



Abstract
A human body part, such as a foot, may be observed from an egocentric perspective (consistent with looking at one's own body, e.g. top of the foot, toes pointing up) or from an allocentric perspective (only consistent with looking at someone else, e.g. top of the foot, toes pointing downwards). We found that the right extrastriate body area (EBA) response to images of body parts was enhanced for body parts presented from an allocentric perspective. Other areas of extrastriate cortex which responded robustly to images of bodies, including the right lateral occipital complex, right MT and left EBA, nevertheless did not distinguish between the two perspectives. A region of primary somatosensory cortex showed the reverse selectivity: the blood oxygen level-dependent response to body parts presented from an allocentric perspective was suppressed. These results help to illuminate the integration of visual and tactile information by which the brain identifies seen body parts as belonging to the self or to another person.
Introduction
The extrastriate body area (EBA) is a region of human visual cortex in which the blood oxygen level-dependent (BOLD) response is significantly higher when the subject is viewing images of human bodies and body parts relative to other categories of visual stimuli, including familiar inanimate objects and even human faces (Downing et al., 2001). One aim of the current study was to further characterize the response of the EBA, and in particular to functionally distinguish the EBA from several other regions of extrastriate cortex that respond robustly, though not selectively, to images of body parts.
Neighbouring the EBA is another visual region, MT, which responds more to moving than to stationary stimuli (e.g. Tootell and Taylor, 1995). Previous studies have noted, however, that the human region MT also responds more to images of human bodies relative to images of familiar non-human objects (Downing et al., 2001), in addition to any effect of implied motion in the image (Kourtzi and Kanwisher, 2000). In the same vicinity of extrastriate cortex is a third functional region, the lateral occipital complex (LO, Malach et al., 1995; Grill-Spector et al., 1998a,b, 2001; Kourtzi and Kanwisher, 2001). The LO responds robustly to images of familiar and unfamiliar objects, including human bodies.
As the preferred stimuli for each of these regions include human body stimuli, it was of interest to determine whether they shared functional or anatomical profiles with EBA. Here, a localizer experiment was used to define regions of interest (ROIs) for each functional region using an appropriate contrast of visual stimuli. Then the location and response to body and body part images was compared across the ROIs.
A second portion of the study, the body perspective experiment, was designed to test whether the EBA distinguishes between body parts presented from egocentric versus allocentric perspectives. Preference for an egocentric perspective would be consistent with a primary role for the EBA in visuomotor control (see Astafiev et al., 2004). Enhanced response to bodies presented from an allocentric perspective, on the other hand, would suggest that the EBA is preferentially attuned to the bodies of others, perhaps as an input to higher-level social cognition.
Materials and Methods
Participants
Ten healthy, right-handed adults volunteered or participated in this study for payment. All subjects had normal or corrected to normal vision and gave informed consent to participate in the study.
Equipment
Subjects were scanned in the Siemens 3 T scanner at the MGH-NMR Center in Charlestown, MA, using a head coil. Standard echoplanar imaging procedures were used (TR = 2 s, TE = 30 ms, flip angle 90°). Twenty-six 4 mm thick axial slices covered the whole brain, excluding the cerebellum.
Materials
Each subject was scanned on a localizer experiment and on the body perspective experiment. The localizer experiment included four conditions. The first two were greyscale photographs of (i) whole bodies, excluding heads, and isolated body parts; and (ii) familiar inanimate objects (e.g. car, drum, tulip) and object parts (e.g. hammer head, screwdriver handle). There were 30 exemplars from each category (15 of whole bodies or objects and 15 of body or object parts). Images were presented at a rate of one every 800 ms (stimulus duration = 500 ms, interstimulus interval = 300 ms).
The other two conditions were concentric dark and light grey rings, 0.4 or 0.6 cycles/degree, shown either (i) expanding and contracting or (ii) changing contrast but not position. In the motion conditions, the rings reversed direction of motion 0.75–1 times per second moving at 1.9 or 2.5°/s. In the contrast conditions, the stationary rings gradually changed contrast from 75% contrast difference to 15% contrast difference, and then back again. The motion localizer used here was slightly different from the standard localizer, which compares neural response to moving and stationary rings with no contrast change. The changing contrast in the current control condition was designed to reduce adaptation to the stationary rings, and so provide a more conservative localizer for human motion selective brain regions.
All stimuli were presented in a central square of the screen that subtended ∼12 × 12°. A black fixation dot was superimposed on each image. Each scan consisted of 16 experimental blocks (two blocks each from each condition, 16 s each) during which the subject passively viewed the images and 16 s fixation-only periods intervening every four blocks and at the beginning and end of the scan. Block order was counterbalanced within and between subjects. Subjects were scanned four times on the localizer experiment.
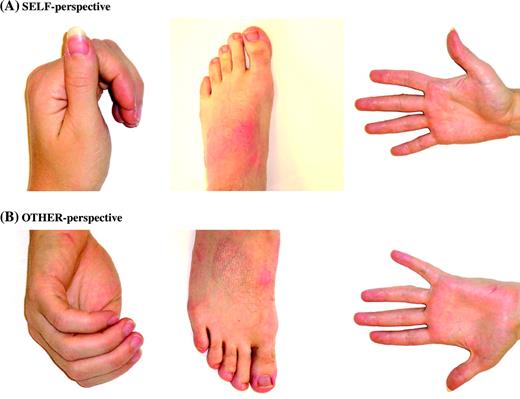
Stimuli for the body perspective experiment were forty full colour pictures of body parts (hands, arms, feet, and legs). Twenty of the images were photographed from the perspective of the viewer (‘egocentric’) and the other 20 were photographed from a perspective from which the viewer's own body would be inaccessible (‘allocentric’, see Fig. 1). Images in the two conditions were matched for luminance. All stimuli were presented in a central square of the screen that subtended ∼12 × 12°. Images were presented at a rate of one every 800 ms (stimulus duration = 500 ms, interstimulus interval = 300 ms). Images were presented in 16 s blocks of 20 images from the same condition. A 16 s fixation block intervened between each experimental block. The scan lasted 336 s. Subjects performed a one-back repetition detection task during the scan, pressing a button whenever the same stimulus appeared twice in immediate succession, which occurred twice per block.
{kind=link}
Sample stimuli for the body perspective experiment. (A) Images of body parts depicted from a perspective accessible to the model (egocentric perspective). (B) Body parts shown from a perspective inaccessible to the model (allocentric perspective). The stimuli in these two conditions have very similar low-level visual properties.
FMRI Analysis
MRI data were analysed using SPM 99 (http://www.fil.ion.ucl.ag.uk/spm/spm99.html) and in-house software. Each subject's data were motion corrected and normalized onto a common brain space (the MNI template). Data were then smoothed using a Gaussian filter (Full Width Half Maximum = 5 mm), and high-pass filtered during analysis. Every experiment used a blocked design, and was modelled using a boxcar regressor. Thus the reported response to a given condition was the average response across a whole block of that condition.
We carried out a whole brain random-effect analysis (P < 0.001 uncorrected, extent threshold = 3 voxels) comparing egocentric with allocentric conditions. Further analyses were conducted using a region of interest (ROI) approach.
Individual-subject ROIs for four regions (right and left EBA, right MT and right LO) were defined for each subject, based on the results of the localizer experiment. All ROIs were defined as the cluster of contiguous voxels in extrastriate cortex that was more active (P < 0.05 corrected) when the subject was looking at one condition compared to another. The EBA ROIs were defined as regions (bilaterally) that responded more to pictures of human bodies and body parts than to pictures of non-human objects and object parts. MT and LO were identified only in the right because (i) both regions were more observed more consistently on the right than on the left, and (ii) these regions were used as controls for the right EBA during the body perspective experiment. MT was defined as the voxels that responded more to moving rings than to stationary rings. The putative LO was defined as the region that responded more to non-human objects and object parts than to stationary concentric rings (contrast change condition). (This region was identified as putative LO, since the standard LO localizer is a contrast between familiar objects and scrambled objects or textures, not concentric rings as used here.)
Within each ROI, the average percent signal change (PSC) relative to fixation baseline, where PSC = 100 × [raw BOLD magnitude for (condition – fixation)]/[raw BOLD magnitude for fixation], was calculated for each condition (averaging across all voxels in the ROI, all TRs in the block, and all blocks of the same condition). This calculation yielded a single grand average PSC value per ROI for each condition. We calculated PSCs for each ROI for both the localizer experiment and the body perceptive experiment.
Results
Localizer Experiment
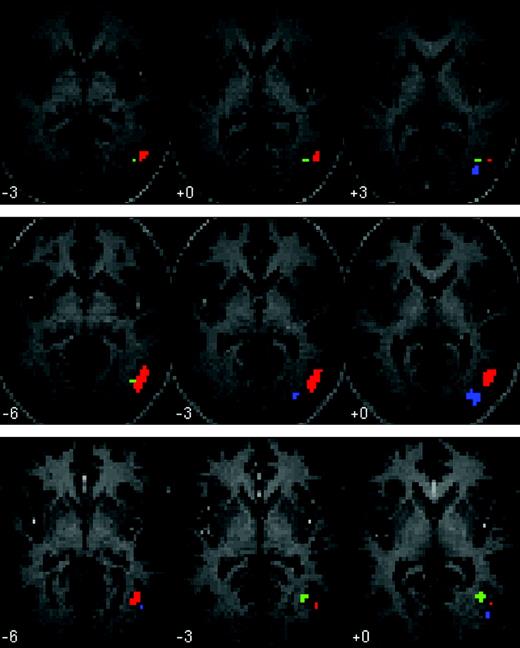
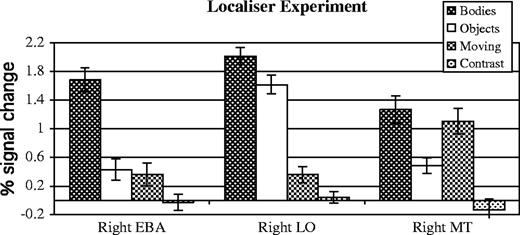
The EBA was defined in all 10 subjects (bilaterally in nine subjects, right only in one subject; see Fig. 2), as the region of extrastriate cortex that responded more when subjects viewed the images of bodies and body parts, relative to the inanimate objects and object parts (P < 0.05, corrected, Fig. 2). The average peak of the right EBA was [54 −66 3]. The average peak of the left EBA was [−45 −72 3]. As expected, the EBA showed a strong categorical response to images of bodies, compared with images of objects (Fig. 3).
{kind=link}
Individual subject ROIs for three sample subjects (P < 0.05 corrected). Three axial slices are shown for each subject. Red = EBA; green = MT; blue = LO. Left is shown on the left of each slice.
{kind=link}
Functional response of three ROIs in the localizer experiment. All of these regions respond robustly to images of bodies, but are clearly functionally distinguishable based on the response to the other three conditions.
Right MT was defined in eight subjects (average peak [48 −69 0]) as the region responding more to moving rings than to stationary rings changing contrast. MT neighboured but was anatomically distinct from the EBA (Fig. 2). The response in MT to the static images of human bodies was as high as the response to moving rings, and significantly higher than the response to images of objects (P < 0.05, paired samples t-test, Fig. 3). By contrast, in both the right and the left EBA, the response to moving rings was no higher than the response to pictures of objects.
The right LO was defined in eight subjects (average peak [45 −81 −3]) as the region responding more to the images of inanimate objects, relative to stationary concentric rings. LO was consistently posterior to the EBA and MT (Fig. 2). The functional profile of LO was qualitatively distinct from both the EBA and MT, showing a high response to bodies and objects, and a low response to the rings, whether moving or stationary (Fig. 3).
Body Perspective Experiment
In the body perspective experiment, full-colour photographs of body parts were presented in alternating blocks: ‘egocentric’ images were photographed from the perspective of the model; ‘allocentric’ images were photographed from a perspective inaccessible to the model, and therefore only consistent with looking at the body of another person. Subjects debriefed after the experiment were not aware of the contrast between egocentric and allocentric photographs. No subject noticed the block structure of the experiment, or was able to guess the manipulated variable. All of the subjects expressed surprise when the contrast of interest was described and then demonstrated.
Each of the regions defined in the localizer experiment was used as a region of interest for the body perspective experiment. As all of these regions responded preferentially to body images, the analysis within the regions of interest was used to detect any differences in the profiles of their responses to body images. Specifically, it would show whether a differential response to egocentric versus allocentric images was unique to one area or shared by all regions that responded to body and body part stimuli.
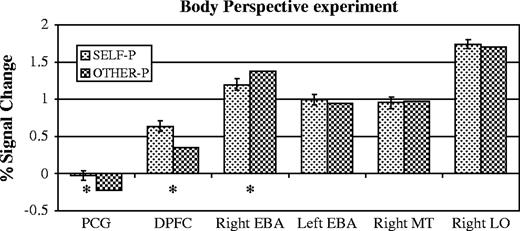
The right EBA response was higher when subjects looked at allocentric images (PSC 1.4) than egocentric (PSC 1.2, P < 0.05, paired-samples t-test, Fig. 4). The left EBA on the other hand did not distinguish between the two conditions (allocentric PSC 1.0, egocentric PSC 1.0, n.s.). A repeated measures ANOVA revealed a trend towards an interaction of condition by hemisphere in the EBA (P = 0.07).
{kind=link}
Results of the body perspective experiment. Asterisks mark comparisons significant in a paired samples t-test, P < 0.05. Bars show standard error of the difference. Independent individual subjects regions of interest were defined for the right EBA, left EBA, right MT and right LO. The data shown here were used to define the regions of interest for the post-central gyrus (PCG) and dorsal prefrontal cortex (DPFC), and so the magnitude of response in those two regions is illustrative only.
The right EBA preference for allocentric over egocentric images was unique in right extrastriate cortex as well. Right MT did not distinguish between the two conditions (allocentric PSC 1.0, egocentric PSC 1.0, n.s., Fig. 4). The interaction between the right EBA and right MT was significant (P < 0.01, repeated measures ANOVA). Right LO also did not distinguish between the two conditions (allocentric PSC 1.7, egocentric PSC 1.7, n.s., Fig. 4), and the interaction between the right EBA and right LO was significant (P < 0.05, repeated measures ANOVA).
Whole brain random effects analyses were also conducted. None of the previously discussed brain regions passed the threshold of (P < 0.001 uncorrected) in the comparison, allocentric > egocentric. The random effects analysis did reveal two brain regions with a higher response to egocentric than to allocentric pictures: one in the left post-central gyrus (PCG, [−12 −33 66]), near the primary somatosensory representation of the leg/foot (C. Moore, personal communication), and one in the right dorsolateral prefrontal cortex (DPFC, [51 27 27]). In the right DPFC, the response to the egocentric perspective was higher than the response to allocentric perspecitve, which was higher than the response to fixation. In the left PCG, however, the response to egocentric images was equal to the response during passive fixation, and the response to allocentric images was suppressed below the fixation baseline (Fig. 4).
Discussion
Regions of extrastriate cortex were identified with preferential responses to human bodies over familiar objects (EBA), to moving over stationary rings (MT), and to familiar objects over stationary rings (putative LO). As in previous studies, these three regions were in neighbouring but not identical anatomical locations. Furthermore, there were robust functional differences between the three regions. The EBA showed a dramatic response to images of human bodies, relative to all other stimulus conditions. MT showed a high response to both moving rings and stationary pictures of human bodies, and low to both stationary rings and familiar inanimate objects. Finally, the LO response was high for both bodies and objects, and low for both moving and stationary rings. These results confirm the existence in the human right lateral occipito-temporal cortex of at least three anatomically and functionally distinct regions, all of which respond robustly to images of human bodies relative to fixation.
Right and Left EBA
The response of the right EBA to still photographs of body parts was enhanced when body parts were displayed from a perspective inconsistent with looking at one's own body (the allocentric perspective; see also A.W-Y. Chan, M. Peelen and P. Downing, unpublished observations). Importantly, the enhancement was selective: neither nearby brain regions (right MT and LO) nor the left EBA differentiated the two perspectives. The functional profile of the right EBA was significantly different from those of the right MT and LO, and the interaction with the left EBA approached significance. These results suggest that the right EBA does genuinely distinguish between images of body parts based on the perspective of the image, responding preferentially to images only consistent with looking at someone else's body.
The apparent laterality of preference for an allocentric perspective in the current results is consistent with previous research suggesting a right-lateralized preference for the perception of others' actions. For instance, Leube et al. (2003) let subjects watch videos of their own hand actions, into which a small temporal delay was added. The response in the right posterior STS (probably neighbouring but not identical with the EBA) was positively correlated with increasing temporal delay. That is, as the observed motion was less similar to the subject's own actions, and so appeared more like the action of another person, the response of the right posterior STS increased.
Both Leube et al.'s findings and the current results are consistent with previous evidence that the left and right hemispheres are preferentially involved in self- and other-recognition respectively (Turk et al., 2002; Brady et al., 2004). In the current study we did not find a preference for the self-perspective in the left EBA, possibly because the egocentric images were not actually pictures of the subject's own body. Also, while the allocentric perspective on the body parts are never accessible to a person looking at her own body (making the stimuli unambiguous), viewers do sometimes see other bodies from an egocentric perspective.
It is notable that in the present study neither the left nor the right EBA showed a preferential response to body parts presented from an egocentric position, the pattern predicted for a region involved in motor control (and actually observed in a region of somatosensory cortex, described below). By contrast, Astafiev et al. (2004) recently reported a small bilateral increase in activation of the EBA when subjects actually moved their own hand, or imagined moving it, even in the absence of visual feedback. Further research will be necessary to reconcile these results, and to characterize the contribution of the EBA to one's own body image.
MT and LO
Right MT responded robustly to pictures of body parts in both the localizer experiment and the body perspective experiment. The magnitude of response to body parts in the MT was approximately as high as the response to moving rings. It seems unlikely that implied motion alone can account for this effect. First, the stimuli used in the body perspective experiment had relatively little implied motion. Second, Kourtzi and Kanwisher (2000) found that the increases in MT response due to implied motion and to a human body in the stimulus were approximately additive.
Importantly, MT did not differentiate between egocentric and allocentric images. The high response to body parts in MT therefore cannot be simply leaking over from the nearby EBA: the two neighbouring regions' responses to body parts have significantly different functional profiles. Thus, the motion sensitive region in right human extrastriate cortex may show a genuinely high response to images of human body parts which is distinct from the preference for body parts observed in the nearby EBA. These results suggest a functional disanalogy between human MT, as defined here, and macaque MT which does not respond preferentially to images of bodies (Tsao et al., 2003).
Unlike MT and the EBA, the BOLD response in the LO was robust for both bodies and objects, consistent with its proposed role in general object recognition. Like MT, the right LO did not show the allocentric preference which was seen in the body-selective right EBA, confirming the anatomical specificity of the effect.
Post-central gyrus
In our data, the BOLD response of the PCG was suppressed below the fixation baseline specifically when subjects viewed body parts from an allocentric perspective. The current results are consistent with previous behavioural, neuroimaging and lesion studies (e.g. Farne et al., 2000; Ladavas et al., 2000; Kennett et al., 2001; Taylor-Clarke et al., 2002), suggesting that visuo-tactile integration may cause differential excitation of primary sensory cortices in response to visual images of body parts presented from a self-consistent or a self-inconsistent visual perspective. For instance, Farne et al. (2000) tested neglect patients who experienced tactile extinction following lesions to the right parietal cortex. Tactile sensation on the dorsal surface of the left hand was extinguished by a visual stimulus just above the patient's own extended right hand, or above a rubber arm and hand positioned so as to be visually consistent with the patient's perspective (i.e. extending forward, palm down, from near the patient's right shoulder). When the rubber hand was rotated, to make it visually incompatible with the subject's own perspective (the allocentric perspective), visual stimuli above the rotated rubber hand no longer interfered with sensation in the patient's left hand. Farne et al. suggested that a visually perceived hand would excite the sensory cortex, and so could compete with actual tactile stimulation, only if it was visually consistent with the patient's perspective on his own body, and not if the visual perspective was inconsistent. The current results precisely support this suggestion: the response of the primary sensory cortex was suppressed when the subjects saw images from a perspective incompatible with looking at their own bodies.
The region of significant activation observed in the primary sensory cortex for egocentric > allocentric perspective was located near the sensory representation of the foot (C. Moore, personal communication). The stimuli used in this study included hands, arms and feet. One possibility is that the stimulus manipulation was most robust for the feet stimuli, since the range of accessible perspectives on one's own (relatively inflexible) feet is much smaller than the range of viewpoints from which it is possible to view one's own (extremely flexible) hands.
Right DPFC
The right dorsolateral prefrontal cortex response was enhanced when subjects viewed body parts from an egocentric perspective. This region was not anatomically near to regions of frontal cortex typically associated with self-identification or self-processing (e.g. Macrae et al., 2004). However, lesions of lateral prefrontal cortex have been associated with uncontrollable imitation of observed actions (De Renzi et al., 1996). One possibility is therefore that this region of DPFC is involved in inhibition of sensorimotor cortex in response to observed body parts of others. Future investigation of the role of this region will be necessary.
Conclusions
In sum, the right EBA of the visual cortex responded more to images of body parts when the photographs were taken from a perspective inaccessible to the model. The opposite pattern of response – suppressed response to images from the perspective of another person – was observed in the PCG and the DPFC. All of these regions may be involved in the visuo-tactile integration by which the brain identifies seen body parts as belonging to the self, as opposed to another person. The right EBA's preference for body parts that could not belong to the subject is also consistent with a role for the right EBA in the perception of other people per se, perhaps as input to subsequent perception of others' actions and reasoning about other minds.
This work was funded by grant NIHM 66696. Our thanks especially to Nancy Kanwisher, without whom this work would not have been possible, and to Yuhong Jiang, Rebecca Schwarzlose and Nick Knouf.
References
Astafiev SV, Stanley C, Shulman G, Corbetta M (
Brady N, Campbell M, Flaherty M (
De Renzi E, Cavalleri F, Facchini S (
Downing PE, Jiang Y, Shuman M, Kanwisher N (
Farne A, Pavani F, Meneghello F, Ladavas E (
Grill-Spector K, Kushnir T, Edelman S, Itzchak Y, Malach R (
Grill-Spector K, Kushnir T, Hendler T, Edelman S, Itzchak Y, Malach R (
Grill-Spector K, Kourtzi Z, Kanwisher N (
Kennett S, Taylor-Clarke M, Haggard P (
Kourtzi Z, Kanwisher N (
Kourtzi Z, Kanwisher N (
Ladavas E, Farne A, Zeloni G, di Pellgrino G (
Leube DT, Knoblich G, Erb M, Grodd W, Bartels M, Kircher TT (
Macrae CN, Moran JM, Heatherton TF, Banfield, Kelley WM (
Malach R, Reppas JB, Benson RR, Kwong KK, Jiang H, Kennedy WA, Ledden PJ, Brady TJ, Rosen BR, Tootell RB (
Taylor-Clarke M, Kennett S, Haggard P (
Tootell RB, Taylor JB (
Tsao DY, Freiwald WA, Knutsen TA, Mandeville JB, Tootell RB (
Author notes
1Psychology Department, Harvard University, Cambridge, MA, USA and 2Department of Brain and Cognitive Science, MIT, Cambridge, MA, USA