





Abstract
The shells of many marine gastropods have ventrally directed serrations (serial projections) at the edge of the adult outer lip. These poorly studied projections arise as extensions either of external spiral cords or of interspaces between cords. This paper describes taxonomic, phylogenetic, architectural and functional aspects of serrations. Cord-associated serrations occur in cerithiids, strombids, the personid Distorsio anus, ocenebrine muricids and some cancellariids. Interspace-associated serrations are phylogenetically much more widespread, and occur in at least 16 family-level groups. The nature of serration may be taxonomically informative in some fissurellids, littorinids, strombids and costellariids, among other groups. Serrated outer lips occur only in gastropods in which the apex points more backward than upward, but the presence of serrations is not a necessary byproduct of the formation of spiral sculptural elements. In hard-bottom gastropods that do not flee from predators, pointed serrations may resist shear when the shell is clamped firmly to the substratum. The functions of serration in other gastropods are less clear, but likely involve defence against predators with soft feeding structures in some cases.
INTRODUCTION
The intricacy and diversity of sculpture on both the exterior and interior of molluscan shells have long been appreciated by collectors, taxonomists and those interested in shell function. The edge of the outer lip, where these external and internal features meet, has on the whole been much less carefully studied. Taxonomic descriptions often indicate whether the edge of a gastropod's outer shell lip is smooth, crenulated, denticulated or serrated, but the mode of formation and the diversity of edge ornamentation have been mentioned only in passing. In gastropods, the only features of the outer lip edge that have been investigated in some detail are abaxially orientated spines in such groups as Strombidae and Muricidae, ventrally directed labral teeth in Late Cretaceous to Recent predatory species, and adult modifications of the apertural rim (see, e.g. Lamy, 1931; Vignon, 1931; Vermeij & Signor, 1992; Vermeij, 2001; Merle, 2005). As a continuation of work on features of the shell margin in bivalves (Vermeij, 2013), this paper represents a descriptive survey and discussion of marginal modifications of the outer-lip edge in gastropods, with special emphasis on serial extensions that are either called serrations or denticulations (see below). Other important states or features of the outer lip are not treated here except insofar as they relate to the serrated condition. These additional traits and structures include an externally reflected lip, an internally thickened or inrolled lip, denticles formed along the inner side of the outer lip, the adapical or anal notch or sinus, the stromboid notch, the siphonal notch or canal, and the slit (characteristic of many early gastropods and pleurotomarioideans) (see Vermeij & Signor, 1992; Wagner, 1999; Vermeij, 2001, 2007). Finally, I contrast the ornamentation of the shell edge of gastropods with that of bivalves, noting that the often quite different functional demands of a univalve and a bivalve shell call for radically different architectures of the shell margin.
MATERIAL AND METHODS
I have surveyed all the major groups of Cenozoic and living gastropods in my research collection, supplemented by observations on specimens in museum collections, especially at Naturalis in Leiden. Descriptions in the taxonomic literature (including many of my own papers) proved generally to be inadequate for determining the nature and formation of sculptural features at the edge of the outer lip. Unless otherwise indicated, all descriptions refer to adult shells. Names traditionally recognized as subgenera have been treated as full genera for the names of species, a practice that is in many cases justified by characters of the shell edge. In genera such as Cerithium where appropriate taxonomic divisions have not yet been accepted, I place quotation marks around the genus to indicate the atypical nature of the species being discussed.
Terminology
I use the following terminology and conventions.
The outer lip (labrum) is the abaxial sector of the rim of the aperture, situated on the labral (or palatal) side of the aperture. The inner lip, on the labial side of the aperture, is the adaxially facing sector of the apertural rim.
The edge of the outer lip of a gastropod shell is the transition between the shell's exterior and interior surfaces.
Continuous external sculptural features oriented perpendicular to the outer-lip edge are referred to as spiral cords or interspaces. Comparable features on the inner surface of the labral (palatal) wall are referred to as lirae. Discontinuously formed external features are spines, nodes, beads and ribs (collabral, axial, opisthocline or prosocline). Their equivalents on the palatal side are denticles or (rarely) beads.
On the inner (labial) side, continuous spiral elements are columellar and parietal folds and, in the case of many species with a siphonal canal, the entrance fold to the canal. Discontinuously formed inner-lip denticles and beads are also known.
The edge of the outer lip is said to be polished if the inner shell layer extends over the edge to the exterior side of the lip; otherwise, the lip is said to be sharp.
Serial extensions of the external spiral elements are referred to as serrations if the projections are oriented ventrally—that is, towards the substratum on which the gastropod rests. Projections may be pointed or even spine-like, or they may be smoothly rounded. Serrations form as extensions of either cords or interspaces, or more rarely they may involve serial extensions of the lip as a whole.
Serial extensions of cords that are laterally directed, parallel to the substratum on which the gastropod rests or oriented dorsally away from the substratum, as referred to as lateral extensions.
A labral spine or tooth is a single (sometimes double) ventral extension of an external cord or interspace on or abapical to the midpoint of the outer lip in some predatory gastropods (Vermeij, 2001). More rarely it forms as a unique extension of the lip as a whole.
If the apertural rim is ontogenetically modified only once, at the termination of spiral growth, the gastropod is said to exhibit determinate growth. If the aperture is ontogenetically modified once or more during spiral growth, as indicated by axial thickenings (varices) oriented parallel to the outer lip's contour, growth is said to be episodic. Whether growth is periodic depends on the time course of growth events. If the apertural rim remains geometrically similar or changes gradually in form as it enlarges through ontogeny, the gastropod is said to exhibit indeterminate growth, even when the rate of growth slows markedly in adulthood. Note that the definition of determinate growth adopted here and by Vermeij & Signor (1992) differs from the definition and criteria set out by Papadopoulos, Todd, & Michel (2004).
DESCRIPTIVE ACCOUNT
Limpets
Cap-shaped shells in which the primary spiral (‘radial’) cords extend laterally as points beyond the overall contour of the margin occur widely among patellogastropods and siphonariids as well as in amathinids (Amathina) and some calyptraeids. In the last-named group, examples include eastern Pacific species of Crucibulum, the New Zealand Maoricrypta costata (Sowerby, 1824), and two species of Bostrycapulus, the Panamic B. urraca Collin, 2005 and the Floridian C. aculeatus (Gmelin, 1791). Among haliotids, the Indian Ocean Haliotis squamosa Gray, 1826 and H. mariae Wood, 1828 have strong spiral ribs extending laterally beyond the main contour of the shell margin. Other haliotids also bear spiral ribs, but these do not consistently extend laterally beyond the margin.
The Caribbean fissurelline fissurellids Cremides angusta (Gmelin, 1791) and C. barbadensis (Gmelin, 1791) show a less extreme lateral scalloping of the margin. No such rib extensions are known in diodorine and emarginuline fissurellids, deep-sea cocculiniforms, the freshwater neritid Septaria, stomatellid trochoideans, the coralliophiline muricid Galeropsis, freshwater pulmonate limpets and monoplacophorans.
Relatively few limpets display the condition in which the ventral shell edge is serrated by the ends of external sculptural elements. Fine serrations formed by tiny extensions of external interspaces between ‘radial’ cords occur in the hipponicid genus Sabia, and are the rule in diodorine and hemitomine fissurellids. They are also known in at least one emarginuline genus (Tugali) and in two fissurellines, the Caribbean Cremides nodosa (Born, 1778) and Clypidella fascicularis (Lamarck, 1822). If current taxonomy and phylogenetic hypotheses for Fissurellidae (McLean & Geiger, 1998; Aktipis, Boehm, & Giribet, 2011; McLean, 2011) are correct, the ventrally serrate condition would have evolved at least three times independently in the family. Based on these and other shell characters, however, I am sceptical that Cr. nodosa and Cl. fascicularis have been correctly classified. Their serrations, sculpture, central apex and ovate form all imply placement in Diodorinae, in which case ventral serration arose at most twice among fissurellids. It is remarkable that serration of this type is unknown in patellogastropods, haliotids, cocculinoideans, neriotoidean limpets, calyptraeids and siphonariids. The presence of spiral sculptural elements, which occur in most marine limpets, therefore guarantees neither lateral or ventral serration in limpet-like gastropods.
Coiled Vetigastropoda
Despite the very widespread occurrence of external spiral sculpture, serration at the edge of the outer lip is extremely rare in the clades Trochoidea and Seguenzioidea (for phylogenies and classification see Kano, 2008; Williams, Karube, & Ozawa, 2008; Williams et al., 2010; Bandel, 2010). The only coiled vetigastropods with a distinctly serrated outer lip I have found are a few turbinids of the Indo-West Pacific genus Marmarostoma (Fig. 1). In M. argyrostoma (Linnaeus, 1758), M. chrysostoma (Linnaeus, 1758) and M. setosum (Gmelin, 1791), external interspaces end in distinct projections, which are especially prominent on the abapical sector of the lip. This condition is also present in C. crassum (Wood, 1828) but is much less well developed, as is the external spiral sculpture. In other turbinids, the outer lip is sharp but not serrated or only slightly so. It is curious that internal lirae, with which interspace-related serrations might have been associated, are absent in Turbinidae. The only lirate vetigastropods I have found are some tropical eastern Pacific species of the tegulid genus Agathistoma, the Indo-West Pacific trochid tribe Trochini, the Indo-West Pacific monodontine trochid genus Monodonta (in which the lirae are very short), some tropical American calliostomatids and the Indo-West Pacific chilodontid seguenzioidean genus Euchelus. None of these taxa exhibits a serrated lip.
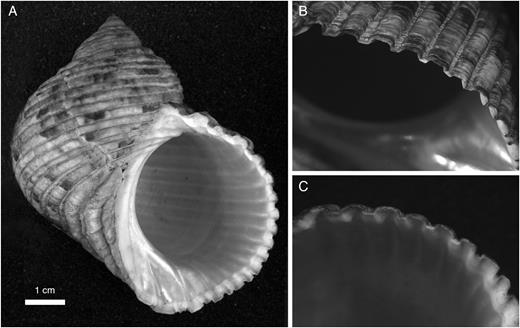
Marmarostoma setosum (Gmelin, 1791), Moluccas (collection Naturalis: ZMA 309371). A. Ventral view of shell. B, C. Close-up of outer lip from outer and inner side respectively, showing abapical serrations formed by external interspaces.
Neritidae
Within Neritimorpha, some members of the Neritidae display distinct extensions of external interspaces. Where present, serrations are usually weak, but they are quite prominent in the Indo-West Pacific Ritena plicata (Linnaeus, 1758), species in the Nerita (s. l.) costata Gmelin, 1791 group, and most members of the genus Cymostyla. Serration is absent in Theliostyla aibicilla (Linnaeus, 1758) and the closely related T. sanguinolenta (Menke, 1829), as well as in Cymostyla maxima (Gmelin, 1791) and all neritid genera outside the Nerita clade (for classification and phylogeny see Frey & Vermeij, 2008). As in other groups, the presence of external or internal spiral elements does not guarantee a serrated lip edge. The western Pacific Theliostyla exuvia (Linnaeus, 1758) and the Indian Ocean T. textilis (Gmelin, 1791) have prominent external spiral cords, but their outer lips are not serrate. Most species in the Nerita clade have denticles on the inner side of the outer lip but, as noted above, many are not serrated.
Littorinidae
The circular to broadly ovate aperture of littorinids is bordered by an outer lip whose edge in the adults of most species is smooth or nearly so. However, several taxa develop small, rounded serrations formed as projections of external interspaces. These serrations are best expressed in the Indo-West Pacific Tectarius pagodus (Linnaeus, 1758) and T. tectumpersicum (Linnaeus, 1758). These high-shore species are unique among living littorinids in having denticles on the inner side of the outer lip and long, widely separated, thin lirae further inside the aperture. In the closely related but smaller-shelled taxa Echininus, Echininiopsis and Tectininus (all three synonymized under Tectarius by Reid, Dyal, & Williams, 2012), the outer lip and labral wall are smooth.
Serration is also present in the Caribbean Cenchritis muricatus (Linnaeus, 1758) and in all species of the Atlantic-East Pacific members of Echinolittorina with nodular sculpture that I have examined. All serrated littorinid species bear external beads, tubercles or spines, but the presence of such protrusions is not invariably associated with serration, as indicated by smooth-edged species of Nodilittorina and the Indo-West Pacific clade of Echinolittorina s. l. (for classification and phylogeny see Williams and Reid, 2004; Reid, Dyal, & Williams, 2012). From the phylogenetic studies of Reid, Dyal, & Williams (2012), it is clear that serration (and liration) is a derived condition in Littorinidae. It is not yet clear how many times serration arose, but liration evolved only once in the family.
Cerithioidea
Although some clades within or close to Cerithioidea lack outer-lip serrations (e.g. Turritellidae, Vermetidae, Dendropomatidae and Modulidae), others do possess these features. In the Cerithiidae, rounded or pointed projections formed at the ends of spiral cords are widespread. They are most prominently expressed in rock-dwelling species from the Indo-West Pacific, such as Cerithium columna (Sowerby, 1855), C. echinatum (Lamarck, 1822), C. mutatum (Sowerby, 1834) and C. nodulosum Bruguière, 1792 (Fig. 2). They occur in reduced form even in many sand-dwelling species of Rhinoclavis and Pseudovertagus, but serration is essentially absent in Rhinoclavis vertagus (Linnaeus, 1758) and in the rock-dwelling diminutive Semivertagus nesioticus (Pilsbry & Vanatta, 1906). In serrated species of Rhinoclavis, the polished, rounded projections do not exhibit the asymmetry of the cords, whose abapical edges are less steeply inclined than their adapical edges. Most American cerithiids have weakly to obsoletely serrated lip edges, as do Indo-West Pacific species of Clypeomorus and many species that Houbrick (1992) allocated to a very broadly defined genus Cerithium.
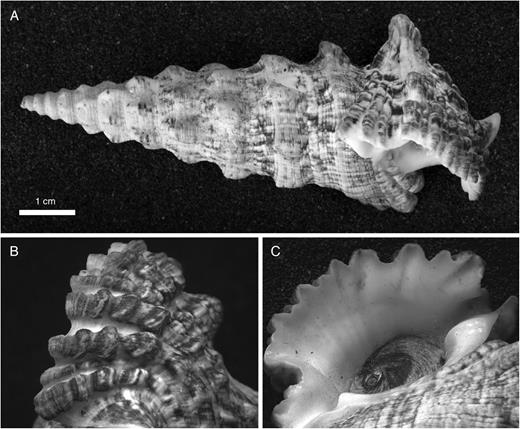
Cerithium nodulosum Bruguière, 1792, Mombasa, Kenya (collection Naturalis: ZMA Moll 161334). A. Whole shell, showing edge of lip. B. Dorsal view of last whorl, showing sculpture. C. Ventral view of outer lip and aperture, showing serrations formed at ends of cords.
Within Potamididae, low rounded projections of external spiral interspaces form on the outer-lip edge in a few Indo-West Pacific species including Terebralia palustris (Linnaeus, 1758), some T. sulcata (Born, 1778) and Telescopium telescopium (Linnaeus, 1758). Other members of the family have smooth lip margins. In Terebralia, rows of strong interior denticles periodically form at the position of external varices, but these denticles are not evident at the outer lip of the adult stage and are therefore not intimately associated with the weak serrations. No such rows of denticles occur in Telescopium, which lacks varices.
Stromboidea
The Stromboidea exhibit a great diversity of conditions of the shell margin, much of which has gone largely unnoticed. The most familiar condition is the presence of laterally directed spines, formed as extensions of major cords. This condition characterizes adults of all members of the Aporrhaidae and also occurs in Indo-West Pacific genera such as the rostellariid Tibia and the strombids Harpago, Lambis and Sinustrombus.
Ventrally directed serrations, lobes or spines occur in many Indo-West Pacific stromboideans. In Tibia, blunt spines associated with obsolete spiral cords are oriented ventrally at the edge of the adult outer lip. In Sinustrombus taurus (Reeve, 1857) and all species of Lambis, strong rounded projections situated abapical to the stromboid notch also form at the ends of cords, whereas in Lentigo lentiginosus (Linnaeus, 1758), similar projections are associated with external interspaces. Harpago chiragra (Linnaeus, 1758), which like Lambis and Sinustrombus has laterally directed spines, lacks these anterior projections. In Tridentarius dentatus (Linnaeus, 1758), there are three ventrally directed spines, one anterior to and two behind the stromboid notch (Kronenberg & Vermeij, 2002). All three spines appear to be extensions of cords, which are developed only near the adult outer lip. The available stromboidean phylogeny (Latiolais et al., 2006) indicates that these configurations arose independently in several lineages of the clade.
The second category of ventrally directed projections comprises broad, rounded lobes or waves on the outer lip posterior to the stromboid notch. These lobes, which appear to be associated with major external cords, are seen in the genera Lentigo, Gibberulus and Lambis. Kronenberg & Vermeij (2002) have already recognized that these waves are absent in all American strombids, which also lack the anterior spines and lobes referred to above. All ventrally directed elaborations occur in species whose adult outer lip is polished.
A few Indo-West Pacific strombids display a series of very small, sharp projections just posterior to the stromboid notch on the main sector of the adult outer lip. I have observed these teeth in Canarium urceus (Linnaeus, 1758) and C. wilsonorum (Abbott, 1967) (for nomenclature see Kronenberg & Dekker, 2000; Willan & Kronenberg, 2004), both with sharp-edged rather than polished lips. No such ventrally directed serrations are present in other species that have been assigned to Canarium, including C. haemastoma (Sowerby, 1842), C. labiatum (Röding, 1798), C. maculatum (Sowerby, 1842), C. microurceus (Kira, 1959), C. mutabile (Swainson, 1821) and C. ochroglottis (Abbott, 1960). Together with other characters, the presence of serrations in C. urceus (the type of Canarium Schumacher, 1817) and related species may point to the necessity of restricting the genus Canarium to these species and of separating those without serrations into a distinct genus, for which the name Conorium Jousseaume, 1880 (type species: Strombus mutabilis Swainson, 1821) is available.
Ovulidae
Most cypraeoideans, including Ovulidae, have an adult outer lip that is thickened and inwardly enrolled. Often, this lip is adorned with serial denticles on its inner side. In a few Indo-West Pacific ovulids, whose lip shows a slight external expansion, these denticles extend to the lip's edge as closely spaced, sharp projections, which in Serratovolva dondani (Cate, 1964) are much more prominent and much more widely separated than the minute spiral threads on the shell's exterior. Similar marginal denticles, presumably formed in the same way, characterize the ovulid genera Dentiovula and Prionovolva (Cate, 1973).
Tonnoidea
Marginal serrations or teeth are common and often prominent in members of the Tonnoidea. Where they occur, they are almost always formed as extensions of external interspaces or of internal lirae or denticles. Sharply pointed edge denticles occur in some Bursidae, including species of Bufonaria, Bursa, Tutufa and Tutufella, but lip edges are smooth or hardly serrated in such other bursids as Aspa, Crossata, Bursa corrugata (Perry, 1811) and B. ranelloides (Reeve, 1844). In Ranellidae, sharply rounded serrations formed as extensions of denticles on the inner side of the outer lip occur widely, but they are absent in the tropical genus Gutturnium and in such cool-water genera as Argobuccinum, Fusitriton and Priene. Exceptionally prominent, sharp and widely separated serrations occur in species of Charonia. Spine-like serrations formed at the ends of external interspaces or as continuations of internal denticles are found in the cassid genera Phalium (abapical sector only) and Casmaria, and in the tonnid genus Malea.
In the personid genus Distorsio, the type species D. anus (Linnaeus, 1758) has up to three rows of ventrally directed projections on its expanded ventral surface (Fig. 3). The marginal lobes form as sharply rounded extensions of external cords. A second row of sharper projections corresponds to extensions of interspaces. On the abapical sector of the adult outer lip, a third inner row often forms, again corresponding to cords. Other species assigned to Distorsio show much less prominent projections. These are formed as extensions of external interspaces in D. decipiens (Reeve, 1844), D. minoruonishii Parth, 1989 and D. smithi von Maltzan, 1884. In still other species such as D. decussata (Valenciennes, 1832) and D. reticularis (Linnaeus, 1758), the edge of the outer lip is nearly smooth. As noted for several other groups, taxonomic reassessment based on these previously unobserved differences in outer-lip features may require the subdivision of Distorsio as currently understood into several genus-level taxa (for current practice see Kronenberg, 1994; Beu, 1988, 1998, 2005, 2010).
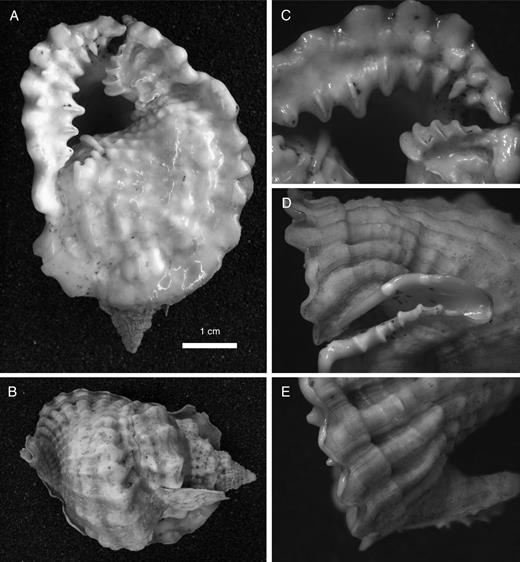
Distorsio anus (Linnaeus, 1758), Mauritius (collection Naturalis: RMNH Moll 193544). A. Ventral view of whole shell. B. Dorsal view. C. Ventral close-up of outer lip. D. Close-up of dorsally recurved siphonal canal. E. Close-up of outer lip from outer side, showing three rows of serrations, the first and third formed as extensions of cords, the second as continuations of interspaces.
Muricidae
Most muricids have ventrally directed serrations on the adult outer lip of the shell. In the clades Muricinae, Ergalataxinae, Coralliophilinae and Rapaninae, as delineated by Barco et al. (2010), these serrations are produced as extensions of interspaces between cords (Fig. 4), whereas in Ocenebrinae they are extensions of the cords themselves. Members of the Haustrinae have poorly developed serrations, whereas most Trophoninae and Typhinae lack them. Serrations are also absent in such sedentary coralliophilines as Magilus antiquus Montfort, 1810, Galeropsis monodonta (Quoy & Gaimard, 1834) and Latiaxis mawae (Griffith & Pidgeon, 1834).
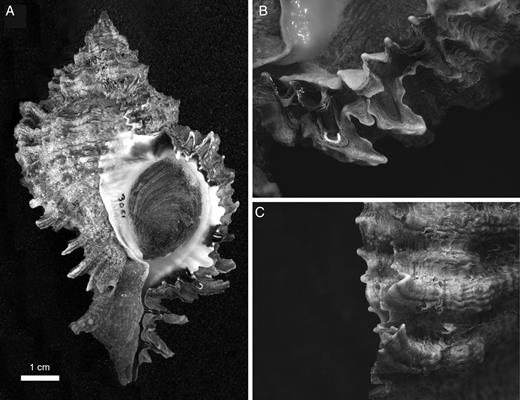
Phyllonotus oculatus (Reeve, 1845), Anse à la Banque, Guadeloupe (collection Naturalis: RMNH Moll.Vink 3081.1). A. Ventral view of shell. B, C. Outer lip close-up from inner and outer side respectively, showing serrations formed as continuations of interspaces.
Buccinoidea
The condition of ventrally directed serial serration of the adult outer lip is very common in the Buccinoidea. Projections formed at the ends of external interspaces are found in nearly all living Fasciolariidae (Snyder et al., 2012; Lyons & Snyder, 2013), Colubrariidae, Melongenidae (mainly on the abapical sector of the lip), Pseudolividae (abapical to the so-called pseudolivid groove, which ends in a labral tooth; see Vermeij, 1998), warm-water Buccinulidae (e.g. Kelletia, Lirabuccinum and Siphonalia) and the buccinid subfamilies Pisaniinae (especially members of the Cantharus group; see Vermeij, 2006) and Photinae. Serration is absent in a few temperate South African fasciolariines (Snyder et al., 2012) and in most cold-water buccinoideans (including Buccininae, Colinae, Buccinulidae and Busyconinae).
In the Nassariidae, serrations occur in some species of one clade in the Indo-West Pacific, and in reduced form in some West African species and in the Californian Caesia fossata (Gould, 1850). Although several taxonomic descriptions refer to denticles at the edge of the abapical sector of the outer lip as extensions of basal cords (Kool, 2003, 2008; Dekker & Dekkers, 2009), my observations indicate that nassariid serrations at the lip's edge (often confined to the abapical sector) form as extensions of denticles or lirae on the inner side of the outer lip, effectively corresponding to external interspaces. Often, these serrations occur along a sector of the lip where no cords (and therefore no interspaces) are visible, as in Nassarius coronatus (Bruguière, 1789), Zeuxis dorsatus (Röding, 1798), ‘Niotha’ distorta (A. Adams, 1852), Z. bicallosus (E. A. Smith, 1876), Z. siquijorensis (A. Adams, 1852), Z. vitiensis (Hombron & Jacquinot, 1854), Telasco luridus (Gould, 1850) and T. sufflatus (Gould, 1860). Particularly prominent spine-like serrations occur in Alectrion papillosus (Linnaeus, 1758) and A. glans (Linnaeus, 1758). The tiny labral tooth found at the abapical end of the outer lip in species of Hebra and in some species of ‘Niotha’ is likewise an extension of an interspace. A genus-level revision of nassariids, taking into account morphological characteristics of the shell as well as molecular sequences, is very much in order. No serration is present in the majority of nassariid species, including all living and fossil tropical American species I have examined. They are also absent in members of the Dorsaninae.
Harpidae
A few species of Harpa are characterized by blunt or tooth-like serrations on the anterior (abapical) sector of the outer lip. They include Harpa harpa (Linnaeus, 1758), H. amouretta Röding, 1798, H. doris Röding, 1798 and H. crenata (Swainson, 1822). Although no strong spiral sculpture is present on the external surface at locations corresponding to these serrations, the serrations appear to arise from the inner side of the lip, implying that the projections are extensions of interspaces. In species of Morum, the thickened outer lip bears faint serrations, which are always extensions of spiral features on the inner side of the outer lip.
Costellariidae
The great majority of Costellariidae have smooth, polished lip margins, but a few species possess fine, rounded projections, which in Vexillum plicarium (Linnaeus, 1758) become sharply pointed on the abapical sector of the lip. In all cases these projections correspond to external interspaces. Species from soft bottoms that have serrated lips include V. plicarium, V. rugosum (Gmelin, 1791), V. regina (Sowerby, 1828), Costellaria deshayesi (Reeve, 1844), C. modesta (Reeve, 1845) and C. pacifica (Reeve, 1845). Hard-bottom species with serrations include Pusia amabilis (Reeve, 1845), P. bernhardina (Röding, 1798), P. crocata (Lamarck, 1811), C. speciosa (Lamarck, 1844) and C. tuberosa (Reeve, 1845). All these species are found in shallow Indo-West Pacific waters. As is the case in Nassariidae, the genus-level taxonomy of Costellariidae is in dire need of revision, because current classification belies great morphological heterogeneity within genera.
Mitridae
A weakly crenulate or serrate margin is a feature common to many mitrids including most species of the genera Nebularia, Neocancilla, Pterygia, Scabricola, Subcancilla and Swainsonia, as well as in some species of the Indo-West Pacific genus Strigatella such as S. auriculoides (Reeve, 1845), S. astricta (Reeve, 1844), S. pellisserpentis (Reeve, 1844) and S. retuse (Lamarck, 1811). In species of Mitra, all from the Indo-West Pacific, the lip bears strong crenulations or spinose denticles, either along its entire length as in M. papalis (Linnaeus, 1758) or on the abapical sector as in M. mitra (Linnaeus, 1758). All projections appear to be formed from the inner side of the outer lip and therefore correspond to external interspaces even when external sculpture is absent. Other mitrids and the related families Pleioptygmatidae and Volutomitridae have smooth lip margins.
Vasidae
Adult shells of the genus Vasum have a polished outer lip with rounded low extensions of the external interspaces, evident mainly on the abapical sector of the lip. These are most prominently expressed in the large Indo-West Pacific V. ceramicum (Linnaeus, 1758).
Cancellariidae
Many members of the Cancellariidae have a serrated outer lip. Serrations are continuations of external cords in the tropical eastern Pacific Aphera tessellata (Sowerby, 1832) and Hertleinia mitriformis (Sowerby, 1832) and more subtly in Bivetiella obesa (Sowerby, 1832). Interspace-associated serrations, which are typically low and closely spaced, occur in the plesiotritonine Indo-West Pacific Plesiotriton vivus Habe & Okutani, 1981, and in the cancellariines Bivetiella simlis (Sowerby, 1832) from West Africa, Bivetopsia chrysostoma (Sowerby, 1832) in the eastern Pacific, and at least some populations of Cancellaria reticulata (Linnaeus, 1758) from the Caribbean, and species of Merica from the Indo-West Pacific including M. boucheti Petit & Harasewych, 1986, M. oblonga (Sowerby, 1832) and M. sinensis (Reeve, 1856). The adult lip is smooth or nearly so in many taxa including species of Nevia, Scalptia, Solatia, Sveltia, Sydaphera, Tribia, Trigonostoma, Trinotaphera, and the plesiotritonines Loxotaphrus and Tritonoharpa. Taxonomic descriptions include mention of serrations also in Africotriton, Nopponaphera, Ovilia and Ventrilia (Bouchet & Petit, 2002; Landau, Petit, & Marquet, 2006; Landau, Petit, & Da Silva, 2007; Landau, Petit, & Da Silva, 2012), but it is not clear from these descriptions how the serrations are formed.
Conoidea
I have not made a thorough survey of conoideans, but it is clear that outer-lip serration is widespread in this large clade. All serrations I have seen represent extensions of interspaces. These features occur in the Indo-West Pacific drilliid Clavus auricularia (Link, 1807), the Indo-West Pacific turrid genera Lophiotoma and Turridrupa, the eastern Pacific pseudomelatomid genus Knefastia, and most strikingly in the Early Miocene to Recent tropical American clathurellid genus Glyphostoma. In the Conidae, serration occurs only on the abapical sector of the outer lip in some Indo-West Pacific species. These are best developed in Conus marmoreus Linnaeus, 1758 (Fig. 5), and its relatives, as well as in Pionoconus catus (Hwass in Bruguière, 1792), P. striatus (Linnaeus, 1758), and Cylindrus (or Dendroconus) textile (Linnaeus, 1758). All these species feed on either fishes or gastropods. Weakly developed rounded projections characterize the abapical sector of the lip in such Indo-West Pacific species as Miliariconus coronatus (Gmelin, 1791), Rhombiconus distans (Hwass in Bruguière, 1792), R. imperialis (Linnaeus, 1758), Puncticulus arenatus (Hwass in Bruguière, 1792), P. pulicarius (Hwass in Bruguière, 1792), Virroconus chaldaeus (Röding, 1798) and V. ebraeus (Linnaeus, 1758). All conids with abapical serrations except the species of Rhombiconus and Virroconus have a relatively broad aperture, although not all wide-apertured conids have serrations, as exemplified by Gastridium geographus (Linnaeus, 1758), G. tulipa (Linnaeus, 1758), and species of the Atlantic-East Pacific genus Chelyconus and the Indo-West Pacific Protostrioconus obscurus (Sowerby, 1833).
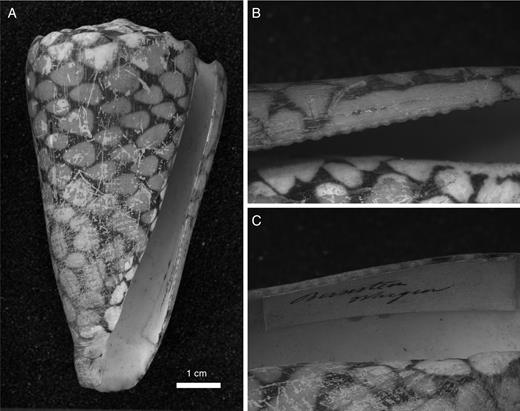
Conus marmoreus Linnaeus, 1758, Waigeo, Indonesia (collection Naturalis: RMNH 220893). A. Apertural view of shell. B, C. Close-up of outer lip from outer and inner side respectively, showing fine serrations on abapical sector formed as extensions of interspaces.
DISCUSSION
Phylogenetic and temporal distribution of serration
From the preceding survey it appears that serration at the edge of the adult outer lip is phylogenetically widespread in marine gastropods. It is especially frequent in the Caenogastropoda, particularly in Riedel's (2000) clade Latrogastropoda, which in the living fauna comprises Neogastropoda and the group that Bandel (1993) called Neomesogastropoda. The phenomenon is unknown in Patellogastropoda, Ptenoglossa, shell-bearing Heterobranchia, most Vetigastropoda (except some Fissurellidae and Turbinidae), and such large neogastropod families as Columbellidae, Cystiscidae, Marginellidae, Olivellidae, Olividae and Volutomitridae.
Strikingly, serrations are very rare and never well expressed in freshwater and terrestrial gastropods. Only a few freshwater cerithioideans, such as species of Lavigeria in Lake Tanganyika, show traces of lip serration. I return to this curious scarcity below.
Downward-facing serial projections on the adult outer lip form either as extensions or cords or as extensions of interspaces between cords (often observable only as denticles and ridges on the inner side of the outer lip). Serrations associated with interspaces are phylogenetically much more widespread than those associated with cords. Among living gastropods, extensions from interspaces occur in fissurellids, some species of Marmarostoma (Turbinidae), neritids, littorinids, potamidids, some strombids, the hipponicid Sabia, most tonnoideans, most muricids, buccinoideans, vasids, costellariids, mitrids, volutoideans, some cancellariids and conoideans. Cord-associated serrations are known only in cerithiids, stromboideans, the personid tonnoidean Distorsio anus, ocenebrine muricids and cancellariids. On the basis of this distribution, serration evolved a minimum of 22 times at the family level, 17 as extensions of interspaces and 5 as extensions of cords. These numbers are certainly underestimates, because serration likely arose multiple times in such clades as Fissurellidae, Buccinoidea, Volutoidea and Conoidea.
The fossil record indicates that serrated margins first appear in the Late Cretaceous period. According to Sohl (1964), serrations (‘crenulations’) occur at the lip edge in such Late Cretaceous genera as Buccinopsis, Deussenia, Hercorhynchus, Hydrotribulus, Longoconcha, Lupira, Ornopsis, Pyropsis [P. perlata (Conrad, 1860)], Volutomorpha, and Woodsella. Bandel & Stinnesbeck (2000) added Concepcionella to this list. Whether these crenulations form at the ends of interspaces or cords is unreported except in the cases of Buccinopsis (Sohl, 1964) and Volutomorpha (Dall, 1907), where the projections are extensions of cords.
The phylogenetic position of most of these genera remains controversial, but it is now clear that Cretaceous taxa generally do not belong to clades and families with living members. Bandel & Dockery (2012), in their most recent review of Late Cretaceous gastropods based on characters of the protoconch, consider Buccinopsis, Hercorhynchus, Hydrotribulus, Lupira and Pyropsis as members of the Moreidae, a family in the neomesogastropod superfamily Sarganoidea. They assign Deussenia and Ornopsis to the pyrifusid subfamily Prifusinae, and Longoconcha and Volutomorpha to the pyrifusid subfamily Volutoderminae. In their view, the Pyrifusoidea (which includes Pyrifusidae and Pseudolividae) are stem neogastropods. If these interpretations are correct, as I suspect they are for the most part, the evolution of serration in Cretaceous gastropods would have been independent of that in the succeeding Cenozoic era.
Whether serration appeared before the Late Cretaceous is difficult to infer from the known fossil record. Very few Paleozoic and Mesozoic gastropod fossils have an intact outer lip. On phylogenetic grounds, however, it seems likely that outer-lip serration was rare before the Late Cretaceous. Serrated members of clades with Early Mesozoic origins such as Fissurellidae, Trochoidea, Neritoidea, Littorinoidea, Cerithioidea and Stromboidea all belong to highly derived Cenozoic subclades. All the other cases discussed in this paper are concentrated in the Latrogastropoda, which Bandel & Dockery (2012) trace back to the Aptian–Albian interval of the mid-Cretaceous.
Constructional aspects
To some observers, it might be tempting to interpret serrations as incidental structural byproducts of spiral sculpture. The evidence, however, contradicts this interpretation. Numerous gastropod shells have external spiral sculpture but lack marginal serration. This condition occurs in nearly all subadult shells of species that have serrations as adults. Moreover, many species with spiral cords lack serrations even as adults, including patellogastropods, haliotids, trichotropids, epitoniids and acteonids, among others. Many gastropods with serrated adult lips lack external spiral sculpture corresponding to the projections, which instead appear to form as extensions of denticles or lirae on the inner side of the outer lip. These internal spiral elements are usually at positions that would correspond to external spiral interspaces.
Shell geometry appears to play a role in whether serrations can form at the edge of the outer lip. All cases of serration of which I am aware occur in gastropod shells in which the axis of coiling lies at a low angle of elevation with respect to the apertural plane, that is, in shells in which the apex points back more than up (Vermeij, 1971). In shells with a high angle of elevation, conforming to the trochoid shape, downward-facing projections on the lip would be expressed largely within the plane of the lip, which itself is in the form of a logarithmic spiral that accretes at the posterior end of the aperture (Vermeij, 2002). This condition occurs in haliotids, most Trochoidea (except some Turbinidae), and trochiform genera of Littorinidae, all of which lack outer-lip serration. In gastropods with a low angle of elevation, ventrally directed projections are extensions of either cords or interspaces in the direction of spiral growth, and thus extend out of the plane of the lip. In short, shells with the apex directed backward are architecturally much less constrained with respect to outer-lip ornament, than those in which the apex is oriented upward (Vermeij, 1971, 2002).
Many marine predatory gastropods have evolved a labral tooth, a single (or sometimes double) projection on the middle or abapical sector of the outer lip (Vermeij, 2001). Like serial serrations, the labral tooth forms as an extension of either a cord or an interspace, or in a few cases as an extension of the outer lip as a whole. In some cases, the labral tooth is little more than an exaggerated serration, as in muricids such as Chicoreus, Hexaplex, Concholepas, Neorapana, Jaton and Ocinebrina; the pisaniine buccinids Cancellopollia, Cantharus, Gemophos basidentatusVermeij, 2006 and Pollia; the nassariid Hebra; members of the Pseudolividae; the Eocene melongenid Cornulina; and the ranellid Argobuccinum pustulosum (Lightfoot, 1786). In other cases, however, the formation of the labral tooth differs from that of serial serrations on the same shell. In many ocenebrine muricids, for example, the labral tooth lies at the end of an external groove (interspace), whereas serrations tend to be expressed as extensions of cords. This is the condition in Pterorytis, Muregina and members of the Acanthina group (Acanthina, Acanthinucella, Chorus, Herminespina and Spinucella). In many fasciolariids, the labral tooth lies at the end of a conspicuous cord (as in Benimakia, Dentifusus and Leucozonia), whereas serial serrations are associated with interspaces. In species of the turbinellid genus Columbarium and in the olivid subfamily Ancillariinae, a cord-associated labral tooth occurs in the absence of serial serrations. Development of the labral tooth can therefore often be independent of the formation of serial projections on the outer lip.
In contrast to bivalves, in which serration along part of the valve margins may be asymmetrical (Vermeij, 2013), the serrations on the gastropod outer lip usually have similar adapical and abapical slopes. Rare exceptions include the denticles on the lip of the nassariid Alectrion papillosus, which are oriented abapically, and the anterior-most denticle of the strombid Tridentarius dentatus, which likewise faces forward. Laterally directed spines or nodes at the outer lip, by contrast, are very often adapically oriented, as in numerous muricids and in strombids of the genera Lambis and Harpago. In gastropods where the spiral cords have a shallower slope abapically than adapically, as in the cerithiid genus Rhinoclavis, the corresponding serrations are symmetrical.
These considerations indicate that extensions of the outer lip such as serial projections and labral teeth are not merely byproducts of spiral sculpture and the corresponding folded mantle margin. Instead, they show considerable architectural independence from the expression of external and internal spiral elements, and therefore invite a functional interpretation.
Functional considerations
The wide ecological distribution of outer-lip serration in gastropods implies that functions of serration are likely to be diverse. In hard-bottom gastropods, serration characterizes species that do not use rapid escape responses to such as slow predators as sea stars and predatory gastropods. Serrated hard-bottom gastropods are slow crawlers that, when confronted by a slow predator, tend to respond either by clamping firmly to the substratum or by withdrawing the soft parts into the shell. For example, whereas non-serrated turbinids and trochids typically adhere weakly to rock or algal substrata, and often exhibit well developed flight responses to sea stars, the species of Marmarostoma (Turbinidae) with a serrated outer lip clamp tightly. In these species of Marmarostoma, as well as in muricids, which also respond to disturbance by clamping forcefully to the rock surface, downward-pointing projections on the outer lip are often substantially worn in older individuals, indicating frequent shearing between the shell and the substratum. An ability to resist shear of this kind would be useful not only against forces associated with intense wave action, as in the Indo-West Pacific Marmarostoma setosum and the Caribbean fissurellid Cremides nodosa, but also against powerful predators such as crabs, spiny lobsters, some muricids, sea stars and some fishes, which are able to dislodge firmly clinging prey. Serrations would presumably improve the grip, or at least the frictional resistance, between the shell and the substratum in tightly clamping gastropods. The serrations in Marmarostoma, many muricids and members of the Cantharus group of pisaniine muricids are distributed along much of the length of the outer lip, but they are best developed along the abapical sector, where resistance against dislodgement may be most critical.
Other hard-bottom gastropods with serrations cling less firmly to the substratum and tend not to exhibit wear at the lip edge. This is the condition in the cerithiid genus Cerithium, many serrated buccinoideans, fasciolariids, costellariids, mitrids, vasids, tonnnoideans and littorinids. The function of these serrations remains obscure, especially when the projections are rounded rather than pointed. In species with sharp projections, as in most peristerniine fasciolariids and many pisaniine buccinids of the Cantharus group, it is possible that the gastropod can move its shell back and forth during an encounter with a soft-bodied predator so that the projections rub against and dislodge the offending enemy.
At least two groups of parasitic gastropods have evolved serration in a few genera. This is the case in three genera of ovulids from the Indo-West Pacific, which presumably parasitize anthozoan cnidarians (although specifics remain to be determined), and scleractinian coral-dwelling coralliophiline muricids. In the latter group, the fine serrations of the sharp adult lip are often conspicuously worn. Frictional resistance may be important to these gastropods, although it must be noted that other parasites of hard corals and gorgonians lack serrations, as in epitoniids, most ovulids, pediculariids and many coralliophilines.
Terrestrial gastropods often establish a tight seal between the shell lip and the substratum, especially when hibernating or aestivating, but they generally lack serrated lip margins. Instead, as noted by Boss (1974), many land snails have a planar, outwardly reflected or expanded, smooth-edged outer lip, which together with the rounded or ovate outline of the aperture and the presence of an operculum or epiphragm makes for a tight seal. Similarly expanded lips characterize some freshwater lymnaeid and planorbid pulmonates, which can also endure significant periods of suspended animation. The same lip characteristics occur in tree-dwelling intertidal potamidid cerithioideans and in some littorinids. Limiting water loss by creating a tight seal may be more important for these gastropods than resisting shear through friction.
In many serrated sand-dwellers, the serrations are concentrated on the abapical sector of the outer lip. This is the condition in most serrated Strombidae, Cassidae, Melongenidae, Nassariidae, Costellariidae, Mitridae, Volutidae, Harpidae and Conidae. Most of these sand-dwelling gastropods are highly mobile, and strombids execute rapid escape responses from predatory gastropods. It is possible that sharp abapical serrations serve a defensive function against predators with soft feeding organs, such as gastropods. Savazzi (1991) suggested that the abapical spines in the strombid Tridentarius dentatus might discourage fishes. Whether a similar interpretation applies to species in which pointed or rounded serration are arrayed along the whole length of the outer lip is unclear. This condition occurs in some species of the cassid genus Casmaria, the tonnid genus Malea, several mitrids including Mitra papalis, some photine buccinids and (as broad waves) in the strombid genus Gibberulus.
Downwardly directed spines on sand-dwelling gastropods are today almost entirely confined to the Indo-West Pacific region (Vermeij, 1999, 2001; Landau & Vermeij, 2011). Examples in tropical America are known mainly in fossil volutids and harpids, and in the living eastern Pacific Malea. It is particularly striking that, whereas pointed serrations occur in many Indo-West Pacific nassariids and conids, they are unknown in any tropical American species in these large groups. This finding is consistent with the greater development of shell defences in the Indo-West Pacific than elsewhere in the world (Vermeij, 1993).
CONCLUSIONS
The study of lip serration in gastropods highlights several directions for further study. Previously overlooked features of the lip edge may be taxonomically informative and imply the possibility of distinguishing hitherto undetected clades. Recognition of diversity at the lip edge invites further research on relationships among shell geometry, shell growth and accretion, and the mantle edge. Finally, interactions between the shell and the environment, including predators, often involve the edge of the outer lip, whose characters must often reflect functions that have scarcely been investigated.
ACKNOWLEDGEMENTS
I am grateful to Frank P. Wesselingh for photographing specimens at Naturalis in Leiden, and to Janice Fong for assembling the images into figures. Alyssa Henry and Edith Zipser provided valuable technical assistence. I thank Marco Oliverio and an anonymous reviewer for helpful comments on the manuscript.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}