ABSTRACT
The impacts of white-rot fungi on altering wood chemistry have been studied mostly in vitro. However, in vivo approaches may enable better assessment of the nature of interactions between saprotrophic fungi and host tree in nature. Hence, decayed and sound wood samples were collected from a naturally infected tree (Carpinus betulus L.). Fruiting bodies of the white rot fungus Trametes versicolor grown on the same tree were identified using rDNA ITS sequencing. Chemical compositions (cellulose and lignin) of both sound and infected wood were studied. FT-IR spectroscopy was used to collect spectra of decayed and un-decayed wood samples. The results of chemical compositions indicated that T. versicolor reduced cellulose and lignin in similar quantities. Fungal activities in decayed wood causes serious decline in pH content. The amount of alcohol-benzene soluble extractives was severely decreased, while a remarkable increase was found in 1% sodium hydroxide soluble and hot water extractive contents in the decayed wood samples, respectively. FT-IR analyses demonstrated that T. versicolor causes simultaneous white rot in the hornbeam tree in vivo which is in line with in vitro experiments.
Key words:
Hornbeam wood; Trametes versicolor; chemical compositions; FT-IR analyses
INTRODUCTION
Major decay process caused by fungi initially starts through wounds and cracks in the stem branches and roots of living trees. Later, when tree dies, other fungi gain access to the woody material and immediately speed up the decomposition process (Stokland et al. 2012STOKLAND JN, SIITONEN J AND JONSSON BG. 2012. Biodiversity in dead wood. Cambridge University Press., Schwarze et al. 2004SCHWARZE FWMR, ENGELS J AND MATTHEK C. 2004. Fungal strategies of wood decay in trees. 2nd ed., Springer, Berlin, Heidelberg, New York. , Schmidt 2006SCHMIDT O. 2006. Wood and Tree Fungi: Biology, Damage, Protection, and Use. Springer, Berlin, Heidelberg.). Several microorganisms may be involved in the degradation. The most efficient wood destroyers are the white-rot and brown-rot fungi among the Basidiomycetes (Eriksson et al. 1990ERIKSSON KEL, BLANCHETTE RA AND ANDER P. 1990. Microbial and enzymatic degradation of wood and wood components. Springer, Berlin., Eaton and Hale 1993EATON RA AND HALE MDC. 1993. Wood: decay, pests and protection. Chapman and Hall, London., Schmidt 2006). In addition, some Ascomycetes and Deuteromycetes cause a different type of wood rot called soft rot. However, some white rot fungi are able to decompose wood in a similar way to soft rot decay (Schwarze et al. 2004, Bari et al. 2017BARI E, KARIM M, OLADI R, TAJICK GHANBARY MA, GHODSKHAH DARYAEI M, SCHMIDT O, BENZ JP AND EMAMINASAB M. 2017. A comparison between decay patterns of the white-rot fungus Pleurotus ostreatus in chestnut-leaved oak (Quercus castaneifolia) shows predominantly simultaneous attack both in vivo and in vitro. For Path http://onlinelibrary.wiley.com/doi/10.1111/efp.12338/abstract
http://onlinelibrary.wiley.com/doi/10.11...
).
The majority of the white-rot fungi use cellulose and hemicelluloses at nearly the same rate; however, lignin is usually utilized at a somewhat faster rate. A few white-rot fungi, on the other hand, remove lignin and hemicelluloses preferentially, but eventually degrade all wood cell wall components. These fungi invade tissues of hardwoods and they rarely have been seen on softwoods (Schmidt 2006SCHMIDT O. 2006. Wood and Tree Fungi: Biology, Damage, Protection, and Use. Springer, Berlin, Heidelberg., Kubicek 2013KUBICEK CP. 2013. Fungi and lignocellulosic biomass. Chichester: John Wiley & Sons, Inc.). White rot has been classified by macroscopic characteristics into simultaneous and selective white rot and (Schmidt 2006, Kubicek 2013). Many white-rot fungi seem to produce just one of these behaviors, but some species can cause alternative forms under different conditions (Martínez et al. 2005MARTÍNEZ AT, SPERANZA M, RUIZ-DUEÑAS FJ, FERREIRA P, CAMARERO S, GUILLÉN F, MARTÍNEZ MJ, GUTIÉRREZ A AND DELRÍO JC. 2005. Biodegradation of lignocellulosics: microbial, chemical, and enzymatic aspects of the fungal attack of lignin. Intern Microbiol 8: 195-204., Bari et al. 2015cBARI E, SCHMIDT O AND OLADI R. 2015c. A histological investigation of Oriental beech wood decayed by Pleurotus ostreatus and Trametes versicolor. For Path 45: 349-357.). Many white-rot fungi occur as dormant spores within the live sapwood and immediately after falling moisture content in dead cells, the mycelium grows and the decomposition starts (Boddy 1994BODDY L. 1994. Latent decay fungi: the hidden foe? Arboric J 18: 113-135.).
The impacts of white-rot fungi on wood chemistry have been studied by various methods (e.g. Martínez et al. 2001MARTÍNEZ AT, CAMARERO S, GUTIÉRREZ A, BOCHINI P AND GALLETI GC. 2001. Studies on wheat lignin degradation by Pleurotus species using analytical pyrolysis. J Anal Appli Pyrol (58-59): 401-411., Mohebby 2005MOHEBBY B. 2005. Attenuated total reflection infrared spectroscopy of white-rot decayed beech wood. Int Biodeterior Biodegr 55: 247-251., Bari et al. 2015aBARI E, NAZARNEZHAD N, KAZEMI SM, TAJICK GHANBARY MA, MOHEBBY B, SCHMIDT O AND CLAUSEN CA. 2015a. Comparison of degradation capabilities of the white rot fungi Pleurotus ostreatus and Trametes versicolor. Int Biodeterior Biodegr 104: 231-237., Karim et al. 2016KARIM M, GHODSKHAH DARYAEI M, TORKAMAN J, OLADI R, TAJICK GHANBARY MA AND BARI E. 2016. In vivo investigation of chemical alteration in Oak wood decayed by Pleurotus ostreatus. Int Biodeterior Biodegr 108: 127-132.) But the most of the above mentioned methods have been carried out in vitro. However, there are inherent difficulties in emulating micro-environmental conditions occurring in the field (Rayner and Webber 1984RAYNER ADM AND WEBBER JF. 1984. Interspecific mycelial interactions - An overview. In: Jennings DH and Rayner ADM (Eds), The Ecology and Physiology of the Fungal Mycelium. Cambridge University Press, Cambridge, UK, p. 383-417.) and in vivo approaches may enable better assessment of the nature of interactions between saprotrophic fungi and the wood (Woodward and Boddy 2008WOODWARD S AND BODDY L. 2008. Interactions between saprotrophic fungi. In: Boddy L, Frankland JC and Van West P (Eds), Ecology of saprotrophic Basidiomycetes, Academic Press, Elsevier, p. 125-141.). In this regards, the white-rot fungus Trametes versicolor is a unique microorganisms that has a widespread host (Eaton and Hale 1993EATON RA AND HALE MDC. 1993. Wood: decay, pests and protection. Chapman and Hall, London., Schmidt 2006SCHMIDT O. 2006. Wood and Tree Fungi: Biology, Damage, Protection, and Use. Springer, Berlin, Heidelberg.) and act as great carbon recycler in the natural forest ecosystems. Hornbeam (Carpinus betulus) is an important tree species in Iran, economically and ecologically which accounts for 30.5% of the standing volume and 30% of the stem number in the Hyrcanian forests of Iran. This species in combination with beech compose 54 and 60% of the stem number and standing volume of the Iranian Northern Forests, respectively (Sagheb Talebi et al. 2014), and therefore, it has been studied from different aspects (Taghiyari and Sarvari Samadi 2010TAGHIYARI HR AND SARVARI SAMADI Y. 2010. Ultimate length for reporting gas permeability of Carpinus betulus wood. Special Topics & Reviews in Porous Media - An International Journal 1(4): 345-351. ).
More than 90% of wood decaying fungi in standing and felled trees in the north of Iran and America are caused by white-rotting fungi (Gilbertson 1980GILBERTSON RL. 1980. Wood-rotting fungi of North America. Mycologia 72: 1-49., Bari et al. 2015aBARI E, NAZARNEZHAD N, KAZEMI SM, TAJICK GHANBARY MA, MOHEBBY B, SCHMIDT O AND CLAUSEN CA. 2015a. Comparison of degradation capabilities of the white rot fungi Pleurotus ostreatus and Trametes versicolor. Int Biodeterior Biodegr 104: 231-237.). Thus, the white rot fungi called as predator and terminator for natural forest ecosystems (Bari et al. 2016) and many studies were carried out on them (Taghiyari et al. 2015TAGHIYARI HR, KALANTARI A, GHORBANI M, BAVANEGHI F AND AKHTARI M. 2015. Effects of fungal exposure on air and liquid permeability of nanosilver-and nanozincoxide impregnated Paulownia wood. Int Biodeterior Biodegr 105: 51-57.). Former studies (Bari et al. 2015a, b, c, d, 2016) demonstrated that the white-rot fungus Trametes versicolor produces simultaneous decay in beech wood in laboratory condition (in vitro) (Bari et al. 2016). In this respect, a question arises that whether the degradation patterns of hornbeam wood decayed by this fungus is the same as in beech or not? In addition, will the fungus produce simultaneous or selective decay? The naturally decomposition of wood gives us new and significant knowledge about the behavior of fungi in nature compared to controlled conditions. Moreover, investigations of decomposition processes in forest ecosystems will be useful from the natural recycling of organic substance point of views (Stokland et al. 2012STOKLAND JN, SIITONEN J AND JONSSON BG. 2012. Biodiversity in dead wood. Cambridge University Press.). Since there is no information about the behavior of this fungus in natural conditions, this study is aimed to assess the chemical composition of wood naturally decayed by T. versicolor. FT-IR spectroscopic analysis was accompanied with conventional chemical analysis to study the chemical changes in hornbeam wood attacked by the white-rot fungus T. versicolor. Comparable studies had been already done with the same fungus and Fagus orientalis wood (Bari et al. 2015a).
MATERIALS AND METHODS
FUNGI IDENTIFICATION
The fruiting bodies of white rot fungus Trametes versicolor (L.: Fr.) Pilát were collected from a fallen hornbeam tree (Carpinus betulus L.) at the Guilan forest, northwest Iran. Then morphology identification was done according to Ryvarden and Gilbertson (1993RYVARDEN L AND GILBERTSON RL. 1993. European Polypores: Volumes 1-2. Oslo: Fungiflora., 1994) and molecular identification was carried out according to Schmidt et al. (2012SCHMIDT O, GAISER O AND DUJESIEFKEN D. 2012. Molecular identification of decay fungi in the wood of urban trees. Eur J For Res 131: 885-891.) and Bari (2014BARI E. 2014. Potential of biological degradation of oriental beech wood by the white-rot fungus Pleurotus ostreatus and the effects on mechanical and chemical properties there of and its comparison with standard the white-rot fungus Trametes versicolor. Master Thesis, Sari Agriculture and Natural Resources University, Sari, Iran. (Unpublished).) with ten replicates. In brief, the DNA of the infected wood and fruiting bodies of fungi were extracted; PCR was amplified with primers ITS1 and ITS4, electrophoresed, purified, and sequenced. Thereafter, the identification was accomplished by the sequence comparison with sequences deposited in the DNA databases using the BLAST in the NCBI. The same tree was used for further chemical analyses (see below).
CHEMICAL ANALYSIS
Un-decayed and parts of the naturally decayed wood specimen were cut, dried at 103±2 °C for 24 hours, and then ground to pass a 40-mesh (420-μm) screen. The preparation of the wood powder for chemical composition analysis was done according to TAPPI standard (T 264 om-88, 1988). Lignin and cellulose content of both kinds of wood samples were measured using T 222 om-98 (1998), and T 17 wd-97 (1997a) procedures, respectively. The chemical percent of wood extractives was evaluated in accordance with TAPPI T 212 om-93 (1993a), TAPPI T 207 om-93 (1993b), and TAPPI T 204 cm-97 (1997b) standard methods, respectively. The pH values of the samples were calculated using a pH meter device. For this purpose, about 1 grams of meshed wood powder was added to 25ml distilled water and they were kept in room condition overnight. Then, the pH meter was calibrated with distilled water and pH values of the samples were then determined. All analyses were done with five replicates.
FT-IR SPECTROSCOPY
Dried samples were milled and passed through a mesh 40 sieve mesh. FT-IR spectra were obtained directly from wood powder. Using a Shimadzu 8400s FT-IR Spectrometer equipped with DLATGS detector, all samples were examined at a spectral resolution of 4 cm-1: Spectra from 30 scans of sample and background scans were measured. The background spectra were collected using an empty collector. A rubber band method was used for baseline correction. The band for CO2 was removed to make a suitable baseline correction (Mohebby 2005MOHEBBY B. 2005. Attenuated total reflection infrared spectroscopy of white-rot decayed beech wood. Int Biodeterior Biodegr 55: 247-251.).
STATISTICAL ANALYSIS
To compare chemical losses, a Student t-test was performed (95% confidence level) between decayed and un-decayed samples. Statistical analysis was performed using the SPSS software program, version 17 (2010).
RESULTS AND DISCUSSION
FUNGI IDENTIFICATION
The results indicated that three fungi i.e. Trametes versicolor, Trichoderma harzianum (Rifai.) and Mortierella elongata (Linnem.) were present in the samples, while T. versicolor was the only found white-rot fungus. On the other hand, T. harzianum and M. elongate as members of the Pezizomycetes and Mucoromycetes might have participated in the decomposition of wooden disc. T. harzianum is known as a mycoparasite and a cellulolytic fungus, while M. elongata is a terrestrial saprophytic fungus inhabiting soil that may use sugar material from wood or other plants (Eaton and Hale 1993EATON RA AND HALE MDC. 1993. Wood: decay, pests and protection. Chapman and Hall, London.).
CHEMICAL ANALYSIS
The average chemical components in un-decayed and naturally-decayed hornbeam samples by T. versicolor are shown in Table I. As depicted in this figure, mean cellulose loss was a bit more pronounced compared to lignin degradation. On the other hand, at the examined stage of decay, cellulose and lignin were degraded approximately at the same rate. T. versicolor contains the full lingocellulosic enzyme system for degrading all wood cell walls layers. Entire decomposition of wood cell walls by fungi will take place only when the fungi have all necessary enzymes for degradation. In this regard, only some wood decaying fungi such as Phanerochaete chrysosporium, P. carnosa and T. versicolor have the full genome for degradation (Canam et al. 2013CANAM T, TOWN J, IROBA K, TABIL L AND DUMONCEAUX T. 2013. Pretreatment of lignocellulosic biomass using microorganisms: approaches, advantages, and limitations. In: Chandel AK and Silva SS (Eds), Sustainable degradation of lignocellulosic biomass- techniques, applications and commercialization, p. 181-206.). However, fungi which are specialized may have different behavior in nature than under controlled conditions. There are also evidences that certain cell wall components may induce the production of the necessary enzymes. Thus cellulose induces the formation of carbohydrolases in white-rotting fungi (Fengel and Wegener 1989FENGEL D AND WEGENER G. 1989. Wood: Chemistry, ultrastructure, reactions, 2nd ed., de Gruyter, Berlin.). Nonetheless, since T. versicolor is a member of the white-rot fungi, these behaviors were expected. Several literatures (e.g. Eriksson et al. 1990ERIKSSON KEL, BLANCHETTE RA AND ANDER P. 1990. Microbial and enzymatic degradation of wood and wood components. Springer, Berlin.) as well as our former studies (Bari et al. 2015aBARI E, NAZARNEZHAD N, KAZEMI SM, TAJICK GHANBARY MA, MOHEBBY B, SCHMIDT O AND CLAUSEN CA. 2015a. Comparison of degradation capabilities of the white rot fungi Pleurotus ostreatus and Trametes versicolor. Int Biodeterior Biodegr 104: 231-237., c) in controlled situation, demonstrated the simultaneous decay behavior of this fungi. The average losses of cellulose and lignin were 32.52% and 26.10%, respectively which were in accordance with former studies and our investigations on Fagus orientalis wood (Bari et al. 2015a). A significant decrease was seen in pH value in naturally decayed wood. According to Humar et al. (2001HUMAR M, PETRIC M, POHLEVEN F. 2001. Changes of the pH value of impregnated wood during exposure to wood rotting fungi. Holz Roh Werkst 59: 288-293.), the fungi are able to severely reduce the pH in wood after advancement of decay. Nonetheless, it has been known that white-rot fungi do not cause such massive decline in pH of wood comparing to brown-rot fungi (Shortle 1990SHORTLE WC. 1990. Ionization of wood during previsual stages of wood decay. In: Llewellyn GC and O’Rear CE (Eds), Biodeterioration Research, Vol 3: mycotoxins, biotoxins, wood decay, air quality, cultural properties, general biodeterioration, and degradation. Springer Science + Business Media New York, p. 333-348.). On the other hand, considerable changes were recorded in extractives constituent values of decayed wood. Reduction of alcohol-benzene soluble extractives was observed in decayed samples. However, 1% NaOH soluble extractives as well as hot-water soluble extractives were increased in decayed wood samples. It was found that the wood decaying fungi hydrolyze the extractives such as styrel esters and waxes into fatty acid and sterol moieties as an important source of carbon for the their growth (Silk et al. 2001SILK PJ, AUBRY C, LONERGAN GC AND MACAULAY JB. 2001. Chlorometabolite production by the ecologically important white rot fungus. Chemosphere 44: 7. ). In this regards, similar observations (e.g. Malakani et al. 2014MALAKANI M, KHADEMIESLAM H, HOSSEINI HASHEMI SK AND ZEINALY F. 2014. Influence of fungal decay on chemi-mechanical properties of beech wood (Fagus orientalis). Cellulose Chem Technol 48: 97-103.) were reported.
Average of percent chemical components in decayed hornbeam wood in sound and naturally infected sample by white-rot fungus Trametes versicolor.
FT-IR EVALUATIONS UNDECAYED WOOD (CONTROL)
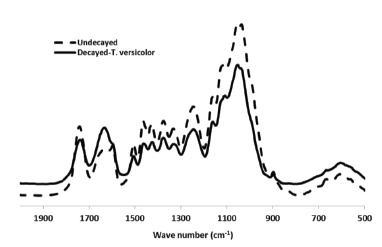
The FT-IR spectrum of un-decayed hornbeam wood is depicted in Figure 1. A strong hydrogen bonded (O-H) stretching absorption was observed at 3500-3300 cm−1 (1) and a prominent C-H stretching absorption around 2896 cm−1 (2). On top of that, there were many well-defined peaks in the fingerprint region between 1800-500 cm−1. The peaks in the fingerprint were assigned in accordance with several studies (e.g. Harrington et al. 1964HARRINGTON KJ, HIGGINS HG AND MICHELL AJ. 1964. Infrared spectra of Eucalypus regnans F. Muell. and Pinus radiata D. Don. Holzforschung 18: 108-113., Hergert 1971HERGERT HL. 1971. Infrared spectra. In: Sarkanen KV and Ludwig CH (Eds), Lignins: Occurrence, Formation, Structure and Reactions. Wiley Interscience, New York, p. 267-297., Schultz and Glasser 1986SCHULTZ TP AND GLASSER WG. 1986. Quantitative structural analysis of lignin by diffuse reflectance fourier transform spectrometry. Holzforschung 40: 37-44., Faix 1992FAIX O. 1992. Fourier transform infrared spectroscopy. In: Lin SY AND Dence CW (Eds), Methods in Lignin Chemistry. Springer, Berlin, p. 83-109., Pandey and Pitman 2003PANDEY KK AND PITMAN AJ. 2003. FTIR studies of the changes in wood following decay by brown-rot and white-rot fungi. Int Biodeterior Biodegr 52: 151-160., Mohebby 2005MOHEBBY B. 2005. Attenuated total reflection infrared spectroscopy of white-rot decayed beech wood. Int Biodeterior Biodegr 55: 247-251.), the assigned peaks in the above mentioned literatures were un-conjugated at 1738 cm−1 (3); conjugated C-O stretching at 1594 cm−1 (4); aromatic skeletal vibration at 1504 cm−1 (5); C-H deformation in lignin and carbohydrates at wave numbers 1461 (6) and 1423 cm−1 (7); C-H deformation in cellulose and hemicelluloses at 1372 cm−1 (8); C-H vibration in cellulose and C1-O vibration in syringyl derivatives at 1325 cm−1 (9); syringyl ring and C-O stretching in lignin and xylan at wave number 1238 cm−1 (10); C-O-C vibration in cellulose and hemicellulose at 1157 cm−1 (11); aromatic skeletal and C-O stretch at 1096 cm−1 (12); C-O stretch in cellulose and hemicelluloses at wave number 1035 cm−1 (13) and C-H deformation in cellulose at 892 cm−1 (14).
FT-IR SPECTRA OF DECAYED WOOD
The result of FT-IR spectra analyses of decayed and un-decayed wood is summarized in Table II. The FT-IR spectra of decayed hornbeam wood (Fig. 2) demonstrated changes of peaks in fingerprint regions at the naturally infected wood by T. versicolor. The comparison between the peaks from the decayed and un-decayed wood revealed that some peaks had severe changes; thus the subtraction of peaks demonstrated that amounts of abortion in the most picks were slightly increased in the fingerprint region. In Fig. 3 the baselines shows high differences between the un-decayed and the decayed wood; while the peaks under the baselines showed an increase of bands and above the baselines indicated prominent declined of the bands.
The molecular chemical composition changes in hornbeam wood in sound and naturally infected sample by white-rot fungus Trametes versicolor.
Subtracted FT-IR spectra of the white-rot decayed hornbeam wood in naturally infected (in vivo) by Trametes versicolor.
The FT-IR spectra of decayed and un-decayed hornbeam wood (Fig. 1) demonstrated changes of the peaks in fingerprint regions of wood decayed by T. versicolor. There were many well-defined peaks in the fingerprint region of 1800-500 cm−1. A severe decline took place at 1738 cm−1 which caused heavily reduction belonging to unconjugated groups in lignin and carboxylic acid ester hemicelluloses (Takahashi et al. 1989TAKAHASHI M, IMAMURA Y AND TANAHASHI M. 1989. Effect of acetylation on decay resistance of wood against brown rot, white rot and soft rot. International Chemistry, Congress of Pacific Basin Societies, Agrochemistry, Sub-symposium on Chemical Modification of lignocellulosic Materials-Chemical Reactions, 16 p.), This behavior was expected for this fungus; because white rot fungi use all three major chemical compositions in the cell wood wall (Eriksson et al. 1990ERIKSSON KEL, BLANCHETTE RA AND ANDER P. 1990. Microbial and enzymatic degradation of wood and wood components. Springer, Berlin., Eaton and Hale 1993EATON RA AND HALE MDC. 1993. Wood: decay, pests and protection. Chapman and Hall, London., Schmidt 2006SCHMIDT O. 2006. Wood and Tree Fungi: Biology, Damage, Protection, and Use. Springer, Berlin, Heidelberg., Kubicek 2013KUBICEK CP. 2013. Fungi and lignocellulosic biomass. Chichester: John Wiley & Sons, Inc.), as a result of fungal activities, a considerable increase at 1594 cm−1 peak was seen which is related to carbon-oxygen bond (Pandey and Pitman 2003PANDEY KK AND PITMAN AJ. 2003. FTIR studies of the changes in wood following decay by brown-rot and white-rot fungi. Int Biodeterior Biodegr 52: 151-160.), a carbon-oxygen bond is a covalent bond between carbon and oxygen (Carey and Sundberg 2007CAREY FA AND SUNDBERG RJ. 2007. Advanced Organic Chemistry, 5th ed., Springer-Verlag, Berlin, New York.), which was increased after the attack by white rot fungi (Fengel and Wegener 1989FENGEL D AND WEGENER G. 1989. Wood: Chemistry, ultrastructure, reactions, 2nd ed., de Gruyter, Berlin.). Microbial cellulolysis is a very complex procedure, especially in view of that portion of the mechanism that relates to the degradation of hydrogen bond-ordered cellulose (Wood and Garcia-Campayo 1994WOOD TM AND GARCIA-CAMPAYO V. 1994. Enzymes and mechanisms involved in microbial cellulolysis. In: Ratledge C (Ed), Biochemistry of microbial degradation. Springer Science + Business Media, B.V., p. 197-231.). Therefore, cellulolysis by white-rot fungi involves the interaction of enzymes defined as exoglucanases and endoglucanases as well as ß- glucosidases (Eriksson and Wood 1985). Carbon and oxygen form terminal double bonds in functional groups are collectively known as carbonyl compounds to which belong such compounds as ketones, esters, carboxylic acids and many more. Compounds with formal C-O triple bonds do not exist except for carbon monoxide, which has a very short, strong bond (reported from the standard bond energies). The wood-inhabiting microorganisms use carbon only from enzymatically easily accessible and digestible substrates, like simply constructed sugars, peptides, or fats, or from the storage material starch in the parenchyma cells. The wood decay fungi use carbon additionally from the complex, main components of the woody cell wall, cellulose, hemicelluloses, and lignin (Fengel and Wegener 1989, Schmidt 2006, Kubicek 2013) decreases in intensity observed at around 1504; 1461; 1423; 1372; 1325; 1238; 1157; 1096 cm−1 were related to aromatic skeletal; benzene ring vibration in lignin, deformation in lignin and carbohydrates as well as benzene ring vibration in lignin; aromatic skeletal vibrations combined with C-H in plane deformation, deformation in cellulose and hemicelluloses and bending vibration in cellulose and hemicelluloses, vibration in cellulose, C1-O in syringyl derivatives, syringyl ring C-O stretch in lignin and xylan, vibration in cellulose and hemicelluloses, aromatic skeletal and C-C stretch and association band in cellulose and hemicelluloses, respectively. This condition of attack by white-rot fungi has been reported in several studies (Pandey and Pitman 2003, Faix et al. 1993FAIX O, BREMER J, SCHMIDT O AND STEVANOVIC T. 1993. Monitoring of chemical changes in white-rot degraded beech wood by pyrolysis gas chromatography and Fourier transform infrared spectroscopy. J Anal Appli Pyrol 21: 147-162., Mohebby 2005MOHEBBY B. 2005. Attenuated total reflection infrared spectroscopy of white-rot decayed beech wood. Int Biodeterior Biodegr 55: 247-251., Yilgor et al. 2013YILGOR N, DOGU D, MOORE R, TERZI E AND KARTAL SN. 2013. Evaluation of fungal deterioration in Liquidambar orientalis Mill. heartwood by FT-IR and light microscopy. BioResources 8: 2805-2826., Bari et al. 2015aBARI E, NAZARNEZHAD N, KAZEMI SM, TAJICK GHANBARY MA, MOHEBBY B, SCHMIDT O AND CLAUSEN CA. 2015a. Comparison of degradation capabilities of the white rot fungi Pleurotus ostreatus and Trametes versicolor. Int Biodeterior Biodegr 104: 231-237.). Different types of hydrolyzing enzyme could be liberated to alter and break linkages in the cell-wall components and release them as small molecules. During this period, many chemical changes occur in these constituents, indicating that the fungus could assimilate them as carbon sources. A strong decrease took place in the carbon-oxygen bonds which was seen at the 1035 cm−1 bond that demonstrated a decrease in primary alcohol. Moreover, a slight decline was observed at 892 cm−1 which belongs to the deformation in cellulose as well as C1 group frequency in cellulose and hemicellulose (Mohebby 2005, Bari et al. 2015a).
CONCLUSIONS
The white-rot fungus Trametes versicolor caused similar chemical components losses in the xylem of hornbeam wood samples at the advanced decay stages in vivo. FT-IR spectroscopy along with chemical analyses evaluated the qualitative and quantitative changes in lignin and carbohydrate components of infected wood by this fungus. The results showed that T. versicolor reduced lignin and carbohydrate nearly at the same rate, with a slight preference for lignin which was reflected in a small reduction in the lignin/carbohydrate peak area ratio as decay progressed. Nonetheless, T. versicolor had the ability to degrade all three cell wall polymers. Overall, this study proved that this fungus induce simultaneous white rot decay in hornbeam wood in natural situation (in vivo).
ACKNOWLEDGMENTS
Thanks are to Dr. Nadre Bahramifar University of Tarbiat Modares for cooperation in preparation samples for FT-IR analyses and Mrs. Hakimeh Ziaie Juibari, Sari Agriculture and Natural Resources University for collaboration in preparation samples of fungal for DNA extraction.
REFERENCES
- BARI E. 2014. Potential of biological degradation of oriental beech wood by the white-rot fungus Pleurotus ostreatus and the effects on mechanical and chemical properties there of and its comparison with standard the white-rot fungus Trametes versicolor. Master Thesis, Sari Agriculture and Natural Resources University, Sari, Iran. (Unpublished).
- BARI E, KARIM M, OLADI R, TAJICK GHANBARY MA, GHODSKHAH DARYAEI M, SCHMIDT O, BENZ JP AND EMAMINASAB M. 2017. A comparison between decay patterns of the white-rot fungus Pleurotus ostreatus in chestnut-leaved oak (Quercus castaneifolia) shows predominantly simultaneous attack both in vivo and in vitro. For Path http://onlinelibrary.wiley.com/doi/10.1111/efp.12338/abstract
» http://onlinelibrary.wiley.com/doi/10.1111/efp.12338/abstract - BARI E, NAZARNEZHAD N, KAZEMI SM, TAJICK GHANBARY MA, MOHEBBY B, SCHMIDT O AND CLAUSEN CA. 2015a. Comparison of degradation capabilities of the white rot fungi Pleurotus ostreatus and Trametes versicolor. Int Biodeterior Biodegr 104: 231-237.
- BARI E, OLADI R, SCHMIDT O, CLAUSEN CA, OHNO K, NICHOLAS DD, GHODSKHAH DARYAEI M AND KARIM M. 2015b. Influence of xylem ray integrity and degree of polymerization on bending strength of beech wood decayed by Pleurotus ostreatus and Trametes versicolor. Int Biodeterior Biodegr 104: 299-306.
- BARI E, SCHMIDT O AND OLADI R. 2015c. A histological investigation of Oriental beech wood decayed by Pleurotus ostreatus and Trametes versicolor. For Path 45: 349-357.
- BARI E, TAGHIYARI HR, MOHEBBY B, CLAUSEN CA, SCHMIDT O AND VASEGHI MJ. 2015d. Mechanical properties and chemical composition of beech wood exposed for 30 and 120 days to white-rot fungi. Holzforschung 69: 587-593.
- BARI E, TAGHIYARI HR, NAJI HR, SCHMIDT O, OHNO MK, CLAUSEN CA AND BAKAR ES. 2016. Assessing the destructive behavior of two white-rot fungi on beech wood. Int Biodeterior Biodegr 114: 129-140.
- BODDY L. 1994. Latent decay fungi: the hidden foe? Arboric J 18: 113-135.
- CANAM T, TOWN J, IROBA K, TABIL L AND DUMONCEAUX T. 2013. Pretreatment of lignocellulosic biomass using microorganisms: approaches, advantages, and limitations. In: Chandel AK and Silva SS (Eds), Sustainable degradation of lignocellulosic biomass- techniques, applications and commercialization, p. 181-206.
- CAREY FA AND SUNDBERG RJ. 2007. Advanced Organic Chemistry, 5th ed., Springer-Verlag, Berlin, New York.
- EATON RA AND HALE MDC. 1993. Wood: decay, pests and protection. Chapman and Hall, London.
- ERIKSSON KEL, BLANCHETTE RA AND ANDER P. 1990. Microbial and enzymatic degradation of wood and wood components. Springer, Berlin.
- ERIKSSON KEL AND WOOD TM. 1985. Biodegradation of cellulose. In: Higuchi T (Ed), Biosynthesis and Biodegradation of Wood Components. Academic Press, London, p. 469-504.
- FAIX O. 1992. Fourier transform infrared spectroscopy. In: Lin SY AND Dence CW (Eds), Methods in Lignin Chemistry. Springer, Berlin, p. 83-109.
- FAIX O, BREMER J, SCHMIDT O AND STEVANOVIC T. 1993. Monitoring of chemical changes in white-rot degraded beech wood by pyrolysis gas chromatography and Fourier transform infrared spectroscopy. J Anal Appli Pyrol 21: 147-162.
- FENGEL D AND WEGENER G. 1989. Wood: Chemistry, ultrastructure, reactions, 2nd ed., de Gruyter, Berlin.
- GILBERTSON RL. 1980. Wood-rotting fungi of North America. Mycologia 72: 1-49.
- HARRINGTON KJ, HIGGINS HG AND MICHELL AJ. 1964. Infrared spectra of Eucalypus regnans F. Muell. and Pinus radiata D. Don. Holzforschung 18: 108-113.
- HERGERT HL. 1971. Infrared spectra. In: Sarkanen KV and Ludwig CH (Eds), Lignins: Occurrence, Formation, Structure and Reactions. Wiley Interscience, New York, p. 267-297.
- HUMAR M, PETRIC M, POHLEVEN F. 2001. Changes of the pH value of impregnated wood during exposure to wood rotting fungi. Holz Roh Werkst 59: 288-293.
- KARIM M, GHODSKHAH DARYAEI M, TORKAMAN J, OLADI R, TAJICK GHANBARY MA AND BARI E. 2016. In vivo investigation of chemical alteration in Oak wood decayed by Pleurotus ostreatus. Int Biodeterior Biodegr 108: 127-132.
- KUBICEK CP. 2013. Fungi and lignocellulosic biomass. Chichester: John Wiley & Sons, Inc.
- MALAKANI M, KHADEMIESLAM H, HOSSEINI HASHEMI SK AND ZEINALY F. 2014. Influence of fungal decay on chemi-mechanical properties of beech wood (Fagus orientalis). Cellulose Chem Technol 48: 97-103.
- MARTÍNEZ AT, CAMARERO S, GUTIÉRREZ A, BOCHINI P AND GALLETI GC. 2001. Studies on wheat lignin degradation by Pleurotus species using analytical pyrolysis. J Anal Appli Pyrol (58-59): 401-411.
- MARTÍNEZ AT, SPERANZA M, RUIZ-DUEÑAS FJ, FERREIRA P, CAMARERO S, GUILLÉN F, MARTÍNEZ MJ, GUTIÉRREZ A AND DELRÍO JC. 2005. Biodegradation of lignocellulosics: microbial, chemical, and enzymatic aspects of the fungal attack of lignin. Intern Microbiol 8: 195-204.
- MOHEBBY B. 2005. Attenuated total reflection infrared spectroscopy of white-rot decayed beech wood. Int Biodeterior Biodegr 55: 247-251.
- PANDEY KK AND PITMAN AJ. 2003. FTIR studies of the changes in wood following decay by brown-rot and white-rot fungi. Int Biodeterior Biodegr 52: 151-160.
- RAYNER ADM AND WEBBER JF. 1984. Interspecific mycelial interactions - An overview. In: Jennings DH and Rayner ADM (Eds), The Ecology and Physiology of the Fungal Mycelium. Cambridge University Press, Cambridge, UK, p. 383-417.
- RYVARDEN L AND GILBERTSON RL. 1993. European Polypores: Volumes 1-2. Oslo: Fungiflora.
- RYVARDEN L AND GILBERTSON RL. 1994. European Polypores: Volumes 2. Oslo: Fungiflora.
- SAGHEB TALEBI K, SAJEDI T AND POURHASHEMI M. 2014. Forests of Iran: A Treasure from the Past, a Hope for the Future Plant and Vegetation, Vol 10. Springer, Netherlands.
- SCHMIDT O. 2006. Wood and Tree Fungi: Biology, Damage, Protection, and Use. Springer, Berlin, Heidelberg.
- SCHMIDT O, GAISER O AND DUJESIEFKEN D. 2012. Molecular identification of decay fungi in the wood of urban trees. Eur J For Res 131: 885-891.
- SCHULTZ TP AND GLASSER WG. 1986. Quantitative structural analysis of lignin by diffuse reflectance fourier transform spectrometry. Holzforschung 40: 37-44.
- SCHWARZE FWMR, ENGELS J AND MATTHEK C. 2004. Fungal strategies of wood decay in trees. 2nd ed., Springer, Berlin, Heidelberg, New York.
- SHORTLE WC. 1990. Ionization of wood during previsual stages of wood decay. In: Llewellyn GC and O’Rear CE (Eds), Biodeterioration Research, Vol 3: mycotoxins, biotoxins, wood decay, air quality, cultural properties, general biodeterioration, and degradation. Springer Science + Business Media New York, p. 333-348.
- SILK PJ, AUBRY C, LONERGAN GC AND MACAULAY JB. 2001. Chlorometabolite production by the ecologically important white rot fungus. Chemosphere 44: 7.
- STOKLAND JN, SIITONEN J AND JONSSON BG. 2012. Biodiversity in dead wood. Cambridge University Press.
- TAGHIYARI HR, KALANTARI A, GHORBANI M, BAVANEGHI F AND AKHTARI M. 2015. Effects of fungal exposure on air and liquid permeability of nanosilver-and nanozincoxide impregnated Paulownia wood. Int Biodeterior Biodegr 105: 51-57.
- TAGHIYARI HR AND SARVARI SAMADI Y. 2010. Ultimate length for reporting gas permeability of Carpinus betulus wood. Special Topics & Reviews in Porous Media - An International Journal 1(4): 345-351.
- TAKAHASHI M, IMAMURA Y AND TANAHASHI M. 1989. Effect of acetylation on decay resistance of wood against brown rot, white rot and soft rot. International Chemistry, Congress of Pacific Basin Societies, Agrochemistry, Sub-symposium on Chemical Modification of lignocellulosic Materials-Chemical Reactions, 16 p.
- TAPPI STANDARD. 1988. Preparation of wood for chemical analysis, T 264 om-88.
- TAPPI STANDARD. 1993a. One percent sodium hydroxide solubility of wood and pulp, T 212 om-93.
- TAPPI STANDARD. 1993b. Water solubility of wood and pulp, T 207 om-93.
- TAPPI STANDARD. 1998. Standard methods for acid-insoluble lignin in wood and pulp, T 222. om-98.
- TAPPI STANDARD. 1997a. Cellulose in wood, T 17 wd-97.
- TAPPI STANDARD. 1997b. Wood extractives in ethanol-benzene mixture, T 204 cm-97.
- WOOD TM AND GARCIA-CAMPAYO V. 1994. Enzymes and mechanisms involved in microbial cellulolysis. In: Ratledge C (Ed), Biochemistry of microbial degradation. Springer Science + Business Media, B.V., p. 197-231.
- WOODWARD S AND BODDY L. 2008. Interactions between saprotrophic fungi. In: Boddy L, Frankland JC and Van West P (Eds), Ecology of saprotrophic Basidiomycetes, Academic Press, Elsevier, p. 125-141.
- YILGOR N, DOGU D, MOORE R, TERZI E AND KARTAL SN. 2013. Evaluation of fungal deterioration in Liquidambar orientalis Mill. heartwood by FT-IR and light microscopy. BioResources 8: 2805-2826.
Publication Dates
-
Publication in this collection
07 Dec 2017 -
Date of issue
Oct-Dec 2017
History
-
Received
17 Oct 2016 -
Accepted
28 Nov 2016