





Abstract
Group foraging allows for individuals to exploit the food discoveries of other group members. If searching for food and searching for exploitation opportunities within a group are mutually exclusive alternatives, the decision to use one or the other is modeled as a producer-scrounger game because the value of each alternative is frequency dependent. Stochastic producer-scrounger models generally assume that producer provides a more variable and uncertain reward than does the scrounger and hence is a riskier foraging alternative. Socially foraging animals that are attempting to reduce their risk of starvation should therefore alter their use of producer and scrounger alternatives in response to changes in energy budget. We observed flocks of nutmeg mannikins (L. punctulata) foraging in an indoor aviary to determine whether their use of producer and scrounger alternatives were risk sensitive. Analyses of the foraging rewards of three flocks of seven birds confirm that producer is a riskier foraging strategy than is scrounger, although the difference in risk is rather small. We then submitted two other flocks to two different energy budgets and observed the foraging decision of four focal birds in each flock. All but one bird increased their relative use of the riskier producer strategy in the low food reserve treatment, but the overall use of producer did not differ significantly between treatments, providing evidence for a small but consistent effect.
Foraging groups provide animals with the opportunity to exploit food discovered or otherwise made available by other group members. This general form of exploitation or joining has been documented in birds, fish, spiders, and mammals (see Barnard, 1984; Giraldeau and Beauchamp, 1999 and references therein). Social foragers may be faced with a decision if they can alternate between searching for food or for joining opportunities, but not do both simultaneously (incompatible). There is evidence in some ground-feeding birds that the two alternatives are incompatible (Coolen et al., 2001; Koops and Giraldeau, 1996), although it is not universal (see Smith et al. 2002). This incompatibility and its degree may depend on the visual, cognitive, or other constraints of the animals that are specific to each foraging system (Fernandez-Juricic et al., 2004). When the two foraging alternatives are incompatible, the best allocation to each strategy can be obtained by using a producer-scrounger (PS) game analysis (Barnard and Sibly, 1981; Giraldeau and Beauchamp, 1999; Giraldeau and Caraco, 2000). The payoff structure of the PS game is such that either strategy pays more than the other when rare. Thus, when scroungers are rare, they should increase in frequency because scrounger pays more than producer. This is because the scroungers can exploit the discoveries of many searching individuals with very little competition. When producers are rare, they will increase because they do better than the numerous scroungers who must then compete among themselves for the few available discoveries. The result is that the payoff curves of producer and scrounger strategists cross at an intermediate frequency of scrounger: the stable equilibrium frequency (SEF; Mottley and Giraldeau, 2000). Deterministic rate-maximizing PS games hypothesize that animals attempt to maximize their expected food intake and predict the SEF of scrounger within groups. Some empirical studies provide qualitative support for their predictions (for reviews, see Giraldeau and Beauchamp 1999; Giraldeau and Caraco, 2000).
For small animals that have a high metabolic rate, however, the probability of surviving some prolonged nonforaging period, such as night for diurnal animals, may be more important than maximizing the expected intake while foraging (Caraco, 1980). The energy budget rule (Stephens, 1981) predicts that animals should prefer a less variable reward option (risk-averse) when their expected energy budget exceeds the requirements to survive the nonforaging period and, conversely, prefer a variable reward option (risk-prone) when expecting to fall below this requirement. There is support for the energy budget rule in small fishes, birds, and mammals (Bautista et al., 2001; for a review, see Kacelnik and Bateson, 1996; Roche et al. 1998), in which individual subjects expecting an energy deficit increased their preference for a variable (risk-prone) over a constant (risk-averse) foraging option with equal means. Likewise, if producer and scrounger strategies have different variability and predictability of rewards associated with each, then the SEF of scrounger in a group may also depend on the energy budget of the foragers (Barta and Giraldeau, 2000; Caraco and Giraldeau, 1991; Giraldeau and Caraco, 2000).
Stochastic PS models assume that producer generally yields a more variable intake than scrounger and hence offers a risk-prone alternative to group foragers (Barta and Giraldeau, 2000; Caraco and Giraldeau, 1991; Giraldeau and Caraco, 2000). Producer is often the more variable option because it yields either no food or a large fraction of a food discovery: the amount eaten before the arrival of scroungers plus the amount shared with scroungers. Scrounger is usually the less variable option because the animal has a more certain provision of scrounging opportunities, each generating a small fraction of the food discovery. Risk-sensitive PS models predict that the group will reach a SEF of scrounger as a consequence of individuals each attempting to minimize their probability of energy shortfall, so that the use of the foraging strategies will vary with the individuals' energy budget.
To date, only one published study experimentally tested a risk-sensitive PS model. Koops and Giraldeau (1996) tested Caraco and Giraldeau's (1991) risk-sensitive PS model by manipulating the energy budget of starlings (Sturnus vulgaris) foraging in a group. Although the starlings responded to altered patch encounter rates as predicted by the model, they did not respond as predicted to different levels of food deprivation. Moreover, no study to date has been able to ascertain the validity of the assumption of the risk-sensitive PS model concerning the risk-prone and risk-aversive qualities of producer and scrounger, respectively. The difficulty in measuring the profitability and the risk associated to the two strategies resides in the fact that although one can count the number of food patches discovered or joined by an individual, the effort invested in each strategy is unknown (Giraldeau and Caraco, 2000).
Coolen et al. (2001) recently developed a behavioral means of continuously monitoring the strategy used by nutmeg mannikins (L. punctulata) independently of whether they actually find feeding opportunities or not. When these birds forage in groups for seeds dispersed on a feeding table, the position of the head while hopping provides an indication of whether the bird is playing producer or scrounger. Hopping with the head down is statistically correlated with patch finding (producer); hopping with the head up, with patch joining (scrounger). Individuals were found to adjust their investment in alternative strategies in response to changes in foraging conditions. When the foraging conditions offered few rich patches that called for a higher SEF of scrounger (Giraldeau et al., 1990), the birds settled on a higher rate of hopping with the head up. In addition, Coolen and Giraldeau (2003) show that increasing the distance between food and cover increases the use of head up in stationary birds, but not the frequency of hopping with the head up. They conclude that the latter is dedicated to the detection of scrounging opportunities.
In the current study, we reproduce the experimental setup of Coolen et al. (2001) as closely as possible and use the birds' hopping with the head up and with the head down to measure their investment in producer and scrounger strategies, respectively. We use these behaviors to investigate the payoff structure and risk associated with both strategies. We then test for risk sensitivity in the birds' use of producer and scrounger strategies when they are placed in conditions of low and high energy budgets by food depriving them.
EXPERIMENT 1: PAYOFF OF PRODUCER AND SCROUNGER STRATEGIES
Methods
Study subject
Nutmeg mannikins are small 13.7 ± 0.2g (±SE, n = 8) granivorous birds originally from southeast Asia. In the wild, they feed mostly by hopping on the ground or climbing tall grasses and twigs in search of the seeds of grasses and weeds (Immelmann, 1965). The sexes are monomorphic, and the birds are social throughout the year.
We formed three flocks of seven randomly selected birds from a colony of 97 wild-caught adult birds, and observed them as they foraged in an indoor aviary (195 × 305 cm, 240 cm high) kept at 22°C–24°C and a 12-h light/12-h dark photoperiod. Each bird was identified with a unique combination of two color leg bands, and before experiments, each was given a different colored acrylic paint mark on the head and tail. Water was available ad libitum at all times, as was a mixture of millet seeds outside experimental periods.
Foraging apparatus
The birds foraged on a grid consisting of two plywood boards joined to form a 2.0 × 1.2-m surface. The grid contained 198 wells in total (mean diameter and depth (±SE, n = 10) of 1.32 ± 0.02 cm and 0.83 ± 0.01 cm, respectively) spaced on average at 10.16 ± 0.05-cm intervals. The grid rested 92 cm off the aviary floor to allow a seated observer to film the birds by using a handheld 8-mm color camcorder. Outside of experimental periods, a sheet of opaque paper covered the grid.
Training
After a 7–10-day habituation period in the aviary, the birds were trained to search for white millet seeds placed in the grid's wells. Because these birds use their crop to store seeds, they had to be food-deprived overnight (12 h) plus a short 3-h period from lights on. The experimenter (G.M.W.) initially placed a small handful of seeds around the center of the grid and an increasing proportion of the seeds directly in the wells. Once the birds were searching in the wells, they were exposed to a dispersed food distribution (three seeds placed in each of 60 randomly pre-selected wells) and then to a clumped food distribution (10 seeds placed in each of 20 randomly preselected wells).
A trial typically started once the experimenter left the aviary and the birds flew down from their perches onto the grid. Trials ended after 3 min of foraging or 1 min after all the birds flew back to their perches, after which the remaining seeds and husks were removed. Training consisted of six trials per day at 30-min intervals. Under the dispersed food distribution, the SEF of scrounger is expected to be low, whereas it should be higher for clumped distributions (Coolen et al., 2001; Giraldeau et al., 1990; Livoreil and Giraldeau, 1997). After 3 days of training in the dispersed food distribution, all birds could find at least five patches per trial in three consecutive trials. After 3 days of training in the clumped food distribution, all birds had joined at least three patches per trial in three consecutive trials, and training was considered complete.
Testing
Testing started the day after complete training and was carried out on consecutive days. The flocks were food deprived and given six trials per day on the clumped distribution as for training. A different focal bird was randomly selected and videotaped in each trial. The testing period was interrupted once for each flock owing to the birds not flying down on the grids to forage or to technical difficulties. Test days that immediately followed an interruption were discarded to reduce possible effects of the interruption. Testing for flocks 1 and 2 was interrupted during two consecutive days, after which the subsequent 2 days of testing were discarded. Tests for flock 3 were interrupted 1 day, after which the following day of testing was discarded. Because tests that yielded less than 45 s of observation were discarded, flocks were observed for unequal number of days depending on their performance. Flocks 1, 2, and 3 were tested for 15, 22, and 19 days, respectively, such that each bird was observed at least six times, on average 8.6 ± 0.3 times (±SE, n = 21).
Behavior patterns
Eight behavior patterns were noted from the video records by using the Observer 3.0 event recorder software (±0.1 s): (1) finding a patch, (2) joining a patch, (3) eating a seed from a patch that was found, (4) eating a seed from a patch another bird found, (5) hopping with the head up, (6) hopping with the head down, (7); stationary with the head up, and (8) stationary with the head down. No aggression was observed among the birds while foraging. A bird's head position was based on a projected line going from its eye through its nares. The head was up when this line pointed to the horizon or higher, and down otherwise (Coolen et al., 2001). A series of hops was treated as continuous when each hop was separated by less than 1.0 s. A bird remaining in one place for more than 1.0 s was recorded as stationary. Hops were categorized a posteriori into pursuit and search hops (sensu Coolen et al., 2001). The two hops immediately before a patch discovery were defined as producer pursuit hops irrespective of head orientation. A continuous series of hops in the same head position immediately before a patch-joining event was defined as scrounger pursuit hops irrespective of head orientation. The remaining hops were categorized as producer search hops if the bird had the head down, and scrounger search hops if the bird had the head up. Hopping rate was calculated by dividing the frequency of hops by their durations. Patch encounter rate (patches per second) and seed intake rate (seeds per second) were calculated for producer and scrounger separately by dividing the number of patches and seeds respectively, by the time invested in each strategy: the sum of search, pursuit, and feeding times recorded for each strategy. We calculated the proportional use of producer and as the proportion of time which the bird had its head down and up while searching, respectively.
Analysis
Analysis was limited to the first 60 s of each trial in order to reduce the effect of patch depletion. From each trial in which a bird was observed, we extracted all available 10-s sequences of investments for producer and scrounger strategies and randomly selected one of each for analysis in order to avoid pseudo-replication. For each selected sequence, we calculated the intake rate for producer or scrounger by dividing the number of seeds from found or joined patches, respectively, by the time invested in the sequence (10 s). We then used these estimates to obtain the mean and coefficient of variation (CV) of intake rate of producer and scrounger for each bird. We compared the mean and CV of intake rates between strategies by using repeated-measures ANOVAs with each bird as an independent data point and blocking by flock. When homoscedasticity could not be achieved among flocks, we analyzed them separately by using paired t tests and combined the probability tests if they were in the same direction (Sokal and Rohlf, 1981). In addition, we used the cumulative frequency distribution of these intake rates to estimate a bird's probability of incurring an energy shortfall as a function of energy budget for producer and scrounger strategies. For each strategy, we averaged the frequency distribution of intake rates of all birds and used the log-likelihood ratio (Zar, 1996) to test for differences in the distribution of intake rates between strategies. Statistical tests were run on SPSS 10.0 and were two-tailed.
Results
General results
A total of 182 trials yielded 335.6 min of foraging for 21 subjects. Only 14 trials (8%) included disruptions, which together lasted 5.8 min, leaving a total of 329.8 min of observation time. The birds flew down on the grid and foraged on average (±SE, n = 21) 110 ± 14 s per trial. They spent 52.9 ± 6.6% of their time feeding in patches, 39.3 ± 6.6% hopping, and 7.8 ± 5.1% stationary. During the average trial, a bird obtained 9.6 ± 5.1 seeds from 1.9 ± 1.1 discovered patches, and 7.7 ± 4.4 seeds from 3.1 ± 1.9 joined patches.
Behavioral indicators of strategy use
The proportion of time a bird had its head up while searching was a strong predictor of the proportion of patches that a bird joined (arcsine-square root transformation) among the patches from which it fed (partial correlation, controlling for flock: r = .71, df = 18, p < .001) (Figure 1). In addition a two-way repeated-measures ANOVA testing for strategy (producer/scrounger) and hop type (search/pursuit) with flocks as a between subject factor, showed that scrounger hops occurred at a higher rate than producer hops (F1,18 = 8.32, p = .01). Birds also hopped at a higher rate during pursuit than during search (F1,18 = 8.02, p = .01), more so for scrounger, but the interaction between the two factors was marginally nonsignificant (F1,18 = 4.33, p = .05).
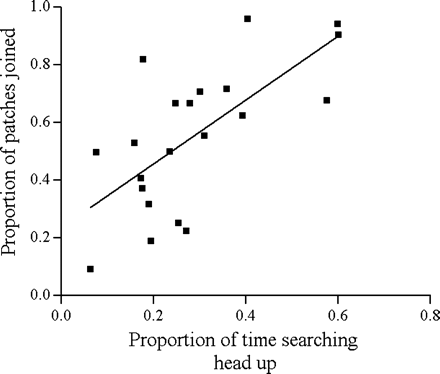
Correlation between the proportion of patches joined by a bird and the proportion of time it had the head up while searching. Each point is the average of six to 10 trials for one bird (n = 21).
Payoffs of producer and scrounger strategies
One bird was excluded from the analysis of payoffs, because it played scrounger too rarely to provide sufficient data for a comparison of strategies. Data from the remaining 20 birds showed that birds had a significantly lower mean (±SE, n = 20) patch encounter rate while playing producer (0.075 ± 0.003 patches/s) than scrounger (0.162 ± 0.007 patches/s) for an equal time investment (repeated-measures ANOVA testing for strategy and flock effects: F1,18 = 153.28, p < .001). On the other hand, birds obtained significantly more seeds from finding a patch (producer, 5.0 ± 0.2 seeds) than joining one (scrounger, 2.6 ± 0.1 seeds; repeated-measures ANOVA testing for strategy and flock effects: F1,18 = 198.85, p < .001). The resulting seed intake rate for producer (0.21 ± 0.02 seeds/s) was lower than for scrounger (0.25 ± 0.01 seeds/s) but not significantly (repeated-measures ANOVA testing for strategy and flock effects: F1,18 = 2.46, p = .14). The variability of intake rates as measured by the mean coefficient of variation (CV) was significantly higher for birds while producing (1.00 ± 0.02) than scrounging (0.55 ± 0.01; combined probability of paired t tests for three flocks:
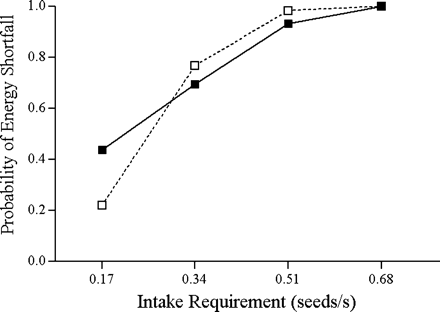
Probability of incurring an energy shortfall relative to the required intake rate for producer (filled squares) and scrounger (open squares) strategies (n = 20).
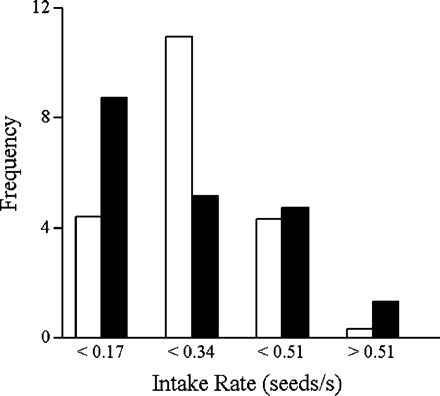
Frequency distribution of intake rates for producer (filled bars) and scrounger (open bars) strategies (n = 20).
EXPERIMENT 2: TESTING FOR A RISK-SENSITIVE DECISION
Methods
Estimating daily food requirement
We estimated the daily food requirement (grams of seeds) of eight L. punctulata randomly chosen from the same pool of 97 birds as used in experiment one. The birds were kept singly in cages (30 × 30 cm, 34 cm high) during 14 days of measurement. The birds maintained visual and auditory contact with others but fed from separate feeders hanging outside their cages. Each feeder contained 10.00 g of white millet seeds, and was changed daily 2 h after lights on. The used feeder was cleared of its empty husks and weighed. The birds were also weighed daily at the same time the feeders were changed.
Manipulation of energy budget
Two flocks of seven individuals were randomly selected from the 68 birds of the colony that had not been used in other parts of this study. They were habituated to an aviary for 14 days and trained to forage on a grid as in experiment 1. Flocks were then given four trials per day. To modify their energy budget, they were subjected to the following two controlled feeding schedules. At the beginning of the second half (6 h) of the day preceding a test, flocks were given either 50% (low energy budget) or 100% (high energy budget) of their estimated food intake for that period. They were then food deprived overnight for 12 h plus 2 h after lights on before testing. Testing always ended after 1.5 h, and birds always had 2 h of ad libitum feeding 30 min after the end of testing. Birds were tested on consecutive days for each treatment, and the treatment order was balanced between flocks. Birds were allowed 7 days of ad libitum feeding and were subject to another 3 days of training between the first and second treatments.
Analysis
We selected four birds randomly in each flock as focal birds, and recorded their behavior during the first 60 s of observation in each trial. We obtained data for each bird by averaging the measures of all trials for each treatment. Repeated-measures ANOVAs tested for effects of energy budget treatment on the birds' foraging behavior, blocking by flock. Proportions were arcsine-square-root transformed to achieve normality and homoscedasticity (Zar, 1996). When no transformation could achieve homoscedasticity between flocks, we analyzed them independently and combined the probability tests if they were in the same direction (Sokal and Rohlf, 1981). Again, all tests were two-tailed.
Results
General
The birds' average (±SE, n = 8) food requirement was 3.7 ± 0.3g of millet per day and was not correlated to their weight (13.7 ± 0.2g). Results of flock foraging are based on 163 trials (approximately 10 trials per bird per treatment), yielding 310.9 min of foraging for all eight birds. Flocks foraged as in the previous experiment except that disruptions occurred more frequently than in experiment 1. A total of 71 trials (44%) included disruptions that lasted a total of 30.4 min. Removing the disruptions left 280.5 min of observation time. The birds flew down on the grid and foraged for 114 ± 3 s per trial. They spent 49.9 ± 2.4% of their time feeding in patches, 37.4 ± 2.7% hopping, and 12.7 ± 2.3% stationary.
Effects of energy budget
Birds hopped at a significantly higher rate in the low energy budget treatment (20 ± 1 hops/s) than in the high energy budget treatment (18 ± 1 hops/s; combined probability of paired t tests for both flocks:
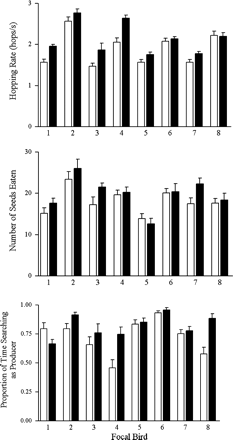
Mean + SE (n = 8–11) hopping rate (a), number of seeds eaten in a trial (b), and proportion of time searching as scrounger (c) in the high (blank) and low (filled) energy budget treatments.
DISCUSSION
The present study provides evidence for risk sensitivity in the PS game in socially foraging nutmeg mannikins. More precisely, results from the first experiment show that producer constitutes a riskier foraging alternative than does scrounger as generally assumed by stochastic PS models. The second experiment reveals that the birds increase their relative use of the producer strategy in response to a decrease in energy budget. We discuss the implications of the two experiments in turn.
Our conclusions rest on the assumption that a bird's head position indicates its foraging strategy. This assumption is based on results provided by Coolen et al. (2001) using the same species of birds foraging in the same experimental apparatus. Moreover, our current results support those of Coolen et al. (2001) because we observed a strong correlation between the proportion of patches that a bird joined and the proportion of time that it had its head up while hopping. In addition, we found that our birds hopped at a higher rate when they had the head up than when they had the head down. The slower hopping with the head down may reflect that seeds are more difficult to detect than a feeding conspecific. It follows that nutmeg mannikins may need to slow down their hopping rate when playing producer and even more so when prey crypticity increases (see Gendron and Staddon, 1983; Pyke, 1981; Speakman, 1986). Thus, our results suggest that an individual's hopping rate may ultimately be used as another indicator of strategy use by nutmeg mannikins. Our estimates of intake rate and risk may not be accurate if the birds could also find food patches or feeding conspecific by using other behaviors. Conceivably, an individual may have been able to detect a feeding conspecific by looking up while remaining in one place. However, we feel this is unlikely given that Coolen and Giraldeau (2003) found a correlation between stationary behaviors with the head up and simulated predation threat but not joining frequency. We are therefore confident that our measure of head position while hopping was an effective indicator of foraging strategy use.
The analyses of producer and scrounger intake rates in experiment 1 show that producer is a riskier foraging strategy than is scrounger in nutmeg mannikins. When playing producer, the birds have a lower patch encounter rate than when playing scrounger, so it follows that they run a higher chance of not obtaining any food at all by playing producer. Our results allow us to go further and show for the first time, that a bird has a higher variability (CV) of intake rate when playing producer than scrounger in this species. This difference in variability constitutes the basis for risk-sensitive foraging (Caraco, 1980; McNamara and Houston, 1992; Stephens, 1981) and is consistent with the common assumption of stochastic PS games (Barta and Giraldeau, 2000; Caraco and Giraldeau, 1991). A study of socially foraging starlings (Koops and Giraldeau, 1996) previously showed that an individual's intake within a patch is more variable when the patch is discovered than joined. The latter however is not sufficient because it did not take into account the effort required to obtain each feeding opportunity, hence the net intake rate. Although the higher variability in producer intake rate in the present study strongly suggests that it is a riskier foraging strategy than is scrounger, it may not be the case if mean intake rates differ greatly. We did not find a significant difference in the mean payoff obtained from using either of the two strategies as predicted by rate-maximizing games for foragers at the SEF (Barnard and Sibly, 1981; Vickery et al., 1991). The observed power of the test (1-β = 0.32), however, does not totally exclude this possibility. A small difference in mean intake rate however, is expected because producer and scrounger birds are predicted to have equal probabilities of falling bellow their requirement, not equal mean intake rates (Barta and Giraldeau, 2000; Caraco and Giraldeau, 1991). The frequency distribution of intake rates also suggests that producer is riskier than is scrounger, but the two distributions did not differ significantly. Overall, our results indicate a potential for a risk-sensitive PS game in our system. The small difference in risk that we observed may not be biologically significant, especially at high required intake rates, at which the differences between strategies are minimal. This may lead to risk-insensitive decisions (Shafir and Trivaks, 2000) or to effects that may be too small to be detectable (Barta and Giraldeau, 2000). In such cases, deterministic rate-maximizing models would appear sufficient to predict the animal's foraging decision.
Despite the small difference in risk noted in the first experiment, the second experiment reveals a limited but consistent increase of producer use with a decrease in energy budget. We discuss possible causes for the small amplitude of the birds' response to energy budgets and the implications of the result for risk-sensitive PS games as well as for the energy budget rule.
The small amplitude of the effect of energy budget on producer use is not likely owing to an ineffective treatment. Although the birds may have increased their food intake during the ad libitum feeding period that preceded the 6 h of low food availability, they were not likely to compensate fully for the following treatment, as these birds ingest approximately 45% of their daily intake in the first half of a 12-h photoperiod (Trudeau LE, unpublished data). In addition, the birds are also not likely to accumulate large food or fat reserves early in the day because the consequent mass gain can impair locomotion and result in an increased vulnerability to predators (see Burns and Ydenberg, 2002; Metcalfe and Ure, 1995). Our birds responded to the low energy budget treatment by hopping at a higher rate and by eating more, indicating that they were hungrier. Instead, the small amplitude of the change in producer use may be owing to the negative frequency dependence of the payoffs in a PS game (Barnard and Sibly, 1981). As an individual increases its use of one strategy, the other strategy will become relatively more profitable. Thus an individual can only shift its use of foraging strategies slightly without a significant trade-off in expected intake rate, which in all cases is a significant factor in survival probability (Barnard et al., 1985). Nonetheless, even small differences in risk may be biologically significant because an energy deficit can have important consequences for the animals, such as the failure to migrate (Bednekoff and Houston, 1994), decreased reproduction (McNamara et al., 1991; Schmitz, 1992), or starvation (Barkan, 1990; McNamara and Houston, 1986). Because our small birds had been food deprived for 14 h in addition to a reduced intake the previous day in the low energy budget treatment, the perceived risk of starvation was likely considerable. Furthermore, small differences in survival probability on a short time scale become significant over an animal's reproductive lifetime (McNamara and Houston, 1982).
The observed increase in preference for risk or producer use with a decreasing energy budget in our small nutmeg mannikins has implications for a number of theories. It is consistent with the energy budget rule and other tests of the model done on small animals choosing between two prey types (Bautista et al., 2001; Kacelnik and Bateson, 1996 and references therein). Our result therefore adds to other studies suggesting that small homeothermic animals respond to energy budget manipulations in a shortfall minimizing manner. In addition, our results indicate that the prediction of the energy budget rule with solitary foragers in mind extends to the choice of producer and scrounger strategies in feeding groups of nutmeg mannikins, as hypothesized by risk-sensitive PS games (Barta and Giraldeau, 2000; Caraco and Giraldeau, 1991). The stochastic PS model (Caraco and Giraldeau, 1991) predicts that a change in the energy budget of group members will affect their use of producer and scrounger strategies depending on group size and the producer's competitive efficiency, the proportion of a food patch that goes to its finder. For the present study, with a group size of seven and a competitive efficiency of 0.59, the model predicts a small decrease of producer use with a decreasing energy budget. Our birds' response was the reverse, but we cannot reject the model because certain assumptions of the model such as sequential patch discoveries and long search times relative to patch time were not met. Nevertheless, it may be that the former model is too simplistic. A dynamic stochastic PS model (Barta and Giraldeau, 2000) assumes that foragers maximize their survival probability throughout the day, and predicts an increase in producer use with a decreasing energy budget when they are very close to immediate starvation. For diurnal animals, this is more likely to occur in the morning, because they cannot feed during night. Our birds foraged during the morning and their response to increased food deprivation is consistent with the prediction of the model. Nevertheless, an explicit test of both risk-sensitive PS models is required to determine which theory is more applicable to socially foraging nutmeg mannikins and to ground feeding birds in general.
We thank Zoltan Barta, Thomas Caraco, and two anonymous reviewers for useful comments on a previous version of the manuscript. This research was conducted in partial fulfillment of the degree of masters in Biology of Concordia University and was financially supported by a Natural Sciences and Engineering Research Council of Canada research grant, as well as by a Fonds pour la Formation de Chercheurs et l'Aide à la Recherche du Québec to L.A.G. G.M.W. received financial support through teaching assistantships provided by the Department of Biology of Concordia University as well as from National Environment Research Council Discovery, FQRNT Team, and PAFARC (UQÀM) research grants to L.A.G. All experiments reported here were conducted under the guidelines of the Canadian Council for Animal Care and were approved by the Concordia University Animal Care Committee.
References
Barkan CPL,
Barnard CJ (ed),
Barnard CJ, Brown CAJ, Houston AI, and McNamara JM,
Barnard CJ, and Sibly RM,
Barta Z, and Giraldeau L-A,
Bautista LM, Martin B, Martinez, L. and Mayo C,
Bednekoff PA, and Houston A,
Burns JG, and Ydenberg RC,
Caraco T, and Giraldeau L-A,
Coolen I, and Giraldeau L-A,
Coolen I, Giraldeau L-A, and Lavoie M,
Fernandez-Juricic E, Erichsen JT, and Kacelnik A,
Gendron R, and Staddon J,
Giraldeau L-A, and Beauchamp G,
Giraldeau L-A, and Caraco T,
Giraldeau L-A, Hogan JA, and Clinchy MJ,
Kacelnik A, and Bateson M,
Koops MA, and Giraldeau L-A,
Livoreil B, and Giraldeau L-A,
McNamara JM, and Houston AI,
McNamara JM, and Houston AI,
McNamara JM, Merad S, and Houston A,
Metcalfe NB, and Ure SE,
Mottley K, and Giraldeau L-A,
Roche JP, Dravet SM, Bolyard K, and Rowland W,
Schmitz OJ,
Shafir S, and Trivaks B,
Smith RD, Ruxton GD, and Cresswell W,
Speakman JR,
Vickery WL, Giraldeau L-A, Templeton JJ, Kramer DL, and Chapman CA,
Author notes
aDepartment of Biology, Concordia University
bDépartement des sciences biologiques, Université du Québec à Montréal, CP 8888, succursale Centre-Ville, Montréal, Quebec H3C 3P8, Canada

{kind=link}
{kind=link}
{kind=link}
{kind=link}