






Summary
The concept of the ‘unique myocardial band’, which proposes that the ventricular myocardial cone is arranged like skeletal muscle, provides an attractive framework for understanding haemodynamics. The original idea was developed by Francisco Torrent-Guasp. Using boiled hearts and blunt dissection, Torrent-Guasp created a single band of ventricular myocardium extending from the pulmonary trunk to the aortic root, with the band thus constructed encircling both ventricular cavities. Cooked hearts can, however, be dissected in many ways. In this review, we show that the band does not exist as an anatomical entity with defined borders. On the contrary, the ventricular cardiomyocytes are aggregated end to end and by their branching produce an intricate meshwork. Across the thickness of the left ventricular wall, the chains of cardiomyocytes exhibit a gradually changing helical angle, with a circumferential zone formed in the middle. There is no abrupt change in helical angle, as could be expected if the wall was constructed of opposing limbs of a single wrapped band, nor does the long axis of the cardiomyocytes consistently match with the long axis of the unique myocardial band. There are, furthermore, no connective tissue structures that could be considered to demarcate its purported boundaries. The unique myocardial band should be consistent with evolution, and although the ventricular wall of fish and reptiles has one or several distinct layers, a single band is not found. In 1965, Lev and Simpkins cautioned that the ventricular muscle mass of a cooked heart can be dissected almost at the whim of the anatomist. We suggest that the unique myocardial band should have ended there.
INTRODUCTION
An accurate description of the architectural arrangement of the cardiomyocytes making up the walls of the left ventricle is fundamental to the understanding of myocardial function. Ventricular mural anatomy has been extensively studied over the last 400 years, but debate continues regarding the precise pattern of the architectural arrangement of the cardiomyocytes aggregated together within the walls. An intriguing concept was introduced by Torrent-Guasp [1]. He postulated the existence of a ‘unique myocardial band’ or ‘helical ventricular myocardial band’ which was wrapped in such a way as to produce a ‘helical heart’ [2]. The concept has since been developed by some so as to provide explanations for many aspects of surgical cardiac disease [3–5], even being cited recently to explain the actions of cardioplegia [6]. None of these multiple publications, however, including the initial studies of Torrent-Guasp, have been validated by histological studies. In contrast, there is a wealth of anatomical evidence, including multiple histological investigations, which demonstrates the cardiomyocytes to be aggregated together to form a 3D mesh. This arrangement is much more complex than the structure envisaged by proponents of the ‘unique myocardial band’. Here, we review the anatomy of the ventricular mass so as to demonstrate the multiple shortcomings of the concept of a unique myocardial band. In an accompanying article, we review several lines of evidence from physiology which, similar to this article, do not lend support for the notion of the unique myocardial band.
THE BEGINNING OF THE ‘UNIQUE MYOCARDIAL BAND’
On the basis of gross dissections, Torrent-Guasp proposed that the walls of the right and left ventricles exist as a continuous myocardial band, which extends from the root of the pulmonary trunk to the root of the aorta [1]. The muscular band thus sculpted from the ventricular mass was held to form 2 loops, which surround the cavities of both the right and left ventricles. The first, or basal, loop commences at the pulmonary valve and consists of the right ventricular free wall. It leads to the outer layer of the left ventricular free wall and hence onto the right ventricular side of the ventricular septum. The band then continues as the second, or apical, loop, which has a descending segment comprising the inner left ventricular free wall and an ascending segment. The latter segment then forms the subepicardial component of the apex and the rightward component of the ventricular septum, terminating at the aortic root. The cardiomyocytes aggregated together within the band were held to be aligned along its long axis, so that the ascending and descending segments crossed in approximately perpendicular fashion within the septum. Cleavage planes, or sliding surfaces between the segments, were alleged to exist, thus allowing the parts of the band to move across one another. The purported physiological consequences of this arrangement were subsequently described in numerous publications [5, 6], including an account of cardiac development [7].
THE DISSECTION PROCESS USED TO CREATE THE ‘UNIQUE MYOCARDIAL BAND’
Torrent-Guasp developed his technique of dissection over a period of 25 years [1, 3]. The hearts were boiled for a prolonged period, softening the myocardium so as to permit subsequent dissection [6]. The anterior free wall of the right ventricle, along the anterior inter-ventricular groove, was prised off the ventricular septum, using the thumb to cut through the cardiomyocyte aggregates and connective tissues. The pulmonary trunk was thus detached from the root of the aorta, with the blunt dissection extended to the apex of the right ventricle to open the right ventricular cavity. At the inferior limit of the right ventricular cavity, at the junction of the right ventricular free wall and ventricular septum, the blunt dissection was continued into the mid-zone of the left ventricular wall, and towards the root of the aorta, thus exposing the inner helical zone. The left fibrous trigone was cut, permitting the creation of a plane in the middle of the ventricular septum. The dissection was continued between the inner and the outer helical zones, followed by cutting the right fibrous trigone and freeing the aorta. Unfolding the dissection thus created an allegedly unique myocardial band, extending from the pulmonary trunk to the aortic root. This concept of a continuous myocardial band, with attachments at the arterial roots, is attractive, because it permits direct comparison with skeletal muscles, whose myocyte aggregates do indeed run between points of origin and insertion formed by connective tissue or bony structures. Most skeletal muscles, however, are enclosed in fibrous sheaths, thus permitting their dissection along identifiable boundaries. As has been emphasized previously [7], this is not the case for the ventricular myocardial cone.
HISTOLOGICAL AND ANATOMICAL PERSPECTIVES
The essence of skeletal muscles is that the extent of each entity can readily be revealed by anatomical dissection. None of the histological studies of the myocardium that we are aware, in contrast, have provided any evidence for an origin and insertion as described for the alleged unique myocardial band [8, 9]. Already in 1864, Pettigrew [10] had emphasized that the myocardial mass was not arranged like skeletal muscle, instead describing the arrangement of cardiomyocytes within the ventricles as aggregated to form multiple interleaving sheets. Nearly 40 years later, Krehl [11] showed how some of the cardiomyocytes were aligned in circumferential fashion within the middle zone of the left ventricular walls. These histological findings were subsequently confirmed by Feneis [8] and by Greenbaum et al. [12]. Dissection of the left ventricle shows the gradual transition in the myocardial grain between the inner, the middle and the outer zones (Fig. 1) [13]. Sanchez-Quintana et al. [14] were able to show the presence of the circumferential cardiomyocytes in human hearts using anatomical dissection (Fig. 2). LeGrice et al. [15] subsequently showed how the cardiomyocytes themselves were aggregated together in sheet-like configurations. These aggregated entities, or lamellar units, however, do not extend across the ventricular wall, as was suggested by the diagrammatic depiction originally provided by LeGrice et al. [15]. Instead, the aggregated cardiomyocytes throughout the ventricular walls are bound together both by their meshed branching and by the endomysial component of the fibrous matrix, being separated by spaces containing loose perimysial tissue, arteries and veins [16]. This arrangement, validated by histology, is incompatible with the concept of the ‘unique myocardial band’. The intercellular spaces and myocytic branches are ubiquitous within the walls, failing to provide the boundaries needed to produce an anatomically discrete myocardial band.
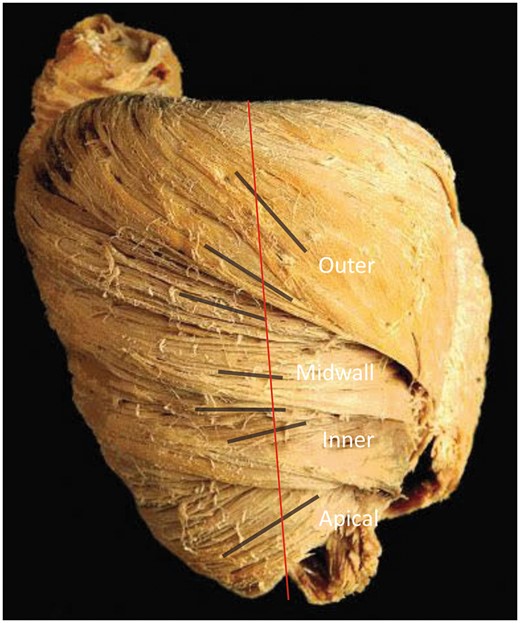
Macroscopic anatomy of the porcine heart. Blunt dissection of the heart shows the gradual transition of the helical angle of the inner, the mid-wall and the outer zones (black lines) compared with the long axis (red line). Figure is provided by P.P. Lunkenheimer and reproduced with permission.
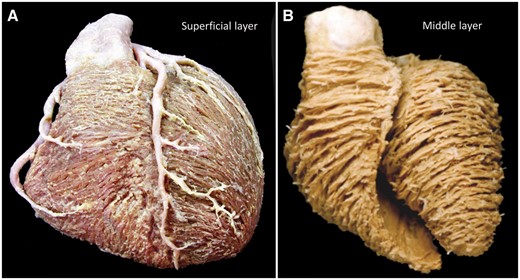
Macroscopic anatomy of the human heart. As seen in (A), the dissection of a human heart reveals that the cardiomyocytes of the superficial layer are common for both ventricles, descending obliquely on the sternocostal and diaphragmatic aspects to the apex, crossing the inter-ventricular grooves. The course of the cardiomyocytes around the right ventricle is more circumferential than in the left ventricle. (B) The grain of the middle layers of both the right and the left ventricles is orientated in a second direction.
Although providing no evidence to validate the notion of the unique myocardial band, the histological evidence does support the presence of helically arranged chains of aggregated cardiomyocytes. Indeed, a well-recognized study had long since demonstrated the progression of such helical angulations when traced through the thickness of the left ventricular walls [17]. The arrangement is that of a left-handed outer helix, with negative angulation relative to the ventricular equator, progressing through a region of zero angulation at the mid-wall and continuing as a right-handed helix, with positive angulation, towards the inner endocardial ventricular surface [17]. This gradual change in helical angulation exists in all the regions of the ventricular walls, including the ventricular septum [17]. In the initial study [17], all the cardiomyocytes, despite their change in helical angulation, were reported to be aligned in more-or-less tangential fashion when assessed relative to the epicardial ventricular surface. Subsequent histological investigations, which used circular knives to cut tissue blocks from the ventricular walls, thereby cancelling the effect of the helical angle on the orientation of the cardiomyocytes within the sections transferred to the microscope slides, revealed that significant numbers of cardiomyocytes deviated from the tangential plane [18, 19]. Investigations using pneumatic dissection of the ventricular walls [20] then showed how it was possible to disrupt the weaker perimysial component of the fibrous matrix. These manoeuvres confirmed that the cardiomyocytes were aggregated together to form an intricate 3D meshwork. Histological findings, therefore, provide further evidence of a complex mural ventricular structure (Fig. 3). None of these investigations have provided any evidence of an alignment of the cardiomyocytes that follows the course of the unique myocardial band. All of the studies, in contrast, have shown a relatively uniform pattern of aggregation throughout the ventricular circumference [8]. The only study that we are aware to have produced the band by following the directions of Torrent-Guasp [1] [2], and then sectioning it histologically, failed to find correlation with the long axes of the band and its contained cardiomyocytes [21].
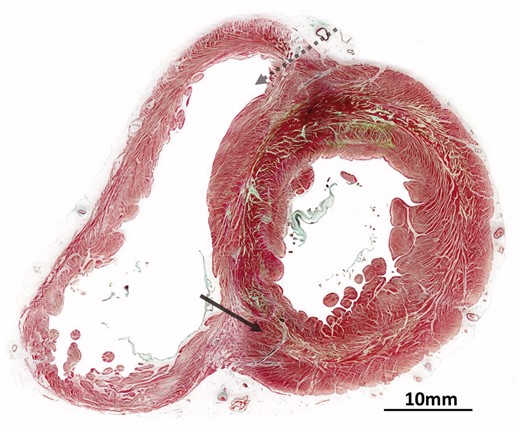
Microscopic anatomy of the human heart. Cross-section of human heart at the equator stained using Masson’s trichrome technique. The dashed arrow indicates the region where the dissection is performed by Torrent-Guasp damages cardiomyocytes while entering the right ventricle. The solid arrow shows the disruption required within the mid-zone of the circumferentially orientated cells. The presence of aggregated units is evident, forming chevron-like structures around most of the ventricular circumference.
HIGH-RESOLUTION COMPUTED TOMOGRAPHY
Recent advances in microcomputed tomography, using iodine enhancement of myocytic and vascular structures, have provided images of sufficient spatial resolution to reveal the alignment (Fig. 4) and dimensions of the chains of individual cardiomyocytes (Fig. 5), along with the pattern of the units produced in consequence of their aggregation within the endomysial weave of the supporting fibrous matrix [16, 22, 23]. The computed tomography images (Fig. 5) confirm the presence of the chevron-like configurations revealed histologically (Fig. 3), which exist within the setting of a relatively smooth helical trans-mural arrangement (Figs 4 and 5). These images do not show the abrupt changes in angulation at the mid-wall that would be expected were the ventricular cone based on the postulated wrapped myocardial band (Figs 4 and 5). Instead, they support the notion of the complex mural mesh, with units exhibiting both helical and trans-mural angles, complex heterogeneous morphologies and multiple connections to adjacent units via myocytic chains [16].
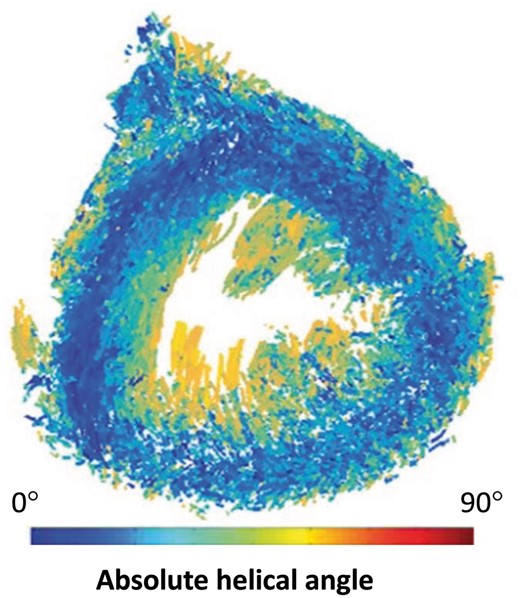
Eigen analysis of the human heart using microcomputed tomography. Angle maps are viewed in short axis, and colour bars indicate cardiomyocyte helical angulation. Note the cardiomyocytes with a helical angle close to zero (blue) encircling the entire circumference of the mid-ventricular wall.
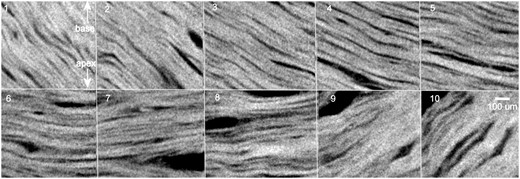
Trans-mural tangential reconstructions using high-resolution microcomputed tomography. Cardiomyocyte aggregates in the subepicardial (1), outer (2–4), mid-wall (5–7), inner (8–9) and subendocardial zones (10) from a rabbit heart are shown. Similar transitions are seen around full circumference of the left ventricle. A gradual transition in angulation that is not compatible with the concept of the ‘unique myocardial band’ is shown. Spatial resolution is approximately 6 µm.
EMBRYOLOGICAL PERSPECTIVE
The ontogenetic development of the human heart is complicated. In a review that sought to correlate development with the notion of the helical heart, we were asked to envisage that ‘a simple and integrated triple figure-eight spiral band, with 3 S-shaped helixes and their apices may correlate the conventional embryologic development of the primitive heart (bulbus cordis, ventricle and arterial outflow vessels)’ [7]. Much has been learned regarding cardiac development since the publication of this review. We now know that the original linear heart tube forms little more than the definitive left ventricle [24]. New material is added at the arterial pole from the heart-forming areas to form the right ventricle and the outflow tract. Similar new growth at the venous pole produces the atrial chambers and the venoatrial connections [25]. In terms of development of the ventricular mass, initially the walls are made up predominantly of a meshwork of luminal trabeculations, with minimal formation of a compact layer. At the early stages of development, subsequent to looping of the heart tube, the atrial chambers connect to the developing left ventricle, whereas the outflow tract is supported above the developing right ventricle. The rightward expansion of the atrioventricular canal then brings the right atrial cavity into communication with the cavity of the right ventricle [26]. After this process, which occurs during the 12th day of development in the mouse, the developing outflow tract, which is beginning its separation into the aortic and pulmonary roots, remains supported by the developing right ventricle. The left ventricle at this stage, therefore, connects to the developing aortic root through the embryonic inter-ventricular communication. It is only subsequent to transfer of the aortic root to the left ventricle that there is closure of the inter-ventricular communication. Even at this stage, which has occurred by the 14th day of murine development, there has been minimal growth of the compact layers of the ventricular walls. The rate of proliferation of the compact myocardium, and the compact component of the ventricular septum, is known at this stage to exceed that of the trabeculated myocardium [27, 28]. At this stage, it is then possible to recognize the aggregation of the individual cardiomyocytes into units of various shapes and dimensions, with the units separated by perimysial spaces throughout the circumference of the walls. When assessed in long axis, many of the units show the sheet-like configuration emphasized by LeGrice et al., although none of the aggregated units extend in full trans-mural fashion (Fig. 6A). When assessed relative to the short axis of the ventricular cone, the aggregates show an obvious circumferential arrangement in the middle component of the wall, with the parietal left ventricular aggregates extending into the ventricular septum (Fig. 6B). The perimysial spaces are not positioned in such a way as to permit unwrapping of the alleged myocardial band. On the contrary, the overall arrangement of the walls is very much that of a complex 3D mesh. Molecular identification of the compact wall and ventricular septum, besides being possible using proliferation markers, can also be made based on expression of Hey2 and N-myc [29]. CHF1/Hey2 plays a pivotal role in left ventricular maturation through suppression of ectopic atrial gene expression [29, 30]. Neither of these genes nor indications of proliferation by Brdu incorporation or expression of Ki-67 and PCNA [25, 30, 31] give any indication of distinct bands in the compact wall compatible with the postulated helical heart. A recent developmental study, furthermore, suggested the anterior ventricular septum to be formed from a merger between the embryonic left ventricle and the outflow tract [32]. This arrangement provides no support for the opening of the ventricular wall in the manner of Torrent-Guasp.
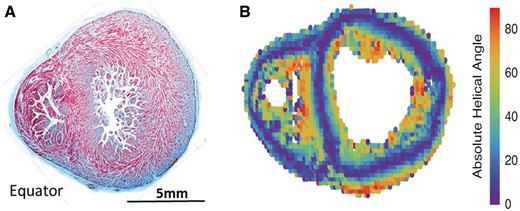
Embryological findings. (A) A haematoxylin–eosin staining cross-section in a human foetus of 20 weeks of development. The changing orientation from radial lamellae to circular orientation, with the formation of chevrons is noted. There is no evidence of the ‘edges’ that would be required to support the notion that the walls are made up of a wrapped band nor evidence of fibrous partitions separating the components of the alleged band. A short-axis map of the helical orientation of the cardiomyocytes using diffusion-tensor magnetic resonance imaging in a human heart at 24 weeks of gestation is shown (B). Colour coding indicates helical angle, such that blue indicates circumferentially orientated cardiomyocytes.
COMPARATIVE ANATOMICAL PERSPECTIVE
It has been suggested that the unique myocardial band is compatible with the overall trends of evolution of the heart [7]. Mammals evolved from ectothermic (cold-blooded) vertebrates, but the orientation of cardiomyocytes in these species has received limited attention [33, 34]. We do know that, in some fish and most amphibians, the compact wall may be so thin that the epicardium almost touches the ventricular lumen. In this setting, the compact wall consists of 1 layer only (Fig. 7). In some highly active animals, such as tuna fish, in contrast, the compact wall is well developed and may consist of 2 or 3 layers. The number of layers generally appears to increase with the width of the compact wall [35]. At least in fish, the cardiomyocytes of the distinct compact layers may be at almost right angles to each other [10, 36, 37], and the different myocardial layers may be so distinct that they are easily separated [35, 38, 39]. In fish, nonetheless, we have never observed findings to support the concept of a myocardial band that connects the atrioventricular orifice with the conoventricular, or bulboventicular, orifice. Reptiles, which may be considered to represent the ancestral state of mammals and birds, have a variable number of compact layers, like in fish, but generally there are 2 or 3 layers [37, 40–42] The innermost layer is the interface between the compact wall and the trabeculated, or spongy, interior wall. It is the thinnest of the compact layers when there is a sizeable compact wall [41, 42]. The ventricular compact wall of many reptile species has 2 zones, distinguished by the orientation of the cardiomyocytes. The smooth progression of the helical angle within the depth of the ventricular walls (Figs. 1, 4 and 5), in contrast, is a feature of all mammalian species studied to date, regardless of their size, which suggests a common geometric environment for the cardiomyocyte. This is because the geometric interplay between the inner and the outer surfaces of the left ventricle, with mural thickening, is an expression of the ratio of wall thickness to chamber size and not their absolute dimensions [13]. The ventricular wall of ectothermic vertebrates may have substantial deviations from the architectural arrangement of 2 distinct layers. Many species, however, do have a bilayered compact wall, but the 2 layers appear largely distinct, not unlike a Russian Matryoshka nesting doll (Fig. 7). Unwinding such layers would produce an outer and an inner shell, not a single band. It follows that the ‘aberrant fibres’ that are initially disrupted in the unwinding of Torrent-Guasp are in fact an evolutionarily old part of the ventricle (Fig. 7). We propose that ‘aberrant fibres’ is a spurious concept, and the disruption of circumferential compact myocardium immediately invalidates the significance of unwinding of the heart.
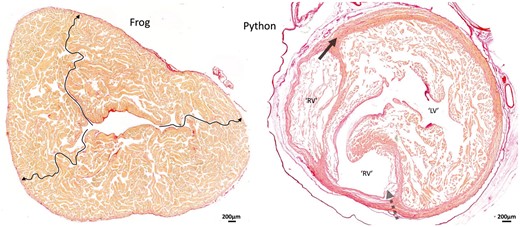
Comparative anatomy. Ventricular architecture in a frog and a snake illustrated by 10-μm-thick transverse sections stained with picrosirius red. The (contracted) ventricle of the Xenopus frog exemplifies the highly trabeculated ventricle found in many fish and amphibians. The arrows show 3 pathways where no myocardium was crossed and by which the extremely thin outer compact layer can be reached from the central lumen. Distinct layers to the compact wall are not recognized in such settings, and even if they are, the functional implications would be proportional to the mass of the layers, i.e. miniscule. The ventricle of pythons has a high-pressure LV and a low-pressure RV surrounded by 2 distinct layers of compact myocardium. (This ventricle was fixed in diastole.) Where the dashed arrow is placed (as in Fig. 2), much of the compact wall is made up of approximately circumferentially oriented myocardium. The presence of this myocardium suggests an old evolutionary origin to the so-called ‘aberrant fibres’ that has to be disrupted initially in the Torrent-Guasp procedure. We propose that ‘aberrant fibres’ is a spurious concept and further propose that the disruption of such commonly found circumferential compact myocardium immediately invalidates the significance of unwinding of the heart. LV: left ventricle; RV: right ventricle.
CONGENITAL HEART DISEASE PERSPECTIVE
There are multiple congenital lesions that point to the lack of credibility of a concept depending on the presence of a unique myocardial band extending from the pulmonary to the aortic roots and encircling both ventricular cavities. In the first instance, it is difficult to envisage how such a concept would be compatible with the development of a heart having double inlet to, and double outlet from, the right ventricle. In this setting, the left ventricle is no more than a hypoplastic apical component. It is equally difficult to envisage how the notion of a band extending from the pulmonary to the aortic roots would be compatible with the presence of a common arterial trunk. It is similarly difficult to explain hearts having either aortic or pulmonary atresia, not to mention the fact that, in Ebstein’s malformation, the location of the alleged passage of the myocardial band from the right to the left ventricle inferiorly can be paper-thin due to atrialization of the inlet component of the right ventricle. The presence of a myocardial band should have important implications for the development of congenital heart disease. We are unable to find any such evidence. Dissections performed in congenitally malformed hearts also confirm the presence of the cardiomyocytes aligned in circumferential fashion, a feature denied by some of the proponents of the band [43]. The circumferential cardiomyocytes, furthermore, were present in the hypertrophied walls of the right ventricle in a heart obtained from a patient with tetralogy of Fallot (Fig. 8).
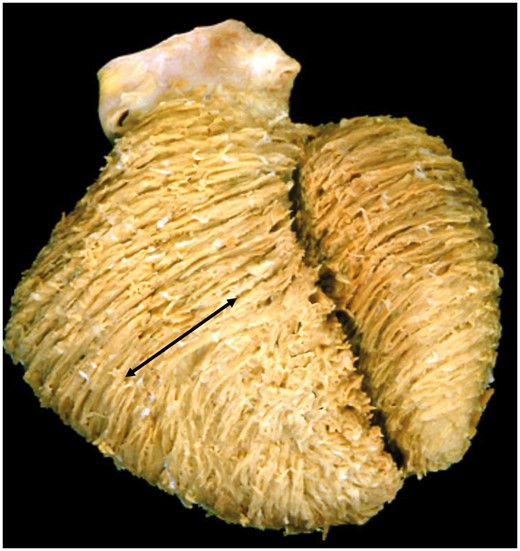
Ventricular architecture in tetralogy of Fallot. The dissection reveals the macroscopic features in the setting of tetralogy of Fallot, showing a middle layer with a circumferential orientation in the right ventricle (arrow). The presence of circumferential cardiomyocytes in a direction perpendicular to the direction of the ‘basal loop’ is incompatible with the concept of a ‘unique myocardial band’.
CURRENT UNDERSTANDING OF MYOCARDIAL STRUCTURE
The wealth of data available from the techniques and approaches discussed above shows that the myocardial walls are made up of cardiomyocytes aggregated together to produce a 3D meshwork of interconnected units. The average orientation of cardiomyocytes shows a gradual change in their helical angulation, with a middle component having approximately zero angulation relative to the ventricular equator. There are no gaps in the assembly of the units that would permit a dissector to begin to reveal the presence of a unique myocardial band. Nor are there planes of cleavage that would permit the tracing of such a band from the pulmonary trunk to the aortic root, particularly when note is taken that the outer wall of the left ventricle is held, by proponents of the band, to contribute to both its basal and apical loops. Lev and Simpkins [44] had already emphasized, in 1965, that under the conditions employed by Torrent-Guasp, the ventricular muscle mass can be dissected almost at the whim of the anatomist. We do not deny that, with skill and practise, it is possible to unravel the heart to produce a continuous myocardial band. Indeed, it is now possible to observe Torrent-Guasp producing the strip on an online video [2]. The important question is whether Torrent-Guasp is producing the band according to his own preconceived notions or on the basis of the accepted techniques for anatomical dissection. When analysing the arrangement of adjacent skeletal muscles, relatively uniform dissections are produced simply by delimiting the boundaries of the individual muscles, with the skeletal myocytes aggregated together within the epimysial sheets. Such an approach is not possible when considering the ventricular cone, because the cardiomyocytes within the ventricular walls are aggregated together by the endomysial components of the fibrous matrix and by their own branched connections via the intercalated discs. The aggregated units themselves are separated by perimysial spaces, with the overall walls enclosed between the epicardial and the endocardial boundaries. There are no obvious planes of cleavage that permit delimitation of anatomically defined tracts or subunits within the walls. Observation of the approach taken by Torrent-Guasp reveals that the initial separation of the right ventricle tears away its parietal wall from the ventricular septum, disrupting what are described as ‘aberrant fibres’ (Fig. 3) [2]. They are, of course, only ‘aberrant fibres’ according to the preconception of the band. In reality, these cardiomyocytes can be shown, by following the grain produced by the aggregated chains, to form a myocardial component common to both ventricles [12, 14]. Having reached the inferior inter-ventricular groove, Torrent-Guasp then alleges to show loops of the band that encircle the left ventricle. The plane developed by Torrent-Guasp is within the aggregated cardiomyocytes that surround the cavity of the left ventricle in circumferential fashion. Such circular cardiomyocytes, denied by current proponents of the band, are well demonstrated by dissections made following the overall alignment of the aggregated units (Fig. 1). Detailed anatomical analysis, therefore, using all available techniques, contradicts the hypothesis of the band.
CONCLUSIONS
There is extensive experimental evidence to show that the ventricular walls are made up of an intricate 3D network of aggregated cardiomyocytes. Apart from the questionable blunt dissections performed by Torrent-Guasp, there are no direct or indirect observational data to support the concept of a compartmentalized ventricular myocardial band that extends from the pulmonary trunk to the aorta (Supplementary Material, Table S1). We submit that the notion of a unique myocardial band is anatomically spurious. Taken with our second review [45], we conclude that the value of the band as an explanation of ventricular structure, function and cardiac pathophysiology has come to an end.
SUPPLEMENTARY MATERIAL
Supplementary material is available at EJCTS online.
ACKNOWLEDGEMENTS
We are grateful to Bastiaan J.D. Boukens, Henggui Zhang and Camilla Omann for reviewing and commenting on the article and to P.P. Lunkenheimer for providing Fig. 1.
Funding
This work was supported in part by the European Union’s Horizon 2020 research and innovation programme [707663].
Conflict of interest: none declared.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}