
Abstract
Animals have varied taurine biosynthesis capability, which was determined by activities of key enzymes including cysteine dioxygenase (CDO) and cysteine sulfinate decarboxylase (CSD). However, whether CDO and CSD are differentially regulated across species remains unexplored. In the present study, we examined the regulations of CDO and CSD in rainbow trout and Japanese flounder, the two fish species with high and low taurine biosynthesis ability respectively. Our results showed that the expression of CDO was lower in rainbow trout but more responsive to cysteine stimulation compared to that in Japanese flounder. On the other hand, both the expression and catalytic efficiency (kcat) of CSD were higher in rainbow trout than those of Japanese flounder. A three-residue substrate recognition motif in rainbow trout CSD with sequence of F126/S146/Y148 was identified to be responsible for high kcat, while that with sequence of F88/N108/F110 in Japanese flounder led to low kcat, as suggested by site-directed mutagenesis studies. In summary, our results determined new aspects of taurine biosynthesis regulation across species.
Similar content being viewed by others

Introduction
Taurine (2-amino ethanesulfonic acid) is one of the most abundant free amino acids in vertebrates1. It plays important roles in multiple physiological processes including bile salt conjugation2, osmoregulation3, calcium homeostasis4, and trophism in the development of central nervous system5. Taurine is synthesized endogenously from sulfur amino acids such as methionine and cysteine. The major pathway of taurine de novo synthesis involves the sequential oxidation of cysteine to cysteine sulfinic acid (CSA) by cysteine dioxygenase (CDO, EC 1.13.11.20), decarboxylation by cysteine sulfinate decarboxylase (CSD, EC 4.1.1.29), and oxidation of the resulting hypotaurine to taurine by a putative hypotaurine dehydrogenase, which remains uncharacterized6,7. In this metabolic pathway, CDO and CSD has been characterized as the key enzymes that determine taurine biosynthesis capability8. Factors that influence taurine biosynthesis enzymes activities include hormone status9,10, development stages11, osmotic conditions12 and diet13,14.
Taurine biosynthesis ability varies greatly among species15,16. Livers from dog and rat have a high concentration of all enzymes required for taurine biosynthesis while those from man, monkey and cat exhibit extremely low activity of CSD1,17,18. A wide range of CSD activities was also obtained in different fish species16. Taurine biosynthesis is high in rainbow trout19 but low in Japanese flounder20 and turbot21. As a result, low biosynthesis ability makes taurine an essential nutrient for many species. In cats, many defects associated with taurine deficiency have been observed, such as retinal degeneration22, impairment of reproduction23, abnormal development24 and dilated cardiomyopathy25. Meanwhile, dietary taurine supplementation stimulated growth on multiple fish species, such as rainbow trout26, Japanese flounder27, turbot21, cobia28, and yellowtail29. In addition, taurine supplementation improved metamorphosis of Solea senegalensis larvae30.
To date, the differential taurine biosynthesis across species has been largely attributed to the activities of CDO and CSD enzymes, but the exact underlying mechanism has not been explored. Dietary sulfur amino acids stimulated taurine biosynthesis in rainbow trout19 but not in Japanese flounder20. Our previous study suggested that the response of CDO activities to dietary sulfur amino acids was less sensitive in turbot than that in mammals21. These results provide clues that the taurine biosynthesis might be differentially regulated among species.
Rainbow trout and Japanese flounder are teleost with high and low taurine biosynthesis respectively in spite of the similar zoological status and feeding habits16, therefore can serve as good model for comparative taurine biosynthesis studies across species. In the present study, the primary sequences of CDO and CSD in these species were identified. The expression and activities of CDO and CSD in fish livers were determined. The responses of CDO to cysteine stimulation were characterized. The kinetics of recombinant CDO and CSD proteins were also investigated.
Results
cDNA Cloning of CDO and CSD in rainbow trout and Japanese flounder
In the present study, the full-length cDNAs of CDO and CSD from rainbow trout and Japanese flounder were cloned. The full-length cDNA of rainbow trout CDO was 817 bp, with an open reading frame (ORF) of 600 bp encoding 200 amino acids (GenBank Accession No. KP739883). The full-length cDNA of Japanese flounder CDO was 747 bp, with an ORF of 603 bp encoding 201 amino acids (GenBank Accession No. KP739882). The CDO amino acid sequence between rainbow trout (Oncorhynchus mykiss) and Japanese flounder (Paralichthys olivaceus) shared 84% identity. Multiple sequence alignment was done to compare CDO sequences across species, including rainbow trout (Oncorhynchus mykiss), Japanese flounder (Paralichthys olivaceus), zebrafish (Danio rerio, Q6NWZ9), amphibian (Xenopus laevis, NP_001083506), sauropsida (Anolis carolinensis, XP_003223077), chicken (Gallus gallus, XP_424964), mouse (Mus musculus, NP_149026), Rat (Rattus norvegicus, AAH70509), and human (Homo sapiens, AAH24241). As shown in Fig. 1, the primary sequence of CDO showed high homology across species. All CDO orthologs from various species contained two cupin motifs. The residues making of Cys-Tyr cofactor, a unique post-translational modification of CDO, are conserved in all species.

Highly conserved residues are indicated with black backgrounds and similar residues are shaded. The cupin superfamily motifs are indicated in the top line. Dashes indicate gaps. Asterisks show highly conserved cysteine and tyrosine residues formation of cross-linked cofactor of CDO. From top to bottom, the sequences are from Paralichthys olivaceus (KP739882), Oncorhynchus mykiss (KP739883), Danio rerio (Q6NWZ9), Mus musculus (NP_149026), Rattus norvegicus (AAH70509), Homo sapiens (AAH24241), and Xenopus laevis (NP_001083506)
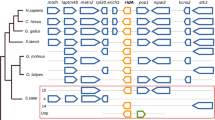
The full-length cDNAs of CSD isolated from rainbow trout and Japanese flounder were 1942 bp and 1751 bp respectively. The ORFs of rainbow trout and Japanese flounder CSD were 1575 bp encoding 525 amino acids (GenBank Accession No. KP739885) and 1461 bp encoding 487 amino acids (GenBank Accession No. KP739884) respectively. The predicted amino acid sequence of rainbow trout and Japanese flounder CSD shared 79% identity. Multiple alignment of amino acid sequences of CSD was conducted across species including rainbow trout, Japanese flounder, zebrafish (Danio rerio, NP_001007349), amphibian (Xenopus laevis, XP_002936687), sauropsida (Anolis carolinensis, XP_003216712), chicken (Gallus gallus, XP_423847), rat (Rattus norvegicus, NP_068518), cat (Felis catus, XP_006933794) and human (Homo sapiens, NP_057073). As shown in Fig. 2, a conserved enzymatic domain of DOPA decarboxylase and a pyridoxal phosphate (PLP) binding sequence (Asn-Pro-His-Lys, NPHK) were found in all species. The N-terminal region of CSD varied across species. As a member of the aspartate aminotransferase fold type I superfamily, a three-residue substrate recognition motif was present in CSD (Fig. 2). The three-residue F/S/Y motif was conserved in most species, including rainbow trout (F126, S146 and Y148). However, this motif was presented as F/N/F in Japanese flounder (F88, N108 and F110), yellowtail (Seriola quinqueradiata), medaka (Oryzias latipes), large yellow croaker (Larimichthys crocea), half-smooth tongue sole (Cynoglossus semilaevis), and bicolor damselfish (Stegastes partitus) (Fig. 3).

Highly conserved residues are indicated with black backgrounds and similar residues are shaded. Dashes indicate gaps. The conserved DOPA decarboxylase domain are indicated in the top line. The substrate recognition motif are indicated in the bottom line. Asterisks show the conserved residues in PLP binding site. From top to bottom, the sequences are from Paralichthys olivaceus (KP739884), Takifugu rubripes (ABF22453), Danio rerio (NP_001007349), Cyprinus carpio (BAE73113), Oncorhynchus mykiss (KP739885), Mus musculus (NP_659191), Rattus norvegicus (NP_068518), and Homo sapiens (NP_057073)

The three key residues in motif are bold. From top to bottom, the sequences are from insect, echinoderm, fish, amphibian, sauropsida, chicken and mammals
Hepatic expression and activities of CDO and CSD in rainbow trout and Japanese flounder
A method for absolute quantification of cDNA using real-time PCR was used to compare the mRNA level of CDO and CSD in livers of rainbow trout and Japanese flounder31. As shown in Fig. 4, the copy number of CDO in per milligram oligo-dT primed cDNA of rainbow trout was 6.8 × 105 while the value of Japanese flounder was 1.57 × 106 (Fig. 4a). On the other hand, CSD mRNA levels in rainbow trout liver (9.8 × 105 copies per mg cDNA) were significantly higher than that in Japanese flounder (2.3 × 105 copies per mg cDNA, Fig. 4b).

Copy numbers of CDO and CSD in livers of rainbow trout (RT) and Japanese flounder (JF) were measured by a method of absolute quantification of cDNAs. The results are expressed as copies per μg oligo-dT primed cDNA and the data are presents as means ± S.E.M.(n = 6) (a–b). Protein abundance of CDO and CSD in fish livers was analysed using western blot. Aliquots of 20 μg protein was loaded in each line and β-tubulin was used as a loading control (c). The relative protein abundance of CDO and CSD in rainbow trout (RT) livers were normalized to the total protein loading and the expression levels in Japanese flounder (JF) were expressed as relative expression values to those in RT group. The data are expressed as means ± S.E.M.(n = 4) (d,e). The differences between experimental groups are tested using independent t tests. **p < 0.01, ***p < 0.001
The protein abundance of CDO and CSD in livers of rainbow trout and Japanese flounder were also detected (Fig. 4c–e). The relative protein level of CDO in rainbow trout was accounted for approximately 27% of that in Japanese flounder. However, the CSD protein level in rainbow trout was approximately 1.9 fold to that in Japanese flounder.
The hepatic activities of CDO and CSD were also examined. As shown in Fig. 5a, the CDO activity in rainbow trout liver was 3.67 ± 0.15 nmol CSA/min/mg protein, which was signigicantly lower than that in Japanese flounder (7.99 ± 0.45 nmol CSA/min/mg protein). On the other hand, the hepatic CSD activity in rainbow trout (0.75 ± 0.04 nmol hypotaurine/min/mg protein) was approximately 3.3 fold to that in Japanese flounder (0.22 ± 0.01 nmol hypotaurine/min/mg protein, Fig. 5b).

The enzyme activities of CDO (a) are expressed as nmol CSA per min per mg protein and the enzyme activities of CSD (b) are expressed as nmol hypotaurine per min per mg protein. The data are expressed as means ± S.E.M.(n = 6). The differences between experimental groups of rainbow trout (RT) and Japanese flounder (JF) are tested using independent t tests. **p < 0.01; ***p < 0.001
Differential regulation of cellular CDO expression
Upon cysteine stimulation, CDO forms the intramolecular Cys-Tyr cofactor (mature form), which appeared as a lower band in SDS-PAGE compared to the form without cofactor (immature form)14. The formation of Cys-Tyr cofactor could increase the catalytic efficiency by approximately 10 fold over the cofactor-free CDO32, which is critical in maintaining cysteine concentration below toxic levels33. In this study, fish CDOs were transfected into HepG2 cells, which contained no endogenous CDO activity34, thus provided a clean background for CDO activity measurement.
Both fish CDOs showed responses after cells were treated with serial cysteine levels in medium. As shown in Fig. 6a, mature form of CDO with Cys-Tyr cofactor (shown as the lower band in SDS-PAGE) induced by cysteine was observed. However, CDO expression among species was differentially regulated. The protein levels of rainbow trout CDO increased 2.9 fold when cysteine levels were increased from 0 to 1 mM (Fig. 6b). On the contrast, the Japanese flounder CDO increased 1.4 fold when cysteine levels were increased from 0 to 0.1 mM and then decreased when cysteine levels from 0.1 to 1 mM (Fig. 6c). More importantly, the changes of CDO expression were mainly attributed to the changes of mature form of CDO with cofactor while the immature form were much more constant. When the concentration of cysteine increased to 1 mM, rainbow trout CDO was mainly expressed as the mature form (~73%) while the ratio of CDO mature form in Japanese flounder was approximately 48% (Fig. 6b,c).

Rainbow trout (RT) and Japanese flounder (JF) CDO were transfected into HepG2 cells and the expression in response to cysteine levels (0, 0.05, 0.1, 0.3, 0.6, 1 mM) was analyzed with western bolt. Aliquots of 20 μg protein was loaded in each line and β-tubulin was used as the loading control. The overexpressed CDO was detected with antibody against FLAG (a). CDO with Cys-Tyr cofactor (mature form, the lower band) and the cofactor-free form (immature form, the upper band) were quantified using NIH Image 1.63 software respectively. Relative protein abundance was expressed as relative expression value to the cysteine-free group (b–c). The data are expressed as means ± S.E.M. (n = 3). At least triplicates were conducted for each data point. The different expression of total CDO in response to cysteine were tested using one-way ANOVA and Tukey’s multiple-range test. Different letters above the bars denote significant differences between groups at the p < 0.05 level
Kinetic characterization of CDO and CSD
Both CDO and CSD proteins from rainbow trout and Japanese flounder were recombinantly expressed in E.coli and purified (Fig. 7a,b). The enzyme kinetics of CDO was determined using a wide range of substrate concentration (0–20 mM cysteine, Fig. 7c). Our result demonstraed a two phase CDO kinetics, a Michaelis-Menten model at low cysteine concentration (0–4 mM, Fig. 7d), and a substrate inhibition model at high cysteine concentration (>4 mM). The Km of rainbow trout CDO and Japanese flounder CDO for cysteine was 0.79 ± 0.09 mM and 1.23 ± 0.15 mM respectively. The value of kcat of CDO was 16.72 ± 0.67 s−1 in rainbow trout and 29.36 ± 1.52 s−1 in Japanese flounder.

Recombinant rainbow trout (RT) and japanese flounder (JF) CDO were separated by 4-20% TruPAGETM Precast Gels (a). The wild-type (wt) and site-mutants (sm) of CSD proteins were separated by 10% SDS-PAGE gel (b). Activity of recombinant rainbow trout (RT) CDO and Japanses flounder (JF) CDO (0.2uM) was investigated using a wide substrate range (0-20 mM), which revealed a two phase kinetics (c). The kinetic parameters of fish CDOs were analyzed according to Michaelis-Menten results of CDO activities at cysteine concentrations below 4 mM (d). Activity of recombinant fish CSDs (0.5uM) at a CSA concentration of 0–15 mM were measured and it was with well-behaved Michaelis-Menten kinetics. Kinetic parameters of the wild type (WT) and the S146N/Y148F mutant of CSD from rainbow trout were analyzed according to Michaelis-Menten results (e). The kinetic parameters of the wild type (WT) and the N108S/F110Y mutant of CSD from Japanese flounder were analyzed according to Michaelis-Menten results (f). The kinetic characterization of recombinant proteins were estimated using nonlinear fitting by Prism 5 software (Graphpad software). The data are expressed as means ± S.E.M. (n = 3)
The CSD activity was measured using a CSA concentration of 0–15 mM. As shown in Fig. 7e,f, according to the Michaelis-Menten equation, the Km of rainbow trout and Japanese flounder CSDs were 2.30 ± 0.24 mM and 3.45 ± 0.38 mM respectively. The value of kcat of CSD in rainbow trout and Japanese flounder was 4.72 ± 0.36 s−1 and 1.52 ± 0.13 s−1 respectively.
Sequences alignment analysis showed that the key residues within the active sites of fish CSDs were not well conserved, F126/S146/Y148 in rainbow trout versus F88/N108/F110 in Japanese flounder. In order to explore their impact on CSD catalytic activity, these key residues were switched between the two fishes by site-directed mutagenesis. The two CSD variants were also recombinantly expressed, purified and kinetic characterized. The Km of rainbow trout CSD mutant (S146N/Y148F) was 2.73 ± 0.27 mM and the kcatwas 1.86 ± 0.07 s−1 (Fig. 7e), which were similar to those of the wild-type Japanese flounder CSD. The Km of Japanese flounder CSD mutant (N108S/F110Y) was 1.83 ± 0.17 mM and its kcatwas 6.26 ± 0.20 s−1 (Fig. 7f), which were similar to those of the wild-type rainbow trout CSD.
Discussion
Taurine biosynthesis is controlled by two key enzymes, CDO and CSD8. To date, CDO and CSD have been identified and characterized from mammals to eubacteria32,35,36,37,38, suggesting the taurine biosynthesis pathway via CDO and CSD is highly conserved during evolution. Given the fact that taurine biosynthesis ability varies greatly among species, it will be interesting to examine whether the regulation of CDO and CSD can be different among species. In the present study, we examined CDO and CSD from rainbow trout and Japanese flounder, two fish species with high and low taurine biosynthesis ability respectively in spite of the similar zoological status and feeding habits16.
We firstly characterized the full primary sequence of both CDO and CSD from rainbow trout and Japanese flounder. CDO is a member of the cupin superfamily composed of a wide range of functional proteins with two conserved cupin motifs for metal binding39,40. Both cupin motifs were identified in CDOs of rainbow trout and Japanese flounder (Fig. 1). In addition, the Cys-Tyr residues for intra-molecular crosslink41,42 were also highly conserved. CSD is a member of the group II pyridoxal 5′ - phosphate (PLP)-dependent amino acid decarboxylases family43,44. The DOPA decarboxylase domain and the NPHK sequence motif35 were both conserved in CSDs of rainbow trout and Japanese flounder.
Previously, taurine biosynthesis ability was only attributed to the activities of CDO and CSD13,45. Here we showed a far more complicated mechanism. Rainbow trout is low in CDO but high in CSD when compared with Japanese flounder, suggesting that CDO and CSD might play differential roles in regulation of taurine biosynthesis. It is known that CDO and CSD were differentially regulated by sulfur amino acids. High levels of dietary sulfur amino acids could markedly increase CDO activities and stimulated taurine biosynthesis46, while decreased CSD activities47. It remains to be investigated whether the different levels of CDO and CSD between rainbow trout and Japanese flounder is the result of evolutional adaptation for diets and osmotic conditions, both of which regulating CDO and CSD levels12,46,47.
CDO is known as one of the most highly regulated metabolic enzymes responding to sulfur amino acids33. Its concentrations increase robustly with elevated cysteine levels in diet47. However, this response seems to be different among species21. In this study, the differential regulations of fish CDOs exogenously expressed in HepG2 cells were examined. The response of rainbow trout CDO to cysteine stimulation increased ~2.9 fold when cysteine levels increased from 0 to 1 mM. This was much more dramatic than that of Japanese flounder (~1.4 fold). An analogous study showed that rat CDO exogenously expressed in HepG2/C3A cells increased 1.6 fold upon the same stimulation34. It is known that CDO concentrations are regulated by cysteine levels via the formation of intramolecular Cys-Tyr cofactor, which could dramatically increase CDO’s catalytic efficiency14,48 and stability49. In this study, the crosslink formation efficiency between the two fishes was different (~73% crosslinked form CDO in rainbow trout while only ~48% in Japanese flounder at 1 mM cysteine). To date, several residues in the active site of CDO that affect the cofactor formation have been characterized14,41,42,48,50,51. Cys-93 and Tyr-157 are involved in cofactor formation directly; His-86, His-88 and His-140 are vital for iron coordination; Arg-60 is the only charged residue in the active site and probably plays a role in hydrogen bonding with cysteine. However, sequence alignment showed that all the previously identified residues in CDO active site were conserved in both rainbow trout and Japanese flounder. The reason for the differential regulation of cofactor formation among species remains to be further explored.
Due to the profound influence of cofactor formation on CDO catalytic efficiency, recombinant CDO proteins and kinetic assays in this study were performed under conditions with iron and oxygen, which favored cofactor formation14. The two fish CDOs showed comparable kinetic parameters, which might be a result of the high identities (84%) of the deduced amino acids sequences. The Km of 0.79 mM for rainbow trout CDO and 1.23 mM for Japanese flounder CDO are in line with the reported kinetic parameters of human (Km = 0.77 mM)48 and rat (Km = 0.45 mM)51. It is known that CDO plays a critical role in regulation of cysteine homeostasis33. The similarity CDO kinetic parameters among species indicate that maintaining cysteine concentration below the toxic level are important for the living organism. However, Ye and colleagues reported a Km of 3.1 mM for human CDO50, a result probably caused by differential enzyme preparations14,50. In addition, our results confirmed CDO was inhibited at high substrate concentrations, similar to what was reported for rat CDO and human CDO14,48,51. Crystal structure studies showed that Cys-164 lined in the only access of substrate to the active site. When cysteine concentrations was high (millimolar levels), cysteine could form a disulfide with Cys-164 and inhibited CDO catalysis51.
Despite of the high identities (79%) of the deduced amino acids sequences of CSD, kinetic characterization of fish CSDs showed a great difference of catalytic efficiency. The kcat of rainbow trout CSD (4.72 ± 0.36 s−1) was about 3.1 fold of that in Japanese flounder CSD (1.52 ± 0.13 s−1). Recently, a three residue substrate recognition motif (Faa19SaaY) has been identified in the active site of human CSD and was shown to dramatically affect the catalytic efficiency32. Sequence alignment of the motif showed that the residues, especially the latter two residues, were less conserved in fishes (Fig. 3). The motif was presented as F/S/Y in rainbow trout but as F/N/F in Japanese flounder and yellowtail52, which also showed a limited taurine biosynthesis capability29. In order to explore the impact of this motif variation on CSD catalytic activity, the key residues within the two fish CSDs were switched. Results of the kinetic characterization showed that the kcat of rainbow trout S146N/Y148F variant (1.86 ± 0.07 s−1) was similar to that of wild-type Japanese flounder CSD, while an analogous result was obtained in Japanese flounder. It indicates that the F/N/F motif in Japanese flounder CSD was responsible for the low catalytic activity and the limited taurine biosynthesis capability. Structural analyses of human CSD (PDB: 2JIS) showed that the phenolic side chain at X3 position of the F/S/Y motif was crucial for CSA binding, which might form a hydrogen bond with the lone pair of electrons on the sulfur atom of CSA32. In addition, the F/N/F motif was also present in many other fishes with unknown taurine biosynthesis capability, such as medaka52, large yellow croaker, half-smooth tongue sole and bicolor damselfish. Their ability of taurine biosynthesis remains to be examined.
In conclusion, our results identified the differential taurine biosynthesis ability between rainbow trout and Japanese flounder and determined the regulation of taurine biosynthesis was a far more sophisticated system than ever thought before.
Materials and Methods
Tissue collection
Rainbow trout (76.4 ± 5.8 g) and Japanese flounder (98.7 ± 7.2 g) at the same age (4 months old) were obtained from local rearing farms (Weifang and Yantai respectively, China). After overnight fasting to remove possible nutritional influences on taurine biosynthesis53,54, fish was anesthetized with 3-aminobenzoic acid ethyl ester (MS222) at 100 μg/ml. Liver, the major site of taurine biosynthesis, was rapidly removed and immediately frozen in liquid nitrogen and stored at −80 °C before use. All procedures performed in study were in strict accordance with the recommendations in the Guide for the Use of Experimental Animals of Ocean University of China. The protocols for animal care and handing used in this study were approved by the Institutional Animal Care and Use Committee of Ocean University of China.
Molecular cloning and characterization of full length cDNAs
Total liver RNA was isolated using TRizol reagent (Invitrogen). The quality and concentration of RNA were measured by Nanodrop 2000 (Thermo Fisher Scientific). First strand cDNA was produced by SMART RACE cDNA Amplification Kit (Clontech) and used as template for 3′ or 5′ rapid amplification of cDNA ends (RACE) PCR. To obtain the full length cDNAs of CDO and CSD, degenerative primers were designed on the conserved regions from different species using CODEHOP online software (http://blocks.fhcrc.org/blocks/codehop.html) firstly. Subsequently, specific primers were further designed for RACE PCR with Primer 5 software. All PCR products were separated by agarose gel and the target band was purified, and then ligated into pEASY-T1 vector (Transgen Biotech). The PCR products were sequenced in Sangon Biotech Co., Ltd (Shanghai). The full-length cDNA sequences were deposited to the GenBank database. All the primers used in this study were listed in Table 1.
Absolute mRNA quantification
In order to compare the mRNA levels of CDO and CSD in livers of juvenile rainbow trout and Japanese flounder, absolute quantification of mRNA using quantitative real-time PCR (qRT-PCR) was conducted as described by Whelan et al. (2003)31. Briefly, the PCR products, obtained from real-time PCR primers for specific target gene, were cloned into the pEASY-T1 vector. The plasmid was further transformed into DH5α competent cells (Takara). Single colonies were grown up in LB-broth medium and plasmid was subsequently purified using the SanPrep Column Plasmid Mini-Preps Kit (Sangon Biotech). Sequence verified plasmids were serially diluted and used for plasmid standards after linearized by Hind III (New England Biolabs). Single-stranded cDNA from fish tissues was synthesized using a PrimeScript® RT Reagent Kit (Takara) and the concentration of cDNA was measured by Nanodrop 2000 (Thermo Fisher Scientific).
The qRT-PCR was carried out in a quantitative thermal cycler Mastercycler ep realplex (Eppendorf). SYBR Green real-time PCR kit (Takara) was used. The melting curve was performed after the amplification phase for confirmation the specificity of production. The plasmid standards and tissue cDNA samples were detected in a same plate and each sample was run in triplicate. Target gene expression levels were quantitated as copy number per microgram of oligo-dT primed cDNA according to the curves of plasmid standards.
Recombinant Protein expression and purification
For the purpose of producing recombinant protein in bacterial expression systems, the open reading frame (ORF) of CDO and CSD from rainbow trout and Japanese flounder were subcloned into the pET-28a expression plasmid (Novagen) using BamHI and HindIII restriction sites. Sequence verified plasmid was transformed into E.coli BL21 (DE3) competent cells (Transgen Biotech) for protein expression.
In order to explore the impact of the residues within the active sites on CSD catalytic activity, two double-sties CSD variants in the pET-28a vector were constructed by the QuikChange Lightning Site-Directed Mutagenesis kit (Stratagene). The S146 and Y148 residues in rainbow trout CSD were mutated to N and F, while the N108, and F110 residues in Japanese flounder CSD were replaced to S and Y, respectively. The mutant expression constructs were sequence verified and transformed into E.coli BL21 (DE3) competent cells for protein expression.
Transformed cells were cultured in LB medium supplemented with 50 μg/mL kanamycin at 37 °C. Expression of CDO was induced with 0.2 mM isopropyl β-D-1-thiogalactopyranoside (IPTG) at OD600 = 0.5, and continued for 4 hours at 37 °C. The cell were harvested by centrifugation at 6,000 g for 20 min at 4 °C. Expression of CSD was induced at OD600 = 0.6 with 0.5 mM IPTG at 25 °C overnight.
All expressed proteins carried an N-terminal 6 × His-tag and were purified using HisTrap HP columns (GE Healthcare) on an ÄKTA FPLC Purifier system (Amersham Biosciences). Due to the loose binding of iron in the active center of CDO, expression media and all purification buffers of CDO were supplemented with 1 mM ammonium iron sulfate to ensure proper iron saturation48. The cell pellets were resuspended in cold lysis buffer (10 mM NaH2PO4, 300 mM NaCl, 20 mM imidazole, pH 8.0). The cell suspensions were kept in an ice-water bath and sonicated (Sonics, VC130) followed by centrifugation at 12000 g for 20 min at 4 °C. The supernatants were filtered using 0.45 μm pore membranes and applied to the HisTrap HP columns. Columns were washed using a gradient elution with Buffer A (10 mM NaH2PO4, 300 mM NaCl, 20 mM imidazole, pH 8.0) and Buffer B (10 mM NaH2PO4, 300 mM NaCl, 500 mM imidazole, pH 8.0). Protein elution fractions were further purified using a Superdex 200 column (GE Healthcare) with activity buffers (CDO buffer: 100 mM NaCl, 20 mM Tris, pH 8.0; CSD buffer: 100 mM NaH2PO4,10 μM pyridoxal 5′ - phosphate, pH 7.4). The purified protein samples were concentrated ~10 fold using Amicon ultra 10 K filters (Millipore). Purified protein samples were stored at −80 °C.
Enzyme assays
Activity assays for tissue samples and recombinant CDO proteins were done as described previously55. For tissues, samples were homogenized in 50 mM 2-(N-Morpholino) ethanesulfonic acid (MES) buffer (pH 6.1) with a homogenizer and centrifuged at 25,000 g for 30 min at 4 °C. The supernatant was used for CDO enzyme assay. The activity assay was conducted in a total volume of 400 μl containing the following components: 62.5 mM MES buffer, 0.3 mM ferrous sulfate, 2 mM NAD, 5 mM hydroxylamine, 5 mM cysteine, and 62.5 μM bathocuproine disulfonate. For purified recombinant CDO proteins, the reaction mixture contained 0.2 μM purified CDO, 62.5 mM MES buffer, 0.3 mM ferrous sulfate, 62.5 μM bathocuproine disulfonate and varying concentrations of L-cysteine (0–20 mM). The reaction was started by addition of cysteine and incubated at 37 °C with vigorous shaking at 900 rpm to ensure proper oxygenation using a thermomixer (Eppendorf). The reaction was terminated by the addition of 200 μl 5% sulfosalicylic acid and the reaction mixtures were placed on ice for 15 min. After centrifugation, the reaction product, CSA, was collected in supernatant and measured by HPLC (HP 1100 system, Agilent Technologies).
CSD activities in tissue samples and recombinant proteins were measured as described previously16,32,56. Briefly, tissue samples were homogenized in 50 mM phosphate buffer (pH 6.8). The homogenates were centrifuged at 21,000 g for 15 min at 4 °C. The supernatant was used for CSD enzyme assay. The activity assay mixture (0.5 ml final volume) contained 15 mM glutamate, 25 mM CSA, 0.8 mM pyridoxal 5′ - phosphate (PLP), 0.55 mM DTT and 0.1 ml tissue homogenates. After incubation at 37 °C for 30 min, the reaction was terminated by 10% trichloroacetic acid. For recombinant CSD proteins, enzymatic assays were conducted in a final volume of 0.2 ml containing 0.5 μM purified CSD, 100 mM NaH2PO4, 10 μM PLP and varying concentrations of CSA (0–15 mM). After incubation at 37 °C for 10 min, the reaction was terminated by 20 μl of 1 M hydrochloric acid. After centrifugation, the reaction product, hypotaurine, was measured by HPLC.
The products of enzyme reactions (CSA and hypotaurine) were pre-column derivatized with o-phthaladehyde (OPA)/2-mercaptoethanol and separated with a 4.6 × 250 mm Zorbax Eclipse C18 column (Agilent Technologies) using a gradient elution. The derivatized product was detected using a fluorescence detector (FLD). The gradient mobile phase and the parameters of FLD were set as described previously55,56.
Cell culture, transfection, and treatment
For the transient expression of CDO protein in mammalian cells, the ORF of CDOs were subcloned into the pGen2.1 vector (GeneScript) using NotI and BamHI restriction sites. Sequence verified plasmid was transformed into DH5α competent cells (TakaRa) and purified by EndoFree Plasmid Maxi Kit (Qiagen). The human hepatoma cells (HepG2 cells) were obtained from the Type Culture Collection of the Chinese Academy of Sciences, Shanghai, China. Cells were maintained in DMEM (Invitrogen) supplemented with 10% FBS (Invitrogen), 1% penicillin-streptomycin, 2 mM GlutaMAXTM and 1 mM sodium pyruvate in a humidified incubator at 37 °C and 5% CO2. Cells were transfected with pGen2.1-CDO constructs using lipofectamine 3000 (Invitrogen) according to the manufacturer’s instructions. Cells transfected with empty pGen2.1 vector were used as control. After 24 hrs, transfected cells were evenly splitted and treated with experimental mediums for 24 hrs before lysis. The experimental medium was made by supplementing sulfur amino acid-free DMEM with 10% FBS, 1% penicillin-streptomycin, 2 mM GlutaMAXTM, 1 mM sodium pyruvate, 0.2 mM L-methionine and varying concentrations of L-cysteine (0, 0.05, 0.1, 0.3, 0.6, or 1 mM). Bathocuproine disulfonate (0.05 mM) was added to reduce the rate of cysteine auto-oxidation to cystine. All cell culture experiments were repeated at least three times.
Immunoblotting analysis
The protein abundance of CDO and CSD in livers of rainbow trout and Japanese flounder were quantified using western blotting. Polyclonal rabbit antibody against CDO (#C6247) was purchased from Sigma, and anti-β-tubulin (#2146) was purchased from Cell Signaling Technology. A rabbit polyclonal antiserum against CSD was generated as described previously11. Frozen tissue samples were homogenized in ice-cold RIPA buffer (50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 1 mM EDTA, 0.5% Nonidet P-40, 0.1% SDS) with protease inhibitor cocktail (Roche Applied Science). The homogenate was centrifuged at 12000 g for 30 min at 4 °C. Protein concentrations in supernatant were determined by a BCA protein assay kit (Beyotime) using bovine serum albumin as a standard. Aliquots of 20 μg protein were subjected to 12% SDS-PAGE and transferred to PVDF membrane. The membrane was blocked with 5% nonfat milk in TBST buffer (20 mM Tris-HCl, 500 mM NaCl and 0.1% Tween 20) and incubated with primary antibody overnight at 4 °C. Horseradish peroxidase (HRP)-conjugated secondary antibodies were detected using ECL reagents (GoodHere Inc., China). The density of the protein bands was quantified using NIH Image 1.63 software.
The expression of CDO in response to cysteine levels was evaluated using monoclonal mouse antibody against FLAG (#A00187, GeneScript). Cells were harvested and lysed in ice-cold RIPA buffer supplemented with protease inhibitors. The lysates were centrifuged at 12,000 g for 20 min at 4 °C. The cleared supernatants were collected and stored at −80 °C. Protein concentrations were determined with a BCA protein assay kit. Aliquots of 20 μg of total protein was loaded and separated by 4–20% TruPAGETM Precast Gels (Sigma) and followed with western bolt analysis. β-Tubulin was used as the loading control. At least triplicates were conducted for each data point.
Statistical analysis
Each value is expressed as means ± S.E.M. Statistical analysis of expression levels and activities of CDO and CSD between rainbow trout and Japanese flounder were performed by independent t tests. The different expression of CDO in response to cysteine stimulation were tested using one-way ANOVA and Tukey’s multiple-range test. The kinetic characterization of recombinant proteins were estimated using nonlinear fitting by Prism 5 software (Graphpad software). A value of P < 0.05 was considered statistically significant.
Additional Information
How to cite this article: Wang, X. et al. Differential regulation of taurine biosynthesis in rainbow trout and Japanese flounder. Sci. Rep. 6, 21231; doi: 10.1038/srep21231 (2016).
References
Jacobsen, J. G. & Smith, L. H. Biochemistry and physiology of taurine and taurine derivatives. Physiol Rev 48, 424–511 (1968).
Huxtable, R. J. Physiological actions of taurine. Physiol Rev 72, 101–163 (1992).
Lambert, I. H. Regulation of the cellular content of the organic osmolyte taurine in mammalian cells. Neurochem Res 29, 27–63 (2004).
Foos, T. M. & Wu, J. Y. The role of taurine in the central nervous system and the modulation of intracellular calcium homeostasis. Neurochem Res 27, 21–26 (2002).
Lima, L., Obregon, F., Cubillos, S., Fazzino, F. & Jaimes, I. Taurine as a micronutrient in development and regeneration of the central nervous system. Nutr Neurosci 4, 439–443 (2001).
Sumizu, K. Oxidation of hypotaurine in rat liver. Biochim Biophys Acta 63, 210–212 (1962).
Huxtable, R. J. Taurine in the central nervous system and the mammalian actions of taurine. Prog Neurobiol 32, 471–533 (1989).
de la Rosa, J. & Stipanuk, M. H. Evidence for a rate-limiting role of cysteinesulfinate decarboxylase activity in taurine biosynthesis in vivo . Comp Biochem Physiol B 81, 565–571 (1985).
Jerkins, A. A. & Steele, R. D. Cysteine sulfinic acid decarboxylase activity in response to thyroid hormone administration in rats. Arch Biochem Biophys 286, 428–432 (1991).
Jerkins, A. A., Jones, D. D. & Kohlhepp, E. A. Cysteine sulfinic acid decarboxylase mRNA abundance decreases in rats fed a high-protein diet. J Nutr 128, 1890–1895 (1998).
Wang, X., He, G., Mai, K., Xu, W. & Zhou, H. Ontogenetic taurine biosynthesis ability in rainbow trout (Oncorhynchus mykiss) . Comp Biochem Physiol B Biochem Mol Biol 185, 10–15 (2015).
Bitoun, M. & Tappaz, M. Gene expression of the transporters and biosynthetic enzymes of the osmolytes in astrocyte primary cultures exposed to hyperosmotic conditions. Glia 32, 165–176 (2000).
Tappaz, M. L. Taurine biosynthetic enzymes and taurine transporter: molecular identification and regulations. Neurochem Res 29, 83–96 (2004).
Dominy, J. E. et al. Synthesis of amino acid cofactor in cysteine dioxygenase is regulated by substrate and represents a novel post-translational regulation of activity. J Biol Chem 283, 12188–12201 (2008).
Hayes, K. C. & Sturman, J. A. Taurine in metabolism. Annu Rev Nutr 1, 401–425 (1981).
Yokoyama, M., Takeuchi, T., Park, G. S. & Nakazoe, J. Hepatic cysteinesulphinate decarboxylase activity in fish. Aquac Res 32, 216–220 (2001).
Hayes, K. C., Stephan, Z. F. & Sturman, J. A. Growth depression in taurine-depleted infant monkeys. J Nutr 110, 2058–2064 (1980).
Sturman, J. & Hayes, K. The biology of taurine in nutrition and development. Adv Nutr Res, PP 231–299 (1980).
Yokoyama, M. & Nakazoe, J.-I. Accumulation and excretion of taurine in rainbow trout (Oncorhynchus mykiss) fed diets supplemented with methionine, cystine and taurine. Comp Biochem Physiol A Physiol 102, 565–568 (1992).
Park, G.-S., Takeuchi, T., Yokoyama, M. & Seikai, T. Optimal dietary taurine level for growth of juvenile Japanese flounder Paralichthys olivaceus. Fisheries Sci 68, 824–829 (2002).
Wang, Q. et al. Dietary sulfur amino acid modulations of taurine biosynthesis in juvenile turbot (Psetta maxima). Aquaculture 422–423, 141–145 (2014).
Hayes, K. C., Carey, R. E. & Schmidt, S. Y. Retinal degeneration associated with taurine deficiency in the cat. Science 188, 949–951 (1975).
Sturman, J. A., Gargano, A. D., Messing, J. M. & Imaki, H. Feline maternal taurine deficiency: effect on mother and offspring. J Nutr 116, 655–667 (1986).
Sturman, J. A. Dietary taurine and feline reproduction and development. J Nutr 121, S166–170 (1991).
Pion, P. D., Kittleson, M. D., Rogers, Q. R. & Morris, J. G. Myocardial failure in cats associated with low plasma taurine: a reversible cardiomyopathy. Science 237, 764–768 (1987).
Gaylord, T. G. et al. Supplementation of taurine and methionine to all-plant protein diets for rainbow trout (Oncorhynchus mykiss). Aquaculture 269, 514–524 (2007).
Kim, S.-K. et al. Effect of different dietary taurine levels on the conjugated bile acid composition and growth performance of juvenile and fingerling Japanese flounder Paralichthys olivaceus . Aquaculture 273, 595–601 (2007).
Lunger, A. N., McLean, E., Gaylord, T. G., Kuhn, D. & Craig, S. R. Taurine supplementation to alternative dietary proteins used in fish meal replacement enhances growth of juvenile cobia (Rachycentron canadum). Aquaculture 271, 401–410 (2007).
Takagi, S. et al. Taurine is an essential nutrient for yellowtail Seriola quinqueradiata fed non-fish meal diets based on soy protein concentrate. Aquaculture 280, 198–205 (2008).
Pinto, W. et al. Dietary taurine supplementation enhances metamorphosis and growth potential of Solea senegalensis larvae. Aquaculture 309, 159–164 (2010).
Whelan, J. A., Russell, N. B. & Whelan, M. A. A method for the absolute quantification of cDNA using real-time PCR. J Immunol Methods 278, 261–269 (2003).
Agnello, G., Chang, L. L., Lamb, C. M., Georgiou, G. & Stone, E. M. Discovery of a substrate selectivity motif in amino acid decarboxylases unveils a taurine biosynthesis pathway in prokaryotes. ACS Chem Biol 8, 2264–2271 (2013).
Stipanuk, M. H., Ueki, I., Dominy, Jr J. E., Simmons, C. R. & Hirschberger, L. L. Cysteine dioxygenase: a robust system for regulation of cellular cysteine levels. Amino Acids 37, 55–63 (2009).
Dominy, Jr J. E., Hwang, J. & Stipanuk, M. H. Overexpression of cysteine dioxygenase reduces intracellular cysteine and glutathione pools in HepG2/C3A cells. Am J Physiol Endocrinol Metab 293, E62–69 (2007).
Kaisakia, P. J., Jerkins, A. A., Goodspeed, D. C. & Steele, R. D. Cloning and characterization of rat cysteine sulfinic acid decarboxylase. Biochim Biophys Acta 1262, 79–82 (1995).
Higuchi, M., Celino, F., Tamai, A., Miura, C. & Miura, T. The synthesis and role of taurine in the Japanese eel testis. Amino Acids 43, 773–781 (2012).
Nagasaki, T. et al. Cysteine dioxygenase and cysteine sulfinate decarboxylase genes of the deep-sea mussel Bathymodiolus septemdierum: possible involvement in hypotaurine synthesis and adaptation to hydrogen sulfide. Amino Acids 47, 571–578 (2015).
Hosokawa, Y., Matsumoto, A., Oka, J., Itakura, H. & Yamaguchi, K. Isolation and characterization of a cDNA for rat liver cysteine dioxygenase. Biochem Biophys Res Commun 168, 473–478 (1990).
Dunwell, J. M., Culham, A., Carter, C. E., Sosa-Aguirre, C. R. & Goodenough, P. W. Evolution of functional diversity in the cupin superfamily. Trends Biochem Sci 26, 740–746 (2001).
Stipanuk, M. H., Simmons, C. R., Karplus, P. A. & Dominy, Jr J. E. Thiol dioxygenases: unique families of cupin proteins. Amino Acids 41, 91–102 (2011).
McCoy, J. G. et al. Structure and mechanism of mouse cysteine dioxygenase. Proc Natl Acad Sci USA 103, 3084–3089 (2006).
Joseph, C. A. & Maroney, M. J. Cysteine dioxygenase: structure and mechanism. Chem Commun (Camb). 3338–3349 (2007).
Christen, P. & Mehta, P. K. From cofactor to enzymes. The molecular evolution of pyridoxal-5′-phosphate-dependent enzymes. Chem Rec 1, 436–447 (2001).
Sandmeier, E., Hale, T. I. & Christen, P. Multiple evolutionary origin of pyridoxal-5′-phosphate-dependent amino acid decarboxylases. Eur J Biochem 221, 997–1002 (1994).
Worden, J. A. & Stipanuk, M. H. A comparison by species, age and sex of cysteinesulfinate decarboxylase activity and taurine concentration in liver and brain of animals. Comp Biochem Physiol B 82, 233–239 (1985).
Bagley, P. J. & Stipanuk, M. H. Rats fed a low protein diet supplemented with sulfur amino acids have increased cysteine dioxygenase activity and increased taurine production in hepatocytes. J Nutr 125, 933–940 (1995).
Bella, D. L., Hahn, C. & Stipanuk, M. H. Effects of nonsulfur and sulfur amino acids on the regulation of hepatic enzymes of cysteine metabolism. Am J Physiol 277, E144–153 (1999).
Arjune, S., Schwarz, G. & Belaidi, A. Involvement of the Cys-Tyr cofactor on iron binding in the active site of human cysteine dioxygenase. Amino Acids 47, 55–63 (2015).
Dominy, Jr J. E., Hirschberger, L. L., Coloso, R. M. & Stipanuk, M. H. Regulation of cysteine dioxygenase degradation is mediated by intracellular cysteine levels and the ubiquitin-26 S proteasome system in the living rat. Biochem J 394, 267–273 (2006).
Ye, S. et al. An insight into the mechanism of human cysteine dioxygenase. Key roles of the thioether-bonded tyrosine-cysteine cofactor. J Biol Chem 282, 3391–3402 (2007).
Simmons, C. R. et al. Crystal structure of mammalian cysteine dioxygenase. A novel mononuclear iron center for cysteine thiol oxidation. J Biol Chem 281, 18723–18733 (2006).
Haga, Y. et al. Isolation, molecular characterization of cysteine sulfinic acid decarboxylase (CSD) of red sea bream Pagrus major and yellowtail Seriola quinqueradiata and expression analysis of CSD from several marine fish species. Aquaculture 449, 8–17 (2015).
Kaushik, S. & Luquet, P. Study of free amino acids in rainbow trout in relation. Ann Hydrobiol 8, 135–144 (1977).
Lee, J.-I., Londono, M., Hirschberger, L. L. & Stipanuk, M. H. Regulation of cysteine dioxygenase and γ-glutamylcysteine synthetase is associated with hepatic cysteine level. J Nutr Biochem 15, 112–122 (2004).
Stipanuk, M. H., Dominy, Jr J. E., Ueki, I. & Hirschberger, L. L. Measurement of Cysteine Dioxygenase Activity and Protein Abundance. Curr Protoc Toxicol 38, 6.15.11–16.15.25 (2008).
Coloso, R. M., Hirschberger, L. L., Dominy, Jr J. E., Lee, J. I. & Stipanuk, M. H. Cysteamine dioxygenase: evidence for the physiological conversion of cysteamine to hypotaurine in rat and mouse tissues. Adv Exp Med Biol 583, 25–36 (2006).
Acknowledgements
This research was supported by the National Scientific Foundation of China grant (No.31172416), the National Basic Research Program of China (973 Program) Grant (No. 2014CB138600) and Fok Ying-Tong Education Foundation grant (131024) to G.H.
Author information
Authors and Affiliations
Contributions
G.H. and K.M. designed the research. G.H. and X.W. conducted the research and wrote the paper. W.X. and H.Z. provided technical assistance and contributed to the preparation of the figures. All authors analyzed the results and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Wang, X., He, G., Mai, K. et al. Differential regulation of taurine biosynthesis in rainbow trout and Japanese flounder. Sci Rep 6, 21231 (2016). https://doi.org/10.1038/srep21231
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep21231
This article is cited by
-
Taurine synthesis via the cysteic acid pathway: effect of dietary cysteic acid on growth, body taurine content, and gene expression of taurine-synthesizing enzymes, growth hormone, and insulin-like growth factor 1 in Japanese flounder Paralichthys olivaceus
Fisheries Science (2021)
-
Roles of dietary taurine in fish nutrition
Marine Life Science & Technology (2020)
-
Stimulatory effect of dietary taurine on growth performance, digestive enzymes activity, antioxidant capacity, and tolerance of common carp, Cyprinus carpio L., fry to salinity stress
Fish Physiology and Biochemistry (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
