
Abstract
The autonomy of segment-specific regulatory domains in the Bithorax complex is conferred by boundary elements and associated Polycomb response elements (PREs). The Fab-6 boundary is located at the junction of the iab-5 and iab-6 domains. Previous studies mapped it to a nuclease hypersensitive region 1 (HS1), while the iab-6 PRE was mapped to a second hypersensitive region HS2 nearly 3 kb away. To analyze the role of HS1 and HS2 in boundary we generated deletions of HS1 or HS1 + HS2 that have attP site for boundary replacement experiments. The 1389 bp HS1 deletion can be rescued by a 529 bp core Fab-6 sequence that includes two CTCF sites. However, Fab-6 HS1 cannot rescue the HS1 + HS2 deletion or substitute for another BX-C boundary – Fab-7. For this it must be combined with a PRE, either Fab-7 HS3, or Fab-6 HS2. These findings suggest that the boundary function of Fab-6 HS1 must be bolstered by a second element that has PRE activity.
Similar content being viewed by others


Introduction
The Drosophila Bithorax complex (BX-C) has three homeotic genes, Ultrabithorax (Ubx), abdominal-A (abd-A) and Abdominal-B (Abd-B), the pattern of expression of which in the posterior parasegments/segments, PS5-14/T3-A9 determines their identity1,2,3. This expression pattern is generated by a unique collection of tissue specific enhancers that are located in nine functionally autonomous cis-regulatory domains. Two of these domains, abx/bx and bxd/pbx, direct Ubx expression in PS5 (T3) and PS6 (A1). Three domains, iab-2, iab-3 and iab-4 control abd-A in PS7-9 (A2-4), respectively. Finally Abd-B expression in PS10-13 (A5-9) is controlled by four regulatory domains, iab-5, iab-6, iab-7 and iab-8,9 respectively3,4 (Fig. 1a).

Deletions in the region of the Fab-6 boundary. (a) Morphology of the male abdominal segments in wild type (wt), F61attP and F61+2attP alleles. The organization of the abd-A and Abd-B regulatory regions in Bithorax complex is shown near wt abdominal segments. The abd-A (blue) and Abd-B (green) promoters are schematically demonstrated by arrows. The abdominal segments and iab-domains that provide their features are shown (more in the text). A7 and A8 are absent in wt adult males. A9 is a part of male genitals. The lines with colored circles mark characterized (Fub, Mcp, Fab-6, Fab-7, and Fab-8) and predicted (Fab-3, and Fab-4) boundaries. The red circles indicate number of the CTCF binding sites in each boundary. F61attP and F61+2attP: the red arc arrows show the direction of segment transformation. (b) Schematic representation of deletions mapped in the region of the Fab-6 boundary. DNAse I hypersensitive sites (HS1 and HS2) are shown as gray boxes. Previously described deletions of the Fab-6 boundary8 are indicated by breaks in the gray lines. The endpoints of F61attP and F61+2attP deletions used in the replacement experiments are indicated by breaks in the black lines. The coordinates of endpoints are according to the complete sequence of BX-C in SEQ89E numbering61.
BX-C regulation is divided into two phases, initiation and maintenance. During the initiation phase, which takes place around the blastoderm stage, a combination of gap, pair-rule gene and maternally derived transcription factors establishes parasegment identity by interacting with special initiator elements in each regulatory domain3,5,6. This interaction sets the activity state, either on or off, of the regulatory domains3,7,8. The BX-C regulatory domains are activated sequentially along the anterior–posterior axis. For example, iab-5 is activated in PS10 and it directs Abd-B expression in this parasegment, while the remaining Abd-B regulatory domains iab-6 - iab-8,9 are off. In PS11, both iab-5 and iab-6 are activated; however, iab-6, which is closer to the Abd-B promoter, is responsible for regulating transcription. Again, the iab-7 and iab-8,9 are off.
Once the gap, pair-rule and maternal gene products disappear during gastrulation, regulation of BX-C switches to the maintenance phase. In this phase the on and off states of the regulatory domains are maintained by Trithorax (Trx) and Polycomb (PcG) group proteins, respectively3,9. Trx and PcG family proteins are highly conserved and regulate enhancer and promoter activity mainly by introducing or removing histone modifications10. Special elements, called PREs (Polycomb Response Element) are responsible for recruiting these maintenance factors11,12. Originally discovered in BX-C, PREs were subsequently found to control many other developmental genes10. PREs in flies map to large nucleosome free regions and have sites for a complex array of DNA binding protein11,12. Included in this group are recognition motifs for the GAGA transcription factor, GAF, and the one known PcG protein that binds DNA, Pleiohomeotic (Pho)13,14. These two proteins cooperate in the recruitment of other PcG factors15,16,17,18,19, which then mark the surrounding nucleosomes with the H3K27me3. In BX-C this histone mark helps establish and maintain the cis-regulatory domains in the off state20,21. When the cis-regulatory domains are in the on state, PcG proteins and the H3K27me3 mark are replaced by Trx proteins and marks of active chromatin22.
Critical to the autonomous activity of the BX-C regulatory domains are another class of elements, called boundaries or insulators23,24. Each of the BX-C regulatory domains is separated from the adjacent domains by boundaries. For example, iab-5 and iab-6 are separated from each other by Fab-6, while iab-6 is separated from iab-7 by the Fab-7 (Fig. 1). With the exception of Mcp, the boundaries in the Abd-B region (Fab-6, Fab-7 and Fab-8) have two critical activities25,26,27,28,29,30,31,32. The first is blocking crosstalk between adjacent domains. Boundary deletions fuse flanking regulatory domains, allowing adventitious interactions between initiators (and PREs) in each domain. Typically, the initiator in the more proximal domain activates the fused domain leading to a gain-of-function (GOF) transformation. When Fab-7 is deleted, for example, the iab-6 initiator activates iab-7 in PS11 where it would normally be off. This results in a transformation of PS11(A6) into PS12 (A7)28. Smaller Fab-7 deletions that remove most of the boundary but retain iab-7 PRE (HS3) give a mixed GOF and LOF (Loss-of-function) transformations33. In this case, the PRE is thought to silence some of the iab-6, iab-7 enhancers. The second function is boundary bypass. This activity is required when there are boundaries between the regulatory domain and its target promoter. As illustrated in Fig. 1a, there are two boundaries, Fab-7 and Fab-8, between iab-6 and Abd-B. In order to drive Abd-B expression in PS11, the tissue specific enhancers in iab-6 must bypass these two boundaries. Studies on Fab-7 and Fab-8 indicate that bypass activity, like insulation, is an intrinsic property25,26,34. However, not all BX-C boundaries have bypass activity, nor do they need this activity. For example, Mcp marks the border between the abd-A and Abd-B regulatory domains. In this location, bypass activity is not needed, and Mcp lacks this function26.
The most thoroughly studied BX-C boundary, Fab-7, consists of four hypersensitive regions, HS*, HS1, HS2 and HS335. Transgene and endogenous replacement experiments indicate that it is composed of multiple, partially redundant elements. For example, in transgene assays, fragments spanning HS* + HS1 + HS2, have enhancer blocking activity36,37. These same sequences are sufficient to confer nearly full boundary function in the context of BX-C: they block iab-6:iab-7 crosstalk and support bypass27,38,39. In transgene assays14,16 and also in genetic experiments33, Fab-7 HS3 functions as a PRE. However, recent experiments showed that in addition to PRE activity, HS3 also has boundary function27,39. In fact, a combination of the distal half of HS1, dHS1, plus HS3, has full function in Fab-7 boundary replacement experiments. Interestingly, a similar configuration of a boundary element and adjacent centromere distal PRE is observed for Mcp, Fab-6 and Fab-8, suggesting that this organization may have some functional significance. On the other hand, for Fab-8, the available evidence indicates that the nearby PREs is not important for full boundary function40.
Here we have used boundary replacement experiments to analyze the functional properties of the Fab-6 boundary. The chromosomal segment that is thought to contain Fab-6 has two DNase I hypersensitive regions, HS1 and HS2 (Fig. 1b). Unlike other boundaries, both hypersensitive regions can function as PREs and silence mini-white reporters in transgene assays41,42. In addition, sequences spanning HS1 region also function as a boundary, blocking the white enhancer from activating white41. Consistent with this observation, there are two binding sites for the chromosome architectural protein dCTCF in HS1 and dCTCF together with CP190 are associated with HS1 in vivo43,44. Moreover, as might be expected, blocking activity of HS1 is partially compromised in dCTCF mutants41. Several other lines of evidence argue that HS1 likely corresponds to Fab-6. First, Iampietro et al.8 found that like other BX-C boundaries, deletion of HS1 results in a mixed GOF/LOF transformation of A5 towards A6 or A4. Second, Kyrchanova et al.45 showed that a 425 bp sequence spanning Fab-6 HS1 functionally interacts with Fab-7, Fab-8 and the promoter region of Abd-B in a transgene pairing assay. Here we report that the core of Fab-6 boundary maps to a 529 bp sequence spanning HS1. While this core sequence can rescue a deletion spanning HS1, it cannot by itself rescue an HS1 + HS2 deletion, nor can it substitute for Fab-7. Instead HS1 must be combined with an HS2 fragment or the Fab-7 HS3. Our studies indicate that Fab-6 has two unusual features. One is the very large size of the sequences required for boundary function: two different DNA fragments of ~ 1 kb each are needed for boundary activity in the context of BX-C. The other is that both of these sequences not only have insulating activity but also function as PcG dependent silencers.
Results
The role of the Fab-6 HS1 in the functioning of the iab-5 and iab-6 regulatory domains
Iampietro et al.8 generated several deletions in the region between iab-5 and iab-6 that were expected to remove sequences critical for boundary function. One of these, Fab-62, removes ~ 8 kb including both HS1 and HS2. A second ~ 2 kb deletion, Fab-63, extends from a site in iab-5 ~ 1.1 kb from HS1 to a site within HS1 located just beyond the CTCF sites (Fig. 1b). Both deletions give a mixture of GOF/LOF phenotypes in A5 (PS10) and A6 (PS11). A third ~ 4,7 kb iab-6Δ7 deletion, that removes HS2, had no apparent phenotypic effects8. Based on these findings and those of Perez-Lluch et al.41, it was proposed that HS1 is the core Fab-6 boundary while HS2 corresponds to the iab-6 PRE.
To more precisely map the sequences required for boundary function, we used CRISPR/Cas9 to generate a 1389 bp deletion, F61attP, that excises all of HS1 plus several hundred bp of surrounding DNA (Fig. 1 and S1). The CRISPR/Cas9 construct carries a dsRed reporter (3 × P3-DsRed) to select for deletion events. Also included is an attP site that can be used for boundary replacement experiments46. In the adult cuticle the HS1 deletion, F61attP, displays a strong GOF transformation of segment A5 towards A6. This transformation is most clearly evident in the ventral sternite. Instead of the characteristic quadrilateral shape (wt: Fig. 1), the A5 sternite has a banana shape like that normally observed in A6. It also differs from wt in that it lacks bristles. On the other hand, there are patches of depigmented tissue in the A5 tergite, which is indicative of a LOF transformation towards A4. Interestingly, we also observed weak LOF transformations in A6. These include small depigmented patches in the A6 tergite, and an occasional bristle in the A6 sternite (F61attP, Fig. 2). The depigmentation seen in the tergite would be consistent with an A6 to A4 LOF transformation.

Mapping minimal sequences in HS1 Fab-6 required for boundary function. (a) Schematic diagram of the Fab-6 replacements. The coordinates of the Fab-6 fragments (F6′) are according to the BX-C sequence61. On the right side: scheme of the replacements after integration in F61attP platform. The mCherry and mini-y genes are indicated by magenta and yellow arrows respectively. (b) Morphology of the male abdominal segments (numbered) in F61attP, F61330, F6873 and F6529. The filled red arrowheads show morphological features indicative of GOF transformations. The empty red arrowheads show LOF transformations. Black arrowheads indicate pigmented spots that are the result of ectopic activation of the mini-y reporter. All other designations are the same as described in Fig. 1.
A similar, though not identical mixture of GOF/LOF cuticle phenotypes in A5 and a weak LOF phenotype in A6 were observed by Iampietro et al.8 in their Fab-63 deletion that removes part of iab-5 and most but not all of HS1 (Fig. 1a). One notable difference is that the GOF transformations of the sternite were much more modest in Fab-63 than in F61attP. Presumably these differences reflect the locations of the deletion breakpoints.
The Fab-6 HS1 cooperates with HS2 in blocking crosstalk between the iab-5 and iab-6 in vivo
To further define sequences important for Fab-6 function we generated three replacements. The largest was 1330 bp and contained nearly the entire sequence deleted in F61attP. The two other replacements were 873 bp and 529 bp (Fig. 2a). All three included the two CTCF sites. We introduced these replacements into F61attP using the φC31 attP/attB integration system46. To monitor blocking activity in the context of BX-C, the replacement included a minimal yellow reporter, mini-y (Fig. 2a). The reporter has a 340 bp yellow promoter linked to a yellow cDNA but lacks the wing, body and bristle enhancers of the endogenous yellow gene. As a result, mini-y expression depends upon enhancers in the neighborhood. The mini-y was placed relative to the test boundary sequences so that it is located in the iab-6 regulatory domain. We also included a second reporter, an mCherry gene, in the replacement construct, which was found to be useless due to high background expression.
In order to recover insertion events and also to monitor blocking activity, we used a y1 genetic background. In flies carrying the null y1 allele, the tan gene is still expressed under the control of Abd-B and A5 and A6 have a light brown-yellow instead of black pigmentation47 (Fig. 2b, wt y1). When mini-y is introduced into the F61attP deletion without a boundary, reporter expression is driven by enhancers in the fused iab-5:iab-6 domain. As can be seen by the dark pigmentation in the A5 and A6 tergites, the enhancers drive mini-y expression in both segments. However, as is observed when expression of the endogenous y gene is driven by Abd-B in the starting F61attP platform, there is “tan” pigmentation along the posterior margin of A5 as well as patches elsewhere in the tergite without pigmentation indicative of a LOF transformation of A5 into A4 (Fig. 2b). In these cells, the fused domain is shut down and mini-y (like Abd-B) is not expressed. A different pattern of mini-y expression is evident when the 1330 bp HS1 sequence is included in the replacement. In this case, mini-y is expressed at high levels throughout the A6 tergite, while expression is (with the exception of variable number of small darkly pigmented dots) absent in A5. Thus, the 1330 bp fragment effectively blocks crosstalk between iab-5 and iab-6. This is also true for the two smaller replacements, 873 bp and 529 bp. Both eliminate mini-y expression in A5 as effectively as the larger fragment. The blocking activity of these replacements is also evident in the morphological phenotypes of A5 in both y1 and y+ flies. Like wt the A5 tergite is covered in trichome hairs, while the sternite has a quadrilateral shape with many bristles. These features indicate that even the smallest DNA sequence effectively blocks crosstalk between iab-5 and iab-6. There was, however, one anomaly. In replacements carrying mini-y the A6 sternite has several bristles that are not seen in wt. However, this is likely due to promoter competition between mini-y and Abd-B for the iab-6 enhancers as it is not observed in the replacements after the mini-y is excised (Fig. 2b).
Fab-6 HS1 cannot substitute for Fab-7
In previous studies a deletion F7attP50 (Fig. 3a) that replaces the four Fab-7 nuclease hypersensitive regions with an attP site was generated27. We used this platform to further assess the functional properties of Fab-6 HS1 (Fig. 3a). For this purpose, we introduced the 1330 bp and 873 bp Fab-6 fragments described above, and a 685 bp fragment that has the same proximal end as the 529 bp fragment.

Fab-6 HS1 cannot substitute for Fab-7 unless it is combined with a PRE. (a) Maps of the Fab-6 and Fab-7 regions. The previously obtained Fab-7attP50 deletion27 is indicated by breaks in the black lines. (b) Morphology of the male abdominal segments in males with different Fab-6 replacements. All other designations are the same as described in Figs. 1 and 2.
The F7attP50 deletion results in a complete GOF transformation, and in males not only A7 but also A6 are absent. Surprisingly, all three Fab-6 HS1 fragments failed to rescue F7attP50 (Fig. 3b). In all three replacements, only a rudimentary A6 tergite is present, while there is no sternite. These findings indicate that Fab-6 HS1 is not sufficient to reconstitute a functional boundary in a different BX-C chromosomal context.
One plausible explanation is that the two deletions we have used to test Fab-6 HS1 boundary function are not equivalent. The Fab-7 deletion removes all of the nuclease hypersensitive regions including the HS3, which has both boundary and PRE activity. By contrast, the Fab-6 deletion only removes HS1 but not the HS2 PRE. If this explanation is correct, then it may be possible to reconstitute Fab-7 by combining DNA fragments that encompass Fab-6 HS1 and HS2.
To test this possibility, we generated two Fab-7 replacements. The first, F61330+3212, has HS1 plus a large fragment extending to either side of HS2. The second, F61330+930, has a smaller HS2 fragment. Figure 3 shows that for both of these replacements, the A6 segment is almost wild type, indicating that they are effective substitutes for Fab-7. In both replacements the tergite is fully formed and the trichome hairs are largely restricted to the anterior and lateral edges as in wt, while the sternite has the appropriate banana shape. However, for both replacements there are two anomalies: there are patches of ectopic trichomes on the A6 tergite, while the sternite has several bristles. These weak LOF defects would suggest that the boundary bypass activity of both F61330+3212 and F61330+930 is not fully effective. Alternatively, since both HS1 and HS2 have PRE activity, the two together could sometimes silence iab-6.
We also tested a 2264 bp fragment that includes the 1339 bp HS1, but extends in the opposite direction towards iab-5. Unlike F61330+3212 or F61330+930, F62264 failed to substitute for Fab-7, suggesting that there are no additional sequences conferring insulator function on the centromere proximal side of Fab-6 HS1.
Combination of Fab-6 HS1 and Fab-7 HS3 substitutes for Fab-7
The finding that Fab-6 HS1 substitutes for Fab-7 when combined with HS2 suggests that that HS2 has both PRE and boundary activity like Fab-7 HS339. If this idea is correct, then Fab-6 HS1 might be able to substitute for Fab-7 when linked to Fab-7 HS3. To explore this idea, we combined four different Fab-6 HS1 fragments (Fig. 3b and Fig. S2) with Fab-7 HS3. The largest was F61330, while the smallest was F642545. We also tested two intermediate Fab-6 fragments, F6744 and F61119 (Fig. 3).
The two larger combinations, F61330 + F7HS3 and F61119 + F7HS3, have similar activities. In both cases they rescue the GOF transformations of the F7attP50 deletion. The sternite has the appropriate banana shape, while the tergite is wt in size (Fig. 3b). However, as observed for F61330+3212, the sternite usually has a few bristles, while there are ectopic trichome patches on the tergite. Surprisingly, the two smaller combinations, F6744 + F7HS3 and F6425 + F7HS3, fail to fully rescue F7attP50. The GOF phenotypes are most pronounced in the F6425 + F7HS3 combination where both the sternites and tergites are significantly reduced in size. In the case of F6744 + F7HS3 the sternites are typically misshapen while the tegrites have nicks or are smaller than normal. As was the case for the larger F6 fragments, weak LOF phenotypes are also observed, likely due to minor defects in bypass activity.
Fab-6 HS1 + HS2 deletions have a more complete GOF phenotype
Our Fab-7 replacement experiments indicate that F61330 (HS1) must be combined either with the Fab-6 HS2 or Fab-7 HS3 to rescue the F7attP50 deletion. To further pursue the role of the Fab-6 HS2, we designed a CRISPR/Cas9 deletion, F61+2attP, which is 5995 bp and removes both HS1 and HS2.
As anticipated from studies on Fab-7, the phenotype of F61+2attP differs from F61attP. As shown in Fig. 1a, there is a nearly complete GOF transformation of A5 towards A6. The difference in the phenotypic effects of F61+2attP and F61attP are most clearly evident in the A5 tergite. While the A5 tergite in F61attP has patches of unpigmented cuticle indicative of an LOF transformation, the A5 tergite in F61+2attP is fully pigmented in > 90% of the males. Similarly, though the A5 sternite in F61attP has an A6-like banana shape, it also has several bristles, which are not present in the A6 sternite (Fig. S3). In F61+2attP the GOF transformation is more complete as the sternite lacks bristles in 70–80% of the males.
Thus, as was observed for Fab-7 deletions which retain or remove HS333, removing both Fab-6 HS1 and HS2 results in much more complete GOF transformation than HS1 only. In this context, we would note that a strong GOF transformation of A5 was not observed by Iampietro et al.8 in the Fab-62 deletion (Fig. 1b) which removes both Fab-6 HS1 and HS2. Instead, they observed a mixed GOF/LOF phenotype not altogether different from their smaller deletion, Fab-63. We suspect that the difference in phenotypes is that their deletion removes a larger segment from iab-5.
HS2 contributes to Fab-6 boundary activity
Next, we generated four F61+2attP replacements. The first replacement, F61330, has only HS1. The second, F61330+3212, has both HS1 and HS2, but lacks 730 bp between the HS1 and HS2. The third, F61330+930, has only HS1 and HS2. The fourth replacement was F62264 which contains the 1330 bp HS1 sequence, but extends into iab-5 (see above). These fragments were introduced into the mini-y replacement vector (Fig. 4).

The Fab-6 HS2 cooperates with HS1 to reconstitute a fully functional boundary. On the top: schematic diagram of the Fab-6 region, the F61+2attP deletion and the different DNA fragments tested for boundary activity. Morphology of the male abdominal segments in males carrying different Fab-6 fragments in F61+2attP. All other designations are the same as described in Figs. 1 and 2.
Figure 4 shows that the two replacements that contain only HS1, F61330 and F62264 are unable to insulate mini-y and it is expressed in A5 and A6. Consistent with ineffective insulation, the sternite in A5 has a banana shape while the arrangement of the trichomes in the A5 tergite resembles that normally observed in A6 (they are restricted to the anterior and dorsal lateral margins). Both of these morphological features are indicative of a GOF transformation of A5 towards A6 and are observed with and without the mini-y reporter. It is worth noting that the GOF transformations are less complete when the mini-y reporter is present. Though the A5 sternite has a banana shape, it has some bristles while there are small patches of trichomes on the tergite. However, the ectopic bristles and trichomes disappear and the flies resemble the starting deletion, F61+2attP after excision of mini-y.
A different result is obtained when the replacement combined Fab-6 HS1 plus HS2. The pigmentation pattern in the A5 and A6 tergites of y1; F61330+3212 mini-y and y1; F61330+930 mini-y indicates that the mini-y is (with the exception of a few small dots of pigmented cuticle in A5: see arrowhead) insulated from enhancers in iab-5. The morphology of the dorsal and ventral cuticle in A5 is also normal. The one possible exception is some gaps in the trichome field in the smaller F61330+930 replacement.
These findings suggest that iab-6 PRE, HS2, contributes to boundary function much like is observed for Fab-7 where HS3 functions both as the iab-7 PRE and as part of the Fab-7 boundary. If this suggestion is correct, then one would predict that Fab-7 HS3 would be able to substitute for Fab-6 HS2. To test this prediction, we combined Fab-6 HS1 with Fab-7 HS3 (F61330 + F7HS3). As shown in Fig. 4, the F61330 + F7HS3 combination also restores boundary function.
Discussion
Parasegment/segment identity depends upon the proper functioning of the different elements in each BX-C regulatory domains3. These elements include parasegment specific initiators that are determine the on/off state of domains early in development, elements that function to maintain the on/off state (PREs/TREs), and enhancers that drive expression of the homeotic genes in patterns appropriate for each parasegment. The regulatory domains are bracketed by chromatin boundaries, which are responsible for insulating regulatory elements within one domain from interactions with regulatory elements in neighboring domains. In addition to insulating activity, some of the BX-C boundaries can mediate interactions between enhancers in the regulatory domains and the relevant target promoter25,26,34. This boundary bypass function requires long distance physical interactions between boundaries and compatible architectural elements associated with the promoters of the three homeotic genes.
Here we have investigated the insulating and bypass activities of the Fab-6 boundary, which is located between iab-5 and iab-6. Though the sequence organization of Fab-6 is very different from that of the Fab-7, there are some parallels in their properties. Deletions that remove Fab-7 HS* + HS1 + HS2 but retain the HS3 iab-7 PRE result in a mixed GOF/LOF transformation of A633. A similar mixed GOF/LOF phenotype, in this case in A5 is observed in deletions that remove only Fab-6 HS1. For the Fab-6 HS1 deletion, the GOF phenotype results from activation of iab-6 in PS10 by the iab-5 initiator, while the LOF phenotype is due to the silencing of iab-5 and iab-6 in PS10 by a mechanism dependent on the iab-6 PRE. Fab-7 deletions that remove all HS sites result in a complete GOF transformation of A6 (PS11) into A7 (PS12)33. A similar strong GOF transformation is observed when both Fab-6 HS1 and HS2 are deleted.
Also, in both cases boundary function is supplemented by elements that have PRE activity. For Fab-7, the iab-7 PRE (HS3) contributes to its boundary function. Fab-6 also depends on elements that have boundary and PRE activity. In transgene studies Perez-Lluch et al.41 found that Fab-6 HS1 and HS2 silenced a white reporter by PcG dependent mechanism. In addition, HS1 also functions as a boundary in an enhancer blocking assays, and this activity depends upon the chromosomal architectural protein dCTCF. The conclusion that HS1 has boundary activity is supported by the studies of Iampietro et al.8 as well as our experiments showing that the F6529 fragment containing CTCF sites can rescue the GOF/LOF transformations of a F61attP deletion. On the other hand, even larger fragments extending to either side of the Fab-6 HS1 region are unable to rescue Fab-7attP50 and F61+2attP deletions. However, Fab-6 HS1 can substitute for Fab-7 when it is combined with HS2. This observation suggests that HS2 functions not only as a PRE silencer, but also as a boundary. Consistent with this idea, we found that the Fab-7 deletion, F7attP50, can also be almost fully rescued by combining Fab-6 HS1 and Fab-7 HS3. Rescuing of the large Fab-6 HS1 + HS2 deletion also requires a combination of Fab-6 HS1 with either HS2 or Fab-7 HS3. In this respect, it is interesting to note that genome-wide studies on Drosophila TADs (topologically associated domains) suggest that PREs are involved in the formation of chromatin loops48,49. It is not clear at this point to what extent this architectural function is due to the PcG silencing activity of these elements as opposed to a distinct boundary-like function.
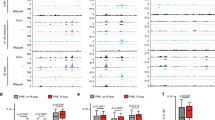
It is interesting to compare Fab-6 with the two other Abd-B boundaries, Fab-7 and Fab-8. Like Fab-8, there are two CTCF motifs in Fab-6 HS1 that bind dCTCF in ChIP experiments43,44,50. In spite of this similarity, Fab-6 HS1 is clearly not equivalent to Fab-8. Unlike Fab-6 HS1, a 337 bp Fab-8 fragment that includes the two CTCF sites can fully substitute for Fab-734,40. While dCTCF does not bind directly to Fab-7, ChIP-seq analysis51,52,53,54 indicate that several chromosomal architectural factors known to be important for Fab-7 boundary functions also appear to be associated with Fab-6 HS1. These common factors include the BEN DNA binding domain factors, Insensitive and Elba, Pita, and several proteins (GAF, Mod(mdg4) and CLAMP) that are thought to be components of the LBC (Late Boundary Complex) (Fig. 5). All of these factors also have recognition sequences in Fab-7 HS*, HS1 or HS225,27,55,56,57 and are known to play a role in their boundary functions. As for Fab-6 HS2, ChIP experiments indicate that the Polycomb DNA binding protein Pleiohomeotic (Pho), and the three LBC proteins GAF, Mod(mdg4) and CLAMP localize to this hypersensitive region. The same proteins are found associated with Fab-7 HS3. These similarities in protein composition would potentially explain why Fab-7 HS3 can substitute for Fab-6 HS2.

Comparison of chromatin protein binding to the Fab-6 and Fab-7 regions. The data are from a public functional genomics data repository GEO. The DNAse I hypersensitive sites (HS) of Fab-6 and Fab-7 are indicated as grey boxes. Binding sites for GAF, Clamp, INSV/ELBA complex, PHO, Zeste, Pita, dCTCF are indicated by short lines that correspond to the color of the protein names.
Our experiments highlight important differences in the properties of Drosophila boundary elements from those that have been reported for boundaries in mammals. All that is thought to be needed for full boundary function in mammals is the binding of a single protein, CTCF, to its 15 bp recognition sequence58,59. While Fab-6 has two dCTCF sites, these two sites are clearly not sufficient for boundary function in the context of BX-C. Instead, boundary activity requires two DNA sequences of ~ 1 kb in length and ChIP experiments indicate that at least eight different DNA binding proteins are associated with Fab-6 HS1 and HS2 in vivo (Fig. 5). Given that ChIPs have been done for only a small fraction of the predicted Drosophila DNA binding proteins60, it reasonable to think that many more proteins will interact with HS1 and HS2 and contribute to their boundary functions. Further analysis of the proteins associated with Fab-6 HS1 and HS2, and mutational studies will be required to confirm and extend these suggestions.
Methods
Generation of F6 1attP and F6 1+2attP deletions
The deletions were obtained by CRISPR/Cas9-induced homologous recombination. As a reporter, we used pHD-DsRed vector that was a gift from Kate O’Connor-Giles (Addgene plasmid # 51434). The plasmid was constructed in the following order: proximal arm-3 × P3:DsRed-distal arm. Arms for homology recombination were amplified by PCR from DNA isolated from Oregon line. For generation of the F61attP deletion, homology arms were obtained by DNA amplification between primers: F6ProxDRI: TATGAATTCCCCGAGACTAAACATAATTCGC; F6ProxRNde: TATCATATGACTGGCACCAGCTAATTGACAA; F6DistDSpe: TTACTAGTCATATTTGGGGATTTCTCTAAGTTTG; F6DistRPsc.
TTTACATGTCCGTGGTCGTTTTTTGTGGTT. For generation of the F61+2attP deletion only distal arm was changed: i6SGII: ATTAGATCTGCAAACTCAGTGGGCTTTTC; i6SXho: ATTCTCGAGCTGGTTGTTGGGATCGGG. The guide RNAs were selected using the program “CRISPR optimal target finder” (O’Connor-Giles Lab): for F61attP deletion - GTGCGCTAAGCACGCATATT and GTGTGTGGTCCGCAATACAG, for F61+2attP - AGTTTGCAAAGACAGTCCGT and GTGTGTGGTCCGCAATACAG. The breakpoints of the designed deletions: F61attP - 3R:16883201..16881813 and F61+2attP - 3R:16887807..16881813 16,869,768 according Genome Release r6.36.
To generate the desired deletions, the plasmid construct was injected into 58492 (Bloomington Drosophila Stock Center) embryos together with two gRNAs. The F0 progeny was crossed with y w; TM6/MKRS flies. Flies with potential deletions were selected on the basis of dsRed-signal in eyes and the posterior part of their abdomens and these flies were crossed with y w; TM6/MKRS flies. All independently obtained flies with dsRed reporter were tested by PCR. The successful deletion events were confirmed by sequencing of PCR products.
Generation of transgenic lines carrying different insertions in the Fab-7 attP50, F6 1attP and F6 1+2attP landing platforms
The Fab-7attP50 landing platform was described previously27. The test replacement fragments were inserted in a plasmid carrying the rosy reporter and an attP site described in27.
For the F61attP and F61+2attP landing platforms the replacement vector was a plasmid with the mini-yellow and mCherry reporter as shown in Fig. S1. The Fab-6 fragments were obtained by PCR amplification. Their coordinates are shown in Figures according to the published sequences of the Bithorax complex61.
Integration of the plasmids in the landing platforms was achieved by injecting the plasmid and a vector expressing the фC31 recombinase into embryos of yw; F61attP/ F61attP or yw; F61+2attP/F61+2attP line. The successful integrations were selected on the basis of expression of yellow reporter in abdominal segments. The integration of the replacement DNA fragments was confirmed by sequencing of PCR fragments.
The yellow and mCherry reporters were excised by Cre-mediated recombination between the lox sites.
All stocks are available upon request.
Cuticle preparations
Cuticle preparations were carried out as described by26.
References
Lewis, E. B. A gene complex controlling segmentation in Drosophila. Nature 276, 565–570 (1978).
Celniker, S. E., Keelan, D. J. & Lewis, E. B. The molecular genetics of the bithorax complex of Drosophila: characterization of the products of the Abdominal-B domain. Genes Dev. 3, 1424–1436 (1989).
Maeda, R. K. & Karch, F. The open for business model of the bithorax complex in Drosophila. Chromosoma 124, 293–307 (2015).
Kyrchanova, O. et al. The boundary paradox in the Bithorax complex. Mech. Dev. 138, 122–132 (2015).
Starr, M. O. et al. Molecular dissection of cis-regulatory modules at the Drosophila bithorax complex reveals critical transcription factor signature motifs. Dev. Biol. 359, 290–302 (2011).
Drewell, R. A. et al. Deciphering the combinatorial architecture of a Drosophila homeotic gene enhancer. Mech. Dev. 131, 68–77 (2014).
Casares, F. & Sánchez-Herrero, E. Regulation of the infraabdominal regions of the bithorax complex of Drosophila by gap genes. Development 121, 1855–1866 (1995).
Iampietro, C., Gummalla, M., Mutero, A., Karch, F. & Maeda, R. K. Initiator elements function to determine the activity state of BX-C enhancers. PLoS Genet. 6, e1001260 (2010).
Simon, J., Chiang, A. & Bender, W. T. different Polycomb group genes are required for spatial control of the abdA and AbdB homeotic products. Development 114, 493–505 (1992).
Schwartz, Y. B. & Pirrotta, V. A new world of Polycombs: unexpected partnerships and emerging functions. Nat. Rev. Genet. 14, 853–864 (2013).
Kassis, J. A., Kennison, J. A. & Tamkun, J. W. Polycomb and trithorax group genes in drosophila. Genetics 206, 1699–1725 (2017).
Kuroda, M. I., Kang, H., De, S. & Kassis, J. A. Dynamic competition of Polycomb and Trithorax in transcriptional programming. Annu. Rev. Biochem. 89, 235–253 (2020).
Brown, J. L., Mucci, D., Whiteley, M., Dirksen, M.-L. & Kassis, J. A. The Drosophila Polycomb group gene pleiohomeotic encodes a DNA binding protein with homology to the transcription factor YY1. Mol. Cell 1, 1057–1064 (1998).
Mishra, R. K. et al. The iab-7 polycomb response element maps to a nucleosome-free region of chromatin and requires both GAGA and pleiohomeotic for silencing activity. Mol. Cell. Biol. 21, 1311–1318 (2001).
Americo, J. et al. A complex array of DNA-binding proteins required for pairing-sensitive silencing by a polycomb group response element from the Drosophila engrailed gene. Genetics 160, 1561–1571 (2002).
Hagstrom, K., Muller, M. & Schedl, P. A Polycomb and GAGA dependent silencer adjoins the Fab-7 boundary in the Drosophila bithorax complex. Genetics 146, 1365–1380 (1997).
Kahn, T. G. et al. Interdependence of PRC1 and PRC2 for recruitment to Polycomb Response Elements. Nucleic Acids Res. 44, 10132–10149 (2016).
Brown, J. L. & Kassis, J. A. Architectural and functional diversity of Polycomb group response elements in Drosophila. Genetics 195, 407–419 (2013).
Ray, P. et al. Combgap contributes to recruitment of Polycomb group proteins in Drosophila. Proc. Natl. Acad. Sci. U. S. A. 113, 3826–3831 (2016).
Cheutin, T. & Cavalli, G. The multiscale effects of polycomb mechanisms on 3D chromatin folding. Crit. Rev. Biochem. Mol. Biol. 54, 399–417 (2019).
Bowman, S. K. et al. H3K27 modifications define segmental regulatory domains in the Drosophila bithorax complex. Elife 3, e02833 (2014).
Ciabrelli, F. et al. Stable Polycomb-dependent transgenerational inheritance of chromatin states in Drosophila. Nat. Genet. 49, 876–886 (2017).
Gyurkovics, H., Gausz, J., Kummer, J. & Karch, F. A new homeotic mutation in the Drosophila bithorax complex removes a boundary separating two domains of regulation. EMBO J. 9, 2579–2585 (1990).
Chetverina, D. et al. Boundaries of loop domains (insulators): determinants of chromosome form and function in multicellular eukaryotes. BioEssays 39, 1600233 (2017).
Kyrchanova, O. et al. Complete reconstitution of bypass and blocking functions in a minimal artificial Fab-7 insulator from Drosophila bithorax complex. Proc. Natl. Acad. Sci. 116, 13462–13467 (2019).
Postika, N. et al. Boundaries mediate long-distance interactions between enhancers and promoters in the Drosophila bithorax complex. PLOS Genet. 14, e1007702 (2018).
Wolle, D. et al. Functional requirements for Fab-7 boundary activity in the bithorax. Mol. Cell. Biol. https://doi.org/10.1128/MCB.00456-15.Address (2015).
Karch, F. et al. Mcp and Fab-7: molecular analysis of putative boundaries of cis-regulatory domains in the bithorax complex of Drosophilamelanogaster. Nucleic Acids Res. 22, 3138–3146 (1994).
Iampietro, C., Cleard, F., Gyurkovics, H., Maeda, R. K. & Karch, F. Boundary swapping in the Drosophila bithorax complex. Development 135, 3983–3987 (2008).
Iampietro, C., Gummalla, M., Mutero, A., Karch, F. & Maeda, R. K. Initiator elements function to determine the activity state of BX-C enhancers. PLoS Genet. https://doi.org/10.1371/journal.pgen.1001260 (2010).
Barges, S. et al. The Fab-8 boundary defines the distal limit of the bithorax complex iab-7 domain and insulates iab-7 from initiation elements and a PRE in the adjacent iab-8 domain. Development 127, 779–790 (2000).
Bender, W. & Lucas, M. The border between the Ultrabithorax and abdominal-A regulatory domains in the Drosophila bithorax complex. Genetics https://doi.org/10.1534/genetics.112.146340 (2013).
Mihaly, J., Hogga, I., Gausz, J., Gyurkovics, H. & Karch, F. In situ dissection of the Fab-7 region of the bithorax complex into a chromatin domain boundary and a Polycomb-response element. Development 124, 1809–1820 (1997).
Kyrchanova, O. et al. Distinct elements confer the blocking and bypass functions of the bithorax Fab-8 boundary. Genetics 213, 865–876 (2019).
Galloni, M., Gyurkovics, H., Schedl, P. & Karch, F. The bluetail transposon: evidence for independent cis-regulatory domains and domain boundaries in the bithorax complex. EMBO J. 12, 1087–1097 (1993).
Hagstrom, K., Muller, M. & Schedl, P. Fab-7 functions as a chromatin domain boundary to ensure proper segment specification by the Drosophila bithorax complex. Genes Dev. 10, 3202–3215 (1996).
Zhou, J., Barolo, S., Szymanski, P. & Levine, M. The Fab-7 element of the bithorax complex attenuates enhancer-promoter interactions in the Drosophila embryo. Genes Dev. 10, 3195–3201 (1996).
Mihaly, J. Dissecting the regulatory landscape of the Abd-B gene of the bithorax complex. Development 133, 2983–2993 (2006).
Kyrchanova, O. et al. The bithorax complex iab-7 Polycomb response element has a novel role in the functioning of the Fab-7 chromatin boundary. PLOS Genet. 14, e1007442 (2018).
Kyrchanova, O. et al. Functional Dissection of the Blocking and Bypass Activities of the Fab-8 Boundary in the Drosophila Bithorax Complex. PLoS Genet. 12, e1006188 (2016).
Perez-Lluch, S., Cuartero, S., Azorin, F. & Espinas, M. L. Characterization of new regulatory elements within the Drosophila bithorax complex. Nucleic Acids Res. 36, 6926–6933 (2008).
Ivlieva, T. A., Georgiev, P. G. & Kyrchanova, O. V. Study of the enhancer-blocking activities of new boundaries in the bithorax complex of Drosophilamelanogaster. Russ. J. Genet. 47, 1184–1189 (2011).
Smith, S. T. et al. Genome wide ChIP-chip analyses reveal important roles for CTCF in Drosophila genome organization. Dev. Biol. 328, 518–528 (2009).
Holohan, E. E. et al. CTCF genomic binding sites in Drosophila and the organisation of the bithorax complex. PLoS Genet. https://doi.org/10.1371/journal.pgen.0030112 (2007).
Kyrchanova, O. et al. Selective interactions of boundaries with upstream region of Abd-B promoter in Drosophila bithorax complex and role of dCTCF in this process. Nucleic Acids Res. 39, 3042–3052 (2011).
Bischof, J., Maeda, R. K., Hediger, M., Karch, F. & Basler, K. An optimized transgenesis system for Drosophila using germ-line-specific phiC31 integrases. Proc. Natl. Acad. Sci. U. S. A. 104, 3312–3317 (2007).
Camino, E. M. et al. The evolutionary origination and diversification of a dimorphic gene regulatory network through parallel innovations in cis and trans. PLoS Genet. 11, e1005136 (2015).
Eagen, K. P., Aiden, E. L. & Kornberg, R. D. Polycomb-mediated chromatin loops revealed by a subkilobase-resolution chromatin interaction map. Proc. Natl. Acad. Sci. 114, 8764–8769 (2017).
Ogiyama, Y., Schuettengruber, B., Papadopoulos, G. L., Chang, J.-M. & Cavalli, G. Polycomb-dependent chromatin looping contributes to gene silencing during drosophila development. Mol. Cell 71, 73-88.e5 (2018).
Bonchuk, A. et al. Functional role of dimerization and CP190 interacting domains of CTCF protein in Drosophilamelanogaster. BMC Biol. 13, 63 (2015).
Maksimenko, O. et al. Two new insulator proteins, Pita and ZIPIC, target CP190 to chromatin. Genome Res. 25, 89–99 (2015).
Ueberschär, M. et al. BEN-solo factors partition active chromatin to ensure proper gene activation in Drosophila. Nat. Commun. 10, 5700 (2019).
Rieder, L. E., Jordan, W. T. & Larschan, E. N. Targeting of the dosage-compensated male X-chromosome during early Drosophila development. Cell Rep. 29, 4268-4275.e2 (2019).
Negre, N. et al. A cis-regulatory map of the Drosophila genome. Nature 471, 527–531 (2011).
Kyrchanova, O. et al. Architectural protein Pita cooperates with dCTCF in organization of functional boundaries in Bithorax complex. Development 144, 2663–2672 (2017).
Kaye, E. G. et al. Drosophila dosage compensation loci associate with a boundary-forming insulator complex. Mol. Cell. Biol. 37, 1–18 (2017).
Fedotova, A. et al. Functional dissection of the developmentally restricted BEN domain chromatin boundary factor Insensitive. Epigenet. Chromatin 12, 2 (2019).
Hansen, A. S. CTCF as a boundary factor for cohesin-mediated loop extrusion: evidence for a multi-step mechanism. Nucleus 11, 132–148 (2020).
Arzate-Mejía, R. G., Recillas-Targa, F. & Corces, V. G. Developing in 3D: the role of CTCF in cell differentiation. Development 145, dev137729 (2018).
Kyrchanova, O. & Georgiev, P. Mechanisms of enhancer-promoter interactions in higher eukaryotes. Int. J. Mol. Sci. 22, 671 (2021).
Martin, C. H. et al. Complete sequence of the bithorax complex of Drosophila. Proc. Natl. Acad. Sci. U. S. A. 92, 8398–8402 (1995).
Acknowledgements
We thank Farhod Hasanov for fly injections. We thank Natalia Klimenko (the Center for Precision Genome Editing and Genetic Technologies for Biomedicine, IGB RAS) for help with bioinformatics.
Funding
This work (all functional and morphological analysis) was supported by the Russian Science Foundation, Project No. 19-14-00103 (to O.K.). Part of this work (genome editing procedure and bioinformatics) was supported by Grant 075-15-2019-1661 from the Ministry of Science and Higher Education of the Russian Federation. PS would like to acknowledge support from NIH R35 GM126975. Funding for open access charge: Russian Science Foundation.
Author information
Authors and Affiliations
Contributions
P.G., O.K. designed experiments. O.K., N.P. performed experiments. P.S., P.G., O.K. wrote the main manuscript text. O.K. prepared figures. All authors reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Postika, N., Schedl, P., Georgiev, P. et al. Mapping of functional elements of the Fab-6 boundary involved in the regulation of the Abd-B hox gene in Drosophila melanogaster. Sci Rep 11, 4156 (2021). https://doi.org/10.1038/s41598-021-83734-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-83734-8
This article is cited by
-
CRISPR/Cas9 and FLP-FRT mediated regulatory dissection of the BX-C of Drosophila melanogaster
Chromosome Research (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
