

Abstract
Neurons typically assume multipolar, bipolar, or unipolar morphologies. Little is known about the mechanisms underlying the development of these basic morphological types. Here, we show that the Krüppel-like transcription factor Dar1 determines the multipolar morphology of postmitotic neurons in Drosophila. Dar1 is specifically expressed in multipolar neurons and loss of dar1 gradually converts multipolar neurons into the bipolar or unipolar morphology without changing neuronal identity. Conversely, misexpression of Dar1 or its mammalian homolog in unipolar and bipolar neurons causes them to assume multipolar morphologies. Dar1 regulates the expression of several dynein genes and nuclear distribution protein C (nudC), which is an essential component of a specialized dynein complex that positions the nucleus in a cell. We further show that these genes are required for Dar1-induced multipolar neuron morphology. Dar1 likely functions as a terminal selector gene for the basic layout of neuron morphology by regulating both dendrite extension and the dendrite–nucleus coupling.
SIGNIFICANCE STATEMENT The three basic morphological types of neurons—unipolar, bipolar, and multipolar—are important for information processing and wiring of neural circuits. Little progress has been made toward understanding the molecular and cellular programs that generate these types since their discovery over a century ago. It is generally assumed that basic morphological types of neurons are determined by the number of dendrites growing out from the cell body. Here, we show that this model alone is insufficient. We introduce the positioning of nucleus as a critical factor in this process and report that the transcription factor Dar1 determines multipolar neuron morphology in postmitotic neurons by regulating genes involved in nuclear positioning.
Introduction
Ramon y Cajal placed neurons into three major morphological types based on the number of dendrites connected to the soma (i.e., primary dendrites): unipolar, bipolar, and multipolar and this classification system is universally applicable to different species throughout evolution (Cajal, 1995). Multipolar neurons, like mammalian pyramidal neurons, develop more than one primary dendrite. In contrast, bipolar neurons are defined as having a single primary dendrite that may (e.g., cerebellar Purkinje cells) or may not (e.g., photoreceptors) branch out into an elaborate dendritic arbor. Finally, unipolar neurons such as DRG neurons in vertebrates and the majority of CNS neurons in invertebrates extend a single primary neurite, which usually bifurcates into dendritic and axonal branches.
Multipolar morphology separates the dendritic arbor into distinct fields around the soma (Spruston, 2008), which has an impact, not only on the passive current spread and processing of electrical signals in the neuron (Rall, 1964), but also on the types of synaptic or sensory inputs that the neuron receives (Spruston, 2008). In addition, the three basic morphologies of neurons are relevant to the distinct organizational principles used in both the nervous systems of different animal species and in different parts of a single nervous system. Although all three morphological types are found in different species throughout evolution, the majority of neurons in invertebrates are unipolar, whereas the majority of those in vertebrates are multipolar (Strausfeld, 1976; Laurent, 1999; Grueber et al., 2005). In the insect CNS, unipolar neurons extend a single process from the soma to a synapse-enriched neuropil and then bifurcate into dendrites that arborize locally and an axon that typically projects to other neuropil areas or target tissues (Strausfeld, 1976). Unipolar organization of neuronal processes allows the formation of synaptic connections away from the location of the neuronal cell body, so it is likely an alternative strategy for neuronal migration, which is rare in the insect CNS (Harris, 2001) but common in the vertebrate CNS (Hatten, 1999).
Despite the importance of these fundamental organizations of neuronal processes, very little progress has been made toward understanding the molecular and cellular programs that lead postmitotic neurons to develop multipolar, bipolar, or unipolar morphologies since their description a century ago.
In this study, we show that the transcription factor Dar1 determines the multipolar morphology of postmitotic neurons in Drosophila. Dar1 is selectively expressed in postmitotic multipolar neurons and is required for these neurons to assume the multipolar morphology. Ectopic expression in unipolar or bipolar neurons leads to multipolar morphology. Dar1 regulates the expression of several dynein genes and nudC, which is an essential component of a specialized dynein complex that positions the nucleus in a cell. We further show that this evolutionarily conserved complex is required for multipolar morphology of neurons. These results suggest that dar1 likely functions as a terminal selector gene for the basic layout of neuron morphology.
Materials and Methods
Fly stocks and cDNA constructs.
Fly stocks include: dar13232, FRT2A (Ye et al., 2011); dar13010, FRT2A (Ye et al., 2011); CtC145, FRT19A (Grueber et al., 2003); hiwΔN, FRT19A (Wang et al., 2013); UAS-Dar1 (Ye et al., 2011); UAS-Nod::βGal (Clark et al., 1994), ppk-CD4::tdTomato (Han et al., 2011); RN2-Gal4 (Bloomington Stock Center #7475; Fujioka et al., 2003); PDF-Gal4 (Taghert et al., 2001); UAS-GluedDN (Allen et al., 1999); swts (Boylan and Hays, 2002); UAS-NudE (Arthur et al., 2015); nudE11R2, FRT2A (Hayashi et al., 2014); and UAS-monomeric RFP (Ye et al., 2007). The following RNAi lines were obtained from the Vienna Drosophila RNAi Center: v105898 (CG9492 RNAi), v23504 (CG8407 RNAi), v8737 (CG14763 RNAi), and v35154 (CG6971 RNAi). The following lines were obtained from the Bloomington Stock Center: NudE RNAi (BL41860), NudC RNAi (BL33382), and NudC PL00045, FRT2A. The DNA construct for making the UAS-KLF7 transgenic lines was generated by inserting the mouse KLF7 cDNA (tagged with a V5-epitope at the C terminus) into the pUAST vector.
MARCM and flip-out analyses.
The mosaic analysis with a repressible cell marker (MARCM) experiments were performed as described previously (Ye et al., 2011). For MARCM analysis of dar1 mutations in different classes of dendritic arborization (da) neurons and external sensory (es) neurons, the hs-FLP; Gal421-7, UAS-mCD8::GFP; tubP-Gal80, FRT2A virgins were mated with males of dar13232, FRT2A or dar13010, FRT2A. For MARCM analysis of dar1 mutations with the C4da neuropil marker ppk-CD4::tdTomato, the hs-FLP; ppk-Gal4, UAS-mCD8::GFP; tubP-Gal80, FRT2A virgins were mated with males of ppk-CD4::tdTomato; dar13232, FRT2A. For MARCM analysis of NudE overexpression in dar1−/− neurons, the hs-FLP; Gal421-7, UAS-mCD8::GFP; tubP-Gal80, FRT2A virgins were mated with males of UAS-NudE; dar13232, FRT2A or UAS-NudE; dar13010, FRT2A. For flip-out analysis of Dar1 overexpression in es neurons, the UAS-Dar1 virgins were mated with males of hs-FLP; Gal421-7; UAS-FRT-CD2stop-FRT-CD8::GFP.
Immunostaining and confocal imaging.
Third-instar larvae were dissected and immunostained as described previously (Ye et al., 2011). The primary antibodies include: mouse anti-GFP (Invitrogen, 1:1000), chick anti-GFP (Aves Laboratories, 1:1000), rabbit anti-RFP (Rockland, 1:1000), rat anti-Elav (Developmental Studies Hybridoma Bank, 1:500), guinea pig anti-Dar1 (Ye et al., 2011, 1:1000), guinea pig anti-Knot (gift from Adrian Moore, 1:1000), guinea pig anti-Spineless (Kim et al., 2006, 1:1000), rabbit anti-Abrupt (gift from Stephen Crews, 1:200), rat anti-Cut (Blochlinger et al., 1988, 1:1000), and mouse anti-βGAL (Developmental Studies Hybridoma Bank, 1:100). Confocal imaging was performed with a Leica TCS SP5 confocal microscope.
Microarray with purified Drosophila PNS neurons and microarray analysis.
PNS neurons were labeled with Gal421-7, UAS-mCD8::GFP and were purified with FACS as described previously (Ye et al., 2011). For each microarray sample (n = 3 for wild-type or dar1 mutant), total RNA was extracted from ∼20,000 GFP-positive cells using Trizol (Invitrogen), followed by purification with the RNeasy Micro Kit (Qiagen). cDNA was synthesized and amplified with WT-Ovation Pico RNA amplification System (NuGEN Technologies), followed by biotin labeling with the Encore Biotin Module (NuGEN Technologies). The biotin-labeled cDNA were hybridized to the Drosophila Genome 2.0 Array (Affymetrix).
To detect differentially expressed genes, Bayesian tests were used and implemented in the limma R package (Smyth, 2004). Genes with false discovery rate (FDR) < 0.05 were considered differentially expressed.
For functional annotation of differential expressed genes, we used the DAVID bioinformatics resource (da Huang et al., 2009). Cellular component gene ontology (GO) terms with Benjamini FDR < 0.05 were considered as significantly enriched.
qRT-PCR.
Total RNA was extracted from purified PNS neurons of embryos 16 h after egg-laying using Trizol (Invitrogen). First-strand cDNA was synthesized using SuperScript III First-Strand Synthesis SuperMix (Invitrogen). cDNA from RNA extracted from approximately 250 neurons was used for each real-time reaction. qRT-PCR and the quantification were performed as described previously (Kim et al., 2013).
Primer sets used were as following: chmp1, 5′-AAAGGCCAAGAAGGC GATTC-3′ and 5′-GGGCACTCATCCTGAGGTAGTT-3′; CG9492, 5′-AGATGGATGGACTTGTGCCT-3′ and 5′-CGATCACTCCGCAATATAAATCCT-3′; CG8407, 5′-GCCATTGAACTGTCCATTACC-3′, and 5′-ACCTCAAAGCCGAACCCT-3′; CG14763, 5′-ACAACTACGACAGATATCGCT-3′ and 5′-CTGTAGGTCACAAATCCGTC-3′; nudC set 1, 5′-GACGATGAATCCGATAAGAGTG-3′ and 5′-GAATCTTGAGCTCCACCTCC-3′; nudC set 2, 5′-AGGTGGAGCTCAAGATTCC-3′ and 5′-TCGATGATGGGCGTTTGAC-3′.
Quantifications and statistical analysis.
The number of primary dendrites was counted in 3D z-stacks. RN2+ neurons in which the primary dendrites overlapped with neurites of neighboring neurons were unquantifiable and were excluded. For pigment dispersing factor (PDF) neurons, neurites shorter than 10 μm were excluded. In all bar charts of quantification, values and error bars indicate mean ± SEM. Sample numbers are indicated either in the bars or in the legends. ANOVA was used for comparing three or more groups. Two-tailed unpaired Student's t test was used for comparing two groups. p values are indicated in the bar charts as follows: not significant (NS) p > 0.05, *p < 0.05, **p < 0.01, and ***p < 0.001.
Results
Dar1 is required for multipolar morphology in postmitotic neurons
The Drosophila nervous system offers a simple platform for studying the mechanisms that underlie the basic morphological types of neurons. In Drosophila, the CNS neurons are predominantly unipolar (Strausfeld, 1976; Laurent, 1999; Grueber et al., 2005), whereas the PNS neurons are either multipolar or bipolar (Orgogozo and Grueber, 2005). In the PNS, the da neurons are multipolar (Grueber et al., 2002), whereas the es and chordotonal (ch) neurons are bipolar (Orgogozo and Grueber, 2005).
We previously identified the dendritic arbor reduction 1 (dar1) gene, which encodes a member of the Krüppel-like factor (KLF) family of transcriptional regulators, as a specific regulator of dendritic, but not axonal, growth (Ye et al., 2011). In neurons that lack dar1 function, total length and branch number of dendrites in da neurons are reduced. Conversely, overexpressing Dar1 in these neurons increases dendrite length and branch number. Strikingly, in the embryonic and larval nervous system, Dar1 was exclusively expressed in multipolar neurons and was undetectable in bipolar and unipolar neurons (Ye et al., 2011; Fig. 1A,B). As a comparison, other transcription factors known to regulate dendritic development in multidendritic (md) neurons, such as Knot, Cut, Abrupt, and Spineless, are expressed either in a subset of or in all PNS and CNS neurons (Grueber et al., 2003; Sugimura et al., 2004; Kim et al., 2006; Hattori et al., 2007; Jinushi-Nakao et al., 2007; Crozatier and Vincent, 2008 and data not shown). The unique and selective expression of Dar1 in multipolar neurons led us to hypothesize that Dar1 determines the multipolar layout of neuronal dendrites.

Dar1 is required for multipolar neuronal morphology but not cell fates in postmitotic neurons. A, B, Dar1 is uniquely and selectively expressed in multipolar neurons in the Drosophila nervous system. A, Dar1 is exclusively expressed in multipolar neurons in the PNS. The schematic shows larval PNS neurons in the dorsal cluster, including multipolar neurons (green) and bipolar neurons (blue). The images show PNS neurons labeled with antibodies against the pan-neuronal membrane marker HRP (left), the neuron-specific marker Elav (middle), and Dar1 (right). Green arrows indicate multipolar neurons and blue arrows indicate bipolar es neurons. Scale bar, 10 μm. B, Dar1 is absent in the CNS. Shown are larval VNC double-labeled for the pan-neuronal protein Elav and Dar1. Scale bar, 100 μm. C–F, Loss of dar1 converts the multipolar da neurons into unipolar or bipolar morphology. C, Shown are wild-type (top) and dar13232 (middle and bottom) MARCM clones of the class I (C1) da neurons ddaE. Dar1 mutant ddaE neurons exhibit bipolar (middle) or unipolar (bottom) morphology. D, Wild-type (left) and dar13232 (right) MARCM clones of the class IV (C4) da neurons ddaC. Yellow solid triangles are primary dendrites; magenta solid triangles, axons; open triangle, the primary neurite in unipolar neurons. Scale bar, 20 μm. E, Bar chart showing quantification of the number of primary dendrites in wild-type, dar13232 and dar13010 MARCM multipolar (green) and bipolar (blue) neurons. F, Bar chart of the percentage of neurons with single dendrites in wild-type, dar13232, and dar13010 MARCM multipolar (green) and bipolar (blue) neurons. G, Loss of dar1 does not alter axon targeting of C4da neurons. The axon terminal of a representative dar13232 MARCM clone of the C4da ddaC neuron is shown. The axon terminal is labeled with CD8::GFP and the C4da neuropil is labeled with ppk-CD4-tdTomato. H, Dar1 is expressed postmitotically. Elav (green) and Dar1 (magenta) were stained in wild-type embryos. Although Elav is already expressed in peripheral neurons at stage 10 (top, left), Dar1 is only beginning to become visible at this stage (top, middle). By Stage 13, Dar1 is robustly expressed in identifiable da neurons (bottom, middle) that will henceforth become multipolar. I, Loss of dar1 does not change the class identity of da neurons. Knot and Cut expression in dar1−/− MARCM clones remain identical to wild-type. A single dar1−/− ddaC neuron is marked by GFP expression. In the PNS dorsal cluster, Knot is expressed in C4da (ddaC) neurons, whereas Cut is expressed in C3da (ddaF is shown) and C4da (ddaC) neurons (yellow arrows). Anti-HRP antibody labels all PNS neurons and was used to help the identification of neurons based on the position of cell bodies in the cluster. Scale bar, 10 μm.
To test this hypothesis, we first introduced dar1-null mutations into single da, es, and ch neurons using the MARCM technique (Lee and Luo, 1999; Fig. 1C,D). The wild-type class I-IV (C1–4) da neurons typically extend two or three primary dendrites from the soma. In sharp contrast, the majority of dar1 mutant neurons became bipolar with a single primary dendrite and an axon. Loss of dar1 even converted some C1da neurons into unipolar morphology (Fig. 1C), which was not observed in wild-type neurons. Consistent with the observation that Dar1 is absent in bipolar neurons (Fig. 1A; Ye et al., 2011), bipolar es neurons with dar1 mutations were unchanged, projecting a single primary dendrite, as was seen in wild-type es neurons (data not shown). Two different null alleles of dar1, dar13232 and dar13010, exhibited similar defects (Fig. 1E,F), suggesting that the loss of multipolar morphology in mutant neurons is caused specifically by loss of dar1.
Despite their dendritic defects, the axons of mutant neurons were correctly targeted to the C4da neuron-specific neuropil and formed normal terminal branches (Fig. 1G; Ye et al., 2011; Yang et al., 2014). Moreover, Dar1 is expressed in postmitotic neurons after the expression of the neuronal marker Elav started (Fig. 1H). Furthermore, neuronal type-specific markers, including Knot (Hattori et al., 2007; Jinushi-Nakao et al., 2007), Cut (Grueber et al., 2003), Abrupt (Sugimura et al., 2004), and Spineless (Kim et al., 2006), were expressed appropriately in dar1−/− da neurons (Fig. 1I and data not shown). Therefore, dar1 functions in postmitotic neurons and loss of dar1 does not change the type of neurons.
Together, these results demonstrate that Dar1 is required for the multipolar morphology in Drosophila neurons.
Misexpression of Dar1 in unipolar neurons induces multipolar morphology
Next, we investigated whether misexpressing Dar1 in unipolar neurons would cause them to assume a multipolar morphology. We overexpressed Dar1 in a small subset of unipolar motor neurons labeled by the RN2-Gal4 driver (RN2+ neurons; Fujioka et al., 2003; Ghabrial et al., 2003) and examined their morphology. Dar1-overexpressing RN2+ neurons extended ectopic neurites directly from the soma and consequently became multipolar (Fig. 2A,B). The ectopic primary neurites induced by Dar1 misexpression in these neurons were positive for Nod::βGal, a marker that labels subsets of dendritic branches (Zheng et al., 2008). Similarly, misexpression of Dar1 caused the unipolar PDF neurons in adult brain and the bipolar es neurons in larval PNS to assume multipolar morphology (Fig. 2C–E and data not shown). Moreover, overexpressing the mouse homolog of Dar1, KLF7, in larval RN2+ neurons and adult PDF neurons also led to a dramatic increase in the number of primary dendrites (Fig. 2B–E), suggesting an evolutionarily conserved function of Dar1 and KLF7.
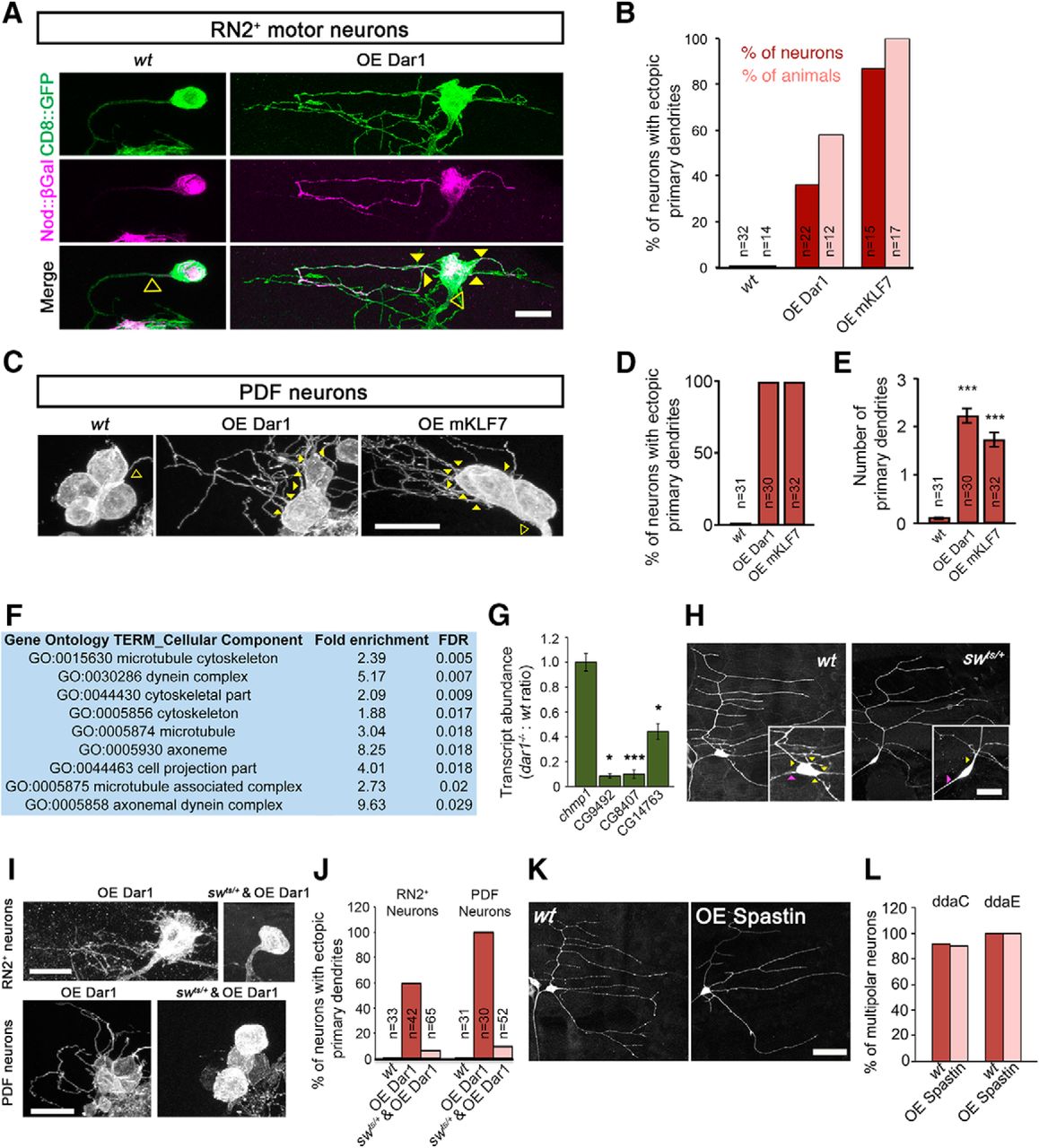
Dar1 requires the dynein complex to induce multipolar morphology of neurons. A, B, Misexpression of Dar1 or mKLF7 converts the unipolar RN2+ motor neurons in larva VNC into multipolar morphology. A, RN2+ neurons (Fujioka et al., 2003; Ghabrial et al., 2003) of wild-type and those overexpressing Dar1 (OE Dar1). Many of the ectopic primary neurites in RN2+ motor neurons are positive for Nod::βGal, a marker that labels subsets of dendritic branches (Zheng et al., 2008). Yellow solid triangles are primary dendrites; open triangles, the primary neurites in unipolar neurons. Scale bar, 10 μm. B, Bar chart showing the percentage of neurons and larvae with ectopic primary dendrites in RN2+ neurons of the indicated genotypes. C–E, Misexpression of Dar1 or mKLF7 converts the unipolar PDF neurons in adult brain into multipolar morphology. C, Representative images wild-type, Dar1-overexpressing, and mKLF7-overexpressing (OE mKLF7) PDF neurons. Yellow solid triangles are primary dendrites; open triangles, the primary neurites in unipolar neurons. Scale bar, 10 μm. D, E, Bar charts showing the percentage of neurons with ectopic primary dendrites and the number of primary dendrites in PDF neurons. F, G, Dar1 regulates the expression of dynein genes. F, Significantly enriched GO terms (cellular components) of which the FDR is <0.05 between wild-type and dar1 mutant microarrays. G, Bar chart of quantifications of relative transcript levels of chmp1 (control, n = 5), CG9492 (n = 4), CG8407 (n = 5), and CG14763 (n = 5) from wt and dar1 mutant purified PNS neurons measured by qRT-PCR. H, The dynein complex is required for multipolar morphology of Drosophila da neurons. Multipolar C1da neurons (ddaE) became bipolar in a temperature-sensitive mutant of the dynein intermediate chain, swts/+, at nonpermissive temperature (25°C). Yellow triangles are primary dendrites; magenta triangles, axons. Scale bar, 20 μm. I, J, Disrupting dynein function suppresses the appearance of ectopic primary dendrites induced by Dar1 misexpression. I, Defective dynein function caused by swts/+ (25°C) blocked the multipolar morphology induced by Dar1 overexpression in RN2+ and PDF neurons. Scale bar, 10 μm. J, Bar chart showing quantification of percentages of RN2+ or PDF neurons with ectopic primary dendrites denoted genotypes in the indicated genotypes. K, L, Roles of Dar1 in dendrite branching and in determining the multipolar layout of da neurons are mediated by distinct mechanisms. K, Wild-type and Spastin-overexpressing (OE Spastin) C1da (ddaE) neurons. Scale bar, 25 μm. L, Quantifications of the percentage of multipolar C4da (ddaC) and C1da (ddaE) neurons. p > 0.05, NS, t test.
dar1 requires dynein genes to determine multipolar morphology of neurons
How might Dar1 determine multipolar organizations of neuronal processes? To answer this question, we first conducted microarray analysis to identify genes with expression that was regulated by Dar1. Transcripts were extracted from wild-type and dar1−/− PNS neurons purified with FACS and used for microarray analysis. GO-based functional annotation (da Huang et al., 2009) of the differentially expressed genes showed that cellular components that were significantly enriched in dar1−/− neurons are mostly related to microtubules (Fig. 2F). Strikingly, the transcript levels of several genes in the dynein complex were consistently reduced in dar1 mutant neurons, which was confirmed by qRT-PCR (Fig. 2G). We thus examined the role of the dynein complex in forming the multiple morphology of neurons by suppressing dynein functions with a temperature-sensitive mutant of the dynein intermediate chain short wing (sw), swts (Boylan and Hays, 2002). Rearing swts/+ larvae at nonpermissive temperatures led to the appearance of bipolar-shaped C1da neurons (Fig. 2H), which are virtually absent in wild-type controls, without altering the number of dendritic branches (data not shown).
Next, we tested whether Dar1 requires the dynein complex to induce primary dendrite formation by introducing a heterozygous swts mutation in RN2+ and PDF neurons that misexpressed Dar1. In animals raised at a nonpermissive temperature, the swts mutation potently reduced the supernumerary primary dendrites induced by Dar1 misexpression (Fig. 2I,J), suggesting that dar1 requires the dynein complex to induce multipolar morphology of neurons.
We showed previously that Dar1 regulates dendrite branching by suppressing the expression of Spastin (Ye et al., 2011). We thus examined the possible role of Spastin in determining the basic layout of neuronal morphology. Although overexpressing Spastin reduced dendritic growth as previously reported, it did not alter the multipolar morphological type of da neurons (Fig. 2K,L). These results suggest that the roles of Dar1 in dendrite branching and in determining the multipolar layout of da neurons are mediated by distinct mechanisms. Consistent with this notion, whereas dendritic branching was reduced in neurons lacking the transcription factor Cut (Grueber et al., 2003) or the E3 ubiquitin ligase Highwire (Wang et al., 2013), these neurons still formed the normal number of primary dendrites despite having simplified higher-order branches (data not shown). Therefore, factors regulating dendritic growth or branching are not necessarily determinants of the multipolar morphology.
Nuclear positioning complex is required for the multipolar morphology of neurons
Loss of dar1 gradually converted multipolar neurons into bipolar or unipolar morphologies. At early stages of development, dar1 mutant neurons are multipolar despite appreciable defects in dendritic growth (Fig. 3A). These multipolar neurons gradually became bipolar, whereas the overall pattern of the dendritic arbors remained unaffected (Fig. 3A,B), indicating that loss of dar1 led to defective coupling between the primary dendrites and the cell nucleus.

Nuclear positioning complex is required for multipolar morphology of neurons. A, B, Loss of dar1 causes multipolar neurons to gradually become bipolar during development. A, Representative images of C4da (ddaC) neurons at different stages of larval development. At 24 h after egg laying, all dar1 mutant ddaCs are multipolar, although dendritic growth defects are already visible. At 96 h after egg laying, many multipolar ddaCs become bipolar, whereas the overall dendritic pattern is maintained. B, Line plot showing the percentage of multipolar ddaCs that are wild-type or dar1 mutant at different developmental times. C, Quantification of relative transcript levels of chmp1 and nudC from purified wt and dar1−/− PNS neurons. qRT-PCR with two distinct sets of nudC primers showed consistent reduction in PCR products in the cDNAs from dar13232 da neurons. D–F, Knocking down the Drosophila homologs of NudE and NudC caused C1da neurons (ddaE) to assume bipolar morphology. D, Cell bodies (arrows) of some NudE or NudC knockdown neurons are connected with the dendritic arbors by a single primary dendrite, which does not occur in normal ddaE neurons. The first branch point of the dendritic arbor is indicated by a triangle. The inset in each panel shows the magnified view of cell body containing the nucleus. The neurons were labeled by the membrane marker mCD8-GFP, which is absent in the cell nucleus. Scale bar, 50 μm. E, Bar chart showing the percentage of neurons with single primary dendrites of denoted genotypes. F, Bar chart showing that the number of dendrite termini is unaffected in NudE or NudC knockdown neurons.
Previous studies have demonstrated that a specialized dynein complex is important for positioning the nucleus in the cell. The genes encoding the factors in the nuclear positioning complex, termed nuclear distribution (nud) genes, were first identified in filamentous fungi and later found to be well conserved evolutionarily (Morris, 2000). For example, in mammals, the NudF homolog called LIS1 and the NudE homolog called NudE-like protein (or NUDEL) are required for coupling the leading process of the migrating neurons with its centrosome and nucleus (Reiner et al., 1993; Niethammer et al., 2000; Shu et al., 2004; Tsai et al., 2005; Ayala et al., 2007; Tsai et al., 2007; Zhang et al., 2009; Hippenmeyer et al., 2010). Interestingly, our microarray analysis also showed a significant reduction in the levels of the nudC gene, which encodes an essential component of the nuclear positioning complex (Morris, 2000; data not shown). qRT-PCR further confirmed that nudC transcripts were dramatically reduced in dar1−/− da neurons (Fig. 3C). We thus examined the role of the nuclear positioning complex in the multipolar layout of neuronal morphology.
Knocking down the Drosophila homolog of NudC, NudE, or introducing mutations in nudC in da neurons by MARCM transformed the multipolar da neurons into bipolar morphology (Fig. 3D,E). Strikingly, whereas the overall morphology and branch numbers of the dendritic arbors were indistinguishable between the mutant and wild-type neurons (Fig. 3F), the nuclei of mutant neurons were several micrometers distant from the dendritic arbors (Fig. 3D). These results suggest that regulators of nuclear positioning are essential for the multipolar layout of neuronal dendrites in Drosophila.
To determine whether regulators of nuclear positioning are essential for mediating Dar1's function in determining the multipolar morphology of neurons, we performed epistasis analysis between dar1 and nudE. NudE overexpression in dar1−/− da neurons partially rescued the increased number of bipolar da neurons caused by loss of dar1 (Fig. 4A,B). Moreover, significantly fewer Dar1-overexpressing RN2+ and PDF neurons displayed multipolar morphology when nudE (nudE11R2/+) or nudC (NudC RNAi) function is reduced (Fig. 4C–F). These results suggest that Dar1 likely acts via the nuclear positioning complex to induce multipolar morphology of neurons.

Dar1 requires nuclear positioning complex to induce multipolar morphology of neurons. A, B, NudE overexpression reduces the percentage of dar1 mutant C4da neurons that assume the bipolar morphology. Results of wild-type, dar3232, and dar3010 neurons in B are the same as those in Figure 1F. The solid triangles indicate primary dendrites and open triangles indicate axons. Scale bar, 10 μm. C, D, Dar1 requires nudE to induce multipolar morphology in adult PDF neurons. C, Representative images of PDF neurons that are wild-type, Dar1-overexpression, nudE11R2/+, and nudE11R2/+ with Dar1 overexpression. Solid triangles indicate primary dendrites and open triangles indicate the primary neurites in unipolar neurons. Scale bar, 10 μm. D, Bar chart showing the percentage of neurons with ectopic primary dendrites in PDF neurons of denoted genotypes. E, F, Dar1 requires nudE to induce multipolar morphology in larval RN2+ motor neurons. E, Representative images of RN2+ neurons that are wild-type, Dar1-overexpression, nudE11R2/+, nudE11R2/+ with Dar1-overexpression, RFP-overexpression, and NudC RNAi with Dar1-overexpression. Solid triangles indicate primary dendrites and open triangles indicate the primary neurites in unipolar neurons. Scale bar, 10 μm. F, Bar chart showing the percentage of neurons with ectopic primary dendrites in RN2+ neurons of denoted genotypes. G, Model explaining the roles of Dar1 in determining multipolar neuron morphology. The nuclear positioning complex couples the nucleus with primary dendrites. An opposing force may exist to pull the nucleus toward the axon. Top, When the coupling between nucleus and primary dendrites is weakened, as in the case of loss of dar1 or loss of nuclear positioning complex components, the nucleus is pulled toward the axon, changing the morphology from multipolar to bipolar. Bottom, When Dar1 is misexpressed in a unipolar neuron, it promotes both dendritic extension from the cell body and coupling between the primary dendrites and the nucleus, leading to multipolar morphology.
Discussion
The universal morphological organization of neuronal dendrites and axons—in the form of the unipolar, bipolar, and multipolar morphologies—is important for information processing in neurons and for the wiring of neural circuits. It has generally been assumed that the formation of the basic morphological types of neurons is determined by the number of dendrites growing out from the cell body (the “outgrowth model”). In this study, we show that this model alone is insufficient to explain the formation of multipolar morphology. We introduce nuclear positioning as a factor in determining the multipolar neuron morphology (Fig. 4G) and propose that Dar1 determines multipolar morphology by regulating both dendrite extension and primary dendrite–nucleus coupling.
Neuronal fate versus neuronal morphogenesis
We report a novel, instructive role for Dar1 in determining the multipolar morphology of postmitotic neurons without changing cell fate. First, despite the dendritic defects, axon morphology (Ye et al., 2011) and targeting (Fig. 1G) are unchanged in dar1 mutant neurons. Second, ectopic expression of Dar1 in postmitotic neurons leads to supernumerary primary dendrites (Fig. 2A–E). Third, the remaining dendrites in dar1−/− neurons still follow the branching pattern assumed by wild-type neurons (Ye et al., 2011; Fig. 1C,D). Fourth, dar1 mutations do not affect the expression of neuron type-specific markers (Fig. 1I and data not shown).
Based on extensive studies in C. elegans, Hobert proposed the concept of terminal selector genes (Hobert, 2008). A terminal selector gene is required for determining specific aspects of a neuron's identity by regulating the expression of genes responsible for these characteristics such as those encoding neurotransmitter receptors, enzymes in a neurotransmitter synthesis pathway, and structural proteins. Loss of a terminal selector gene results in the loss of a specific aspect of the neuron type without affecting the overall neuronal identity. Dar1 plays such a function in the basic layout of neuronal morphology and thus is likely a “terminal selector gene” for neuronal morphology.
Role of dendrite outgrowth and dendrite–nucleus coupling in generating basic morphological types of neurons
Based on the findings in this study, we propose that generating neuronal multipolar morphology requires, not only dendritic extension, but also a coupling mechanism between the nucleus and the dendrites (Fig. 4G). Dar1 promotes both dendritic growth and dendrite–nucleus coupling. Therefore, its misexpression converts unipolar neurons into neurons with multipolar morphology (Fig. 2A–E).
The results presented here raise the interesting possibility that a specialized dynein complex in multipolar neurons with components that are transcriptionally regulated by Dar1 couples the nucleus with the primary dendrites. If the primary dendrite–nucleus coupling is weakened, then the nucleus may move to a different location in relation to the dendrites and axons. We speculate that there might also be an active or passive force that pulls the nucleus toward the axon, opposing the force that couples the primary dendrites and the nucleus. Consistent with this model, the remaining single primary dendrites of all bipolar-shaped da neurons—caused by loss of dar1, reduced functions of dynein, or nuclear positioning complex—are those that project in the direction opposite the axon. This observation again rules out the possibility that the reduction in number of primary dendrites of dar1−/− da neurons is the result of reduction in dendrite growth. If that were the case, then the remaining single primary dendrites would likely project in random directions and not solely away from the axon. Further studies are needed to determine the cellular and molecular basis of the dendrite–nucleus coupling.
Several prior studies have demonstrated that neurons switch between different morphological types during development (Altman, 1972; Cajal, 1995; Nadarajah et al., 2001; Sotelo and Dusart, 2009). These observations suggest that the acquisition of basic morphological types in many neurons includes intermediate morphologies with nucleus–primary dendrite relationships that are different from those seen in the mature neurons. It will be interesting to investigate whether the activity of the nuclear positioning complex and its regulators play a role in these developmental changes in morphological type.
In summary, this study offers a novel model for understanding the establishment of the three basic morphological types of neurons. Starting from genetic analysis of the KLF transcription factor Dar1, we not only uncover an instructive factor that determines the multipolar morphology of neurons, but also provide a mechanistic model showing that the position of the nucleus is critical for establishing multipolar neuron morphology. This study also demonstrates, for the first time to our knowledge, that the basic morphological types are determined by intrinsic molecular mechanisms in postmitotic neurons rather than in precursor cells. The model presented here may also be applied to explaining the changes in basic morphological type during neuron development (Altman, 1972; Matsuda et al., 1996). This study therefore opens the door for a unifying theory of basic structural organization in neurons.
Footnotes
This work was supported by the National Institutes of Health (Grant R01MH091186 to B.Y., Grant T32-GM007315 to M.W.Z., and Grant T32-NS076401 to G.S.) and the Pew Scholars Program in the Biological Sciences (B.Y.) and used the resources of the Drosophila Aging Core of the Nathan Shock Center of Excellence in the Biology of Aging funded by the National Institute of Aging–National Institutes of Health (Grant P30-AG-013283). We thank Sige Zou and Melih Acar for advice on the microarray analysis; Liqun Luo for his support during the revision of this paper; Adrian Moore, Jill Wildonger, and Stephen Crews for generously sharing reagents; and Yukiko Yamashita, Catherine Collins, Kenneth Kwan, Cheng-Yu Lee, and Hisashi Umemori for critical comments on earlier versions of the manuscript.
The authors declare no competing financial interests.
- Correspondence should be addressed to Bing Ye, Life Sciences Institute and Department of Cell and Developmental Biology, University of Michigan, 210 Washtenaw Avenue, Room 5183A, Ann Arbor, MI 48109. bingye{at}umich.edu





{kind=link}
{kind=link}
{kind=link}
{kind=link}