

Abstract
Background: Schizophrenia is a highly heritable neurodevelopmental disorder. A genetic variant of microRNA-137 (miR-137) has yielded significant genome-wide association with schizophrenia, suggesting that this miRNA plays a key role in its etiology. Therefore, a molecular network of interacting miR-137 targets may provide insights into the biological processes underlying schizophrenia.
Methods: We first used bioinformatics tools to obtain and analyze predicted human and mouse miR-137 targets. We then determined miR-137 levels in rat barrel cortex after environmental enrichment (EE), a neuronal plasticity model that induces upregulation of several predicted miR-137 targets. Subsequently, expression changes of these predicted targets were examined through loss of miR-137 function experiments in rat cortical neurons. Finally, we conducted bioinformatics and literature analyses to examine the targets that were upregulated upon miR-137 downregulation.
Results: Predicted human and mouse miR-137 targets were enriched in neuronal processes, such as axon guidance, neuritogenesis and neurotransmission. The miR-137 levels were significantly downregulated after EE, and we identified 5 novel miR-137 targets through loss of miR-137 function experiments. These targets fit into a glucocorticoid receptor–dependent signalling network that also includes 3 known miR-137 targets with genome-wide significant association with schizophrenia.
Limitations: The bioinformatics analyses involved predicted human and mouse miR-137 targets owing to lack of information on predicted rat miR-137 targets, whereas follow-up experiments were performed with rats. Furthermore, indirect effects in the loss of miR-137 function experiments cannot be excluded.
Conclusion: We have identified a miR-137-regulated protein network that contributes to our understanding of the molecular basis of schizophrenia and provides clues for future research into psycho-pharmacological treatments for schizophrenia.
Introduction
Schizophrenia is a serious psychiatric condition with a lifetime prevalence of approximately 1%.1 Although its etiology is still largely unknown, schizophrenia is thought to result from a complex interplay of a subset of genetic and environmental factors.2,3 Despite its high genetic component — estimated to be approximately 80%1 — the identification and replication of genetic susceptibility factors for schizophrenia have proven challenging owing to the genetic and phenotypic heterogeneity that characterizes this disorder.1,2
In addition to multiple protein-coding genes that are implicated in schizophrenia pathophysiology, recent evidence indicates that dysregulation of small, noncoding RNAs, such as microRNAs (miRNAs), may play an important role in the etiology of the disorder.4 MicroRNA-encoding genes are highly conserved across evolution and have been shown to be involved in various fundamental biological processes.5 They regulate the expression of target messenger RNAs (mRNAs) by binding to their 3′-untranslated region (UTR), causing either mRNA degradation or protein synthesis inhibition.6,7 Each miRNA affects many mRNAs, while individual mRNAs can be targeted by several miRNAs. Some miRNAs are highly enriched in the brain or are even brain-specific, suggesting their unique regulatory roles in neuronal function,8 including the modulation of neuronal and synaptic plasticity9,10 and neuronal circuitry.11,12 In support of this notion, the miRNA target site density in synaptic mRNAs is twice as high as in nonsynaptic mRNAs.13 Furthermore, there have been numerous studies implicating brain- enriched miRNAs in the etiology of schizophrenia. For example, a subset of brain-enriched miRNA genes located within an imprinted cluster on chromosomal region 14q32 was differentially expressed in a large cohort of patients with schizophrenia.14 In addition, a microdeletion of chromosomal region 22q11.2, harboring the brain-enriched miR-185, has been consistently linked to schizophrenia, and deletion of the corresponding region in the mouse resulted in an altered development of neuronal dendrites and spines.15 The most robust genetic evidence for the involvement of a miRNA in schizophrenia has emerged from a genome-wide association study (GWAS) with a sample of approximately 50 000.16 In this study, a single-nucleotide polymorphism (SNP; rs1625579) located within an intron of the miR-137 host gene (MIR137HG) achieved genome-wide significance for association with schizophrenia.
Notably, rs1625579 (and hence miR-137) has also been genetically associated with a number of schizophrenia endophenotypes, including cognitive deficits, negative symptoms,17 brain activation,18,19 brain structure and age at disease onset.20 Moreover, apart from having a strong genetic link with schizophrenia, miR-137 is highly abundant in hippocampal and cortical neurons, where it regulates neuronal maturation and dendritic spine morphogenesis,21 and more specifically in the synaptic compartment.22 In addition, miR-137 modulates the proliferation and differentiation of murine adult neural stem cells,23 and its overexpression results in decreased dendritic spine size.21 Since disturbances in synaptic function and plasticity as well as in neuronal circuitry have also been associated with the onset and progression of schizophrenia,13,24,25 we hypothesized that miR-137 is involved in modulating gene expression during synaptic plasticity and that miR-137-mediated regulation of plasticity-related genes is disturbed in patients with schizophrenia. Therefore, we conducted bioinformatics and literature analyses to generate a molecular network of interacting miR-137 targets. In addition, transcriptomics data were derived from a rat neuronal plasticity model of somatosensory experience, and loss of miR-137 function experiments in primary cortical neurons were performed to validate these interacting miR-137 targets.
Methods
All experimental protocols and procedures were approved by the Animal Ethics Committee of the Radboud University Nijmegen (the Netherlands) and were in accordance with the Dutch legislation. Supplementary descriptions of the procedures we used are provided in the Appendix (available at jpn.ca).
Human and mouse miR-137 target prediction
A particularly powerful aspect of miRNA function is the ability of individual miRNAs to regulate a multitude of mRNAs coding for proteins with related cellular functions. Therefore, we explored the possibility that miR-137 has putative target genes with similar neuronal functions. Mature miR-137 is highly conserved across mammals, having exactly the same nucleotide sequence in humans, mice and rats. We used DIANA-microT 3.0, a computational algorithm for miRNA target prediction (www.microrna.gr/microT), to generate a list of putative human and mouse miR-137 targets (i.e., hsa-miR-137, mmu-miR-137). The DIANA-micro T 3.0 algorithm is based on several parameters calculated individually for each miRNA, and, as opposed to other commonly used miRNA prediction tools that use only conserved miRNA recognition/binding sites (e.g., TargetScan), this algorithm combines both conserved and nonconserved miRNA binding sites into a final prediction score, which correlates with the signal to noise ratio (SNR) and hence provides an indication of the false-positive rate of the prediction.26,27 The higher the prediction score, the lower the amount of false positives, and as a broad estimate, prediction scores of 19, 7.3 and 5 correspond to SNRs of 3, 2 and 1.5 and false-positive rates of 33%, 50% and 66%, respectively.26 In this respect, the DIANA algorithm is the only currently available miRNA target prediction tool that also allows for a statistical prediction of the extent to which a predicted target is a true target. Hence, to obtain a list of predicted human and mouse miR-137 targets with a predicted false-positive rate of 50% at the most, we used a threshold of at least 7.3 for the prediction score.
Functional classification of predicted human and mouse miR-137 targets
To detect significantly enriched gene categories in the predicted miR-137 target genes/mRNAs, we used the Ingenuity Pathway Analysis software package (www.ingenuity.com). It uses the Ingenuity Knowledge Base, which is based on information from published literature and many other sources, including gene expression and gene annotation databases, to assign genes to different groups and categories of functionally related genes. Each of these categories can be further divided into subcategories. Ingenuity calculates single p values for the enrichment of each gene category using the right-tailed Fisher exact test and taking into consideration both the total number of molecules from the analyzed data set and the total number of molecules linked to the same gene category according to the Ingenuity Knowledge Base. Furthermore, for each gene category, a multiple testing corrected p value of enrichment, calculated using the Benjamini–Hochberg correction, is provided.
Measurement of miR-137 levels in a rat model of neuronal plasticity
Our initial studies have shown significant enrichment of miR-137 in synaptic compartments of the mouse brain.22 To further examine the significance of this finding and assess if miR-137 levels are altered by synaptic/neuronal plasticity, we used a widely accepted in vivo model, the rat barrel cortex,28 to study experience-dependent changes in miR-137 expression. In the barrel cortex, plasticity can be induced by modifying somatosensory input through simple manipulations, such as exposing the animals to an enriched environment (EE), thus overstimulating the whisker-to-barrel pathway.29 We used RNA samples from a previous study, derived from primary somatosensory (barrel) cortical samples from adult rats that had been placed into an EE to elicit cortical plasticity (see Vallès and colleagues30 for details). Precursor (pre-) and mature (mat-) miR-137 expression levels in barrel cortex were determined by quantitative polymerase chain reaction (qPCR) immediately following the EE session and compared with levels measured in control rats. Significant differences in miR-137 expression between EE and control rats were determined with a Student t test at a significance level of p < 0.05 using SPSS software version 17.0 (SPSS Inc.).
Identification of putative rat miR-137 targets involved in neuronal plasticity
We analyzed a list of putative miR-137 targets involved in neuronal plasticity by comparing the putative list of human and mouse targets with the list of mRNAs that we previously found to be regulated in our rat neuronal plasticity model. This list of mRNAs regulated in rat barrel cortex after EE (170 mRNAs up-regulated and 31 mRNAs downregulated) was obtained using microarray analysis (significance analysis of microarrays q ≤ 0.05 and fold change ≥ |1.25|) in our previous study.30 As opposed to human and mouse miRNAs, the DIANA-microT 3.0 algorithm is not available for rat miRNAs. Therefore, in order to estimate which of the putative human and mouse miR-137 targets are also predicted to be rat miR-137 targets, we subsequently searched the miRWalk database31 for the “combined prediction score” of all putative miR-137 targets that achieved a DIANA miR-137 prediction score of at least 7.3 in humans and/or mice. This combined prediction score ranges from 1 to 4 and is composed of the individual prediction scores — 0 if the target is not predicted and 1 if the target is predicted — from the 4 currently available rat microRNA target prediction tools (i.e., miRanda, miRDB, TargetScan and miRWalk; www.umm.uni-heidelberg.de/apps/zmf/mirwalk).
Regulation of neuronal plasticity–involved targets by miR-137 in rat cortical neurons
To determine whether the observed experience-dependent changes in the expression of the putative miR-137 targets are mediated, at least in part, by miR-137, we transfected sequence -specific anti-miR-137 and nontargeting anti-miR control (NT) into rat primary cortical neurons. Twenty-four hours after transfection, expression levels of all EE-regulated mRNAs that are also putative miR-137 targets were analyzed by qPCR in neurons transfected with anti-miR-137 or NT. The effects of anti-miR137 treatment on mature miR-137 levels were also assessed in a comparable parallel experiment. Significant differences in expression between neurons transfected with anti-miR-137 and NT were analyzed with a Student t test at a significance level of p < 0.05 using SPSS software version 17.0. To correct p values for multiple testing32 and to protect against type 1 error, p values with a false discovery rate (FDR) of 0.05 were calculated using an Excel macro.33
Generation of molecular network of neuronal plasticity genes regulated by miR-137
We generated a molecular network by integrating the results of the bioinformatics analyses with systematic literature searches for the function of the neuronal plasticity targets whose expression was putatively regulated by miR-137 in rat cortical neurons. This was achieved through the molecular network building approach that we have previously applied to study the molecular basis of various neurodevelopmental disorders, including attention-deficit/hyperactivity disorder34 and autism spectrum disorders (ASDs).35
Results
Human and mouse miR-137 target prediction and functional classification of predicted targets
As described in the Methods section, using DIANA-micro T 3.0. with a cut-off of 7.3, we obtained a list of 1049 predicted human miR-137 targets and 814 predicted mouse miR-137 targets. The Ingenuity tool was subsequently used to identify significantly enriched gene categories within the predicted miR-137 targets. Table 1 lists the 10 most significantly enriched biological functions and canonical pathways within the predicted human and mouse miR-137 targets. In both species, several of the identified enriched gene categories are essential for (normal) neuronal development and function, including neuritogenesis, action potential of neurons and ephrin receptor signalling within the predicted human miR-137 targets and neurotransmission, action potential of neurons, ephrin receptor signalling and axonal guidance signalling within the predicted mouse miR-137 targets. These results are in accordance with those from a recent study by Wright and colleagues36 in which bioinformatics analyses were conducted on all human miR-137 targets predicted by the TargetScan tool. However, the criteria they used to designate a gene/mRNA as a putative human miR-137 target were much less stringent than the DIANA cut-off that we used in that they did not consider any statistical or other cut-off but included all genes/mRNAs that contained at least 1 conserved seed-binding site for human miR-137 in their subsequent bioinformatics analyses, as revealed by the TargetScan target predictions website. This approach suggests that a (much) larger proportion of genes serving as a starting point for analyses in the study by Wright and colleagues36 than our study may have been false positives.
Enrichment analyses of the genes/mRNAs predicted to be targets of human and mouse miR-137 using the DIANA tool*
Regulation of miR-137 expression and putative miR-137 targets in the rat barrel cortex following EE
We detected significantly reduced levels of pre-miR-137 (~66% reduction; t test, p = 0.006) and mat-miR-137 (~50% reduction; t test, p = 0.034) in the barrel cortex of rats exposed to EE compared with pre- and mat-miR-137 levels in the barrel cortex of control animals (Fig. 1A). Through applying the prediction criteria for human, mouse and rat miR-137 targets (see the Methods section), we identified 14 putative miR-137 target mRNAs that we previously found to be regulated among the transcriptomes examined in a rat neuronal plasticity model following EE30 (Table 2). The fact that all these predicted targets were upregulated in their mRNA levels is in keeping with the present finding that miR-137 expression was downregulated following EE.
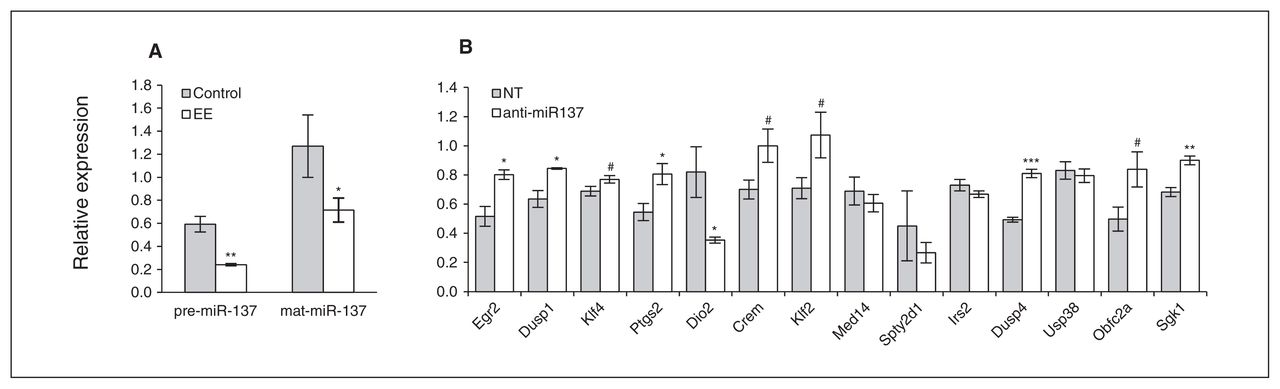
(A) Expression levels of microRNA-137 (miR-137) in the barrel cortex upon exposure of the rats to environmental enrichment (EE). We used a quantitative polymerase chain reaction (qPCR) assay to assess levels of both precursor (pre-) and mature (mat-) miR-137 in total RNA samples from the barrel cortex of rats exposed to EE and control animals. Levels of pre- and mat-miR-137 are expressed relative to U6 small nuclear RNA, used as an internal control; the scale of relative expression is ×10−5. Error bars represent the standard error of the mean (SEM) for control (n = 6) and EE (n = 12) measurements. Results are from 2 and 4 independent pooled samples (n = 2–3 animals per pool), respectively, assessed in triplicate using unpaired 2-tailed Student t tests (*p < 0.05, **p < 0.01). (B) Expression levels of the putative miR-137 target messenger RNAs (mRNAs) in a miRNA loss of function experiment. Primary rat cortical neurons were transfected with anti-miR-137 or, as a control, nontargeting short oligo-nucleotides (NT). Expression levels of predicted miR-137 target mRNAs were quantified by qPCR analysis 24 hours after oligonucleotide transfection. The putative miR-137 target mRNAs were previously found to be associated with activity-dependent plasticity in the rat barrel cortex.30 Error bars represent the SEM for independ ent experiments (n = 4); unpaired 2-tailed Student t test followed by Benjamini–Hochberg false discovery rate (FDR) multiple testing correction,32 #p < 0.10, FDR-corrected; *p < 0.05, FDR-corrected; **p < 0.01, FDR-corrected; ***p < 0.001, FDR-corrected.
Fourteen putative miR-137 target genes upregulated by enriched environment in the rat barrel cortex
Effect of endogenous miR-137 silencing on the expression of EE-regulated putative miR-137 targets
To examine the influence of miR-137 on the expression of the putative miR-137 targets identified in the EE neuronal plasticity model, we have repressed endogenous miR-137 function in primary cortical neurons in vitro using a sequence-specific anti-miR-137 inhibitor. This inhibitor significantly reduces miR-137 levels (see the Appendix, Fig. S1). After miR-137 decrease, we found a significant increase in the levels of 5 out of the 14 (36%) predicted and tested mRNA targets, namely Egr2, Dusp1, Ptgs2, Dusp4 and Sgk1 (t test followed by FDR multiple testing correction; all p < 0.05, corrected; Fig. 1B). Klf4, Crem, Klf2, and Obfc2a levels increased to a lesser and borderline significant extent (t test followed by FDR multiple testing correction, all p < 0.10), while the mRNA levels of Med14, Spty2d1, Irs2 and Usp38 remained unchanged. The finding that Dio2 mRNA levels were significantly decreased in neurons transfected with anti-miR-137 (t test followed by FDR multiple testing correction; p < 0.05, corrected) may be due to additional posttranscriptional regulatory mechanisms potentially exerting secondary signalling effects. These results suggest that miR-137, at least partially, modulates the expression of a significant number of genes in the rat barrel cortex upon induction of neuronal plasticity by EE.
Generation of molecular network of neuronal plasticity genes regulated by miR-137
Bioinformatics analyses of the 5 putative miR-137 targets up-regulated in both the EE neuronal plasticity model and following loss-of function of endogenous miR-137 in vitro showed that some of the most significantly enriched functional gene categories are directly related to the normal development and/or function of the nervous system (i.e., the biological functions epileptic seizure, behaviour and spatial memory; Table 3). In addition, 2 of the 6 significantly enriched canonical pathways (gluco-corticoid receptor signalling and aldosterone signalling in epithelial cells) point toward a role for these miR-137 targets in corticosteroid-related signalling. Furthermore, glucocorticoid receptor signalling — the most significantly enriched canonical pathway within the 5 targets — and protein kinase A signalling are the only 2 pathways that were significantly enriched in all 3 gene enrichment analyses that we conducted (i.e., the analyses of the predicted human and mouse miR-137 targets and of the 5 putative miR-137 targets that were upregulated following the antagomir mediated loss of function of miR-137 levels in primary neurons in vitro; Tables 1 and 3).
Bioinformatics-based enrichment analyses of the 5 predicted miR-137 target genes that are upregulated following enriched environment in the rat barrel cortex (Dusp1, Dusp4, Egr2, Ptgs2 and Sgk1)*
By integrating the results of the bioinformatics analyses with systematic literature searches for the functions of the proteins encoded by the 5 confirmed miR-137 target mRNAs, we built a molecular network that contains putative neuronal plasticity–related miR-137 targets (i.e., COX2 [encoded by PTGS2], DUSP1, DUSP4, EGR2, SGK1) and that may be involved in regulating neuronal plasticity through modulating glucocorticoid receptor–dependent signalling (Fig. 2). This outcome provides a testable model that requires further confirmation in follow-up studies. In addition to the 5 miR-137 targets, we placed the proteins encoded by 3 known miR-137 targets that have previously yielded genome-wide significant association with schizophrenia (i.e., CACNA1C, TCF4 and ZNF804A),37,38 as well as 5 neuronal plasticity–related proteins that functionally link the other network components (i.e., BDNF, DDIT3, EGR1, the glucocorticoid receptor itself and GRIN1) in the generated network (Fig. 2). Interestingly, the DIANA tool revealed that the glucocorticoid receptor (encoded by NR3C1) is a predicted target of both human miR- 137 (prediction score: 7.72) and mouse miR-137 (prediction score: 18.41).
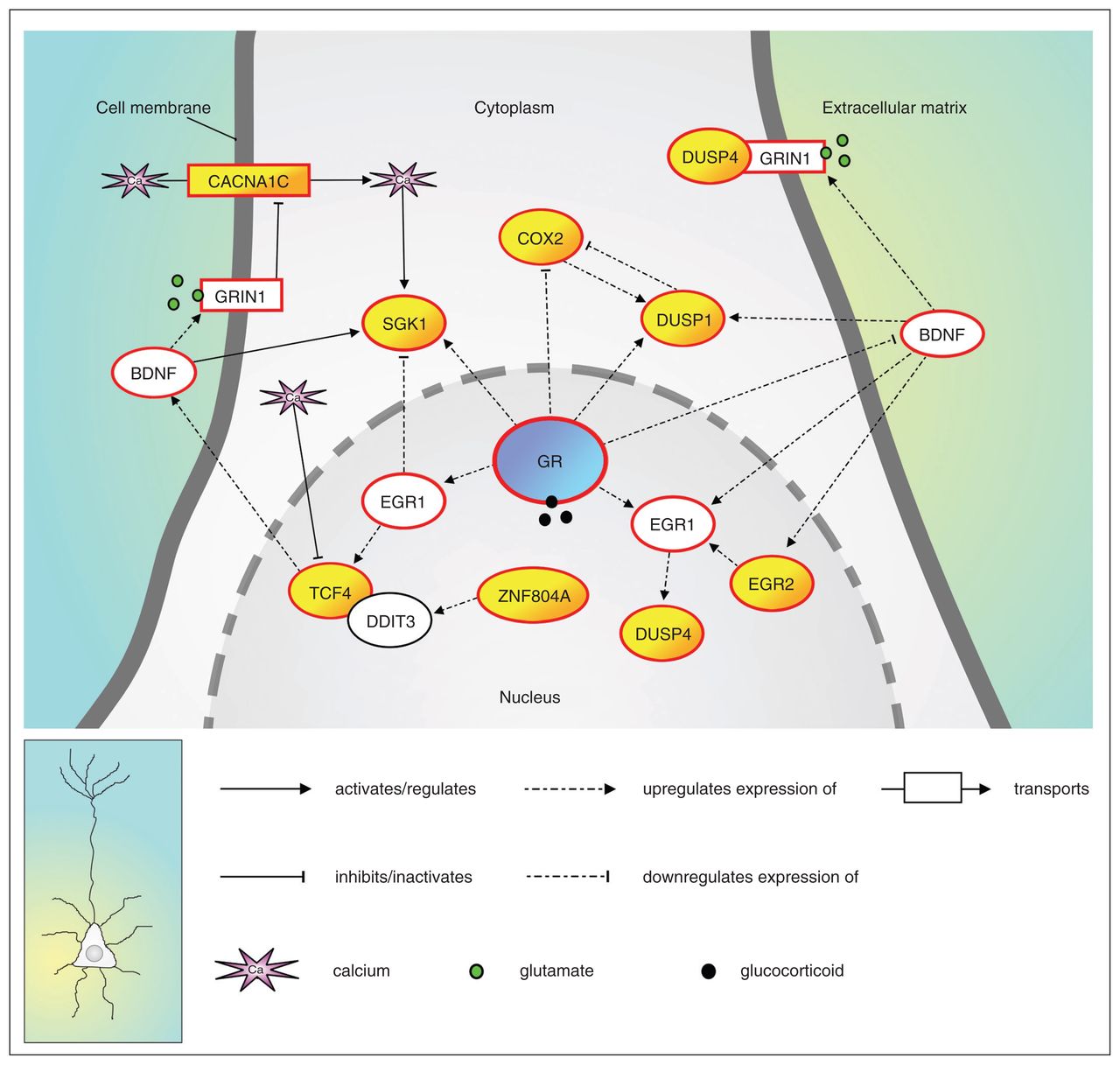
A microRNA-137 (miR-137)–regulated protein network that is implicated in neuronal plasticity through modulating glucocorticoid receptor–dependent signalling. The 5 novel miR-137 targets from this study (DUSP1; DUSP4; EGR2; PTGS2, which encodes the COX2 protein; and SGK1), as well as 3 previously validated miR-137 targets encoded by genes with a genome-wide significant association with schizophrenia (CACNA1C, TCF4 and ZNF804A), are depicted in yellow. The glucocorticoid receptor (GR; shown in blue) protein (encoded by NR3C1) is a predicted target of miR-137 in humans (DIANA prediction score: 7.72) and mice (DIANA prediction score: 18.41). Furthermore, all proteins with a red border have been implicated in schizophrenia through direct genetic evidence and/or messenger RNA/protein expression data (see the Appendix, Table S1). The network is described in full detail in the Appendix, and the current knowledge about the functions of the network proteins is presented.
The network is described in detail in the Appendix, and the current knowledge about the functions of the individual network proteins is delineated. Remarkably, except for DDIT3, all proteins in the network have been implicated in the etiology of schizophrenia through genetic evidence and/or mRNA/protein expression data (see the Appendix, Table S1), which provides a further functional and molecular link between miR-137- dependent dysregulation and schizophrenia.
Discussion
One of the main cognitive deficits observed in patients with schizophrenia is altered perception, resulting in hallucinations and delusions.39 While visual and auditory perception deficits in these patients have been well described, recent studies suggest that deficits in all the other sensory systems (olfactory, gustative, somatosensory) are also associated with schizophrenia pathology.40 Interestingly, sensory training improves cognitive abilities in patients with schizophrenia and results in changes of peripheral biomarker levels of neuronal plasticity.41 Accordingly, sensory stimulation through EE counteracted behaviours related to schizophrenia (e.g., stereotypical behaviour, impaired prepulse inhibition) in a mouse model of the disorder.42 Since the somatosensory system is the dominant sensory system in rodents, with a clear somatotopic representation of the whiskers (the main somatosensory organs) in the primary somatosensory cortex, we chose it to study the molecular basis of altered perception induced by sensory training in rodents.28
In the present study, we used bioinformatics and literature analyses to generate a molecular network of potentially interacting miR-137 targets during the induction of plasticity in the rat barrel cortex. We also identified 5 putative miR-137 target mRNAs by combining transcriptomics data derived from the rat neuronal plasticity model of somatosensory experience with the results of loss of miR-137 function in rat primary cortical neurons. The downregulation of miR-137 following EE that we observed is in keeping with other findings demonstrating that miRNA levels can be modulated upon the induction of long-term synaptic plasticity43 (e.g., for miR-13244 and for miR-485 during homeostatic plasticity45). Furthermore, a number of miRNAs are capable of fine-tuning the expression of synaptic plasticity proteins involved in actin cytoskeleton dynamics, or mRNA transport, resulting in spine morphogenesis.46–48
The molecular network that we have constructed includes the proteins encoded by the 5 miR-137 target mRNAs found to be upregulated in the rat barrel cortex following EE, as well as 3 previously validated miR-137 target proteins. Notably, all but 1 protein (DDIT3) within this network have been linked to schizophrenia through direct genetic evidence and/or expression data. Moreover, the proposed miR-137-regulated network is involved in experience-dependent neuronal plasticity through modulating glucocorticoid receptor–dependent signalling, a pathway that recently has been functionally linked to synaptic/neuronal plasticity49–52 and directly associated with schizophrenia.53,54 The glucocorticoid receptor pathway has been shown previously to be regulated both in a direct and indirect fashion by miR-124a and miR-18,55 whereas our results suggests that this pathway, at least partially, is regulated through miR-137. Furthermore, combined with the aforementioned findings from other groups, our results suggest a direct functional link between impaired miR-137- dependent regulation and schizophrenia through glucocorticoid receptor signalling.
Interestingly, although less strongly, miR-137 has been genetically linked to a number of other brain disorders, namely Alzheimer disease56 and 2 neurodevelopmental disorders other than schizophrenia, intellectual disability22 and ASDs.57 Notably, all these disorders are genetically and functionally associated with dysregulated stress and hypothalamic–pituitary– adrenal axis responses, which may involve, among other pathways, altered glucocorticoid receptor signalling50,58–61 and impaired synaptic plasticity.62–65 This suggests that dysregulation of miR-137 — and possibly impaired glucocorticoid receptor signalling — may lead to impaired synaptic plasticity, representing a common mechanism underlying cognitive deficits associated with clinically distinct neurologic disorders.
Finally, our finding that EE-mediated sensory experience caused a significant downregulation of miR-137 expression in the rat barrel cortex, along with the finding that EE counteracts schizophrenia-relevant behaviours in a mouse schizophrenia model (e.g., stereotypical behaviour, impaired pre-pulse inhibition),42 suggests that reducing brain miR-137 levels may lower the risk for schizophrenia-related behaviour. In this respect, it is of interest to note that in the human pre-frontal cortex miR-137 levels significantly decrease with age, with the most prominent decline in expression being observed in or just after late adolescence,66 which is the window of onset for schizophrenia. This notion indicates that insufficient decline in miR-137 during adolescence may contribute to the pathoetiology of schizophrenia. In support of this hypothesis, a recent study found that the schizophrenia risk allele of rs1625579 (the miR-137-linked SNP) is significantly associated with a lower miR-137 expression level in postmortem prefrontal cortex tissue of the control group but not the schizophrenia group.67 Importantly, beneficial effects of EE have also been reported in rodent models of Alzheimer disease,68,69 intellectual disability70 and ASDs,71,72 raising the intriguing possibility that modulating miR-137 levels may result in altered phenotypes of these 3 miR-137-linked neurologic disorders.22,56,57
Limitations
We used the DIANA algorithm to generate a list of predicted human and mouse miR-137 targets. Although the algorithm is one of the best currently available tools to predict miRNA targets26,27 it does not predict miRNA targets in rats, our animal model of choice to study the expression of miR-137 and its predicted targets in the context of neuronal plasticity. Hence, to estimate which of the putative human and mouse miR-137 targets are also predicted to be targets in rats, we used the miRWalk database to obtain a combined prediction score using the 4 currently available tools that predict targets of rat miRNA. This score indicated that 2 of the 5 predicted miR-137 targets regulated by neuronal plasticity and upregulated after loss of miR-137 function experiments, Ptgs2 and Dusp4, are not predicted targets in rats, which implies that the upregulation of these targets may be due to indirect effects. Therefore, follow-up studies should be performed to assess whether these 2 predicted targets are indeed direct miR-137 targets in the rat. In line with this, gain of miR-137 function experiments may also add relevant information. Although these points should be taken into consideration, our findings are very much in keeping with those of other studies and strongly support the important role of miR-137 in neuronal plasticity and the etiology of schizophrenia.
Conclusion
The outcomes of our studies suggest a critical role of miR-137 in modulating experience-dependent cortical plasticity via a glucocorticoid receptor signalling pathway. As such, the data provide a scope for developing novel molecular therapies aimed at lowering brain levels of miR-137 in order to treat schizophrenia and other miR-137-linked disorders.
Acknowledgements
We thank Laurens Kirkels, Mahshid Gazorpak, Aron Kos and Nikkie Olde Loohuis for technical assistance. This work was supported by VICI grant 453_04_002 from the Netherlands Foundation of Scientific Research (NWO) to P. De Weerd, a Dutch Top Institute Pharma grant (T5–209) to G.J.M Martens, and the Donders Center for Neuroscience fellowship award of the Radboud University Nijmegen Medical Centre and the FP7-Marie Curie International Reintegration Grant to A. Aschrafi.
Footnotes
↵* The authors contributed equally to this work.
Competing interests: None declared.
Contributors: A. Vallès, G. Poelmans and A. Aschrafi designed the study. A. Vallès acquired the data, which all authors analyzed. All authors wrote and reviewed the article and approved the final version for publication.
- Received November 24, 2013.
- Revision received January 15, 2014.
- Accepted February 14, 2014.
References
In this issue

{kind=link}
{kind=link}
Article tools






Related Articles
Cited By...
- No citing articles found.