
Abstract
Rodent and nonhuman primate studies indicate that developmental programming by reduced perinatal nutrition negatively impacts life course cardio-metabolic health. We have developed a baboon model in which we feed control mothers (CON) ad libitum while nutrient restricted mothers are fed 70% of ad libitum global feed in pregnancy and lactation. Offspring of nutrient restricted mothers are intrauterine growth restricted (IUGR) at term. By 3.5 years IUGR baboons showed signs of insulin resistance, indicating a pre-diabetic phenotype, in contrast to healthy CON offspring. We hypothesized that a novel breath analysis approach would provide markers of the altered cardio-metabolic state in a non-invasive manner. Here we assess whether exhaled breath volatile organic compounds (VOCs) collected from this unique cohort of juvenile baboons with documented cardio-metabolic dysfunction resulting from in utero programming can be detected from their breath signatures. Breath was collected from male and female CON and IUGR baboons at 4.8 ± 0.2 years (human equivalent ∼13 years). Breath VOCs were quantified using a two-dimensional gas chromatography mass spectrometer. Two-way ANOVA, on 76 biologically relevant VOCs identified 27 VOCs (p < 0.05) with altered abundances between groups (sex, birthweight, and sex x birthweight). The 27 VOCs included 2-pentanone, 2-octanone, 2,2,7,7-tetramethyloctane and 3-methyl-1-heptene, which have not previously been associated with cardio-metabolic disease. Unsupervised principal component analysis of these VOCs could discriminate the four clusters defining males, females, CON and IUGR. This study, which is the first to assess quantifiable breath signatures associated with cardio-metabolic programing for any model of IUGR, demonstrates the translational value of this unique model to identify metabolites of programmed cardio-metabolic dysfunction in breath signatures. Future studies are required to validate the translatability of these findings to humans.
Export citation and abstract BibTeX RIS

Original content from this work may be used under the terms of the Creative Commons Attribution 3.0 licence. Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
Introduction
Breath analysis of volatile organic compounds (VOCs) has potential for providing non-invasive biosamples for assessment of metabolic diseases [1]. From single-celled microbes [2] to complex organisms including mammals [3], many organisms produce VOCs. Quantifying VOCs yields signatures, which may indicate the metabolic state of the organism. VOCs, which are present in all bodily fluids [4], can be products of cellular metabolism [3, 5]. An intriguing aspect of VOC research is the potential to identify disease status by comparing samples from disease and non-disease states. This approach has been applied to a number of human diseases including lung [6–8], digestive [9–12], and metabolic diseases [13–15].
Human diseases have complex origins including genetics and the environment and their interactions. Given their anatomical, genetic, and physiological similarities to humans, nonhuman primates (NHP) serve as excellent model organisms for studying human disease. NHPs naturally develop obesity, diabetes, hypertension and cardiovascular disease similar to humans, and it is possible to tightly control breeding, diet, and environment [16]. Baboons, an Old World NHP species, have been used to address developmental programming, specifically how maternal health and the intrauterine environment impact health and development of offspring in utero with many outcomes persisting postnatally [17, 18]. Intrauterine growth restriction (IUGR), is defined by low birthweight [17], and can be caused by multiple challenges during fetal development including moderate maternal caloric restriction (MCR) and insufficient placental nutrient transport [17, 19]. Additionally, IUGR has also been associated with increased risk of disease later in life including cardio-metabolic diseases [20–22].
Recent evidence shows that IUGR impacts fetal development and postnatal health [23–27]. Analysis of baboon fetal heart tissue revealed that MCR altered mitochondrial DNA content, mRNA expression of oxidative phosphorylation mitochondrial genes and oxidative stress as indicated by increased levels of lipid peroxidation [28]. Juvenile IUGR baboons have increased insulin resistance and β-cell responsiveness indicative of emerging diabetes [23]. As young adults, they have sub-optimal right and left ventricular cardiac function [27, 29]. Adult male IUGR rats were more susceptible to hypoxia, evident through decreased myocardial mitochondrial function and contractility of the right ventricle [26]. These studies indicate that IUGR has a major impact on cardio-metabolic function, altering the trajectory of health and accelerating disease development. With this evidence of cardio-metabolic state by early adulthood, we hypothesized that markers indicative of metabolic differences between IUGR and control (CON) baboons would be present and quantifiable in biosamples prior to the development of observable, overt, clinical disease. To establish any changes, we collected breath samples from juvenile male and female IUGR and CON baboons, the closest experimental model for human translation, to determine feasibility of breath analysis and identification of biomarkers of sex and IUGR status.
Methods
Animal model
All animal procedures were approved by the Texas Biomedical Research Institute's Institutional Animal Care and Use Committee and conducted in AAALAC approved facilities. Baboons (Papio hamadryas) were maintained at the Southwest National Primate Research Center in outdoor group housing, which provided full social environment, physical activity and feeding using an individual feeding system described previously in detail [30]. From 0.16 gestation, age- and weight-matched pregnant female baboons were randomly assigned to be fed either ad libitum during pregnancy and lactation or a diet that was globally reduced to 70% of feed eaten by controls at the same stage of gestation [31]. Offspring of these mothers, IUGR and CON, lived in the same group housing and were fed the same diet (12% energy from fat, 18% from protein, and 69% from carbohydrate consisting of 0.29% glucose and 0.32% fructose; Monkey Diet 5038, Purina, St Louis, Missouri) after weaning at 9 months. This study assessed IUGR and CON baboons as juveniles, which included male (n = 3, M-CON and n = 3, M-IUGR) and female (n = 4, F-CON and n = 3, F-IUGR) baboons with an average age of 4.8 ± 0.2 (human equivalent ∼13 years of age).
Sample collection
Animals were fasted overnight prior to breath collections. Anesthesia was induced with ketamine hydrochloride (10 mg kg−1 I.M.) and continued with isoflurane (1%–2%). Breath samples were collected in the same clinical room for all animals with the same scientific and veterinary staff present for each collection. Each sample was collected over 120 s from intubated baboons. Using a breath collection apparatus (Menssana Research, Inc., Fort Lee, NJ), exhaled breath was pulled across sorbent tubes (Anasorb GCB1, SKC, Eighty Four, PA) through a T-line that was inserted into the exhalation line. Samples were collected at a rate of 0.5 l min−1. Air-lines were flushed for a minimum of 120 s between animals. Tubes were sealed and stored at 4 °C until analysis.
GC × GC-TOF MS analysis
Sample tubes were thermally desorbed using a Unity 2 thermal desorber (Markes International Inc., Gold River, CA) onto a Pegasus 4D GC × GC-TOF MS system equipped with an Agilent 6890 gas chromatograph (Agilent Technologies, Santa Clara, CA) in line with a LECO time-of-flight mass spectrometer (LECO Corp, St Joseph, MI). Primary column was a 30 m × 0.25 mm i.d. × 0.25 μm df Rtx-5MS (Restek Corp., Belefonte, PA) and secondary column a 2 m × 0.18 mm i.d. × 0.18 μm df DB-17M (Agilent Technologies, Santa Clara, CA). Carrier gas was helium and runs were performed in splitless mode. The initial temperature was 35 °C for 1 min, followed by ramping to 100 °C at a rate of 5 °C min−1 and ramping to 250 °C at a rate of 8 °C min−1 followed by a 30 s hold. Secondary column and modulator temperature offsets were each set to +20 °C. The thermal modulation periods were set to 5 s , hot pulse time 1 s and cool time between stages 1.5 s. Column flow was set to 2.0 ml min−1 and the total run time was set to 34 min. The transfer line was set to 280 °C and ion source 200 °C. Acquisition rate was set to 200 spectra s−1 with a 120 s delay and mass range of 35–400 amu.
VOCs identification
ChromaToF software with Statistical Compare (LECO Corp., St Joseph, MI) was utilized to process the total ion chromatograms including baseline correction, deconvolution, peak calling, spectral matching base on set configurations and VOCs alignment. Only peaks with a minimum spectral match score of 700 (Forward score) to the NIST 2011 library (National Institute of Standards and Technology, 2011) were included in the analysis. Statistical Compare allows for both retention time alignments in addition to spectral matching across samples. Each VOC was required to be present in all samples within a group to be included in further analyses. Alignments were exported for further processing. Peak abundances were used to create a list of consolidated VOC IDs and sorted based on the presence or absence across animal groups. VOCs were further processed through searching available published literature and metabolite databases (supplementary table 4 is available online at stacks.iop.org/JBR/12/036016/mmedia) to determine biological relevance or know contamination (phthalates, siloxanes, silanes, organohalides, styrenes, etc). For our MS platform we assigned them 'level 2—identified compounds', e.g. fragmentation/MS2 spectra consistent with spectra and fragmentation ions or accurate masses with proposed structures, as described previously [32]. Raw datasets are provided as supplementary tables 1 and 2.
Normalization and statistical analysis
Sorted VOC abundances present for 13 baboon samples were imported into Partek Genomics Suite (Partek, Inc., St. Louis, MO). The dataset was quantile normalized [33] and subjected to a two-way ANOVA. Sex, birthweight, and interactions between sex and birthweight were assessed to compute fold changes and p-values. We report VOCs for both nominal (p < 0.05) and adjusted p-values (FDR correction of 0.05). A Venn diagram was created using R package VennDiagram [33]. All normalized and statistically analyzed data are provided as supplementary table 3.
Results
Identification of VOCs in all groups
Collection of breath VOCs from baboons under isoflurane sedation poses challenges to chromatographic separation of metabolites due to the high concentrations of isoflurane contained in the mixed exhaled breath sample. This challenge was overcome by using the chromatographic resolution conferred by GCxGC to effectively separate breath VOCs from the isoflurane metabolites (figure 1). Our initial alignment for each group produced 1059 VOCs assigned to at least one of the four experimental groups (figure 2). Through a data filtering workflow (figure 3), 148 identified VOCs were present in all groups. 76 VOCs remain after the removal of known contaminants and non-biological VOCs (supplemental table 2). Statistical analysis was performed on these 76 metabolites only, resulting in 27 VOCs with significantly different abundances.
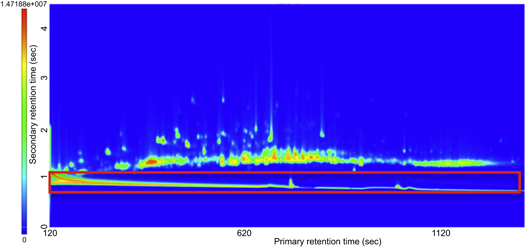
Figure 1. GC × GC-MS total ion chromatogram for breath metabolomics in NHP breath samples collected in a heavy matrix of isoflurane. A contour plot of GC × GC chromatogram as a retention plane of both dimensions and a clear separation of the isoflurane (highlighted red box) across the lower primary dimension (x -axis) from the breath metabolites, which elute in the second dimension (y axis).
Download figure:
Standard image High-resolution image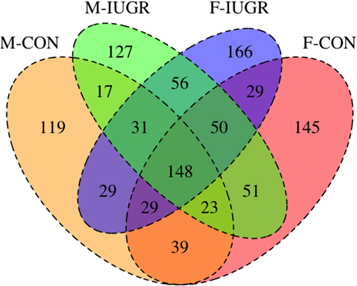
Figure 2. A Venn diagram representing aligned VOCs. A four group Venn diagram showing VOCs present in individual baboon groups and overlapping groups. 148 VOCs were present in all samples.
Download figure:
Standard image High-resolution image
Figure 3. Data filtering workflow. VOCs present in all groups, having potential biological relevance and statistically significant differences (two-way ANOVA, nominal and adjusted p < 0.05) reduced from 1059 to 27 discriminatory VOCs.
Download figure:
Standard image High-resolution imageBreath VOCs indicative of sex differences and birthweight
Two-way ANOVA of VOC abundances revealed putative VOCs, which differed significantly (nominal and adjusted p < 0.05) by sex (table 1) and birthweight (table 2). Listed in table 1 are thirteen VOCs that differed between male and female baboons. Of the VOCs that were significantly different by sex, eight were novel to documented breath analysis including 2-decanone, 2,2,7,7-tetramethyloctane, 1-hydroxy-cyclohexanecarboxylic acid, 1,5-diisopropyl-2,3-dimethyl-cyclohexane, 1,5-diethyl-2,3-dimethyl-cyclohexane, (E,E)-2,4-Heptadiene, 3-ethyl-phenol, and 2,3,6-trimethyl-heptane. Six of the VOCs were lower and seven were higher in female compared to male baboons. Eight of the thirteen compounds are hydrocarbons with two branched alkanes, 2,2,6-trimethyl-octane and 3,7-dimethyldecane having 10-fold and 12-fold lower levels respectively, while 2,3,6-trimethyl-heptane was approximately 40-fold higher in females compared to males. Additionally, three ketones, 2-pentanone, 2-octanone, and 2-decanone where significantly different between males and females.
Table 1. Differentially expressed VOCs by sex.
| Identification | Chemical class | Nominal p-value | Adjusted p-value | Fold change (female versus male) |
|---|---|---|---|---|
| 1,5-diisopropyl-2,3-dimethyl-cyclohexane | Cyclic alkane | 1.07E-04 | 5.51E-03 | −4.0 |
| 2,5,6-trimethyl-decane | Branched alkane | 1.04E-03 | 1.58E-02 | −3.8 |
| 2,2,6-trimethyl-octane | Branched alkane | 1.80E-02 | 1.47E-01 | −10.1 |
| 1,5-diethyl-2,3-dimethyl-cyclohexane | Cyclic alkane | 1.94E-02 | 1.47E-01 | 4.0 |
| (E,E)-2,4-Heptadiene | Alkene | 3.50E-02 | 4.96E-01 | 1.8 |
| 3-ethyl-phenol | Cyclic alcohol | 4.24E-02 | 2.63E-01 | 2.2 |
| 2,3,6-trimethyl-heptane | Branched alkane | 4.50E-02 | 2.63E-01 | 39.4 |
| 2,2,7,7-tetramethyloctane | Branched alkane | 2.11E-03 | 6.62E-01 | −4.9 |
| 2-pentanone | Ketone | 3.79E-04 | 2.67E-02 | 6.7 |
| 1-hydroxy-cyclohexanecarboxylic acid | Carboxylic acid | 3.16E-03 | 3.43E-02 | 1.8 |
| 2-decanone | Ketone | 1.45E-04 | 5.51E-03 | 1.1 |
| 2-octanone | Ketone | 1.72E-02 | 9.59E03 | −8.6 |
| 3,7-dimethyl-decane | Branched alkane | 7.21E-04 | 1.37E-02 | −12.0 |
Table 2. Differentially expressed VOCs by IUGR.
| Identification | Chemical class | Nominal p-value | Adjusted p-value | Fold change (IUGR versus CON) |
|---|---|---|---|---|
| 1,3-dimethyl-benzene | Cyclic alkene | 4.63E-03 | 8.80E-02 | −4.0 |
| 1,2-diethyl-cyclooctane | Cyclic alkane | 7.14E-03 | 1.06E-01 | 5.6 |
| 1,7-Dimethyl-4-(1-methylethyl)cyclodecane | Cyclic alkane | 1.28E-02 | 3.85E-01 | −2.6 |
| trans-1,3-dimethyl-2-methylene-cyclohexane | Cyclic alkane | 1.38E-02 | 1.06E-01 | −6.6 |
| 2,6-dimethyl-octane | Branched alkane | 1.39E-02 | 1.06E-01 | −2.2 |
| 3-methyl-1-heptene | Branched alkene | 1.61E-02 | 1.11E-01 | 5.9 |
| camphene | Monoterpene | 2.74E-02 | 1.74E-01 | 1.9 |
| 2-pentanone | Ketone | 2.43E-03 | 3.01E-02 | −3.6 |
| 1-hydroxy-cyclohexanecarboxylic acid | Carboxylic acid | 3.94E-03 | 8.80E-02 | 1.7 |
| 2-octanone | Ketone | 1.04E-02 | 8.80E-02 | 15.3 |
| 6-ethyl-2-methyl-octane | Branched alkane | 1.37E-02 | 1.06E-01 | 2.4 |
| 2,2,7,7-tetramethyloctane | Branched alkane | 3.96E-04 | 4.84E-01 | 12.1 |
Listed in table 2 are twelve VOCs that differed between IUGR and CON baboons. Five novel hydrocarbons were detected with three, 2,2,7,7-tetramethyloctane, 1,2-diethyl-cyclooctane and 3-methyl-1-heptene, having greater levels in IUGR baboons than CON and two others, trans-1,3-dimethyl-2-methylene-cyclohexane and 1,7-dimethyl-4-(1-methylethyl)cyclodecane, having lower levels in IUGR compared to CON. Three additional hydrocarbons, 6-ethyl-2-methyl-octane, 1,3-dimethyl-benzene and 2,6-dimethyl-octane had lower levels in IUGR compared to CON baboons and both 2-octanone, and camphene had higher levels in IUGR compared to CON baboons.
In addition to assessing VOCs profile differences by sex and birthweight, we analyzed sex by birthweight and found 13 VOCs that were significantly altered (nominal and adjusted p <0.05) (table 3). Of the 13 compounds, three were ketones including 2-octanone, 2-pentanone and 3-decanone, one was an aldehyde 2-ethyl-hexanal, one was an alcohol 2-methylbutan-2-ol, and the remaining seven were hydrocarbons. We observed increased levels of 2-octanone for M-IUGR baboons compared to the 3 other groups and increased level of 2-pentanone in F-CON compared to the other 3 groups (supplementary table 3). Additionally, 2-ethyl-hexanal was observed to lower in F-IUGR compared to F-CON and M-IUGR (supplementary table 3). Combining these VOCs with tables 1 and 2 produced a list of 27 discriminately identified VOCs.
Table 3. Differentially expressed VOCs for the discrimination of sex x birthweight.
| Identification | Chemical class | Nominal p-value (sex) | Adjusted p-value (sex) | Nominal p-value (birthweight) | Adjusted p-value (birthweight) | Nominal p-value (sex x birthweight) | Adjusted p-value (sex x birthweight) |
|---|---|---|---|---|---|---|---|
| 2,2,7,7-tetramethyloctane | Branched Alkane | 2.11E-03 | 6.62E-01 | 3.96E-04 | 4.84E-01 | 9.15E-04 | 8.68E-01 |
| 2-pentanone | Ketone | 3.79E-04 | 2.67E-02 | 2.43E-03 | 3.01E-02 | 2.58E-03 | 6.95E-02 |
| 1-hydroxy-cyclo-hexanecarboxylic acid | Carboxylic Acid | 3.16E-03 | 3.43E-02 | 3.94E-03 | 8.80E-02 | 7.43E-03 | 1.14E-01 |
| (E)-4-octene | Alkene | 9.88E-01 | 6.49E-01 | 8.82E-01 | 4.31E-01 | 7.76E-03 | 7.00E-01 |
| 4-nonene | Alkene | 5.96E-01 | 9.88E-01 | 5.59E-02 | 9.06E-01 | 7.99E-03 | 1.14E-01 |
| 2-decanone | Ketone | 1.45E-04 | 5.51E-03 | 4.57E-01 | 6.43E-01 | 9.02E-03 | 1.14E-01 |
| 2-octanone | Ketone | 1.72E-02 | 9.59E-03 | 1.04E-02 | 8.80E-02 | 1.17E-02 | 9.81E-02 |
| 2,3,6,7-tetramethyl-octane | Branched alkane | 8.99E-01 | 9.62E-01 | 5.92E-02 | 2.96E-01 | 1.49E-02 | 1.41E-01 |
| 2-ethyl-hexanal | Aldehyde | 1.70E-01 | 4.96E-01 | 6.73E-01 | 7.79E-01 | 2.37E-02 | 2.00E-01 |
| 3,7-dimethyl-decane | Branched alkane | 7.21E-04 | 1.37E-02 | 7.62E-02 | 2.96E-01 | 3.42E-02 | 2.40E-01 |
| 2,6,8-trimethyl-decane | Branched alkane | 9.25E-01 | 9.64E-01 | 7.20E-01 | 7.90E-01 | 3.57E-02 | 2.40E-01 |
| 6-ethyl-2-methyl-octane | Branched alkane | 3.74E-01 | 6.38E-01 | 1.37E-02 | 1.06E-01 | 3.85E-02 | 2.40E-01 |
| 2-methyl-2-butanol | Alcohol | 8.83E-01 | 6.49E-01 | 6.37E-01 | 7.79E-01 | 4.10E-02 | 6.68E-01 |
Principal component analysis (PCA) indicative of sex and birthweight
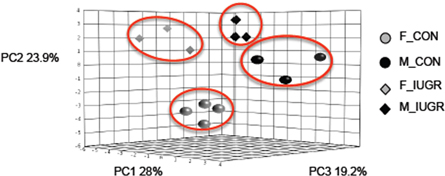
Utilizing an unsupervised PCA of abundances from 27 VOCs listed in tables 1–3 discriminated samples into four distinct clusters (figure 4). This analysis explained 71.1% of the total variation in the data based on the first three principal components (PC), PC1 (28%), PC2 (23.9%) and PC3 (19.2%).
{kind=link}
{kind=link}
{kind=link}
Figure 4. PCA reveals breath signatures that can discriminate sex and birthweight. PCA of 27 discriminately identified VOCs cluster into four specific groups (highlighted red ovals) explaining 71.1% of the total variation in the data, a PC1 (28%), PC2 (23.9%) and PC3 (19.2%).
Download figure:
Standard image High-resolution image{kind=link}
Discussion
Demonstration that breath VOCs can provide an accurate measure of metabolic status non-invasively would significantly improve human health care by providing biomarkers for early detection of pre-clinical conditions in early pathophysiological stages. While VOC identification in human breath is an active area of investigation, human studies present challenges due to uncontrollable environmental and dietary factors that influence breath VOCs. Recent NHP studies have shown feasibility of breath collection while controlling many environmental factors in ways not possible in humans [34]. Our study was designed to show feasibility of identifying robust molecular signatures related to animals' developmental history and sex in NHPs with tightly controlled environmental conditions. Our long-term goal is to use NHP metabolic signatures to inform signatures that may be embedded in noisier human samples.
Our study included baboon offspring with two different developmental histories and both sexes—those from normal pregnancies and those from sub-optimal pregnancies resulting in IUGR. To our knowledge, this is the first study assessing NHP breath VOCs to identify molecular signatures associated with sex and birthweight that may also be indicators of metabolic status. Although some studies have shown that IUGR is associated with childhood and adult morbidity, a recent meta-analysis of current birthweight standards shows that IUGR does not consistently predict offspring health outcomes [35]. These findings highlight the need to develop more sensitive indicators of an individual's health status at birth.
IUGR baboons included in this study had moderately lower birthweights—approximately 12% reduction compared to CON baboons [27, 31]. Approximately one year before breath samples were collected, the IUGR baboons in this cohort showed signs of a pre-diabetic phenotype, evident through increased insulin resistance and β-cell responsiveness [23]. In addition, one year after breath samples were collected, these IUGR baboons showed evidence of impaired cardiac function [27, 29, 36, 37]. Taken together, these observations indicate that the metabolic status of the IUGR baboons differs from CON baboons.
Our results revealed VOCs present in all four groups with known biological relevance and previously reported in breath [4, 6, 14, 34, 38, 39]; however, we also found many VOCs novel breath samples, which have no known relevance to biochemical pathways or physiological function. Although only a subset of the 76 VOCs were significantly different among the four groups, identification of previously reported compounds in humans strengthens the potential translational value of our study in baboons while providing a platform for unbiased analysis to identify novel VOCs associated with an animal's metabolic state.
We also found from our unbiased assessment, twelve VOCs that were different between IUGR and CON, six of which were previously reported in human breath [4, 6, 39–42]. The differences in ketone and branched hydrocarbons suggests that insulin resistance observed one year prior to breath collection in IUGR offspring may be associated with low grade inflammation and oxidative stress. 3-pentanone, a ketone structurally similar to 2-pentanone and 2-octanone, has been implicated in inflammatory conditions [5]. Therefore, the 15-fold increase in 2-octanone in IUGR compared to CON may be due to an inflammatory state in IUGR offspring.
IUGR has been shown to also increase oxidative stress in offspring [43]. Oxidative stress has been linked to altered breath profiles of alkanes [44]. Seven of the twelve metabolites that differed between IUGR and CON are hydrocarbons, many of which are methylated alkanes including 2,2,7,7-tetramethylocatane and 3-methyl-1-heptene; these profiles likely differ between IUGR and CON baboons due to differences in oxidative states.
We identified VOCs camphene (2,2-dimethyl-3-methylene-bicyclo[2.2.1]heptane) that differed between IUGR and CON. Camphene is derived from plants and has been shown to have antioxidant potential, impacting oxidative stress [45] and hyperlipidemia [46]. The source of this compound may be dietary with altered levels as result of different metabolic states between CON and IUGR baboons. Four novel breath hydrocarbons were revealed that differ between CON and IUGR baboons—1,2-diethyl-cyclooctane, 3-methyl-1-heptene, 1,3-dimethyl-2-methylene-cyclohexane and 1,7-dimethyl-4-(1-methylethyl)cyclodecane. Identification of altered exhaled breath VOCs associated with birthweight support feasibility of identifying robust breath signatures from NHPs—future studies in a larger number of NHP are required for validation.
Our study revealed novel sex differences in identified breath VOCs through an unbiased approach. Identifying sex-specific signatures is important given that samples were collected prior to puberty, an age at which current clinical tests may not be able to decipher sex-dependent outcomes. Particular diseases of interest may also have a sex-specific impact. To date, exhaled breath isoprene is the only reported VOC to show sex differences [47]. We did not detect isoprene in all of our samples and therefore did not include isoprene in our statistical analysis, but we did identify thirteen metabolites, seven of which were hydrocarbons that were significantly different by sex (table 1). Studies on fetal baboons reveal sex-specific differences in fetal weight [31], adipocyte differentiation [48], renal mitochondrial gene expression [49], and gene expression in adipose and liver tissues [50]. Sex differences in NHPs are partially mediated by the action of sex hormones, which have the potential to regulate metabolism in a number of different tissues including adipose tissue [51]. In male NHPs testosterone is necessary for normal white adipose tissue (WAT) morphology while in females NHPs, estrogen has a protective effect on WAT during excessive lipid exposure [51]. These differences in normal male and female physiology may lead to altered concentrations of circulating compounds involved in beta oxidation, inflammation and lipid peroxidation impacting end-products detected in exhaled breath. Five of the VOCs detected, 2-pentanone, 2-octanone, 3,7-dimethyl-decane, 2,5,6-trimethyl-decane and 2,2,6-trimethyl-octane, have previously been reported in human breath samples [4, 6, 52, 53]; the fact that they are expressed differentially between males and females is likely due to different physiological states. Ketones and methylated hydrocarbons related to those reported in this study have been implicated in altered metabolic states [5, 6, 39, 44]. Although animals were on the same diet at the time of the breath collection, differences in nutrient utilization and social behaviors have the potential to impact metabolite output. Our results reveal novel VOCs that distinguish these important physiological states; however, the origin of these metabolites as they relate to specific-sex metabolic status needs to be investigated further.
Given that impact of the sex of an organism can influence its metabolic state, we further examined whether each independent factor may interact, i.e. sex and birthweight, revealing additional VOCs of interest, which may extend our ability to identify signatures indicative of metabolic differences among groups. The interaction of sex x birthweight revealed 13 VOCs that are significantly altered (table 3). Nine of the compounds were hydrocarbons, ketones, and a carboxylic acid that were significantly different for sex or birthweight alone. Seven compounds are significantly different for the sex and birthweight interaction that were not significantly altered for sex or birthweight alone including 4-nonene, 2,3,6,7-tetramethyl-octane, 3,7-dimethyl-decane (E)-4-octene, 2-ethyl-hexanal, 2,6,8-trimethyldecane and 2-methyl-2butanol. Interestingly, 2,6,8-trimethyldecane, which was elevated in IUGR baboons, has been shown to be a potential biomarker of type 2 diabetes mellitus (T2DM) [14]. Combined with other alcohols, ketones and hydrocarbon 2,6,8-trimethyldecane was able to discriminate human T2DM from healthy control. It is worth mentioning that the age ranged from 14 to 85 years old which may suggesting 2,6,8-trimethyldecane could be an early indicator of a prediabetic state [14]. Aldehydes have also been implicated as indicators of increased oxidative stress [38]. Although 2-ethyl-hexanal was the only aldehyde with significantly altered levels (table 3), a number of aldehydes in the study were identified including hexanal, nonanal, octanal pentanal, and heptanal (supplementary table 3). These precise signatures provide a means to discriminate according to sex and birthweight and show the capacity of using exhaled breath profiles to discriminate sex and birthweight interactions that differ from sex or birthweight alone. This process would not be possible by assessing individual compounds, as it is an analysis of spectra from many compounds that increase our ability to define a breath signature for each animal group.
For over two decades, breath has been proposed for assessment of cardiac function and therefore a potential source of cardio-metabolic biomarkers [54–58]. As young adults (average age 5.6 ± 0.2 years) our IUGR baboons show cardiac dysfunction, including myocardial remodeling and reduced systolic and diastolic function [27]. IUGR baboons had rEF compared with CON [27] and interestingly, in human studies ketone bodies correlated with heart failure with reduced ejection fractions (rEF) [57]. One human study determined altered ketone bodies in exhaled breath of patients with congestive heart failure (CHF) compared to patients with heart disease but without CHF, and to healthy controls [54]. More recently a study assessing cardio-metabolic biomarkers, showed ketone bodies were elevated in heart failure patients compared to control patients, providing diagnostic potential of breath analysis [59]. A follow up assessment of the prognostic potential of exhaled breath acetone concluded that higher levels were associated with poor prognosis of heart failure patients with rEF [58]. Ketone bodies are normal products of cellular metabolism [60]. They are also implicated in protection of organs and as a fuel source for many organs, including the heart [61]. Different concentrations of ketone bodies may be evidence of altered utilization and it is possible that the altered concentrations are a result of insufficient energy output by a specific organ thereby signaling a need for increased energy production. Although we did not detect differences in previously reported cardio-metabolic markers associated with cardiovascular disease in adults [54, 56–58], we did detect differences in a number VOCs including ketones, alcohols, and hydrocarbons through our unbiased analysis, which may be representative of altered cardiovascular function associated with IUGR and pre-puberty; approximately one year post breath collection, these animals showed impaired cardiac function [27]. Given that these VOCs were collected from juvenile baboons prior to their growth spurt and prior to the cardiac MRI analysis on this cohort [27], we do not know whether these animals had altered rEF and cardiac function and at the time of breath collection. However, since they had shown clear signs of cardio-metabolic function less than one year post breath collection, it is possible that the animals were experiencing signs of altered cardiac function reflected in the altered metabolic breath profiles observed.
A potential limitation of this feasibility study is the sample size. We addressed this limitation by imposing strict alignment software constraints, requiring spectral matches and two retention time matches for all groups of samples, and requiring all samples contain the identified VOCs to reduce the probability of false positives. With this study demonstrating feasibility of NHP breath analysis, future studies will include analysis of longitudinal samples from a greater number of CON and IUGR baboons through their remaining life course, to establish breath signatures of metabolic status and aging.
Conclusion
In summary, this study is the first to identify VOCs in exhaled NHP breath associated with sex and birthweight. Discovery of novel breath VOCs in a NHP that is metabolically very similar to humans shows the utility of this model to identify breath signatures associated with metabolic status. In addition, identification of VOCs in NHP previously identified in humans supports the feasibility of translating findings from NHP breath signatures of metabolic status to informing metabolic status in humans.
Acknowledgments
All authors and co-authors disclose no conflict of interest.
We would like to acknowledge Menssana Research Inc. for providing GC × GC-TOF MS analysis.
This study was supported by the Southwest National Primate Research Center pilot study program (NIH P51 RR013986), donors to the Healthy Babies fund at Texas Biomedical Research Institute, and an Internal Research and Development Grant at Southwest Research Institute (R8223). This investigation used resources supported by the Southwest National Primate Research Center (NIH P51OD011133) and was conducted in facilities constructed with support from NIH ORIP grants (C06 RR14578, C06 RR15456, C06 RR013556, and C06 RR017515).