

Abstract
Animals select their optimal environmental temperature, even when faced with alternatives that differ only slightly. This behavior is critical as small differences in temperature of only several degrees can have a profound effect on the survival and rate of development of poikilothermic animals, such as the fruit fly. Here, we demonstrate that Drosophila larvae choose their preferred temperature of 17.5°C over slightly cooler temperatures (14−16°C) through activation of chordotonal neurons. Mutations affecting a transient receptor potential (TRP) vanilloid channel, Inactive (Iav), which is expressed specifically in chordotonal neurons, eliminated the ability to choose 17.5°C over 14−16°C. The impairment in selecting 17.5°C resulted from absence of an avoidance response, which is normally mediated by an increase in turns at the lower temperatures. We conclude that the decision to select the preferred over slightly cooler temperatures requires iav and is achieved by activating chordotonal neurons, which in turn induces repulsive behaviors, due to an increase in high angle turns.
Introduction
Animals are capable of discerning small differences above and below their preferred ambient temperature, and this ability is especially important for organisms, such as insects, whose internal body temperature rapidly equilibrates with the environmental temperature. However, the molecular and cellular bases by which animals survey their thermal landscapes and decide on how to respond behaviorally to different temperature alternatives are incompletely understood.
A key class of temperature sensors is an evolutionarily conserved set of cation channels, referred to as transient receptor potential (TRP) channels (Venkatachalam and Montell, 2007). These proteins are global mediators of sensory input and thereby control a variety of animal behaviors. In Drosophila three TRP channels participate in the responses to warm or hot temperatures in the noxious range (Tracey et al., 2003; Lee et al., 2005; Rosenzweig et al., 2005; Hamada et al., 2008). We have shown recently that Drosophila larvae are sensitive to small deviations from their preferred temperature of ∼17.5−18°C, and TRPA1 is critical for enabling larvae to select these optimal temperatures over slightly warmer temperatures (≤24°C) (Kwon et al., 2008). In addition, two TRPC channels, TRP and TRPL have been reported to be involved in choosing the preferred temperature over slightly cooler temperatures (Rosenzweig et al., 2008).
Here, we conducted a comprehensive screen for Drosophila TRP channels that enable larvae to choose their optimal temperature of 17.5°C over mildly cooler temperatures (14−16°C). We found that a TRP vanilloid channel, Iav, was required in chordotonal neurons for selecting 17.5 over 14−16°C. Moreover, both Iav and the chordotonal neurons were required for selecting the optimal temperatures by inducing the larvae to increase turning angles, thereby avoiding the cooler temperatures.
Materials and Methods
Preparing third instar larvae for thermotaxis assay.
We prepared larvae for the thermotaxis assay as described previously (Kwon et al., 2008): http://www.natureprotocols.com/2008/07/28/assaying_thermotaxis_behavior.php. Briefly, the flies and larvae were reared at 25°C and were not exposed to CO2 at any time. We removed the third instar larvae from the food and washed them with 15% sucrose to remove remaining food, and then with H2O to remove the sucrose. We allowed the larvae to recover for 30 min at room temperature (∼22°C) before assaying thermotactic behavior. We applied moisture to avoid desiccation.
Thermotactic behavioral assay.
We analyzed thermotactic behavior as described previously (Kwon et al., 2008): http://www.natureprotocols.com/2008/07/28/assaying_thermotaxis_behavior.php. Briefly, we added 40–100 larvae (∼95% third instar) to the test plates. Unless specified otherwise, we placed the larvae along the middle of the plates containing 2% agarose. The experiments were conducted under ambient light. After 15 min, we photographed the plates and calculated the preference index (P.I.): P.I. = {(number of larvae on 17.5°C side) − (number of larvae on the other side with the variable temperature)} ÷ (total number of larvae on each side of the test plate).
Light–dark choice assays using animals expressing channelrhodopsin 2.
We reared the larvae on standard food at 25°C with or without 50 μm all-trans-retinal (Sigma, R2500). We avoided exposing the larvae to light by wrapping vials with foil. To conduct the light–dark choice assays, we prepared the third instar larvae as described above and as detailed previously (Kwon et al., 2008): http://www.natureprotocols.com/2008/07/28/assaying_thermotaxis_behavior.php, except that we collected and washed the larvae under dim light to minimize light exposure. Ten to 15 larvae were placed in the middle of a 3.5 cm cell culture dish covered with 2 ml of 2% agarose (see Fig. 4A,B). To avoid excessive heat, we illuminated half the dish with 20 pulses of light (5 s each; 0.03 mW/mm2), interspersed by 2.5 s dark intervals. The light was a 100 W quartz tungsten halogen bulb housed in an Oriel Apex QTH projector (Newport). The programming was performed using LabChart software in conjunction with PowerLab 4/30 (ADInstruments). The larvae on the light and dark sides were counted and the P.I. values calculated: {(number of larvae on dark side) − (number of larvae on light side)} ÷ (total number of larvae on each side of the test plate). The larvae were prepared and the assays were performed at room temperature (∼22°C).
Constructs for generating transgenic flies.
To generate UAS-iav flies, we amplified the coding region of iav by RT-PCR and subcloned the cDNA between the NotI and XbaI sites of pUAST. To generate the iav-GAL4 transgenic line, we excised the GAL4 sequence and the hsp70 3′ UTR from pGATB (Brand and Perrimon, 1993) using BamHI and SpeI and subcloned the DNA between the BamHI and XbaI sites of pCasper4-GAL4. We amplified ∼3 kb flanking the 5′ end of iav, which we subcloned between the NotI and BamHI sites of pCasper4-GAL4.
Movement trace analysis.
To determine the turning angles (θ) and the total vectoral movement, we tracked movement on temperature preference test plates using a digital camera. We processed the movies using QuickTime Pro 7. The intervals between consecutive frames were 5.1 s. We manually tracked the position of the individual larval head region between the end of the first and tenth frames (45.9 s) using NIH ImageJ software and MTrackJ plugin (Rasband WS, ImageJ, U.S. National Institutes of Health, Bethesda, MD, http://rsb.info.nih.gov/ij/, 1997–2009). The larvae remained on the test plates 3 min before initiating tracking. We discarded the trace if movement was disturbed by the test plate border or by interactions with other larvae. We calculated each θ by establishing the vectoral direction (by comparing larval head positions) between the beginning of two consecutive frames [P(N−1) and P(N) and P(N+1)] (see Fig. 4F) and the beginning of the P(N) and P(N+1) frames. We analyzed 88 individual directional deviations each.
To determine the movement speed (mm/s), we used 11 traces. We first determined the total distance traveled (mm) by adding together the individual connections between 10 consecutive frames. We divided the total distance traveled by the seconds (45.9) between the end of the first and 10th frames. The vectoral travel distance was established by connecting the positions at the end of the first and 10th frames in a trace with a single line.
Fly stocks.
The wild-type strain was Canton S. We obtained the following flies from the Bloomington Stock Center (stock numbers): iav1 (6029), trplMB03075 (23512) UAS-ChR2 (9681) (Schroll et al., 2006) btvBG01771 (12589) and Df(1)pod1Δ17 (8749). We obtained nan36a from C. Kim, Chonnam National University, Gwangju, Korea, pyx3 from J. Kim, Korea Advanced Institute of Science and Technology, Daejeon, Korea (Lee et al., 2005), pain2 from W. D. Tracey, Duke University Medical Center, Durham, NC (Tracey et al., 2003), the trpA1-GAL4 line from P. A. Garrity, Brandeis University, Waltham, MA (Rosenzweig et al., 2005), iav1;P[iav+] from J. Hirsh, University of Virginia, Charlottesville, VA, and atoW from B. A. Hassan, VIB, Katholieke Universiteit, Leuven, Belgium (Choi et al., 2009). The following trp stocks were described previously: trpP343 (Wang et al., 2005), trp47 (Wang et al., 2005) trp302 (Niemeyer et al., 1996), trpA11 (Kwon et al., 2008), pyx3 (Lee et al., 2005), pain2 (Tracey et al., 2003), iav1 (Gong et al., 2004), iav3621 (Gong et al., 2004), nan36a (Kim et al., 2003), trpml1 (Venkatachalam et al., 2008), amo1 (Watnick et al., 2003), nompC2, nompC3 (Walker et al., 2000) and trpm1 (Hofmann et al., 2010). The unpublished trpγ1 and wtrwex mutants were provided by Q. Ren and X. Wang, respectively, Johns Hopkins University School of Medicine, Baltimore, MD.
Statistics.
For single pair comparisons, we used the unpaired Student's t test in Excel 2007 (Microsoft). For multiple comparisons between wild-type and test groups, we used one-way ANOVA with the Dunnett's method using the Minitab 15 (Microsoft) default parameters (one-way ANOVA unstacked, family error rate: 5). For other multiple comparisons, we used one-way ANOVA with the Tukey or Tukey–Kramer test (with unequal sample numbers) in Origin 7.0 (OriginLab).
Results
Requirement for a TRPV channel, IAV, for selecting the optimal temperature over slightly cool but not cold temperatures
To systematically address the requirements for TRP channels for selecting 17.5°C over mildly cooler temperatures, we examined mutations affecting each of the 13 Drosophila TRP channels by performing two-way choice tests. We released larvae at the center of plates between 14° and 17.5°C zones and allowed them to choose between the two temperatures for 15 min (Fig. 1A). If all larvae migrated to the 17.5°C side, the P.I. would be 1.0, while a complete preference for 14°C would result in a P.I. of −1.0. The P.I. would be 0 if there were an equal number of larvae on both sides of the test plate.

Requirement for iav for discriminating 17.5°C from modestly cooler temperatures. A, Schematic of the test plate. The formula for the P.I. is shown. B, Survey of TRPs functioning in the discrimination between 17.5° and 14°C. Preferences for 17.5° and 14°C would result in positive and negative P.I. values, respectively. If there were no preference, the P.I. would be 0. C, Selection of 17.5°C required trpl. D, Thermotaxis did not require trp. E, Comparison of movement speeds. The movement speeds (mm/s) were calculated by dividing the total distance traveled (mm) over 9 video frames (45.9 s). F, The 17.5° versus 14°C P.I. values after releasing the larvae in the indicated zones (A, B, or C; see A). G, Characterization of 17.5° versus 14°C preferences of iav alleles and iav1 in trans with a deficiency (Df) chromosome (Bloomington stock 8749). P[iav+] is the wild-type iav genomic rescue transgene. The asterisk indicates significant differences (p < 0.05) between the indicated pairs of data. If there are no brackets, the asterisk indicates significant differences with wild type.
Elimination of any of the four TRPA channels, TRPM, TRPP (AMO), TRPN (NOMPC) or TRPML did not reduce the preference for 17.5°C (Fig. 1B). Mutation of the gene encoding the third TRPC channel, TRPγ, caused a slight, but statistically insignificant decrease in the P.I. (Fig. 1B). As reported previously, disruption of trpl abolished the preference for 17.5°C (Rosenzweig et al., 2008) (Fig. 1B,C). However, the temperature preferences of two strong trp alleles, trpP343 and trp47, were indistinguishable from wild-type larvae (Fig. 1B,D). Nevertheless, the previous finding that trp contributed to cool sensation was not definitive since thermotaxis was normal in larvae harboring the trp mutation in trans with a deficiency that removed the trp locus (Rosenzweig et al., 2008).
We found that mutations that disrupted either of the TRPV genes, iav1 or nan36a, eliminated the ability of the larvae to discriminate between 14° and 17.5°C (Fig. 1B). These results were surprising since mammalian TRPM and TRPA channels are known or implicated to function in the sensation of low temperatures, while TRPV channels participate in the responses to modestly warm temperatures, or to noxious heat (Venkatachalam and Montell, 2007).
Since iav and nan mutants are sedentary, the impairments in thermotaxis could be due to the reduced movements rather than to defects in temperature sensation. Indeed, both iav1 and nan36a moved more slowly than wild-type larvae at either 14° or 17.5°C (Fig. 1E). Therefore, we tested the effects on the P.I. after changing the release zone on the temperature selection test plate. Instead of placing the larvae in the middle of the test plate, we introduced the larvae at the extreme ends of the 14°C or the 17.5°C sides (B and C sites, Fig. 1A). Wild-type larvae showed a preference for 17.5°C, regardless of the release site, although the P.I. was reduced slightly if they were placed at the end of the 14°C side, presumably due to slower migration at this temperature (Fig. 1A,F). However, the P.I. of nan36a larvae was strongly dependent on their initial placement site on the test plate. When we introduced nan36a larvae at the end of the 14° or 17.5°C sides, most of larvae stayed in the 14° or 17.5°C zones respectively (Fig. 1F). Consequently, we could not discern whether or not nan was impaired in the selection of 17.5° over 14°C. Contrary to these results, the iav1 larvae were randomly distributed on the test plates (P.I. values near 0), regardless of where they were placed on the test plates (Fig. 1A,F). These findings indicate that iav larvae were impaired in the discrimination of the optimal temperature over 14°C.
To provide further evidence that iav was required for choosing 17.5° over 14°C, we tested another iav allele (iav3621). In addition, we analyzed both iav1 and iav3621 in trans with a deficiency (Df) that spanned the iav locus. We found that iav3621, iav1/Df and iav3621/Df larvae all showed the same inability to select 17.5° over 14°C (Fig. 1G). Furthermore, introduction of a wild-type genomic transgene (P[iav+]) rescued the thermotaxis defect in the iav1 mutant (Fig. 1G). The combination of these data demonstrated that iav was required for thermotaxis in the cool range.
In addition to selecting 17.5° over 14°C, wild-type larvae chose 17.5°C over other cool temperatures, at least down to 12°C (Fig. 2A,B). Discrimination of temperatures lower than 12°C was difficult to assess due to diminishing larval migration at lower temperatures (data not shown). Therefore, we tested whether iav was required for selecting 17.5°C over 12° and 16°C. We found that the ability of iav1 to choose 17.5° over 16°C was also eliminated, as well as the discrimination between two cool temperatures (14.5° vs 16°C; Fig. 2A). However, the mutant animals exhibited a normal preference for 17.5° over 12°C (Fig. 2B). As we have shown recently, iav1 larvae displayed normal selection of 17.5°C over comfortably warm temperatures, such as 22° and 24°C (Fig. 2C) (Kwon et al., 2008). These latter results were not surprising given the requirement for trpA1 for thermotactic behavior in this temperature range (Fig. 2C) (Kwon et al., 2008). Thus, iav was required for larvae to sense the preferred temperature, 17.5°C, over moderately cool temperatures (14− 16°C), but was dispensable for choosing 17.5°C over cooler (12°C) or comfortably warm temperatures.

iav is not required for sensing cold and warm temperatures. A, iav1 is defective in choosing 17.5° over 14−16°C. B, iav and the chordotonal organs are not required for discrimination of 17.5° over 12°C. C, iav and the chordotonal neurons are not required for selecting 17.5°C over comfortably warm temperatures. Asterisks indicate significant differences (p < 0.05) with wild type.
Larval chordotonal neurons function in temperature discrimination in the cool but not the cold range
Several observations indicated that chordotonal neurons were the cells required for discriminating 17.5°C from slightly cooler temperatures (14−16°C), but not other temperatures. First, we generated an iav-GAL4 line, which drove UAS-mCD8-GFP expression exclusively in chordotonal neurons (Fig. 3A), consistent with a previous report that expression of iav is specific to chordotonal neurons (Gong et al., 2004). To test whether the chordotonal neurons were critical for cool sensation, we inhibited synaptic transmission in these cells by combining the iav-GAL4 with UAS-TNT (tetanus toxin), and found that it prevented temperature discrimination between 14° and 17.5°C (Fig. 3B). However, there was no impairment on selection of 17.5°C over other temperatures such as 12° or 24°C (Fig. 2B,C). Expression of the inactive form of tetanus toxin had no effect on 17.5° versus 14°C selection (UAS-IMPTNT; Fig. 3B). Second, the combination of UAS-iav and iav-GAL4 was sufficient to rescue the iav phenotype (Fig. 3B). Third, we found that expression of UAS-iav under control of either of two additional chordotonal GAL4 lines (nan-GAL4 and pain-GAL4) (Tracey et al., 2003; Gong et al., 2004) rescued the deficit in cool temperature discrimination in iav1 larvae (Fig. 3B). Fourth, mutations (atoW and btvBG01771) that impaired the normal development of chordotonal neurons caused a defect in discriminating 14° and 17.5°C (Fig. 3B). Reintroduction of UAS-iav using the only available terminal organ GAL4 (GH86-GAL4) did not rescue the iav1 phenotype, although this result was not reliable since we found that expression of the GH86-GAL4 alone impaired temperature selection (data not shown). Nevertheless, the four lines of evidence presented here support the conclusion that the chordotonal neurons are critical for selection of the optimal over cool temperatures.
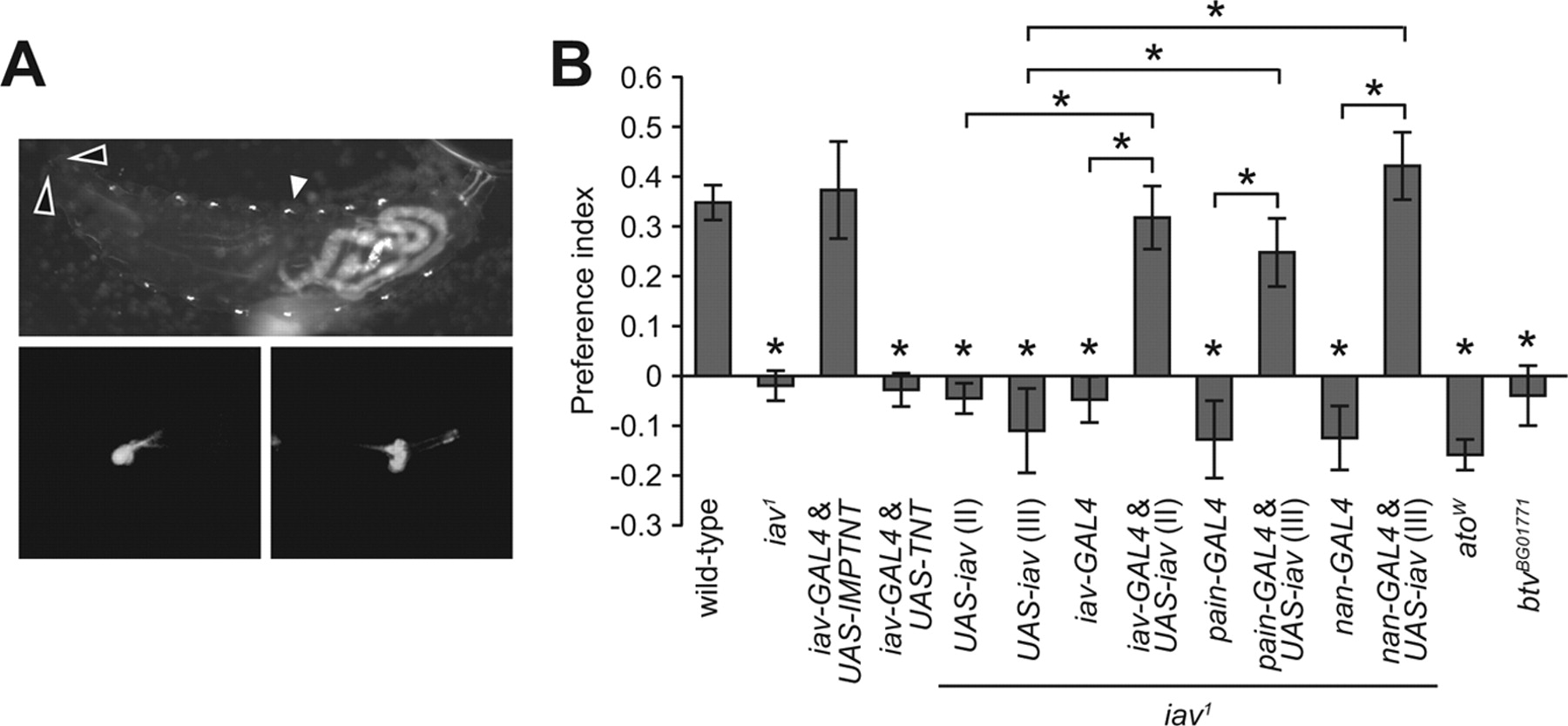
Requirement for chordotonal neurons for cool (14−16°C) avoidance behavior. A, iav reporter expression in larvae. The top shows GFP expression in chordotonal neurons (white arrowhead) in first instar larva using the iav-GAL4 and UAS-mCD8-GFP transgenes. The open arrowheads indicate the position of the terminal organs, which are not labeled. Anterior is to the left. The gut and trachea at the posterior end showed autofluorescence. The bottom shows higher-resolution images of iav reporter expression in the chordotonal neurons of the third instar larva. B, iav expression in chordotonal neurons required for cool temperature avoidance. Expression of UAS-TNT using the iav-GAL4-impaired 17.5° versus 14°C selection. The impairment in selecting 17.5° over 14°C in iav1 was rescued by expressing UAS-iav in chordotonal neurons. atow and btvBG01771 showed defects in selection of 17.5° over 14°C. Asterisks indicate significant differences (p < 0.05) with wild type.
Chordotonal neurons and iav contribute to thermal avoidance and increased turning behavior at cool temperatures
There are at least two mechanisms that potentially underlie the iav-dependent temperature preference behavior. The chordotonal neurons and iav could be required for attraction to the preferred temperature (17.5−18°C) or for avoidance of modestly cool temperatures (14−16°C). To distinguish between these possibilities, we attempted to stimulate iav-expressing neurons, independent of changes in temperatures, and examined whether this induced attractive or avoidance behavior. To activate iav-expressing cells, we expressed channelrhodopsin 2 (UAS-ChR2) (Schroll et al., 2006) under the control of the iav-GAL4. ChR2 is a blue light-activated cation channel, so stimulation with blue but not red light leads to depolarization of neurons. Wild-type larvae express rhodopsins (Sprecher et al., 2007), which are classical G-protein coupled receptors. Consequently, in the absence of ChR2, wild-type larvae display an aversive response to light (Hassan et al., 2000). To eliminate the endogenous light response, we tested the effects of introducing the iav-GAL4 and UAS-ChR2 in a norpAP24 background, which disrupts a phospholipase C critical for the larval photoresponse (Hassan et al., 2000).
To determine the behavioral consequences resulting from stimulating iav-expressing neurons with ChR2, we placed the larvae in the middle of a Petri dish, half of which was kept in the dark, and the other half was exposed to light (Fig. 4A–D). Wild-type larvae avoided blue light, since they express rhodopsins that absorb blue light, including Rh5 (Fig. 4E) (Sprecher et al., 2007). However, norpAP24 larvae did not show a preference for either the dark or light zones. In contrast, expression of UAS-ChR2 under the control of iav-GAL4 restored the ability of norpAP24 larvae to move away from the light (Fig. 4E). This behavior required blue light, consistent with the spectral sensitivity of ChR2. If the norpA P24;iav-GAL4/+;UAS-ChR2/+ larvae were fed on food, which was free of all-trans-retinal, they were unable to discriminate between the dark and light sides (Fig. 4E). We obtained indistinguishable results using the nan-GAL4 to direct expression of UAS-ChR2 in a norpAP24 background (Fig. 4E). Since stimulation of iav- or nan-expressing cells with light induced avoidance responses, these data indicated that wild-type larvae preferred 17.5° over 14−16°C through an avoidance response that required chordotonal neurons.

Requirement for iav for cool temperature-induced turning. A–D, Setup for testing whether temperature independent activation of chordotonal neurons induced an attractive or avoidance response. We expressed a ChR2 transgene (UAS-ChR2 transgene) under the control of the iav-GAL4 or the nan-GAL4 in a norpAP24 background. Blue- but not red light-activated ChR2. As controls, we analyzed wild-type and norpAP24 flies in the presence of blue or red light. Experiments with blue light are depicted. A, Side view of the setup. A semicircle (3.5 cm in diameter) was removed from the top lid of a black box to allow one side of the Petri dish (3.5 cm; 2% agarose), which was inserted into the box, to be exposed to illumination. B, Larvae were released along the middle of the dish. C, norpAP24 larvae randomly distributed throughout the dish in response to blue light. D, norpAP24;iav-GAL4/+;UAS-Chr2/+ larvae and norpAP24;nan-GAL4/UAS-Chr2 larvae avoided blue light when they were fed all-trans-retinal. E, Involvement of iav-expressing neurons in avoidance behavior. We eliminated the intrinsic light response using the norpAP24 mutation. The larvae were reared on food with or without all-trans-retinal and exposed to blue or red light as indicated. Complete preferences for the dark and light sides would produce P.I. values of 1.0 and −1.0, respectively. A lack of preference would result in a P.I. of 0. F, Measurement of turning angles (θ). The θ values were calculated using coordinates (P(N−1), P(N), and P(N+1)) in three consecutive frames. The initial linear movement (initial trajectory) was the vector connecting P(N−1) to P(N). The θ was based on the initial trajectory and the vector connecting P(N) to P(N+1). G, Average θ at 14° and 17.5°C. H, I, Percentage distribution of θ (H, wild type; I, iav1). J, The percentage of frames in which larvae made a turn ≥45°. Asterisks indicate significant differences (p < 0.05) with wild type, unless indicated with a bracket.
To explore further the behavioral basis for the iav-dependent avoidance of 14°C, we tested whether larvae displayed increased turning at the lower temperature. To test this proposal, we tracked the turning behavior of individual larvae at 17.5° and 14°C. After establishing the initial movement trajectory (initial trajectory, Fig. 4F), we determined the angle of deviation (θ; turning angle) from the initial trajectory over the 5.1 s interval between frames (Fig. 4F). We found that wild-type larvae showed an increase in the average turning angle at 14°C compared with 17.5°C (Fig. 4G). The percentage of lower angle turns (θ = 0–45°) decreased while the percentage of medium angle turns (45–90°) increased at 14°C (Fig. 4H). However, the percentage of high angle turns (θ = 90–180°) were similar at 14° and 17.5°C (Fig. 4H). Moreover, the total number of turns (θ ≥ 45°) nearly doubled (Fig. 4J; supplemental Fig. 1, available at www.jneurosci.org as supplemental material).
In contrast to wild-type, in iav1 mutant larvae, the total number of turns (θ ≥ 45°) and the average turning angles did not increase at 14°C (Fig. 4G,J; supplemental Fig. 1, available at www.jneurosci.org as supplemental material). At 17.5°C, iav1 larvae displayed average turning angles similar to wild-type larvae (Fig. 4G). This value did not increase at 14°C (Fig. 4G), and the percentages of low, medium and high angle turns were unchanged at the two temperatures (Fig. 4I). Even though iav1 larvae moved slower than wild-type larvae, the vectoral distance between the starting and ending points was similar to that of wild type at 14°C, due to the lower turning angles (supplemental Fig. 2A, available at www.jneurosci.org as supplemental material). Thus, iav1 larvae traveled as far as wild-type larvae and were able to survey as large a proportion of the thermal environment as the wild type. This is in contrast to nan36a mutant larvae, which traveled half the vectoral distance of wild-type and iav1 larvae (supplemental Fig. 2A, available at www.jneurosci.org as supplemental material) due to a combination of slow movement and high turning at both 14° and 17.5°C (supplemental Fig. 2B,C). Introduction of the wild-type iav+ transgene in iav1 restored the wild-type increase in the number of turns and average turning angle at 14°C (Fig. 4G; supplemental Fig. 2D, available at www.jneurosci.org as supplemental material). Based on these analyses, we suggest that iav is required for choosing the preferred temperature (17.5°C) over the cool temperature (14°C) through avoidance of 14°C, and this behavior is mediated by increases in the number of turns and turning angles at this lower temperature.
Discussion
A key goal in behavioral neurobiology is to define the cellular and molecular determinants that enable animals to decide between two or more alternative actions based on environmental cues. Thermotactic discrimination in Drosophila larvae represents a simple model to address the interplay between sensory input and choice selection. In the current study, we defined the cellular and molecular requirements for selection of slightly cool over the optimal temperature, and demonstrated how the cells and TRP channels contribute to thermotactic behavior. Specifically, we found that the chordotonal neurons functioned in the discrimination of 17.5°C and slightly cooler temperatures, and this required a TRPV channel, Iav. In support of a requirement for the chordotonal neurons, the effect due to loss of iav was reversed by introduction of the wild-type gene in chordotonal neurons. Moreover, expression of tetanus toxin in chordotonal neurons suppressed the ability to select 17.5 over 14°C, consistent with a requirement for synaptic transmission from the chordotonal neurons for cool temperature discrimination.
It has been reported previously that the terminal organs are involved in selecting 18°C over 11°C (Liu et al., 2003). We found that the chordotonal neurons and iav were not required for selecting the optimal temperature over temperatures cooler than 14°C. We propose that the chordotonal organ functions in the discrimination of 17.5°C versus slightly cooler temperatures (14−16°C), while the terminal organ functions in the selection of 17.5°C over 12°C and cooler temperatures.
The finding that Iav was required for choosing the optimal temperatures over slightly cool temperatures underscores the broad evolutionary role for TRPV channels in temperature selection. However, the requirement for Iav is distinct from mammalian TRPVs, several of which function in the responses to warm or hot temperatures (Bandell et al., 2007; Venkatachalam and Montell, 2007). Whether the other Drosophila TRPV, Nan, also contributes to temperature selection is unclear, since the thermotaxis assay was complicated by the greater sedentary behavior than iav, and the abnormally high turning even at 17.5°C. As a result, the nan larvae traveled small distances and remained near the initial site in which they were placed. Nevertheless, as is the case for Iav, the Nan channel is also expressed in chordotonal neurons, and therefore could potentially function in cool sensation (Gong et al., 2004). However, at least one other TRP channel, Pain, is expressed in chordotonal neurons (Tracey et al., 2003), but is not required for choosing 17.5°C over mildly cool temperatures.
In addition to Iav, another TRP channel, TRPL, functions in cool sensation (Rosenzweig et al., 2008). As was previously reported for TRPL (Rosenzweig et al., 2008), expression of Iav in oocytes did not elicit a cool-activated current (data not shown). Thus, it is not clear whether activation of either of these channels by thermal cool is direct or indirect. In contrast to iav and trpl, we did not detect a requirement for trp in temperature discrimination in the cool range, which differs from a previous report (Rosenzweig et al., 2008). However, a role for trp was less clear, given the lack of phenotype when the trp mutation was placed in trans with a deficiency, which uncovered the gene (Rosenzweig et al., 2008). Nevertheless, given that the previous and current studies focused on early and late stage larvae respectively, we cannot exclude that there are developmental differences in requirements for the TRP channel for sensing mildly cool temperatures.
Even though wild-type third instar larvae prefer 17.5°C over any other temperature, we found that the P.I. increased significantly in proportion to the alternative temperature (∼0.2, 12°C; ∼0.35, 14°C; ∼0.7, 24°C). We suggest that these differences are only partially due to the slower movements at the lower temperatures. The variations in the average P.I. values may reflect differences in the molecules, mechanisms and cell types involved in discriminating the optimal temperature (17.5°C) from mildly cool (14−16°C), cool (12°C) and comfortably warm (22−24°C) temperatures. In support of this proposal, inhibition of neurotransmission in chordotonal neurons specifically impaired the ability to discriminate between the preferred temperature over 14°C (Fig. 2B,C), and did not reduce selection of 17.5° over either 12°C or 24°C. Furthermore, a Gq/phospholipase C/TRPA1 thermosensory signaling cascade participates in choosing the optimal temperature over other temperatures in the comfortable range (20−24°C) (Kwon et al., 2008), while Iav and TRPL are required for opting for 17.5°C over mildly cool temperatures.
A critical question is how the thermotactic selection of the optimal over mildly cooler temperatures is accomplished via the chordotonal neurons and iav. The following three observations lead us to propose a model. First, iav mutant animals retain the ability to select the optimal temperature over very cool and comfortably warm temperatures. These results argue against a requirement for iav for positive selection of 17.5°C only, since this optimal temperature is still selected versus some temperatures. Second, stimulation of iav-expressing neurons with channelrhodopsin induces an avoidance response to light. Third, the modestly cooler temperature increased the average turning angles and the total number of turns (θ ≥ 45°) relative to wild type. Thus, we conclude that iav-dependent decision to choose the optimal over slightly cooler temperatures is mediated by increased activity of chordotonal neurons, which stimulates an avoidance response by increasing the number and magnitude of turning angles.
Footnotes
This work was supported by grants from the National Eye Institute (EY010852) and the National Institute of General Medical Sciences (GM085335).
- Correspondence should be addressed to Craig Montell at the above address. cmontell{at}jhmi.edu





{kind=link}
{kind=link}
{kind=link}
{kind=link}