

Abstract
The ability of neurons to fire rapid action potential relies on the expression of voltage-gated sodium channels; the onset of the transcription of genes that encode these channels occurs during early neuronal development. The factors that direct and regulate the specific expression of ion channels are not well understood. Repressor element-1 silencing transcription/neuron-restrictive silencer factor (REST/NRSF) is a transcriptional regulator characterized as a repressor of the expression of NaV1.2, the gene encoding the voltage-gated sodium channel most abundantly expressed in the CNS, as well as of the expression of numerous other neuronal genes. In mammals, REST/NRSF is expressed mostly in non-neural cell types and immature neurons, and it is downregulated on neural maturation. To understand the mechanisms that govern sodium channel gene transcription and to explore the role of REST/NRSF in vivo, we inhibited REST/NRSF action in developing Xenopuslaevis embryos by means of a dominant negative protein or antisense oligonucleotides. Contrary to what was expected, these maneuvers result in the decrease of the expression of theNaV1.2 gene, as well as of other neuronal genes in the primary spinal neurons and cranial ganglia, without overt perturbation of neurogenesis. These results, together with the demonstration of robust REST/NRSF expression in primary spinal neurons, suggest that REST/NRSF is required for the acquisition of the differentiated functional neuronal phenotype during early development. Furthermore, they suggest that REST/NRSF may be used to activate or repress transcription of neuronal genes in distinct cellular and developmental contexts.
- primary spinal neurons
- neuronal differentiation
- repressor element silencer of transcription
- dominant negative
- antisense oligonucleotides
- sodium channels
The generation of rapid action potentials relies on the adequate expression of particular voltage-gated ion channels. During embryonic development ofXenopus laevis, primary spinal neurons transit from an electrically silent state to a condition in which they can fire mature neuronal action potentials (Spitzer and Lamborghini, 1976). Underlying this transition is the onset of the expression of voltage-gated channels (O'Dowd et al., 1988; Desarmenien et al., 1993;Olson, 1996), particularly the beginning of the expression of voltage-gated sodium currents and the rapid increase in the density of potassium currents. This process is inhibited by blockers of RNA synthesis, indicating that activation of transcription is fundamental in the acquisition of electrical activity in neurons (Ribera and Spitzer, 1989). The identity of the potassium channels upregulated during these stages has been unveiled (Ribera, 1990; Burger and Ribera, 1996; Gurantz et al., 1996; Vincent et al., 2000), but the mechanisms that govern the transcription of ion channels are unknown.
Repressor element silencing transcription/neuron-restrictive silencer factor (REST/NRSF) was identified as a factor that influences the expression of voltage-gated sodium channels (Chong et al., 1995) and also as a regulator of the expression of other genes (Schoenherr and Anderson, 1995). REST/NRSF binds a DNA element (RE-1) contained in the regulatory regions of numerous neuronal genes, includingNaV1.2 (Kraner et al., 1992; Mori et al., 1992; Schoenherr et al., 1996). The extensive characterization of REST/NRSF has led to the current view of this protein as a repressor of neural-specific gene expression; REST/NRSF is expressed in non-neural cells and undifferentiated neurons and downregulated on differentiation. REST/NRSF interacts with the corepressors Sin3 and CoREST, assembling complexes that recruit histone deacetylase activity to regions of genes that bear the RE-1 sequence (Andrés et al., 1999; Huang et al., 1999; Naruse et al., 1999; Grimes et al., 2000; Ballas et al., 2001). The in vivo role of REST/NRSF has been difficult to ascertain, given the early embryonic lethality of mice bearing a homozygous deletion of the REST/NRSF gene (Chen et al., 1998). Expression of the DNA-binding domain (DBD) of REST/NRSF in chicken embryos associates with derepression of neural genes (Chen et al., 1998), whereas constitutive expression of the murine isoform in developing neurons associates with repression of some neuronal genes and defects in axon guidance (Paquette et al., 2000). Some studies have found the expression of REST/NRSF mRNA in adult neurons (Palm et al., 1998); furthermore, it has been shown that it can act as a transcriptional activator for some genes (Bessis et al., 1997;Kallunki et al., 1998; Seth and Majzoub, 2001). Some reports also indicate that REST/NRSF may be involved in the regulation of the expression of non-neural genes (Scholl et al., 1996; Kuwahara et al., 2001). These data suggest that the actual function of REST/NRSF may be more complex than originally envisioned and that it could vary according to the cellular context in which it is studied.
MATERIALS AND METHODS
xREST-DBD construction. The DNA-binding domain of xREST (GenBank accession AF096301), comprising the sequence that encodes K138 to Q440, was amplified by PCR with the primers GCG GAATTCAAAAAGGACACCCAGCCCAAC and CGC CTCGAGTCATTGCTTATTTGCATCAACATCTG, adding oneEcoRI and one XhoI restriction sites (underlined in the primers sequences). The PCR product was cloned into the plasmid CS2+NLSMT, kindly provided by Dr. D. Turner (University of Michigan, Ann Arbor, MI). This vector adds a nuclear localization signal and six Myc epitopes to the N-terminal coding region.
Embryo manipulation, injection, and in situhybridization. X. laevis embryos were generated byin vitro fertilization, dejellied, and incubated following standard procedures. A portion (0.5–2 ng) of in vitrotranscribed xREST-DBD RNA or 2–20 ng of a morpholino-modified antisense oligonucleotide (sequence ACTGGTTGACCATTTGAGTGGC) (Gene Tools, Philomath, OR) were injected in a total volume of 10 nl, in one cell at the two cell stage or in two cells at the four cell stage. The RNA encoding β-galactosidase (0.5 ng) was coinjected in all cases. Embryos were incubated at 18°C until neurula stage and then prefixed; the injected side was detected by developing the β-galactosidase reaction, and the whole-mount in situ hybridization (ISH) procedure, using 11-UTP digoxigenin (Boehringer Mannheim, Mannheim, Germany)-labeled probes, was followed according to described methods (Sive et al., 2000).
ISH probes. xNaV1.2 was generated by reverse transcription-PCR (RT-PCR) using primers designed on the basis of the conserved sequences in vertebrate homologs and RNA isolated from X. laevis neural tube as input. The primers sequence was CCIRYIAAYGGIAARATGA and GGIGCIYTNGCNTTYGARGAY. PCR fragments were cloned (Topo-TA; Invitrogen, Carlsbad, CA) and sequenced (GenBank access code AY121368). The X. laevisSCG10 (GenBank X71434) probe was also generated by RT-PCR with the primers GCTCGCTACCCTCTCCACT and CATACTGATATCGCATGATCCGT. Other probes were obtained from the sources indicated: Xslug, Xsox2, and N-βII tubulin from Roberto Mayor (Universidad de Chile, Santiago, Chile); Hox11L2 from Paul Krieg (University of Arizona Health Sciences Center, Tucson, AZ); Pax3 from Andre Brandli (Swiss Federal Institute of Technology, Zurich, Switzerland); and xREST8 from Gail Mandel (Howard Hughes Medical Institute, State University of New York, Stony Brook, NY).
RT-PCR, cell culture, and single-cell RT-PCR. RT-PCR detection of the REST/NRSF transcript was performed using total RNA isolated from whole embryos or dissected tissues and the following primers: F1, AGAACGACCATATCAGTGTATTC; R2, CTGAGTAGATGACAAGCGCC; R1, AATCAGATTTTCTTTTCACACAACTAGA.
Neural plate cell cultures were prepared, and subsequent single-cell RT-PCR of morphologically differentiated neurons was conducted as described (Ribera and Spitzer, 1989; Gurantz et al., 1996). Two different nested PCR reactions were performed with the samples obtained from one cell, one for neuronal tubulin (N-tubulin) mRNA (231 bp product) and the other for xREST/NRSF. The primers for N-tubulin were as follows: F1, GCCAGTGCGGTAACCAAATTG; F2, ACACGGCATTGATCCTACAG; R2, AGCTCCTTCGGTGTAATGAC; R1, TCATGATTCGGTCTGGGTACTC.
To discard contamination with genomic DNA, xREST/NRSF primers were designed flanking an intron; amplification of genomic DNA results in fragments of ∼2 kb, in contrast with the 557 and 660 bp fragments resulting from the amplification of cDNA. PCR products were cloned and sequenced to confirm their identities.
Electrophoretical mobility shift assays. Gel retardation assays were performed using 0.1–1 ng of a 3′-end32P-labeled DNA fragment containing two RE-1 motifs in tandem, kindly provided by Dr. G. Mandel (Chong et al., 1995). Crude protein extracts were prepared by freon extraction of uninjected or injected embryos homogenized in the presence of a mixture of protease inhibitors. The protein extracts and the antibody against the Myc epitope (monoclonal antibody 9E10; Sigma, St. Louis, MO) were added to the mix and incubated at 4°C for 15 min. Then, the labeled probe was added, and the reactions were incubated at 30°C for 25 min. The resulting complexes were analyzed in 4% polyacrylamide gels containing 5% glycerol and 0.5 × Tris-borate-EDTA buffer. The gels were dried, and the complexes were visualized by autoradiography.
RESULTS
REST and NaV1.2 are expressed in primary neurons during neurulation
To study the regulation of the expression of sodium channel genes by REST/NRSF during neurulation, we generated a probe for the X. laevis NaV1.2 ortholog (xNaV1.2). xNaV1.2 is expressed from stage 16 in neural plate, as evaluated by RT-PCR, whereas ISH detects transcripts from stage 18 in spinal neurons, which is consistent with functional studies of the onset of sodium current expression in this system (Fig.1a—d,g). Analysis of sections shows the expression of this transcript restricted to the neural tube, particularly to its lateral and ventral regions (Fig.1d). At later stages, xNaV1.2 is also expressed in the cranial ganglia primordia (Fig. 1c).

NaV1.2 and REST/NRSF are expressed in neural tissues during development of X. laevis.a–d, ISH for xNaV1.2: expression is restricted to primary spinal neurons (sn), cranial ganglia primordia (cg), and olfactory placodes (op). In a section of a stage 18 embryo, xNaV 1.2 expression is observed in the lateral and ventral regions of the neural tube. a, Dorsal view, stage 18; b, lateral view, stage 18;c, anterior view, stage 24; d, central transversal section, stage 18; n, notochord;m, presomitic mesoderm; e,left, ISH showing diffuse expression of REST during neurulation (stage 18), including neural folds; right, a sense REST probe does not produce significant labeling in a stage 18 embryo; f, at stage 35, REST/NRSF expression is stronger in the anterior neural tissue; g, RT-PCR showing the coexpression of xNaV1.2 and REST/NRSF in dissected tissues at the stages annotated; np, neural plate; nt, neural tube; pm, presomitic mesoderm; s, somites. The constitutively expressed transcript EF1α is shown as a control.
We then examined the expression pattern of REST/NRSF. X. laevis REST/NRSF mRNA is widely expressed in the developing embryo; at the neurula stage, it is found in the neural folds and neural tube, paraxial mesoderm, and other structures, as revealed by ISH and RT-PCR using RNA obtained from dissected tissues as input (Fig.1e,g). It is of interest that during neurulation, the transcripts encoding xREST/NRSF and xNaV1.2 are coexpressed in developing neural tissues. At later stages of development, during differentiation of secondary neurons (stage 35), xREST/NRSF expression is prominent in the nervous system (Fig. 1f). To confirm that the expression of REST/NRSF detected by ISH actually occurs in neurons, we analyzed its presence in identified mature neurons in culture by single-cell RT-PCR. These neurons display fully differentiated functional and morphological phenotype. In 6 of 14 cells positively identified as neurons both by morphology and by the expression of the transcript encoding neuronal tubulin, we detected the presence of REST/NRSF transcripts (data not shown).
Inhibition of REST/NRSF leads to decreased expression of NaV1.2 and other neuronal genes
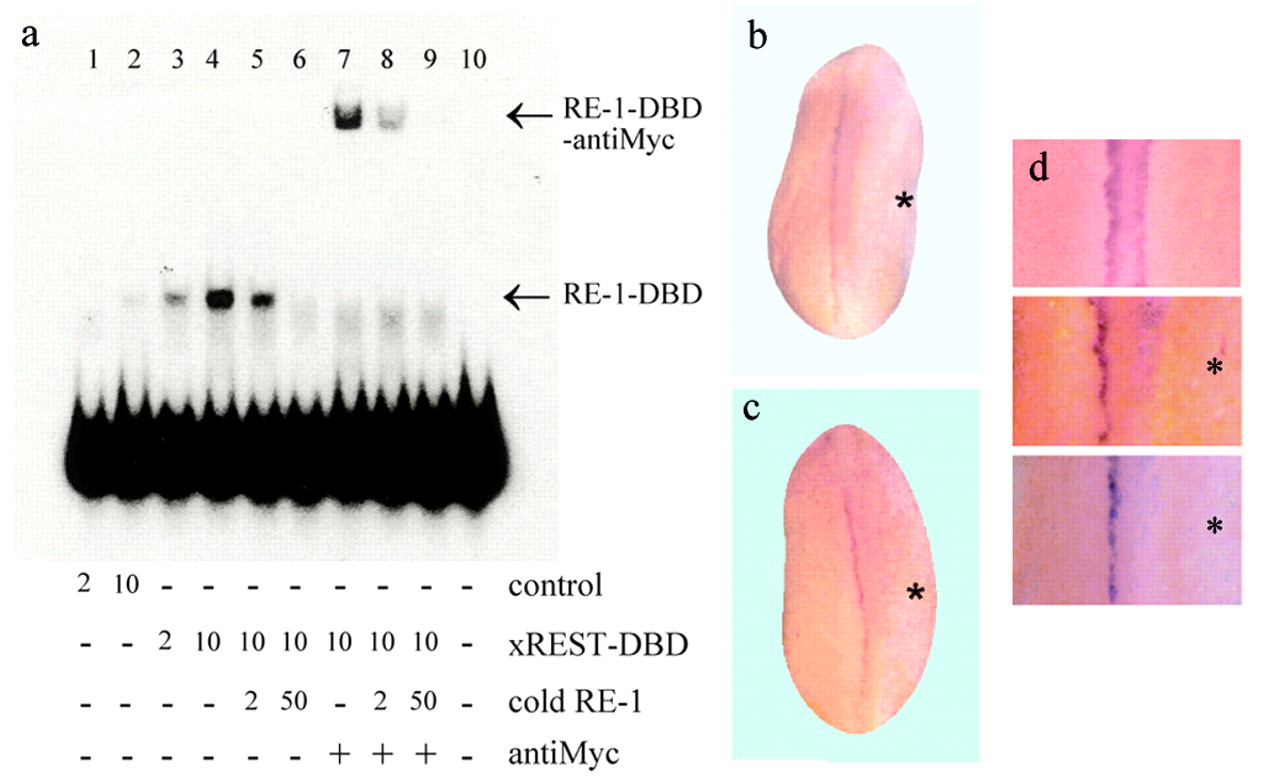
We constructed a mutated version of X. laevis REST/NRSF cDNA, which encodes a protein devoid of the transrepressor domains described for the mammalian ortholog. This protein comprises only the DBD (xREST-DBD), which shows an identity of 85% with the human and murine orthologs, including the amino acids that are critical for binding RE-1. According to the biochemical characterization of REST/NRSF, this domain can compete with the endogenous REST/NRSF for the RE-1 binding sequence, antagonizing the transcriptional effects of the wild-type protein (Chong et al., 1995). Injection of the in vitro-transcribed xREST-DBD RNA into embryos at the one or two cell stages resulted in the expression of the recombinant protein throughout early development and included neurulation, as recognized by Western analysis (data not shown). We then assayed the ability of the expressed xREST-DBD protein to bind RE-1. As shown in Figure2a, crude extracts prepared from stage 18 embryos injected with the xREST-DBD RNA contain a marked RE-1 binding activity. Extracts from uninjected embryos also appear to contain this activity, although in a much lesser amount (Lane 2). The supershift induced by the incubation with the anti-Myc antibody indicates that the observed RE-1 binding activity is because of the expressed recombinant form.

DBD-xREST binds specifically to the RE-1 and inhibits xNaV1.2 expression. a, Electrophoretical mobility shift assay analysis of the complex formation by the use of protein extracts of noninjected embryos (lanes 1–2) and embryos injected with the DBD-xREST RNA (lanes 3–9). Two or 10 μg of total protein was used in each assay. DBD-xREST binds to RE-1 (lanes 3–6). The addition of antibody against the Myc epitope produces the super-retardation of the complex (lanes 7–9). The binding specificity of the complex was evaluated by the addition of 2- or 50-fold excess of cold probe (lanes 5–6, 8–9). The probe alone is shown inLane 10. Arrows indicate the complexes.b, Expression of xREST-DBD inhibits xNaV1.2 expression during neurulation. ISH for xNaV1.2; the injected side is marked by anasterisk. Notice a single line of xNaV1.2 labeling, in contrast with Figure 1a. c, Injection of a morpholino-modified antisense oligonucleotide targeting xREST/NRSF inhibits xNaV 1.2 expression at the time of neurulation.d, Detail of a noninjected embryo (top panel) and embryos injected with xREST-DBD RNA (middle panel) or the antisense oligonucleotide (bottom panel) in one of two cells.
Expression of xREST-DBD resulted in a significant decrease of the expression of the xNaV1.2 RNA in the spinal territory of the injected side; this effect was observed in 49% of the studied embryos (n = 105; three experiments). We did not observe ectopic expression of xNaV 1.2 or increased normotopic expression, as compared with the uninjected side (Fig. 2b, middle panel in d). To test in a mechanistically independent manner the functional role of REST/NRSFin vivo, we injected a morpholino-modified antisense oligonucleotide against the xREST/NRSF into one cell at the two-cell stage embryos and examined xNaV1.2 expression at the neural tube stage. We observed a dose-dependent decrease of the transcript encoding this sodium channel in spinal neurons; at the maximal dose of the oligonucleotide tested (20 ng/injection), 50% of the embryos displayed defects in the expression of xNaV1.2 (n = 130; three experiments); we did not observe extraneural expression of this transcript (Fig. 2c, bottom panel ind).
Because many genes contain RE-1 sequences in their regulatory regions, we assayed the effect of xREST-DBD on the expression of other neuronal-specific genes known to contain RE-1 sequences in other species. We found a consistent decrease in the expression of the N-tubulin and SCG10 transcripts in the primary neural territories. Thus, 58% of the embryos injected with the xREST-DBD RNA displayed partial or complete absence of the N-tubulin transcript in the injected side (n = 112; four experiments), whereas absence of SCG10 was observed in 61% of the cases (n = 98; three experiments) (Fig. 3a–d).

xREST-DBD decreases the expression of other neuronal genes but does not perturb neural induction.a–c, ISH for N-tubulin; the injected side is marked by an asterisk; b, both lateral views;c, anterior view of the same embryo, where thearrowheads signal the label of the primary spinal neurons (sn) and cranial ganglia (cg).d, xREST-DBD represses SCG10 expression.Doublearrowheads signal the expected normal position of N-tubulin or SCG10 expression. The spotted labeling in the injected side (a) and thediffuse hue in c result from β-galactosidase activity staining. e–h, xREST-DBD does not affect neural induction and early differentiation. Expression patterns of Sox2 (e), Slug (f), Hox11L12 (g), and Pax-3 (h). e, h, Anterior views; f, g, dorsal views.
Inhibition of REST function does not affect neurogenesis
The decreased expression of NaV1.2, N-tubulin, and SCG10 associated to the perturbation of xREST/NRSF function may be explained by decreased transcription of these neural-specific genes in a committed neuron as well as by a failure in neuronal differentiation, leading to the lack of neurons able to express these transcripts. To differentiate between these alternatives, we evaluated the effect of xREST-DBD on the expression patterns of genes used as markers of neural induction and differentiation. We did not find significant perturbations in the patterns of expression of the neural plate marker XSox2, the neural crest marker xSlug, and the homeobox genesHox11L2 and Pax-3, which are expressed in Rohon-Beard and a subset of spinal interneurons, respectively (Fig.3e–h).
DISCUSSION
To advance our understanding of the molecular mechanisms that underlie the concerted and specific expression of ion channels, we considered the detailed physiological characterization of the development of excitability performed in Xenopus primary spinal neurons. Although conservation of general mechanisms through phylogeny is expected, comparison of our results with those obtained in mammalians must consider that in amphibians, a subpopulation of neurons (primary neurons) withdraws from the mitotic cycle during early embryogenesis and forms a circuit that controls early larval behavior. In Xenopus embryos, primary neurons undergo their last round of division during neurulation (around stage 15) and then differentiate (Hartenstein, 1989). Our study focuses exclusively on this population. At the time at which we examine expression of xNaV1.2 transcripts, primary spinal neurons are in final stages of their differentiation, although the embryo is still undergoing developmental processes previous to organogenesis. In contrast, secondary neuron precursors continue replicating until stage 35 or later. Much less is known about the physiological development of this population.
We explored the role of REST/NRSF in the onset of the expression of xNaV1.2. Considering the current view about REST/NRSF function, and the widespread expression of xREST/NRSF before the onset of the expression of xNaV1.2, we postulated that a decrease in the expression of REST at the time of primary neuron differentiation would allow the function of activators of the transcription of this and other neural genes. However, we observed a maintained expression of the REST/NRSF transcript throughout neurogenesis, which does not preclude the expression of neural-specific transcripts and the acquisition of the differentiated phenotype in primary spinal neurons. In contrast, conditional expression of the full-length REST/NRSF protein in PC12 cells inhibits the neuronal differentiation normally triggered by nerve growth factor in this cell line (Ballas et al., 2001). Furthermore, we observed that the expression of xREST-DBD associates with a marked repression of three different neural genes in primary spinal neurons ofXenopus embryos, and we did not observe ectopic expression of the studied genes. The results obtained by the injection of morpholino-modified antisense oligonucleotides targeted against xREST/NRSF support the possibility that the repression is a consequence of decreased activity of the endogenous protein.
Because these results contrast with those obtained in mice embryos bearing the homozygous deletion of the REST/NRSF gene, it is necessary to emphasize that the developmental stages that we analyzed are comparable with murine embryonic day (E) 7.5–8 and that studies of REST/NRSF expression in mouse embryos have focused on later stages (Chong et al., 1995; Schoenherr and Anderson, 1995; Chen et al., 1998). Significant abnormalities in REST/NRSF-deficient embryos are observed starting at E9.5–10, and 100% lethality is observed at E11.5. Therefore, early different roles of REST/NRSF may have been missed.
A plausible mechanism that can reconcile the well described repressor role and our observations is the differential recruitment of corepressor or coactivator complexes to the promoters that bear the RE-1 motif. This differential recruitment could depend on the cell type or developmental stage-specific expression of the activators or repressors, the differential accessibility or conformation of chromatin, which in turn may allosterically modulate the function of the transcription factor, or a combination of them. Although there are antecedents for the bifunctionality of transcription factors (Rao, 2001), a detailed biochemical analysis demonstrating this for REST/NRSF has not been communicated. Nevertheless, REST/NRSF can interact with the nuclear receptor corepressor (NCoR), and in turn recruitment of HDAC3 by NCoR has been associated to activation of transcription (Jepsen et al., 2000). Regarding the differences in expression patterns of possible components of complexes that include REST/NRSF, it must be noted that CoREST, described as a REST-associated corepressor, displays a predominantly neural expression pattern at very early stages of both murine and amphibian development (Grimes et al., 2000; De la Calle-Mustienes et al., 2002), although at later stages, non-neural expression is observed. This fact is compatible with the idea that REST-CoREST complexes may play a role in gene expression in the very early nervous system different from the role of repressors in non-neural cells at later stages.
The capability of REST/NRSF to ensemble diverse complexes able to modulate transcription may be used to restrict the expression of some genes to neurons, as well as to silence their non-neural expression. This general mechanism may account for the regulated expression of sodium channels and, thus, the onset of electrical excitability in developing neurons.
Footnotes
↵* R.A. and R.F. contributed equally to this work.
This work was supported by Fondo Nacional de Desarrollo Cientı́fico y Tecnológico 2980063, 4000031, 1961084, and 1000824; Fundación G. Puelma; fellowships from The Company of Biologists Ltd. and Fundación de Estudios Biomédicos Avanzados; and a graduate Comisión Nacional de Deserollo Cientı́fico y Tecnológico Scholarship (R.A.). We thank Dr. G. Mandel and J. Sierralta for continuous support, A. Figueroa and F. Vergara for excellent technical assistance, and Dr. M. E. Andrés for comments on this manuscript.
Correspondence should be addressed to Manuel Kukuljan, Programa de Fisiologı́a y Biofı́sica, Instituto de Ciencias Biomédicas, Facultad de Medicina, Universidad de Chile, Independencia 1027, Santiago, Chile. E-mail:kukuljan{at}neuro.med.uchile.cl.





{kind=link}
{kind=link}
{kind=link}