.orggeochem.2015.09.003
Steinle, L., Graves, C. A., Treude, T., Ferre, B., Biastoch, A., Bussmann, I., et al. (2015). Water column methanotrophy controlled by a rapid oceanographic switch. Nat. Geosci. 8, 378–382. doi: 10.1038/ngeo2420
Stetten, E., Baudin, F., Reyss, J.-L., Martinez, P., Charlier, K., Schnyder, J., et al. (2015). Organic matter characterization and distribution in sediments of the terminal lobes of the Congo deep-sea fan: evidence for the direct influence of the Congo River. Mar. Geol. 369, 182–195. doi: 10.1016/j.margeo.2015.08.020
Talbot, H. M., Handley, L., Spencer-Jones, C. L., Dinga, B. J., Schefuß, E., Mann, P. J., et al. (2014). Variability in aerobic methane oxidation over the past 1.2 Myrs recorded in microbial biomarker signatures from Congo fan sediments. Geochim. Cosmochim. Acta 133, 387–401. doi: 10.1016/j.gca.2014.02.035
Tavormina, P. L., Hatzenpichler, R., McGlynn, S., Chadwick, G., Dawson, K. S., Connon, S. A., et al. (2015). Methyloprofundus sedimenti gen. nov., sp. nov., an obligate methanotroph from ocean sediment belonging to the ‘deep sea-1’ clade of marine methanotrophs. Int. J. Syst. Evol. Microbiol. 65, 251–259. doi: 10.1099/ijs.0.062927-0
Tavormina, P. L., Ussler, W., Joye, S. B., Harrison, B. K., and Orphan, V. J. (2010). Distributions of putative aerobic methanotrophs in diverse pelagic marine environments. ISME J. 4, 700–710. doi: 10.1038/ismej.2009.155
Tavormina, P. L., Ussler, W., and Orphan, V. J. (2008). Planktonic and sediment-associated aerobic methanotrophs in two seep systems along the North American margin. Appl. Environ. Microbiol. 74, 3985–3995. doi: 10.1128/AEM.00069-08
Thioulouse, J., Chessel, D., and Champely, S. (1997). Multivariate analysis of spatial patterns: a unified approach to local and global structures. Environ. Ecol. Stat. 2, 1–14. doi: 10.1007/bf00452928
Thompson, J. D., Higgins, D. G., and Gibson, T. J. (1994). CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 22, 4673–4680. doi: 10.1093/nar/22.22.4673
Thurber, A. R., Levin, L. A., Rowden, A. A., Sommer, S., Linke, P., and Kröger, K. (2013). Microbes, macrofauna, and methane: a novel seep community fueled by aerobic methanotrophy. Limnol. Oceanogr. 58, 1640–1656. doi: 10.4319/lo.2013.58.5.1640
Wagner, T., Kallweit, W., Talbot, H. M., Mollenhauer, G., Boom, A., and Zabel, M. (2014). Microbial biomarkers support organic carbon transport from methane-rich Amazon wetlands to the shelf and deep sea fan during recent and glacial climate conditions. Org. Geochem. 67, 85–98. doi: 10.1016/j.orggeochem.2013.12.003
Whittenbury, R., Davies, S. L., and Davey, J. (1970). Exospores and cysts formed by methane-utilizing bacteria. Microbiology 61, 219–226. doi: 10.1099/00221287-61-2-219
Whittenbury, R., and Krieg, N. R. (1984). “Genus II. Methylomonas,” in Bergey’s Manual of Systematic Bacteriology, 1st Edn, Vol. 1, eds N. R. Krieg and J. G. Holt (Baltimore, MD: Williams & Wilkins Co), 260–261.
Keywords: aerobic methane-oxidizing bacteria, pmoA, organic-rich sediment, methane seeps, Congo deep-sea fan
Citation: Bessette S, Moalic Y, Gautey S, Lesongeur F, Godfroy A and Toffin L (2017) Relative Abundance and Diversity of Bacterial Methanotrophs at the Oxic–Anoxic Interface of the Congo Deep-Sea Fan. Front. Microbiol. 8:715. doi: 10.3389/fmicb.2017.00715
Received: 04 January 2017; Accepted: 06 April 2017;
Published: 25 April 2017.
Edited by:
Peter Dunfield, University of Calgary, CanadaReviewed by:
S. Emil Ruff, University of Calgary, CanadaMatthew Stott, GNS Science, New Zealand
Hisako Hirayama, Japan Agency for Marine-Earth Science and Technology, Japan
Copyright © 2017 Bessette, Moalic, Gautey, Lesongeur, Godfroy and Toffin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Laurent Toffin, laurent.toffin@ifremer.fr