

Abstract
Empirical studies and habitat suitability modeling project significant shifts in species distributions in response to climate change. Because habitat fragmentation can impede species range shifts, wildlife corridors may have increasing importance in enhancing climate resilience for species persistence. While habitat connectivity has been studied for over four decades, the design of connectivity specifically to facilitate species movement in response to climate change is a relatively new challenge. We conducted a systematic review of 116 relevant papers from 1996–2017. Research focused on assessing the utility of habitat connectivity for climate change adaptation by species (N = 29) and modeling and mapping climate-wise connectivity for planning purposes (N = 55). Others addressed fundamental questions of connectivity related to climate adaptation (N = 31). Based on empirical data and computer simulations examining species range shifts in response to climate change at leading edges of current distributions; it is clear that large protected areas connected through linkages, and stepping stones embedded in a permeable matrix promote population persistence and facilitate range expansion. We identified 13 approaches to modeling climate-wise connectivity based on either focal species or landscape structure. When prioritizing areas for connectivity conservation, approaches include focusing on connecting areas of low climate velocity, refugia, climate analogs, or linking current to future suitable habitats. Riparian corridors should be considered in connectivity plans because of their importance as natural movement corridors, climate gradients, and refugia. Guidance is provided on selecting the best methods for connectivity design depending on the objectives, available data, and landscape context. Future research needs to evaluate the functionality of climate-wise connectivity models for facilitating range shifts and compare connectivity outcomes across modeling approaches.
Export citation and abstract BibTeX RIS

Original content from this work may be used under the terms of the Creative Commons Attribution 3.0 licence.
Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
1. Introduction
Climate change presents a fundamental threat to biodiversity. Widespread biotic responses have already been documented, including severe range contractions, local and global extinctions, shifts in phenology causing disruptions of species interactions, but also adaptations to warmer conditions, shifts in species distributions, and changes in resource use and dispersal capabilities at the leading edges of range margins (Parmesan 2006). In the face of climate change, existing obstacles to species range shifts are becoming increasingly problematic. These include landscape fragmentation from roads, intensive agriculture, residential development (Opdam and Wascher 2004, Kitzes and Merenlender 2014); and, increasingly, the velocity of changing climate, associated direct and indirect ecological effects (Loarie et al 2009), and mismatches with dispersal capabilities (Schloss et al 2012).
Conservation planning has focused on advancing landscape connectivity through identification and protection of linkages in the form of corridors, ecological networks, greenbelts, and other landscape features (Heller and Zavaleta 2009) to facilitate movements of genes, propagules (e.g. pollen and seeds), individuals, and populations, and maintain ecological processes (Hilty et al 2006, Heller and Zavaleta 2009). However, while many models exist to estimate the extent of remaining habitat connectivity and researchers have recognized the need for climate-wise connectivity for some time (Peters and Darling 1985, Hunter et al 1988), only recently has there been progress in the design of climate-wise connectivity—connectivity that specifically facilitates animal and plant movement in response to climate change. This is despite the fact that of a number of conservation strategies for planning and developing climate resilient landscapes (Schmitz et al 2015) this is one of the most commonly cited conservation strategy for climate adaptation (Heller and Zavaleta 2009).
Historically, habitat connectivity aimed to functionally connect suitable habitats such that individual organisms can move within their current range. The species that most often required improvements in habitat connectivity tended to be wide-ranging, or exist as meta-populations (Beier et al 2008). Climate-wise connectivity expands the need for connected landscapes: it aims to also connect current habitat to habitat that will become suitable in the future (Hodgson et al 2011). Furthermore, connectivity to facilitate range shifts will likely be essential for all species that can adapt to the changing climate by shifting their ranges (Heller and Zavaleta 2009, Mawdsley et al 2009, Krosby et al 2010, Hannah 2011). Because range shifts will likely occur over generations, climate-wise connectivity needs to provide sufficient habitat for individuals to live in, and find resources throughout their entire life cycles, not just for daily, dispersal, or migratory movements which can occur through habitat that may only be suitable for movement or feeding (Hannah 2011). Further, as species shift their ranges toward future suitable climate conditions, climate-wise connectivity is directional, following temperature and moisture gradients (Parmesan 2006, Killeen and Solorzano 2008). For these reasons, geophysical features that create a diversity of microclimates are important to focus on as they can buffer the effects of climate change, giving species more opportunities and time to track the changing climate (Hannah et al 2014, Anderson et al 2015).
In this paper, we discuss concepts that underpin climate-wise connectivity rooted in how species adapt to climate change. In particular, the role of habitat connectivity has been assessed for some species and communities and leads to conclusions regarding effective conservation strategies for protecting climate resilience landscapes. We share these strategies along with approaches to mapping and modeling climate-wise connectivity to help design corridors and reserve networks. We conclude by connecting the available modeling methods to improve climate-wise connectivity on the ground and suggest topics for future research.
Three concepts are important to explore for consideration in climate-wise connectivity planning: climate velocity, refugia, and population dynamics at the leading and trailing range edges. The concept of climate velocity is to quantify the rate of climate change over time in relation to spatial gradients of climatic heterogeneity across a landscape (Loarie et al 2009). For temperature, climate velocity expresses the speed at which a population would have to move to keep up with the changing climate on the landscape; velocity is calculated as the ratio of rate of climate change (°C year−1) divided by the spatial gradient (°C km−1) to reveal the speed of change as km year−1. Global studies of animal diversity show greater diversity of narrow range species in areas with low velocity (mainly mountainous regions; Sandel et al 2011), supporting the view that rugged topography could help buffer the biotic impacts of climate change by allowing species to move shorter distances, thereby mitigating changing conditions (Ackerly et al 2010). On the other hand, an analysis of montane environments revealed that shortest distance paths connecting present and future suitable climate may often traverse climatically dissimilar landscapes, presenting potential barriers to movement and offsetting the value of heterogeneous topography (Dobrowski and Parks 2016).
The concept of refugia has first been explored in paleoecology with strong evidence for the importance of Pleistocene refugia—areas of endemism—that provided source populations for future species range expansions when the climate began warming (Keppel et al 2012). With respect to modern day climate change, climate refugia are places of lower climate velocity relative to the surrounding. The importance of refugia to buffer species and communities against deleterious effects of climate change has been recognized as one approach to guide protected area planning for climate change resilience (Keppel et al 2012, Keppel and Wardell-Johnson 2012, Hannah et al 2014, Keppel et al 2015). Different types of refugia have been identified. Some researchers make a distinction between macrorefugia and microrefugia. Whereas the former represents larger areas where the climate remains favorable, the latter are 'spread across inhospitable terrains where the refugiated species survive in restricted spots due to the local occurrence of unusual microclimatic conditions' (Rull 2014). Identifying microrefugia requires very high-resolution climate data often including field measurements (Ashcroft 2010). Microrefugia that can facilitate species range shifts are sometimes referred to as stepping stones, and are integrated into corridor planning theory (Hannah et al 2014). Microclimate refugia are microsites with a lower rate of climate change. They can play a role in promoting long-term persistence by slowing climate velocity that is experienced by species. These refugia are expected to be more common in areas with high topographic diversity (Anderson et al 2016). In-situ refugia are locations that will remain, at least temporarily, suitable for a species under climate change either because the climate will minimally change compared to the surrounding, because the temperatures are cooler than the surrounding, because precipitation patterns buffer increasing temperatures (Ashcroft 2010, Maher et al 2017), or because climate change is within the range of suitable climate for the species in that location (Carroll et al 2017). Ex-situ refugia are sites that are currently unsuitable for a species but will become suitable in the future due to forecasted warming scenarios (Ashcroft 2010). Less frequently identified types of refugia are drought refugia (areas in arid and semi-arid regions characterized by relatively high plant abundance; Rouget et al 2003), evolutionary refugia (areas where certain organisms are able to persist despite long-term climate changes; Klein et al 2009), and hydrologic refugia (areas of high relative water availability; McLaughlin et al 2017). Epps et al (2006) defined genetic refugia as in-situ refugia that are well connected and thereby promote the maintenance of high genetic diversity. It is important to note that refugia of any kind are not entirely immune to climate change over a century, and hence refugia should not imply permanence in the face of climate change (Hannah et al 2014). However, sites where the velocity of change is slower could buy some species more time to adapt (Heller et al 2015), making them important to include in a protected area network for increased climate resilience.
For many species and habitat types, a changing climate is expected to reduce climatic suitability for some locations or populations, and increase it in others. These declining and improving locations are referred to as trailing and leading edges, respectively. Along temperature gradients, when the climate is warming, trailing edges will generally be found at warmer (southern or lower elevation) locations, while leading edges will occur at cooler (northern, coastal, or higher elevation) locations where new populations may have a chance to establish beyond a species' current distribution (Thomas 2010). In heterogeneous landscapes, trailing- and leading-edge populations may occur in less obvious locations. For example, conditions may deteriorate on south-facing slopes, while new populations might establish in valley bottoms due to warming of cold-air pools and enhanced moisture availability (Ackerly 2003, Morin and Lechowicz 2008). Connecting trailing edges to the main range and leading edges to future suitable habitat by improving landscape connectivity may be essential to facilitating migration and the adaptation of native species to climate change. Locations with similar climates across current conditions and future scenarios are referred to as climate analogs (Veloz et al 2012); when modeling structural connectivity corridors can be designed to connect one area to another estimated to be a climate analog in the future.
There are different and creative approaches to incorporating these concepts into connectivity assessment and planning, including several that do not and some that do incorporate projections of future climate. Here, we synthesize empirical studies and simulations that assess habitat connectivity for climate adaptation (3.1), map out the existing methods for modeling climate-wise connectivity (3.2), and provide guidance for adopting these approaches to advance climate resilient landscape-scale conservation (4.0).
2. Literature search methods
We systematically searched the online databases ISI Web of Science and Scopus (articles published until September 20, 2017) with a combination of climate-related and connectivity-related terms to find papers at the intersection of connectivity and climate change (Pullin and Stewart 2006):
(('climat⁎ change' OR 'climate velocity' OR 'velocity of climate' OR climat⁎ OR 'global warming' OR 'global change') AND ((corridor⁎ AND connectivity) OR ('range shift⁎' AND connectivity) OR ('conservation planning' AND connectivity) OR ('land facet⁎') OR (linkage⁎ AND connectivity) OR ('coarse filter approach' AND connectivity) OR (wildlife AND (connectivity OR linkage OR corridor OR 'stepping stone')) OR (riparian AND (connectivity OR linkage OR corridor OR 'stepping stone')) OR ('trailing edge⁎') OR (refugia AND (connectivity OR linkage OR corridor OR 'stepping stone')))).
The search terms were iteratively tested and modified using a list of references (see appendix S-1 in supporting information available at stacks.iop.org/ERL/13/073002/mmedia). When reading the papers included in the review from the database search, we looked for additional relevant studies in the citations and included them in the review.
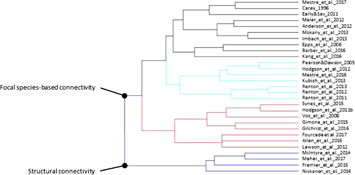
Figure 1. Results of the cluster analysis: studies assessing connectivity for climate change adaptation.
Download figure:
Standard image High-resolution imageWe included all research papers, reviews, and essays that integrate climate and connectivity in their methods and results sections. We filtered the resulting references on three criteria to determine their relevance to our research questions. First, based on the titles and abstracts, ATHK assigned papers to one of three categories: include, exclude based on no inclusion of climate and connectivity in the methods or results sections, or possibly include. Second, titles and abstracts of papers in the 3rd category were reviewed by AMM and DDA. If either investigator considered a paper relevant, it was included in the next step. Third, we performed full text filtering to remove references that may have appeared relevant from the abstract but upon closer inspection did not meet the inclusion criteria. We estimated the comprehensiveness of the database search by calculating the proportion of papers that we obtained through the bibliographies but missed in the database search.
We developed a data extraction spread-sheet to collect metadata from each paper (appendix S-2). Using a subset of the metadata, we conducted a hierarchical cluster analysis (R Core Team 2014) to facilitate grouping the papers based on the metadata collected. We conducted a clustering analysis for heuristic purposes and iterated the analysis to arrive at a useful solution, i.e. one in which the categories correspond to the most important features of the models in relation to our questions about objectives, input, output, corridor end points, and temporal planning horizon. We studied the subgroups to develop climate-wise design approach categories.
3. Results and synthesis
The literature search returned 1333 papers, 103 of which we considered relevant (appendix S-3). We added an additional 12 papers that we obtained through the bibliographies. Of the additional papers, four are reports or book chapters that would not have been encountered in the formal literature search. Without these, the proportion of journal articles that we obtained through the bibliographies but missed in the database search is 6.9%. Of the 115 papers included in the review, 84 were original studies and 31 were review papers or essays. The final cluster analysis split the papers into two main groups: those that assess the utility of habitat connectivity for climate change adaptation by species (figure 1) and those focused on modeling and mapping habitat connectivity to aid in the design of climate-wise connectivity networks (figure 2). In both clusters, the papers were then grouped by whether they focus on focal species connectivity or on structural connectivity. Focal species connectivity is based on information about a single or a set of focal species; structural connectivity describes the physical characteristics of a landscape that are generally understood to facilitate species movement. Within the design cluster, we identified 13 climate-wise design approach categories: six categories capture the studies taking a focal species approach, and seven categories organize the papers taking a structural connectivity approach, although not all clusters correspond with exactly one category (i.e. papers on conservation network planning based on focal species are in two separate clusters). Below, we organize our review assessment based on these two major categories.
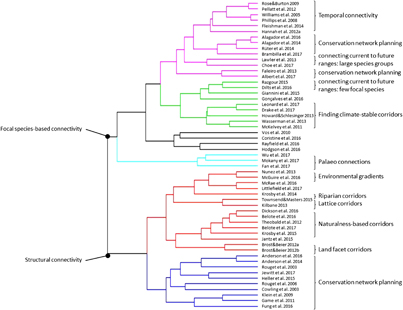
Figure 2. Results of the cluster analysis: studies designing climate-wise connectivity.
Download figure:
Standard image High-resolution imageTable 1. Advantages and disadvantages of strategies to improve climate-wise connectivity.
| Strategy | Advantages | Disadvantages |
|---|---|---|
| Increasing the amount of habitat throughout the landscape | Increases speed of range shifts in fragmented landscapes; benefits most species | |
| Concentrating habitat in few, large areas | Increases species persistence for some species | Slows speed of range shifts |
| Adding corridors between natural or protected areas | Increases speed of range shifts in fragmented landscapes | Trade-off with increasing protected area system; most effective for species with medium dispersal capabilities in moderately fragmented landscapes with lower climate velocity |
| Creating small stepping stones embedded in the matrix | Increases speed of range shifts in fragmented landscapes | Benefits few species |
| Increasing the size of existing protected areas | Increases species persistence; improves temporal connectivity for some species | |
| Improving the permeability of the matrix | Increases speed of range shifts in fragmented landscapes; benefits many species | Unlikely to serve specialist species unless significant habitat restoration is undertaken |
| Maintaining naturally isolated habitats at a density that permits exchange between habitats | Enables dispersal; ensures species persistence; creates genetic refugia |
3.1. Assessing the utility of habitat connectivity for climate change adaptation
For range shifts to occur individual organisms need to physically move to suitable habitat beyond the current distribution (Lawson et al 2012, Fourcade and Öckinger 2017). Several studies assess the effects of landscape permeability determined by the extent and configuration of land use/cover on range shifts at the leading edge of species distributions (table 1). A simulation study examining the effectiveness of different conservation strategies to promote range expansion in a real landscape on eight modeled species with a range of life history strategies found that increasing the amount of habitat throughout the landscape is one of the most effective strategies to facilitate movement (Synes et al 2015). However, concentrating habitat in few large areas reduces the capacity for rapid range shifts for modeled species with varying dispersal capabilities (Hodgson et al 2012). Several simulation and empirical studies have also shown that adding corridors between natural or protected areas can be an effective strategy to facilitate range expansion, but the effectiveness depends on the overall size of the corridor and the elevational gradient (Imbach et al 2013), the degree of landscape fragmentation (Renton et al 2012, Mokany et al 2013, Gimona et al 2015), the amount of available habitat (Collingham and Huntley 2000, Hodgson et al 2011, Synes et al 2015), climate velocity (Renton et al 2012), species' dispersal ability (Meier et al 2012, Kubisch et al 2013, Mokany et al 2013, Gimona et al 2015) and habitat preferences (Hodgson et al 2011).
In more intact landscapes, for plant species with long distance dispersal abilities the quantity of suitable habitat may be more important than the spatial arrangement of suitable habitat for determining successful dispersal (Pearson and Dawson 2005, Renton et al 2012). However, in highly fragmented landscapes where less than 20% of the habitat remains some plant species will not be able to shift their ranges under moderate climate change scenarios, even with strong dispersal capabilities (Renton et al 2013). Conserving or restoring connectivity between suitable habitat to facilitate range expansion is most effective for species with medium dispersal capabilities in moderately fragmented landscapes with lower climate velocity (Pearson and Dawson 2005, Meier et al 2012, Renton et al 2013, Gimona et al 2015).
Corridors that cover large areas and have high altitudinal gradients were found to benefit the greatest number of plant species (Imbach et al 2013), whereas small stepping stones embedded in the matrix are beneficial only to a few species according to studies that examined this for wind-dispersed trees, butterflies, and simulated species with different life histories (Collingham and Huntley 2000, Lawson et al 2012, Synes et al 2015, Gilchrist et al 2016). If habitats are naturally isolated (e.g. vernal pools or microrefugia), they need to exist at sufficient density to keep the distance between them short enough to enable dispersal of species as varied as desert bighorn sheep, water fowl, and arctic plants (Epps et al 2006, McIntyre et al 2014, Niskanen et al 2017). The effectiveness of both, increasing the amount of habitat in the landscape and adding small but critical corridors, often creates a trade-off in conservation strategies because of the commonly high costs of critical corridors (Hodgson et al 2011).
Other strategies of improving landscape connectivity include specifically increasing the size of existing protected areas, adding new protected areas, and enhancing and diversifying the matrix (Donald and Evans 2006, Groves et al 2012, Allen et al 2016, Donaldson et al 2017). Simulation studies designed to compare the effectiveness of these different strategies in facilitating range shifts of modeled species with different life histories differed in their conclusions. While Synes et al (2015) concluded that creation of new habitat adjacent to existing small patches gives the most consistent benefit across species, Hodgson et al (2011) found that adding new habitat randomly provided the most consistent increases in the speed of predicted range expansion. Mokany et al (2013) determined that the best strategy depends on a plant species' dispersal ability and the region, and recommended implementing a mix of landscape configuration strategies. Because different species have different ecological requirements with respect to landscape configuration, landscapes containing large protected areas connected through linkages, or stepping stones embedded in a permeable matrix will promote population persistence and facilitate range expansion at the leading edge for the greatest number of species (Collingham and Huntley 2000, Donald and Evans 2006, Synes et al 2015).
3.2. Climate-wise connectivity design
As this review demonstrates there are many different and creative approaches to climate-wise connectivity planning under exploration. Some variation in approach reflects different conservation objectives (figure 3). Species recovery, a common conservation objective, is the focus of studies designing corridors or protected area networks for focal species (e.g. Dilts et al 2016). Many other approaches aim to facilitate the general movement of species that are projected to respond to climate change with range expansions or range shifts. These designs are based on ensuring connectivity for a large group of species (e.g. Choe et al 2017), a suite of carefully selected focal species (e.g. Albert et al 2017), or focus on structural connectivity (e.g. Littlefield et al 2017). While we present focal species based and structural approaches to connectivity design as distinct, some studies contain elements of both (e.g. Fleishman et al 2014). One effort tries to leverage carbon mitigation to identify lands that will maximize both the amount of carbon sequestered and provide species corridors (Jantz et al 2014). Depending on the objectives, the spatial scale of connectivity planning varies from local to continental, resulting in outputs ranging from corridor designs at the parcel level ready for on-the-ground implementation (e.g. Howard and Schlesinger 2013) to conceptual movement flow patterns across continents (e.g. Lawler et al 2013).

{kind=link}
{kind=link}
Figure 3. Node types (areas that are being connected) and outcomes associated with (a) structural and (b) species-focused climate-wise connectivity modeling approaches. Conservation objectives addressed include connecting protected areas, selecting new sites for protection or planning a conservation network, increasing landscape permeability, conservation of species of concern, and prioritizing regions for connectivity conservation. ˆ Data-intensive models, ⁎ models that require down-scaled climate data, (⁎) models that benefit from down-scaled climate data.
Download figure:
Standard image High-resolution image{kind=link}
3.2.1. Structural connectivity
Planning for structural connectivity aims to accommodate the movement needs of a wide range of species, making it theoretically an efficient planning process. The modeling approaches incorporate different concepts of climate-wise connectivity, and range in design from simple to complex.
3.2.1.1. Riparian corridors
Riparian corridors are commonly used as movement corridors by many species of animals and plants (including terrestrial and aquatic species), support important ecological processes, provide cooler and moister microclimates than the immediate surrounding (especially important in summer or dry seasons), and tend to span climatic gradients as they are oriented along elevational gradients (Beier 2012, Krosby et al 2014). In addition, riparian areas often enjoy popular support for water quality and recreation benefits, and do not require modeling, making them easy to convey for community conservation efforts (Townsend and Masters 2015). In many places, riparian zones often already have some legal protection (Fremier et al 2015), though the legal requirements may not be wide enough to support a full suite of species that could potentially benefit from the corridors.
For these reasons, riparian corridors are often a priority for climate change resiliency. Applying fixed buffers around riparian areas that connect desired termini has been suggested as a simple method to design riparian corridors (Rouget et al 2003, Brost and Beier 2012b). In cases where no specific termini need to be connected, Krosby et al (2014) developed a method for prioritizing riparian areas for climate adaptation based on the temperature gradient the river spans, the width of the riparian area, and the levels of canopy cover, solar insolation, and human modification. The information is combined in an index of climate-corridor quality to estimate the climate adaptation potential for each of the different segments from the headwaters to downstream reaches.
3.2.1.2. Environmental gradients
Environmental gradients influence the distribution of plants and animals (Lawler 2009). Therefore, designing corridors to follow temperature and precipitation gradients may assist individuals tracking suitable climates regardless of the magnitude of climate change (Pearson and Dawson 2005). For topographically diverse areas, climate gradient corridors have been designed that connect presently warm to cool areas in a unidirectional way, prioritizing gradual change in temperatures as well as areas of high naturalness (Nuñez et al 2013, McGuire et al 2016). Mapping connectivity between protected areas to maintain floristic diversity under climate change was done in one case based on land cover permeability and weighted by areas of high species turnover which reflect temperature, precipitation, and soil gradients (Jewitt et al 2017). Another example of mapping environmental gradients included edaphic interfaces, upland-lowland interfaces, and macroclimatic gradients as surrogates for key ecological and evolutionary processes that will ensure resilience to climate change (Rouget et al 2003).
3.2.1.3. Naturalness-based corridors
Naturalness refers to the level of human impact on landscape elements. Naturalness-based corridors for climate change prioritize corridors in areas with the least amount of human development, reasoning that species that are sensitive to human disturbance will be able to more easily traverse such areas (Belote et al 2016). Connectivity is either modeled between protected areas, or on a continuous landscape. Several studies prioritize connectivity between climate analog sites to account for the tendency of species to move towards areas that will provide suitable climates in the future (Nuñez et al 2013, McGuire et al 2016, Littlefield et al 2017). While the focus is on human modification additional information such as slope (Dickson et al 2016), an index of wildness (Belote et al 2016), or ecological integrity, ecosystem representation, and biodiversity priority (Belote et al 2017) may also be included. While corridors designed based on naturalness alone do not minimize climate velocity or connect climate analog sites, the approach can be combined with methods to achieve climate-wise connectivity (Nuñez et al 2013, Littlefield et al 2017).
3.2.1.4. Lattice-work corridors
Ensuring the persistence of a regular crisscrossed array of corridors (lattice-work corridor systems), is an approach to provide comprehensive connectivity without the need of developing complex models. Kilbane (2012) solicited community involvement to modify a systematic trans-continental network of corridors across Australia to match local conditions. Townsend and Masters (2015) recommended involving stakeholders in delineating and protecting connectivity in tropical mountain ecosystems by protecting elevational connectivity along rivers which can enable range shifts to higher elevations, and promoting population viability in perpendicular elevational bands through conservation-friendly land uses. These approaches increase landscape permeability, can incorporate environmental gradients, and are simple in design which makes involvement by the local communities easier, an essential component for successful connectivity conservation implementation (Keeley et al 2018).
3.2.1.5. Land facet corridors
The land facet corridor approach defines landscape units (called land facets, enduring features, geophysical settings, or ecological land units) by topography and soil and aims to maximize in the corridors the continuity and diversity of landscape units found in the neighboring natural areas (Beier and Brost 2010, Brost and Beier 2012b). The rationale behind this concept is that the corridors will support movement by species associated with particular land facets, and even if the suites of species in an area change with a change in climate, biodiversity will remain high because the diversity of landscape units is protected, a concept commonly referred to as 'conserving nature's stage' (Beier et al 2015, Lawler et al 2015). While other approaches to climate-wise connectivity modeling aim to enable species range shifts, land facet corridors are designed to facilitate movement between protected areas of the majority of species that occur there now or will occur there in the future.
3.2.1.6. Conservation network planning
Climate-wise connectivity has been incorporated into systematic conservation planning approaches in different ways. To include a climate-wise connectivity component into a reserve network design that is maximizing representation of different geophysical features as surrogates of biodiversity, microclimatically diverse grid cells in each geophysical category were prioritized (Anderson et al 2014, 2016). To maximize connectivity, priority was given to cells with a high degree of similarity between a focal cell and its neighboring cells with respect to land cover and degree of development based on a resistance kernel analysis (Anderson et al 2014). Anderson et al (2016) further prioritized regional movement pathways that increase in altitude and latitude by integrating landforms into a resistance surface and assigning decreased resistance to upslope movements, and by letting the current in a wall-to-wall Circuitscape approach mostly run in a north-south direction.
Taking advantage of the innate connective properties of streams (see discussion above), other systematic conservation models have approached the climate-wise connectivity goal by prioritizing land units that are located near streams while balancing it with acquisition cost (Klein et al 2009). Temporal connectivity can also be incorporated into systematic conservation planning by prioritizing spatial vicinity between different—cooler and warmer, or drier and moister—habitats, a proxy thought to facilitate species persistence through time. Game et al (2011) accomplished this using the conservation planning software Marxan by requiring a high boundary length modifier which minimizes the difference between topo-edaphic and climate variables in adjacent areas; Heller et al (2015) instead used Marxan to maximize hydro-climate diversity in the reserve network thereby capturing the diversity of climate types in the planning region.
Alternatively, climate-wise corridors can be incorporated into systematic conservation plans by specifying surrogates for key ecological and evolutionary processes such as upland-lowland and macroclimatic gradients as conservation targets (Cowling et al 2003). To capture these gradients in regional-scale corridors, Rouget et al (2006) combined least-cost path analysis with a systematic conservation planning approach that targeted ecosystem representation, suitable wildlife areas, irreplaceable vegetation types, and considered current and future land-use patterns.
While expert-defined corridors are commonly found in habitat connectivity plans (e.g. Penrod 2001), experts were asked to site climate-wise corridors in only two studies (Vos et al 2010, Fung et al 2017). Avoiding the challenge of siting corridors in a systematic conservation modeling framework and including the benefit of familiarity with the socio-economic environment, stakeholders were asked to designate climate-wise pathways based on extensive data generated in the reserve modeling effort, including information on climate velocity (Fung et al 2017), and potential for international connectivity (Vos et al 2010).
3.2.1.7. Carbon stock corridors
While all the connectivity design approaches so far address climate adaptation, carbon stock corridors, aiming to maximize the amount of biomass contained in the corridor, are a climate mitigation strategy (Jantz et al 2014). With funding available for climate mitigation projects in the Reducing Emissions from Deforestation and Forest Degradation (REDD+) framework (Jantz et al 2014) and in Cap and Trade programs (e.g. Balmes 2014), carbon stock corridors add biodiversity conservation benefits by guiding the spatial arrangement of climate mitigation projects.
3.2.2. Connectivity based on focal species
Approaches to connectivity based on focal species all include models of species distributions for at least two points in time with most focusing on current and future distributions, but a few reaching back to the historic record (e.g. Fan et al 2017). Some focus on how present-day connectivity for a species can also contribute to movement for climate adaptation whereas others identify connectivity between current and future ranges across the present landscape or habitat suitability as it changes through time.
3.2.2.1. Finding climate-stable corridors
Several studies characterize how connectivity is changing for focal species under predicted climate scenarios (McKelvey et al 2011, Howard and Schlesinger 2013, Wasserman et al 2013, Giannini et al 2015, Drake et al 2017, Leonard et al 2017). A common conclusion was that suitable habitat area will decrease and fragmentation will increase in the future as a result of climate change and land-use change, but these studies also reveal regions of relatively minor difference between current and future connectivity value that, if protected or restored, could continue to facilitate movement despite climate change.
3.2.2.2. Connecting current to future ranges
While the previous approach aims to protect currently functional corridors because they will remain functional in the future, this approach identifies corridors that will become important for range expansion. It models current and predicted species distributions or suitable habitat and, if spatially disconnected, find the best connection between them. Studies applying this approach identify factors that could either limit movement (such as anthropogenic land uses), or facilitate movement (such as species-specific habitat suitability), and parameterize models to preferentially move through areas with the least impediments to reach future suitable areas. Some studies focus on single species or a small suite of focal species, which makes it possible to also incorporate fine-scale variables such as human population density, land cover type, topography, and species dispersal distances into the species distribution models (Razgour 2015, Coristine et al 2016, Dilts et al 2016, Gonçalves et al 2016). Other studies summarize this information for large groups of species. Lawler et al (2013) modeled current and predicted species distributions for 2903 vertebrate species in the Americas and found areas with projected high densities of climate-driven movements. Based on species distribution models of 2297 plant species, Choe et al (2017) generated climate meta-corridors for species groups with similar spatial distribution patterns that will facilitate range shifts of multiple species. A corridor's suitability can then be assessed by how many species' ranges it intersects.
3.2.2.3. Temporal corridors
Instead of only modeling connectivity between current and future habitat in one time step temporal corridors are designed to ensure that species can actually reach the new climatically suitable areas through time. To this end, several studies model how a species' climatic envelope (suitable temperature and moisture regime) moves across a landscape over several periods of simulated climate change. The predicted corridor is the chain of locations that are quantified during the simulation to be climatically contiguous for enough time to support range shifts, with new populations becoming established in locations that transition into the envelope while other populations go extinct (Williams et al 2005, Phillips et al 2008, Rose and Burton 2009, Hannah et al 2012, Pellatt et al 2012, Fleishman et al 2014). The approach therefore considers temporal connectivity in addition to spatial connectivity. To account for temporal connectivity for plants in conservation planning, Phillips et al (2008) recommended increasing the size of protected areas to encompass adjacent areas important for continued species persistence; Alagador et al (2014, 2016) conversely suggest to instead shift the location of protected areas as focal species of conservation concern are shifting their ranges.
3.2.2.4. Conservation network planning
Taking into account core habitat areas, systematic conservation planning approaches for focal species integrate climate-wise connectivity by (1) extending reserve selection algorithms to account for shifting habitat while minimizing cost and maximizing persistence of corridor targets (Alagador et al 2014, 2016), (2) minimizing the geographic distance between current and future species distributions according to focal species' dispersal ability (Faleiro et al 2013), (3) designing habitat networks that account for short-range and long-range connectivity by giving priority to pixels of high quality that contribute to local and regional connectivity (Rayfield et al 2016, Albert et al 2017), and (4) optimizing the spatial arrangement of habitat for range expansion by retaining habitat patches characterized by high movement flow and adding patches that will alleviate bottlenecks in the habitat network taking into account a species' dispersal ability (Hodgson et al 2016).
Some have argued that existing plans for reserve networks need to be vetted for climate change (Vos et al 2010, Rüter et al 2014). To do so, they developed metapopulation models that take into account current and future suitable habitat, species-specific dispersal and colonization abilities, species-specific landscape permeability, and carrying capacity for a suite of species. The purpose is to identify locations in existing networks that need to be strengthened to maintain spatial cohesion under climate change. By focusing on climate adaptation zones, benefit for species adaptation is maximized and the amount of land required to be protected is minimized (Vos et al 2010). Similarly, Brambilla et al (2017) took the approach of prioritizing habitat areas with more stable climate space (in-situ refugia) and adding habitat areas predicted to become suitable (ex-situ refugia) as well as connectivity areas for a set of alpine bird species to ultimately protect areas that will function for these species into the future.
3.2.2.5. Paleo-connections
Instead of modeling connectivity areas based on current and future species distributions, the paleo-connections approach identifies regions that likely functioned as biodiversity corridors under past climates. While there is a large body of literature about pathways of migration following the ice ages (e.g. Hewitt 2000, McLachlan et al 2005), three papers that resulted from the systematic literature search argue that the areas that connected populations under past climate regimes will also be important under future climate changes. In the case of Wu et al (2017), current bird distribution data were used to describe current patterns of diversity and these patterns were compared with simulated species richness patterns under paleoclimate models. This comparison allowed the authors to assess changes in species richness over time and delineate areas that bridged major biotas in the past. A different approach relies on a landscape genetic framework to identify dispersal corridors of a set of plant species in the late Quaternary (Fan et al 2017). Past, current, and future refugia for the paleoendemic flora also provided the basis for identifying overlapping areas of spatiotemporal connectivity (Mokany et al 2017).
3.3. Caveats
Spatial models and the resulting maps have become important in conservation planning, but it is important to understand the advantages and disadvantages (table 2). Models can be conceptually sound but depend on other highly uncertain data or modeled outcomes, including projections of future carbon emissions, how the atmosphere and oceans respond to these emissions, climate model downscaling, landcover change projections, climate envelope models for the focal species, and dispersal abilities of these species (Beier et al 2009, Rudnick et al 2012). Inherent to most models is that they cannot account for all factors driving the response. For example, even models designing corridors for individual plants to shift their ranges, do not account for specific factors such as soil type, seasonally varying soil properties like wetness (Pellatt et al 2012), or the habitat requirements and dispersal characteristics of animals that disperse the plants. Perhaps the most challenging aspect of these models is that, under climate change, novel types of climates are predicted, and forecasting how suitable these novel climates will be for existing species cannot be reliably determined (Capinha et al 2014). Because the models are predicting species' potential climate space or refugia in the future, it is not possible to validate modeled events by empirical tests.
Table 2. Advantages and disadvantages of climate-wise connectivity modeling approaches.
| Advantages | Disadvantages | |
|---|---|---|
| Focal-species based corridors | • can take advantage of newly available, fine scale climate data and georeferenced individual detections for a wide variety of species• can provide detailed guidance for species conservation and recovery planning by modeling relationships between physical variables and species occurrences | • based on uncertain species habitat preferences, life history characteristics, and climate models• often forecast species distributions for novel climates for which no empirical data can be incorporated in the original model—violating model assumptions for extrapolation• can oversimplify species requirements and miss variables that may be correlated with those included in the model (e.g. soil type, competition, symbiotic species)• intensive data requirements• to maintain computational feasibility, spatial resolution of data is frequently limited resulting in underestimating likelihood of species to persist |
| Finding climate-stable corridors | • finds areas for conservation action that will remain valuable even with climate change | • relies on the assumption that habitat conditions will exist to allow for species occupancy |
| Connecting current to future ranges: large species groups | • results in coarse-scale movement flow patterns | • relies on species ability to move through unsuitable climate space to reach final destination |
| Connecting current to future habitat: one or a few focal species | • useful for endangered species conservation | • does not account for gradual changes in climate |
| Temporal corridors | • accounts for changes in climate through time and thereby ensures that species can actually reach the new climatically suitable areas | • not applicable for connecting protected areas |
| Conservation network planning | • protect area networks are designed to function for focal species into the future• useful for vetting existing reserve network plans for climate change | • network designed to protect focal species, unclear how the rest of the community will use the resulting network |
| Paleo-connections | • long time perspective | • high uncertainty due to modeling past and current climates, and past species distributions |
| Structural connectivity | • good proxy for movement patterns of a wide range of species which makes it an efficient planning process• low levels of model uncertainty• low cost approach because input data are often readily accessible | • may not address the connectivity needs of all species• may miss opportunities in the landscape to protect connectivity because species' responses to land-use are oversimplified |
| Riparian corridors | • commonly used as movement corridors by many species of animals and plants• support important ecological processes• provide cooler and moister microclimates than the immediate surrounding• tend to span climatic gradients as they are oriented along elevational gradients• enjoy popular support for water quality and recreation benefits,• do not require modeling• method has been developed to prioritize riparian corridor for conservation• can easily be applied in combination with other approaches• resulting corridor designs are ready for on-the-ground implementation | • only covers one type of land facet and may not suffice alone for habitat connectivity demands especially in areas such as deserts where water ways are extremely ephemeral |
| Environmental gradients | • climate-gradient corridors accommodate the need for gradual change in temperatures• the climate-gradient corridor approach has been operationalized (Climate Linkage Mapper and Linkage Priority Tool)• environmental gradients are surrogates for key ecological and evolutionary processes that will ensure resilience to climate change• resulting corridor designs are ready for on-the-ground implementation | • climate-gradient corridors are not applicable in landscapes with little topography |
| Naturalness-based corridors | • flexible with respect to termini (either protected areas or node-less approach)• can incorporate different additional concepts (e.g. climate analogs, slope, wildness, ecological integrity, ecosystem representation)• human land use map exists for entire United States; software to operationalize the approach exists and is being improved• resulting corridor designs can guide on-the-ground implementation | • influence of naturalness on resistance to movement likely varies greatly between species• when operationalized using circuit theoretical algorithm, interpretation of dispersed flow in highly permeable area is difficult; output can be difficult to explain to stakeholders |
| Land facet corridors | • does not require climate modeling• corridors will support movement by species associated with particular land facets even if the suite of species in an area changes with climate change• based on globally available data (digital elevation models)• represents connectivity for focal species reasonably well• applies least cost path algorithm which is intuitive• resulting corridor designs are ready for on-the-ground implementation | • few studies have applied land facets and approach output is not well studied• small species with poor dispersal capabilities or species with narrow distributions may not be served well |
| Lattice-work corridors | • does not require a GPS, or modeling• intuitive, easy to convey to stakeholders and therefore conducive to implementation• comprehensive coverage that includes connectivity at all scales | • relies on stakeholder input• may miss important linkages |
| Conservation network planning | • landscape-wide conservation planning: prioritizes protected areas in addition to delineating corridors• flexible with respect to ways of incorporating climate-wise connectivity• resulting network designs can be ready for on-the-ground implementation | • complex approach• siting corridors in a systematic conservation modeling framework is challenging |
| Carbon-stock corridors | • approach for climate mitigation• can take advantage of funding sources tied to climate mitigation• resulting corridor designs are ready for on-the-ground implementation | • not focusing on facilitating range shifts |
Species-focused models assume that species will disperse more successfully through suitable habitat. This may be valid for many species; however, studies have shown that during dispersal and mating-related movement, many animals readily cross land cover types avoided during daily movements (Keeley et al 2017). Another assumption is that plant movement can be promoted by maintaining habitat connectivity. Rare long-distance movements of poor dispersers have been documented that indicate that there may be chance events that move species beyond their current range that do not require habitat connectivity. Similarly, wind-dispersed species may be able to jump across unsuitable habitat (Pearson and Dawson 2005, Anderson et al 2016). Simultaneously, poorly dispersing specialist species may not be able to shift ranges as there may not be suitable habitat nearby.
Studies modeling species distributions into the future make several assumptions. Current distributions of species are assumed to be in equilibrium with climate, and bioclimatic envelopes are assumed to be constant, meaning that species cannot evolve to tolerate new climates. Species are anticipated to move directionally towards locations with analogous climates. Species-specific models of range shifts at the leading edge expect that species disperse, survive, and reproduce in each generation and consider extinction rates as negligible, assuming that newly suitable patches will become occupied within one time step (Hodgson et al 2016). These assumptions oversimplify reality, adding uncertainty into the results.
Bioclimatic envelope models often omit several aspects of biology that would affect model results. Because species' capacity for plastic or genetic response to climate change is generally not known, models do not take into account species adaptation potential (Razgour 2015), even though empirical studies indicate that some species can adapt to the changing climate (Parmesan 2006). Bioclimatic envelope models are also simplistic in that they do not consider species' interactions. While species-specific climate-wise connectivity models can be useful for informing conservation action on endangered species (e.g. California Desert Biological Conservation Framework 2016), they have high data requirements. Generally, a lack of realistic biological information decreases model accuracy. However, the species-specific approach is increasingly taking advantage of newly available, fine scale climate data and georeferenced individual detection rates for a wide variety of species. These data sets allow for very detailed modeling of the relationships between physical variables and species occurrences that could be used to provide detailed guidance for species conservation and recovery planning (Midgley et al 2010, Schumaker et al 2014, Pérez‐García et al 2017).
Structural connectivity approaches assume that areas with minimal human impact facilitate species range shifts. While high human modification will limit movement for species moving in response to changes in climate, the influence of naturalness on resistance to movement likely varies greatly between species (Krosby et al 2015). Hence, structural connectivity modeling, may not address the connectivity needs of all species or may miss opportunities in the landscape to protect connectivity because species' responses to land-use are oversimplified. And finally, in all modeling approaches, different parameter selection and selected threshold points will yield different outcomes (e.g. Belote et al 2016) emphasizing the importance of sensitivity testing in any modeling endeavor.
Coarse resolution of input data often associated with studies that cover a large geographic extent can limit the usefulness of model outputs for conservation planning and implementation on the ground (e.g. Hannah et al 2012, Alagador et al 2014, Gonçalves et al 2016). In many cases to maintain computational feasibility, spatial resolution of the data is limited resulting in underestimating the likelihood of many climate limited species to persist in cooler microhabitats (McGuire et al 2016). Because of the limited resolution of data, some studies find that species ranges will shift hundreds of kilometers (e.g. Theobald et al 2012, Lawler et al 2013, McGuire et al 2016). While it is important to conceptually consider climate-wise connectivity at this large extent, landscape planning for climate adaptation generally takes place more locally.
4. Putting models into practice
4.1. Selecting a connectivity modeling approach
Selecting the best methods for connectivity design depends on the types of nodes or areas to be connected, the conservation objectives, the desired outcome, and available data (figure 3). If protected areas need to be connected for entire ecosystems to thrive and adapt, we suggest starting with structural connectivity designs to capture the connectivity needs of the majority of species. In regions with high topographic diversity, land facet corridors (Brost and Beier 2012a), and environmental gradients (Nuñez et al 2013) are two good options. While the land facet corridor approach usually yields several corridor strands between protected areas harboring suitable habitat for the species adapted to the designated land facet, environmental gradient corridors generally result in one optimally stable climate corridor. In regions with low topographic diversity, microclimatically diverse corridors that, if possible, connect protected areas with an altitudinal or latitudinal differential should be prioritized (Anderson et al 2016). If climate mitigation through retention of above ground carbon and future sequestration is the primary objective, then carbon-stock corridors can be modeled (Jantz et al 2014).
When selecting new sites for protection or to plan a conservation network the best is to take a systematic conservation network planning approach that incorporates refugia, maximizes representation of geophysical settings, prioritizes microclimatically diverse and locally connected grid cells in each geophysical category (Game et al 2011, Heller et al 2015, Anderson et al 2016), or includes macro-climatic gradients as conservation targets (Rouget et al 2006). In topographically complex regions, naturalness-based models that include information on climate analog sites and prioritize topoclimatically diverse cells (Littlefield et al 2017) are an alternative option. To increase overall landscape permeability, conservation action can be prioritized using naturalness-based connectivity models or lattice work corridors. The latter are especially effective on mountain slopes. If it is sensible to base planning on focal species, we recommend selecting a suite of species that represents different life history strategies and ecological functions (Beier et al 2008), and design climate-wise corridors based on the connectivity needs of these species.
For conservation action that addresses the conservation or recovery of species of conservation concern within their current range, there needs to be a focus on protecting functional connectivity that will retain suitable climate conditions for as long as possible within or adjacent to the existing distribution of the species. At the leading edge of the range, corridors should be designed that will connect current to future suitable habitat and climate conditions. For poor dispersers modeling temporal connectivity that addresses short distance movement on decadal timesteps will be important (Phillips et al 2008, Hannah et al 2012), whereas good dispersers may only need models that include one longer range time step (e.g. from 2000–2070; Pellatt et al 2012). All of these species-based approaches are data-intensive in that they require species distribution data and down-scaled current and predicted climate models.
Riparian corridors should be included in connectivity plans in most environments because of their importance as natural movement corridors, climate gradients, and refugia (Beier 2012). By combining results from structural connectivity and species-focused approaches entire ecosystems can be addressed as well as particular focal species of interest, or species whose connectivity needs are not covered by the structural corridors.
For all designs, it is important to provide live-in habitat in the corridors (Beier et al 2008, Mackey et al 2008, Beier 2012), implying that wide landscape linkages (e.g. > 1 km) will be more functional than narrow corridors. Making corridors as wide as possible is a simple way to ensure that they contain a diverse topography that provides micro-refugial sites for species persistence (Jewitt et al 2017). Quantifying the impact of natural and anthropogenic barriers on possible range shifts could inform management strategies within corridors.
4.2. Including refugia in climate-wise connectivity design
Due to the importance of refugia for species persistence during climate change several approaches have been developed to incorporate refugia into connectivity designs. When modeling climate-wise corridors between protected areas, refugia can be integrated into the corridor models by mapping the rate of climate change to identify in-situ refugia. Coristine et al (2016) determined the climate vulnerability by assessing the change and variability of several climate variables relevant to pollinator species over the past decades. Grid cells with lower rates and reduced climatic variability (refugia) were assigned lower resistance values than cells with higher rates and increased variability of climate change to favor the conservation of corridors with refugial value. A different way to include refugia when designing climate-wise connectivity is to identify in-situ refugia and model connectivity between them. Maher et al (2017) applied this approach to mountain meadows, characterizing connectivity based on distance, topography, watercourses, and roads.
During systematic conservation planning different types of refugia (e.g. riparian, drought, or evolutionary refugia), can be identified and included as targets in systematic conservation planning efforts (Cowling et al 2003, Klein et al 2009). Alternatively, the landscape can be characterized based on topographic and climatic factors. Mapped grid cells with minimal difference between current and future conditions can then be categorized as in-situ refugia and assigned a high value. When using a systematic conservation planning algorithm cells with higher values are more likely to be included in an optimal reserve network (Game et al 2011). Similarly, by prioritizing sites characterized by high topographic diversity and elevation gradients and connected by natural cover, Anderson et al (2014) integrated microclimate refugia and connectivity into systematic conservation plans.
The various modeling approaches can be combined in some instances. For example, for modeling structural connectivity in the Mojave Desert, Gallo and Greene (2018) assigned lower resistance values to cells with lower rates of climate change over time as well as higher priority to linkages that connected core refugia areas. In this case, the refugia locations were identified as a function of climate stability and physical parameters including terrain ruggedness, solar radiation, riparian vegetation, waterbodies, distance to water, and spring locations.
For species-specific connectivity models, refugia can be identified based on environmental variables meaningful for the focal species. To determine areas where wolverines are predicted to persist in the western United States and Canada, McKelvey et al (2011) modeled persistent spring snow pack, an environmental condition critical to the wolverine's life cycle, under future climate scenarios and found the least cost paths between the refugia. Bioclimatic envelope modeling has been applied to predict geographic ranges of organisms as a function of climate (Rose and Burton 2009, Pellatt et al 2012). When these models are combined with dispersal models, connections to ex-situ refugia can be modeled for particular species (Vos et al 2008). Mapping refugia can be applied to protected area prioritization, optimizing networks of in-situ refugia, as well as guide management decisions with respect to increasing connectivity to facilitate colonization of new habitat (Vos et al 2008, Pellatt et al 2012, Brambilla et al 2017).
4.3. Tools for climate-wise connectivity modeling
There are numerous software programs available to operationalize the connectivity models (conservationcorridor.org). Here, we present a brief, but by no means complete, overview of tools that have been applied to or developed specifically for climate-wise connectivity modeling focused on tools that were used in the publications reviewed here.
In many of the reviewed studies that modeled either structural or focal species-based connectivity, least cost path and circuit theory-based analyses were used to identify corridors (e.g. Brost and Beier 2012b, Belote et al 2016, Coristine et al 2016). Based on a GIS layer of termini (e.g. protected areas, climate-analog areas, or current and future suitable habitat core areas,) and resistance surfaces which specify the degree to which a grid cell facilitates or inhibits movement, these algorithms highlight areas estimated to have relatively high probability of use as movement corridors. Several software programs are available such as Circuitscape (McRae 2006), the CorridorDesigner ArcGIS toolbox in ArcGIS (Majka et al 2007), and Linkage Mapper (www.circuitscape.org/linkagemapper) to assist with these types of spatial analyses. The latter also now includes two modules which operationalize the climate gradient corridor approach: Climate Linkage Mapper optimizes the route of linkages to minimize the climate gradient traversed (Nuñez et al 2013), and the Linkage Priority Tool prioritizes linkages with climate gradients expected to facilitate species range shifts (Gallo and Greene 2018). The new software Gflow (Leonard et al 2016) extends the utility of Circuitscape to compute connectivity in large-extent and high-resolution landscapes by computing circuit theory-based connectivity simultaneously on many processors, and new advances in Circuitscape include the ability to connect climate analogs and climate gradients (Littlefield et al 2017).
Unicor (Landguth et al 2012) can be used to prioritize corridors that promote species persistence by predicting the importance of locations across the landscape for providing long-distance dispersal connectivity among core habitat patches (Wasserman et al 2013). The packages Conefor (Saura and Torne 2009), Graphab (Foltête et al 2012), and IGRAPH (Csardi and Nepusz 2006) use the principles of graph theory to rank the importance of core areas and linkages for maintaining or improving connectivity across a network (Dilts et al 2016, Kang et al 2016, Rayfield et al 2016). The new Linkage Priority Tool in Linkage Mapper (Gallo and Greene 2018) is designed to assign and map high priority linkages connecting one or more core areas with high climate refugia value, characterized by high topographic heterogeneity and/or high climate stability.
Metapopulation models, especially the habitat network assessment tool LARCH (Landscape Ecological Rules for the Configuration of Habitat, Verboom and Pouwels 2004) have been applied to evaluate landscape connectivity for focal species in fragmented landscapes and design a network of existing and proposed areas that will facilitate focal species range shifts (Vos et al 2010). These models take into account habitat quality, species' traits such as dispersal distance, reproductive potential and life span, and the effects of barriers and stepping stones.
For systematic conservation planning, software programs commonly adjusted to include climate-wise components are Zonation (Moilanen et al 2005) and Marxan (Ball et al 2009). Zonation retains the most valuable cells for multiple species while also accounting for connectivity. Papers in this review that have applied Zonation include Fleishman et al (2014) and Albert et al (2017). Marxan, applied among others by Fung et al (2017) and Heller et al (2015), designs cost-effective reserve systems based on the principles of comprehensiveness, representativeness and adequacy.
5. Conclusion
There are many different approaches to climate-wise connectivity planning that already exist. Several papers that compare the results of older and newer methods show that, based on study-defined criteria, the new methods are often an improvement (Rouget et al 2006, Alagador et al 2014, Heller et al 2015, Drake et al 2017). However, very few studies compare the results of two models to each other, and there is a need for studies that compare different approaches to elucidate differences and resolve which approaches are most appropriate in which contexts. Useful comparisons across methods are presented in Brost and Beier (2012a) and Krosby et al (2015), who compare focal species connectivity designs to land facet and naturalness-based corridors, respectively. In general, the naturalness and land facet corridors represented connectivity for focal species reasonably well, although small species with poor dispersal capabilities or species with narrow distributions were not served well. On the other hand, focal species linkages did not capture connectivity for the land facets well. Both studies concluded that structural connectivity approaches are a good proxy for species movements. However, if data are available for a suite of representative species (Beier et al 2008), corridors based on functional connectivity may better capture movement needs of all species (Krosby et al 2015), while the land facet corridors should be designed as a complement to the focal species corridors (Brost and Beier 2012a). While structural connectivity models make fewer assumptions than species-focused models and have less uncertainty, studies evaluating their functionality for facilitating range shifts of different species types are needed. Because structural connectivity is being designed for facilitating future range shifts, simulation studies taking advantage of data-intensive species models may be an option.
It is important to note that most approaches to climate-wise connectivity modeling depend on high-resolution GIS data layers characterizing the landscape based on, for example, human influence, current climates, downscaled climate models, climate velocity, topographic diversity, and geodiversity. While these layers are available for some regions, increasing world-wide coverage of higher resolution data to facilitate connectivity planning are vital. In sum, while there are existing methods and applicable data for many places, there needs to be a conscious effort to incorporate climate-wise concepts and modeling approaches into landscape planning efforts to ensure that connectivity conservation action will result in more climate resilient landscapes.
Acknowledgments
Funding was provided by the California Natural Resources Agency as part of California's Fourth Climate Change Assessment. We thank Lee Hannah and Sam Veloz for early input, two anonymous reviewers for thoughtful comments that improved the manuscript, and the Berkeley Connectivity Journal Club for helpful discussions. Publication was made possible in part by support from the Berkeley Research Impact Initiative (BRII) sponsored by the UC Berkeley Library.