
Abstract
Ethical responsibility, legal requirements and the need to improve the quality of research create a growing interest in the welfare of laboratory animals. Judging the welfare of animals requires readout parameters, which are valid and sensitive as well as specific to assess distress after different interventions. In the present study, we evaluated the sensitivity and specificity of different non-invasive parameters (body weight change, faecal corticosterone metabolites concentration, burrowing and nesting activity) by receiver operating characteristic curves and judged the merit of a multi-parametric analysis by logistic regression. Chronic pancreatitis as well as laparotomy caused significant changes in all parameters. However, the accuracy of these parameters was different between the two animal models. In both animal models, the multi-parametric analysis relying on all the readout parameters had the highest accuracy when predicting distress. This multi-parametric analysis revealed that C57BL/6 mice during the course of chronic pancreatitis often experienced less distress than mice after laparotomy. Interestingly these data also suggest that distress does not steadily increase during chronic pancreatitis. In conclusion, combining these non-invasive methods for severity assessment represents a reliable approach to evaluate animal distress in models such as chronic pancreatitis.
Similar content being viewed by others

Introduction
Laboratory animals represent a valuable tool for mimicking physiological conditions in humans and are, therefore, still widely used for biomedical research in the pursuit of understanding the pathophysiology of a disease and exploring novel treatments1,2. On the other hand, improving animal welfare is important in order to alleviate the suffering of animals. Consequently, the 3Rs (replacement, reduction and refinement of animal experiments) have been coined by Russell and Burch3 more than 50 years ago and are now the basis of national and international regulations of animal experiments. In accordance with this concept, the European Union Directive 2010/63/EU and many local institutional animal care and use committees in the USA demand to provide adequate information to assess the potential distress and pain that animals might experience during an experiment4,5. This represents an essential component of the harm-benefit analysis when evaluating any project that involves animal experiments4. Distress is often defined as an aversive state in which the animal is unable to adapt completely to stressors and the resulting stress and shows maladaptive behaviour6,7. Indeed, pain can be seen as just one possible cause of distress, thus suggesting that quantifying distress of animals might be of greater importance for animal welfare than focusing just on the quantification of pain8. However, it has been suggested that distress is not always a consequence of pain9; e.g. it might be possible that short time pain does not cause maladaptive behaviour. In this study, we follow the concept of Koch et al. that one should focus on distress (both pain-related and non-pain-related) when evaluating animal welfare8.
The European Union Directive 2010/63/EU provides examples of severity grading for some interventions. For instance, the directive suggests that laparotomy causes moderate distress. Thus, such a model was used in this study as a reference model for evaluating the degree of distress experienced by mice during chronic pancreatitis (CP). Importantly, only few publications provide a profound distress analysis of a specific animal model10,11,12, and there is a crucial need for more objective severity grading of procedures13. Additionally, in order to judge the severity of animal models, it is important, first, to validate methods used for distress analysis and, second, to combine these methods to pursue a multi-parametric analysis of distress. Finally, the distress caused by each animal model needs to be compared to a reference.
Animal distress associated can be assessed by various distress indicators. For example, burrowing or nesting activity may reflect the well-being of mice14,15. In addition, body weight is used as an indicator of well-being, since animals in pain can have reduced food and water intake16. Moreover, many research groups have described that the concentration of faecal corticosterone metabolites (FCM)17,18,19 and plasma corticosterone concentration20 may indicate animal distress. Thus, many publications describe multiple methods to score distress14,15,17,18,19. However, little is known about which readout parameters are best suited to diagnose distress. For judging the diagnostic ability of tests and readout parameters, receiver operating characteristic (ROC) curves are widely used in clinical21,22,23 and in experimental settings24 Moreover, ROC curves have also been widely used to define the accuracy of psychological distress scores for cancer patients25,26.
The pathophysiology of CP remains poorly understood and is still inadequately treated in humans27. Moreover, CP inflicts abdominal pain on patients, which affects daily activities as the disease progresses28,29. In addition to pain, CP can also lead to endocrine and exocrine pancreatic insufficiency with severe complications29. These facts altogether create a crucial need for extensive scientific research in the field of CP in order to explore pathophysiological mechanisms and novel therapeutic approaches. Interestingly, while some animal models for CP do not reproduce all histopathological features of this disease and some animal models have a relatively high mortality rate, repetitive i.p cerulein injections very reliably cause CP30,31.
In humans, pain of CP is managed primarily by paracetamol, nonsteroidal anti-inflammatory drugs or opioid analgesics such as tramadol32. Nevertheless, pain is still often insufficiently controlled33. Recently, microRNAs have been implicated in modulating pain perception34. For example, microRNA-21 has been described to contribute to neuropathic pain in mice35,36. Since neuropathic pain is considered to be an inherent component of pain in CP37,38, inhibition of microRNA-21 might support analgesic regimens and mitigate animal distress associated with CP.
It was the aim of this project to evaluate physical, behavioural and physiological animal distress parameters during CP and to compare their ability to predict distress. We also aimed at exploring the benefit of multi-parametric analysis and compared the distress animals experience during CP to distress after laparotomy.
Results
Pathological features and distress caused by CP
Plasma lipase activity, indicating damage to the pancreas, was significantly elevated during the early and middle phase of CP (Fig. 1A). However, it was reduced during the middle and late phase when compared to the early phase of CP (Fig. 1A). The pancreas to body weight ratio, as a measurement of tissue atrophy, was significantly reduced in cerulein-injected mice when compared to healthy mice (Fig. 1B). Another feature of CP, fibrosis, was also observed in cerulein-injected animals (Fig. 1C,D). The quantification of collagen I deposition in pancreatic tissue confirmed increased fibrosis in mice with CP (Fig. 1E). Thus, repetitive cerulein injection induced key characteristics of CP in mice.

Evaluation of chronic pancreatitis. (A) Serum lipase concentration before (pre) and during the early, middle and late phase of cerulein-induced CP; One Way Repeated Measures ANOVA followed by pairwise comparison with Holm-Sidak-Method (*P ≤ 0.041), n = 4. (B) Pancreas to body weight ratio of healthy mice (−CP) and mice at the late phases of CP (+CP); Student’s t-test (*P ≤ 0.001), healthy: n = 12, CP: n = 15. Immunohistochemical detection of collagen I accumulation in (C) healthy mice and (D) mice at the late phase of CP; bar =100 µm. (E) Quantification of collagen I deposition in healthy mice (−CP) and at the late phase of CP (+CP); unpaired t-test with Welch’s correction (*P < 0.0001), healthy: n = 8, CP: n = 5.
We observed a significant reduction of body weight in all phases after induction of CP when compared to the pre-phase, which is the phase before any intervention (Fig. 2A). We also observed a reduction in body weight between the early phase and the subsequent phases, but not between the middle and the late phase (Fig. 2A). Burrowing activity was significantly reduced throughout the course of CP with the deepest reduction in the early phase, thereafter, a slight recovery was observed (Fig. 2B). Nesting activity was significantly reduced from pre-phase to all post induction phases (Fig. 2C). Moreover, the nesting scores were significantly reduced between the early phase and the middle phase, but not between the middle and the late phase. FCM concentration significantly increased in the early phase after induction, followed by a significant decrease in the middle phase (Fig. 2D). We observed no significant differences in any of these readout parameters between animals treated with microRNA-21 inhibitor and animals treated with microRNA-21 control oligonucleotides (Fig. 2A–D). When comparing these distress parameters before and collectively after induction of CP, we noticed a significant reduction in percent body weight change (Fig. 2E), burrowing activity (Fig. 2F) and nesting activity (Fig. 2G) as well as a significant increase in FCM concentration (Fig. 2H). In order to check the validity of the FCM data, we analysed blood corticosterone concentration at different phases of CP. Corticosterone concentration in the blood also significantly increased in the early phase after induction, followed by a significant decrease in the middle phase (Supplementary Fig. S1). However, this hormone remained significantly elevated at all time points after intervention when compared to the pre-phase (Supplementary Fig. S1). There was also a significant increase when collectively comparing blood corticosterone concentration before and after induction of CP (Supplementary Fig. S1). Thus, these data demonstrate that all readout parameters are sensitive enough to score increased distress after induction of CP.

Course of distress during chronic pancreatitis. (A–D) Determination of distress parameters in control and microRNA-21 inhibitor treated mice before (pre) and during the early, middle and late phases of CP; control: n = 8, microRNA-21 inhibitor: n = 8. (A) Percentage of body weight change; Two Way Repeated Measures ANOVA, followed by pairwise comparison with Holm-Sidak-Method (*P ≤ 0.047). (B) Burrowing activity within 2 hours; Two Way Repeated Measures ANOVA, followed by pairwise comparison with Holm-Sidak-Method (*P ≤ 0.002). (C) Nesting activity Two Way Repeated Measures ANOVA, followed by pairwise comparison with Holm-Sidak-Method (*P ≤ 0.021). (D) Concentration of faecal corticosterone metabolites (FCM); Two Way Repeated Measures ANOVA, followed by pairwise comparison with Holm-Sidak-Method (*P ≤ 0.025). (E–H) Comparing distress before (−CP, n = 16) and during the early, middle and late phases of cerulein treatment (+CP, n = 48). (E) Percentage of body weight change; Mann-Whitney Rank Sum Test (*P ≤ 0.001). (F) Evaluation of burrowing activity; Mann-Whitney Rank Sum Test (*P ≤ 0.001). (G) Evaluation of nesting activity; Mann-Whitney Rank Sum Test (*P ≤ 0.001). (H) Assessment of faecal corticosterone metabolites concentration; Mann-Whitney Rank Sum Test (*P = 0.016).
The accuracy of parameters defining distress after CP
The single readout parameters, percentage of body weight change (Fig. 3A), burrowing activity (Fig. 3B), nesting activity (Fig. 3C) and FCM (Fig. 3D) had discriminatory power to differentiate between animals before and after induction of CP. Moreover, the combination of all four parameters also had discriminatory power to differentiate between animals before and after induction of CP (Fig. 3E). The combination of all four parameters produced the largest area under the curve (AUC = 0.98, 0.95–1.00 confidence interval), suggesting that the combination of all readout parameters allows to differentiate between distress assessed before and after induction of CP with higher accuracy than any of the single readout parameters (Fig. 3A–E). When applying Youden’s index to define the best cut off to differentiate between distress of animals before and after induction of CP, the combination of all readout parameters allows differentiating between animals before and after induction of CP with higher accuracy (96.9%, 89.2–99.6% confidence interval) than any of the single readout parameters (Supplementary Table S1).

Receiver operating characteristic curve analysis of distress parameters in the chronic pancreatitis animal model. Evaluation of (A) percentage of body weight change, (B) burrowing activity, (C) nesting activity, (D) concentration of faecal corticosterone metabolites (FCM) and (E) combination of these four parameters after applying binary logistic regression in a multiple model. The area under the curve (AUC) with the respecting confidence interval (CI) and the P value (P) for testing the AUC to be 0.5, are indicated. Before induction of CP: n = 16, after induction of CP: n = 48.
Comparing distress after laparotomy to distress during CP
After laparotomy the body weight was significantly decreased (Fig. 4A). In addition, the amount of pellets burrowed significantly plummeted after the surgical intervention (Fig. 4B) and the nesting score was significantly reduced after laparotomy (Fig. 4C), while FCM concentration significantly increased (Fig. 4D). These data were separated into two data sets by randomisation (Fig. 5). ROC analysis was performed on data set 1 in order to compare the accuracy of these parameters in defining distress (Fig. 6A–D). The single readout parameters, percentage of body weight change and nesting activity (Fig. 6A,C), had only little discriminatory power to differentiate between animals before and after laparotomy (body weight: 0.63, 0.40–0.86 confidence interval, P = 0.2592; nesting: 0.67, 0.45–0.88 confidence interval, P = 0.1439). Burrowing activity (Fig. 6B) and FCM (Fig. 6D) had a high discriminatory power to differentiate between animals before and after laparotomy. The combination of all four parameters (Fig. 6E) produced the largest area under the curve (AUC = 0.99, 0.96–1.00 confidence interval, P < 0.0001), suggesting that the combination of all readout parameters allows differentiating between animals before and after laparotomy with higher accuracy than any of the single readout parameters (Fig. 6A–E). When determining the best cut off to this multi-parametric analysis, the combination of all readout parameters allows to differentiate between animals before and after laparotomy with higher accuracy (96.2%, 80.4–99.9% confidence interval) than any of the single readout parameters (Supplementary Table S2). In order to check the validity of this cut off, we evaluated if this cut off can differentiate between animals before and after laparotomy using a different data set (data set 2). This cut off had 100% specificity, 100% sensitivity and 100% accuracy when applied to data set 2. When applying this cut off to data taken after laparotomy (data set 2), all data collected after laparotomy were allocated to distress level 2 (Table 1). When applying this cut off to data taken after induction of CP, some data were allocated to distress level 2, whereas some data were allocated to distress level 1, which indicates baseline distress of healthy animals (Table 1). Using Fisher’s exact test, a significant difference in the distress levels distribution between laparotomy and CP was observed. This suggests that during CP two different distress levels can be assessed: A distress level similar to distress observed after laparotomy (distress level 2) and a distress level that compares to baseline distress (distress level 1).

Distress before and after laparotomy. Quantification of distress in mice before (−laparotomy) and after laparotomy (+laparotomy). (A) Percentage of body weight change; Paired t-test (*P ≤ 0.001), n = 25. (B) Burrowing activity within 2 hours; Wilcoxon signed-rank test (*P ≤ 0.001), n = 25. (C) Nesting activity; Wilcoxon signed-rank test (*P = 0.002), n = 25. (D) Concentration of faecal corticosterone metabolites (FCM); Paired t-test (*P ≤ 0.001), n = 25.

Procedure of distress evaluation. (1.) Data collected from 25 mice (intervention: laparotomy) were separated into two data sets by randomisation. (2.) Data of data set 1 were used to define the cut off, which separates distress observed before (−) from distress observed after intervention (+). (3.) This cut off was validated using data set 2. (4.) This cut off was also used to compare distress after laparotomy to distress during CP.

Receiver operating characteristic curve analysis of distress parameters (laparotomy data set 1). (A) Percentage of body weight change, (B) burrowing activity, (C) nesting activity, (D) faecal corticosterone metabolites concentration and (F) combination of these four parameters after applying binary logistic regression in a multiple model. The area under the curve (AUC) with the respecting confidence interval (CI) and the P value (P) for testing AUC to be 0.5, are indicated. Before induction of laparotomy: n = 13, after induction of laparotomy: n = 13.
Discussion
This study demonstrates that all distress readout parameters (FCM, body weight change and burrowing as well as nesting activity) are significantly changed by both CP and laparotomy. This suggests their broad applicability to assess distress after completely distinct interventions. However, the accuracy of these parameters when distinguishing between healthy mice and mice after an intervention was different between CP and laparotomy. In the animal model of CP body weight change was a more accurate parameter than FCM (accuracy of parameters: body weight > burrowing > nesting > FCM), whereas after laparotomy FCM was the most accurate parameter to measure distress (accuracy of parameters: FCM > burrowing > nesting > body weight). In addition, this study also demonstrates that in both animal models the multi-parametric analysis relying on all four readout parameters had the highest accuracy when predicting distress. Interestingly, these data also suggest that distress does not steadily increase during CP.
Body weight and burrowing as well as nesting activity have been often evaluated when assessing distress of animals18,19,39. Reduction of body weight was, for example, reported as a good indicator of distress in acute colitis and after general anaesthesia10,11,40. Body weight reduction was also described after induction of CP with cerulein in mice41. Moreover, reduced burrowing or nesting activity were observed after laparotomy42, induction of hippocampal lesion15, isoflurane anaesthesia18, or intra-bone marrow transplantation43. Analysis of FCM is also a well-established non-invasive method to evaluate distress and incurs less stress on mice than the measurement of corticosterone in blood17,39,44. It is less often used in studies than body weight and assessment of animal behaviour, but increased FCM concentration was reported after ketamine and xylazine anaesthesia11 as well as after intra-bone marrow transplantation43. In agreement with the above cited literature FCM, body weight change, burrowing and nesting activity are significantly changed by CP as well as laparotomy (Figs. 2 and 4). This suggests that these readout parameters are sensitive enough to assess distress after completely distinct interventions, i.e. repetitive cerulein injections and laparotomy. Moreover, we also tested the effect of microRNA-21 inhibitor on these distress parameters. MicroRNA-21 inhibition was described as an effective strategy in many therapeutic areas45,46,47. Thus, we hypothesized that microRNA-21 inhibitor can mitigate animal distress that might be caused by neuropathic pain component of CP37,38. This assumption was based on the previously reported contribution of microRNA-21 to neuropathic pain35,36. However, application of microRNA-21 inhibitor did not have any significant effect on any of the four distress readout parameters during CP. These negative results might be attributed, at least partly, to the multifactorial nature of pain in CP33,37.
Although distress parameters such as FCM, body weight, burrowing as well as nesting activity are often used to measure distress10,11,18,43, little is known about the sensitivity, specificity and accuracy of these readout parameters. According to ROC analysis, we found that body weight and burrowing are able to predict distress in CP with a higher accuracy than nesting and FCM. On the other hand, FCM and burrowing were the most accurate single parameters in predicting distress in laparotomy. Thus, although all four readout parameters are broadly applicable to assess distress, the accuracy of these parameters when distinguishing between healthy mice and mice after an intervention was different between CP and laparotomy. This argues for using more than one parameter for evaluating distress of animals. This is especially necessary, when comparing distress caused by distinct interventions. Indeed, our multi-parametric analysis using logistic regression followed by ROC analysis allowed us to estimate the accuracy of multi-parametric analysis using all four parameters. So far, only a few other studies reported multi-parametric analysis with methods such as cluster analysis10.
Animal models with long lasting distress such as CP are important for basic or preclinical research on many chronic diseases48,49,50. However, according to the European Union Directive 2010/63/EU, long lasting distress for animals automatically increases the severity of an animal experiment4. For example, long-lasting moderate distress has to be upgraded to severe distress4. Indeed, we know that long lasting diseases can lead to persistent or even increased distress and pain. This is, for example, true for osteoarthritis or neuropathic pain51,52. However, we also know that long lasting stress can lead to habituation or desensitization. One well accepted example is the hypothalamic–pituitary–adrenal axis, where repetitive stress often leads to decreased release of stress hormones such as corticosterone53,54. Interestingly, we found that percentage of body weight change, burrowing and nesting activity remained steady between the middle and late phase of CP (Fig. 2A–C). Moreover, FCM concentration was even significantly reduced in the middle phase when compared to the early phase of CP (Fig. 2D). A reduction of the stress hormone corticosterone was also observed in the blood when comparing middle and late phase of disease progression to the early phase of CP (Supplementary Fig. S1). These data suggest that redundant cerulein injections, especially during the late stage of CP, do not lead to hyperalgesia. This is consistent with several publications describing habituation in mice after exposure to repetitive stress55,56.
Surprisingly, only few studies assessed distress caused by preclinical in vivo research11,18,43. Thus, there is an urgent need for detailed characterization of distress in most animal models. In order to judge distress caused by cerulein-induced CP, we used distress caused by laparotomy as a reference, since the European Union Directive 2010/63/EU suggests that the distress level after laparotomy is moderate. When pursuing a multi-parametric analysis using FCM, body weight and burrowing as well as nesting activity of data set 1 before and after laparotomy, a best cut off could be defined that differentiates between ‘baseline distress’ (data taken before laparotomy) and ‘up to moderate distress’ (data taken after laparotomy). Subsequently, this cut off was applied on two different post intervention data sets; laparotomy data set 2 and the CP data set. This comparison demonstrates that after CP, 60% (29 from 48) of all data points can be defined as distress level 2 (up to moderate distress), whereas 40% (19 from 48) of all data points can be defined as distress level 1 (baseline distress). This suggests that at any given time point during CP mice experience sometimes baseline distress and sometimes up to moderate distress. Thus, when just considering one time point, mice often experience less distress during CP than after laparotomy. The following points support such a conclusion. First, none of the mice in both models lost more than 20% of original body weight, a margin which is commonly used to indicate more than moderate, thus severe distress16. Second, none of the mice died or had to be euthanized. Consequently, we propose that distress at a certain time point during CP could be defined to be mild to moderate.
However, the European Union Directive 2010/63/EU demands that the continuity of distress has to be taken into account when judging the severity level of an animal model. For example, long term mild distress should be upgraded to an overall moderate distress. Likewise, long term moderate distress should be upgraded to severe distress. Considering these recommendations, the severity level of CP could be defined to be moderate to severe. However, such a classification seems to violate proportionality when one compares CP to animal models and interventions that are suggested to be severe by the European Union Directive 2010/63/EU. For example, this directive suggests to classify toxicity testing where death is the end-point and irradiation with a lethal dose without reconstitution of the immune system as severe4. In these cases a very high percentage of animals would suffer until death or euthanasia. In contrast to these scenarios, CP only induced a maximal loss in body weight of 13% at any time point. Moreover, no mouse died or had to be euthanized during CP. This suggests that we need more than four classes (non-recovery, mild, moderate and severe) of defining severity of animal experiments, in order to generate a fair system that guarantees proportionality in the severity scoring of distinct animal models.
In conclusion, this study suggests that ROC analysis is a feasible tool to compare sensitivity, specificity and accuracy of distinct methods to analyse animal distress. Moreover, multi-parametric analysis of distress with these four parameters has the potential to evaluate distress with a high accuracy and allows a comparison between distress caused by completely different interventions such as induction of CP and laparotomy. This multi-parametric analysis revealed that mice during the course of CP often experienced less distress than mice after laparotomy.
Material and Methods
Ethical statement
This study was conducted in accordance with the German law for animal protection (TierSchG) and the European Union Directive, 2010/63/EU. The public authority (Landesamt für Landwirtschaft, Lebensmittelsicherheit und Fischerei Mecklenburg-Vorpommern, 7221.3–1–002/17, 7221.3–1–019/15) and the §15 (according to the TierSchG) committee of Mecklenburg-Vorpommern approved all experiments and experimental protocols.
Animals
C57Bl/6J mice were housed in type III cages at 12 h dark light cycle (light period: 7:00–19:00) and a temperature of 21 ± 2 °C, with a relative humidity of 60 ± 20 %. Food (pellets, V1534.000, 10 mm, ssniff Spezialdiaeten GmbH, Soest, Germany) and water were provided ad libitum. Autoclaved bedding (Bedding Espe Max 3–5mm granulate, H 0234–500, Abedd, Vienna, Austria) was used. Enrichment was provided by nesting material (Zoonlab GmbH, Castrop-Rauxel, Germany), a fun tunnel (a 75 × 38 mm paper tunnel, H 0528–151, ssniff Spezialdiaeten GmbH,) and a wooden block (Espe size S, 40 × 16 × 10 mm), H0234.NSG, Abedd).
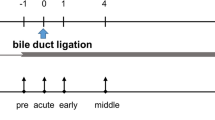
CP was induced by repetitively injecting male 9–15 weeks old C57Bl/6J mice with cerulein. Cerulein (Bachem H-3220.0005, Bubendorf, Switzerland) dissolved in 0.9% sodium chloride was administered by consecutive intraperitoneal (i.p.) injections (50 μg/kg, three hourly injections/day; three days/week (on day 0, 2, 4, 7, 9, 11, 14, 16, 18, 21, 23, 25, 28 and 30). MicroRNA-21 inhibitor (miRCURY LNA™ microRNA-21a-5p inhibitor; cat. #, 339203 YCO0070656, sequence: TCAGTCTGATAAGCT) and its corresponding microRNA-21 control (miRCURY LNA™ microRNA-21a-5p control; cat. #, 339203 YCO0070657, sequence: TCAGTATTAGCAGCT) were purchased from Qiagen (Hilden, Germany). They were resuspended in PBS and injected at a dose of 10mg/kg (s.c. to ensure sustained release of the drug) on day 0 and day 14. For pain relief 1250 mg/l metamizol (Ratiopharm, Ulm, Germany) was provided daily (starting at day −1 until day 33) in the drinking water. The mice were euthanized under anaesthesia by cervical dislocation. The pancreas to body weight ratio of each mouse was recorded on day 33 and the tissue was taken for further analysis. Animals (16 mice split into two separate experiments) were allocated in a non-random manner either to a control or a treatment group by matching their burrowing activity during the pre-phase. Both microRNA-21 control and inhibitor were injected in a non-blinded manner.
In order to execute laparotomy, male C57Bl/6J mice were anaesthetized by 1.2–2.5% isoflurane, which provides general anaesthesia as well as an analgesic effect and reduction of stress. The eyes were protected by applying eye ointment. Within one to two minutes carprofen (Pfizer GmbH, Berlin, Germany) at a dosage of 5 mg/kg was injected (s.c.) before surgery. After shaving and disinfection of the skin, the abdominal cavity was opened by transverse incision. Then, 5 μl of the cell suspension (murine cell line 6606PDA, 2.5 × 105 cells in 5 µl matrigel) were injected slowly with a 25 μl syringe into the pancreas (Hamilton Syringe, Reno, Nev., USA). Thereafter, the cavity and the skin were closed by a coated 5–0 vicryl suture (Johnson & Johnson MEDICAL GmbH, Norderstedt, Germany) and a 5–0 prolene suture (Johnson & Johnson MEDICAL GmbH). The purpose of this cell injection was to generate an orthotopic syngeneic pancreatic cancer model to explore drugs for the treatment of cancer (see e.g. Zechner et al.57 and Zhang et al.58; the FCM data for this model have been created for the manuscript of Kumstel et al. 2019, submitted to Scientific Reports). The data of the laparotomy animal model were randomized to define data set 1 and data set 2 using the random number generator at the official website www.stattrek.com.
Analysis of animal distress
In the laparotomy model, the percentage of body weight change, burrowing activity, nesting activity, FCM concentrations were measured at two time points: before and after surgical intervention. In the CP model, distress readout parameters were measured, before induction of CP (pre-phase) and throughout the different phases of disease progression; early, middle and late phase. Before each experiment, burrowing and nesting assays were performed two times with mice housed in groups of four or six animals to facilitate that the mice learn this behaviour from each other. Thereafter, all the animals were housed individually and burrowing and nesting activity were assessed three times. Mice which burrowed three times less than 10g or which had an average nesting score of less than 2.5 during the pre-phase were excluded from further analysis.
The body weight was measured two times before and one day after laparotomy as well as before induction of CP (pre-phase) and throughout the different phases of disease progression; early (day 3), middle (day 17) and late (day 31). Thus, in all experiments the body weight was determined 24 hours after induction of distress.
The burrowing activity was measured before and on the day of laparotomy as well as before induction of CP (pre-phase) and throughout the different phases of disease progression; early (day 2), middle (day 16) and late (day 30). In order to quantify burrowing activity, a burrowing tube (15 cm length × 6.5 cm diameter) was filled with 200 ± 1 g food pellets (ssniff Spezialdiaeten GmbH) and was put in the cage two and a half to three hours before the dark phase. After two hours, the weight of the food pellets [g] left in the tube was measured and deducted from 200g.
Nesting activity was measured before and on the day of laparotomy as well as before induction of CP (pre-phase) and throughout the different phases of disease progression; early (day 2), middle (day 16) and late (day 30). In order to analyse nest building behaviour, a cotton nestlet (5 cm square of pressed cotton batting, Zoonlab GmbH, Castrop-Rauxel, Germany) was placed in the left back corner of the cage 30 to 60 minutes before the dark phase. The nests were scored at the end of the dark phase ±2 hour, by using a scoring system developed by Deacon15. However, a 6th score point was added to this scoring system. The score 6 defines a perfect nest: The nest looks like a crater and more than 90% of the circumference of the nest wall is higher than the body height of the coiled up mouse.
FCM was assessed in faeces (which was left in the cages within 24 hours) before and on the day of laparotomy as well as before induction of CP (pre-phase) and throughout the different phases of disease progression; early (faeces collected from day 2 to day 3), middle (day 16–17) and late (day 30–31). In the laparotomy model, faeces was collected before and during 24 hours immediately after the surgical intervention. The faeces (200–400 mg) was dried for 4 hours at 65 °C and kept at −20 °C until further processing. Thereafter, 50 mg of dried faeces were extracted with 1 ml 80% methanol for further analysis by 5α-pregnane-3β,11β,21-triol-20-one enzyme immune assay17,59.
Corticosterone concentration was evaluated exclusively in the blood plasma of animals other than those subjected to distress analysis by non-invasive methods (four additional animals). Thus, the analysis of distress by non-invasive methods was not influenced by blood collection. The blood samples were taken before induction of CP without any intervention (pre-phase) and throughout the different phases of disease progression; early (day 2), middle (day 16) and late (day 30) between 11.00 and 15.30 h to refrain from the circadian increase of plasma corticosterone after 16.00 h60,61. Blood was collected 30 minutes after the last cerulein injection by retro-orbital puncture. This time interval is important because the plasma corticosterone concentration is highest 30 minutes after cerulein injection62. Mice were placed into a chamber filled with 5% isoflurane and 100–150µl blood was taken within three minutes of the onset of anaesthesia. The rationale behind using retro-orbital puncture is that this method yields a rather large volume of 100 µl of blood in a short time interval of 10 sec without the need of subsequent manual haemostasis. Blood sampling within three minutes is mandatory in order to exclude that the sampling procedure increases the corticosterone concentration63,64. Retro-orbital puncture is more than seven times faster than saphenous vein puncture and more than 15 times faster than tail vein puncture when collecting the same volume of blood65. Blood sampling was performed by a well experienced researcher. Blood samples were centrifuged at 1200 × g for 10 minutes and stored at −20 °C. Plasma corticosterone concentration was measured using an ELISA-Kit (DEV 9922, Demeditec Diagnostics GmbH, Erfurt, Germany) according to the manufacturer‘s instructions.
Lipase activity and immunohistochemistry
Lipase activity was detected in blood plasma by colorimeter (Cobas C111, Roche diagnostics GmbH, Mannheim, Germany) using a kit (LIPC REF # 05401704, Roche diagnostics GmbH, Mannheim, Germany) according to the manufacturer’s instructions. Immunohistochemistry was performed using a rabbit polyclonal anti-collagen I primary antibody (ab34710, Abcam, Cambridge, UK) at 1:200 dilution and a goat anti-rabbit AP-conjugated secondary antibody at a 1:100 dilution (D0487, Dako, Hamburg, Germany). The pre-treated slides were incubated with permanent red solution (K0640, Dako) prepared according to the manufacturer’s instructions. Image J 1.52a software (National Institutes of Health, USA) was used to quantify collagen I in an automated manner.
Graphs and statistical analysis
All box plots represent the 10th and 90th percentile as whiskers and all line graphs show mean and standard deviation. They were graphed using SigmaPlot 12.0 (SYSTAT Software Inc., San Jose, USA). All biostatistical analysis was done using SigmaPlot 12.0 (SYSTAT Software Inc., San Jose, USA), except for the t-test with Welch's correction, which was performed using GraphPad prism 6.0 (GraphPad Software, San Diego, USA). Significances of differences were evaluated using One Way Repeated Measures ANOVA, Two Way Repeated Measures ANOVA, Student t-test, t-test with Welch’s correction, Mann-Whitney rank sum test, paired t-test or Wilcoxon signed-rank test (see figure legends) according to data characteristics (the decision was based on the fact, if the data were paired and if they passed the Shapiro-Wilk normality test and the Levene median test for equal variance). Differences with p ≤ 0.05 were considered to be significant.
In order to compare the validity of parameters for defining distress, ROC analysis was performed. The data of all animals before and after intervention (either induction of CP or laparotomy) were used for distress prediction and the area under the curve, the 95% confidence intervals, the p-value and the best cut off (based on the highest calculated Youden’s index) were calculated for each parameter. ROC curves present the accuracy of a diagnostic test (an area under the curve of 1.0 means that the parameter is perfect to discriminate between the animals before and after the intervention, whereas a value of 0.5 indicates no discriminative power for this parameter). To analyze the efficacy of the combination of all four parameters, the data sets were combined by binary logistic regression in a multiple model and the ROC curves were calculated afterwards. The best cut off of this multi-parametric distress analysis was calculated using data set 1 of the laparotomy experiment. This cut off was applied to laparotomy data set 2 for verification. The identical cut off was then applied to classify distress after induction of laparotomy or induction of CP in form of a 2 × 2 contingency table. Subsequently, the Fisher‘s exact test was pursued to determine significances in the distribution of two distinct distress levels after laparotomy and cerulein-induced CP.
Data Availability
The datasets used and/or analyzed during the current study are available from the corresponding authors on reasonable request.
References
da Silva Morais, A., Oliveira, J. M. & Reis, R. L. Small Animal Models. Adv Exp Med Biol 1059, 423–439, https://doi.org/10.1007/978-3-319-76735-2_19 (2018).
Campos, G., Fortuna, A., Falcão, A. & Alves, G. In vitro and in vivo experimental models employed in the discovery and development of antiepileptic drugs for pharmacoresistant epilepsy. Epilepsy Res 146, 63–86, https://doi.org/10.1016/j.eplepsyres.2018.07.008 (2018).
Russell, W. M. S. & Burch, R. L. The principles of humane experimental technique (Methuen, London, 1959).
Office, P. Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the protection of animals used for scientific purposesText with EEA relevance. Home page at https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2010:276:0033:0079:en:PDF (2010)
Institutional Animal Care and Use Committee guidebook. 2nd ed. (Office of Laboratory Animal Welfare National Institutes of Health, S.l., 2002).
Netting, J. US dispute over definition of animal distress. Nature 406, 668, https://doi.org/10.1038/35021285 (2000).
National Research Council (US) Committee on Recognition and Alleviation of Distress in Laboratory Animals. Recognition and Alleviation of Distress in Laboratory Animals. (Washington (DC): National Academies Press (US); 2008).
Koch, V. W. Pain and distress: what really matters? Lab Anim (NY) 35, 27–32, https://doi.org/10.1038/laban0506-27 (2006).
Brown, M. et al. Report of the Working Group on Animal Distress in the Laboratory. Lab Anim (NY) 35, 26–30, https://doi.org/10.1038/laban0906-26 (2006).
Häger, C. et al. Running in the wheel: Defining individual severity levels in mice. PLoS Biol 16, e2006159, https://doi.org/10.1371/journal.pbio.2006159 (2018).
Hohlbaum, K. et al. Impact of repeated anesthesia with ketamine and xylazine on the well-being of C57BL/6JRj mice. PloS one 13, e0203559, https://doi.org/10.1371/journal.pone.0203559 (2018).
Hestehave, S. et al. Is there a reasonable excuse for not providing post-operative analgesia when using animal models of peripheral neuropathic pain for research purposes? PloS One 12, e0188113, https://doi.org/10.1371/journal.pone.0188113 (2017).
Bodden, C. et al. Evidence-based severity assessment: Impact of repeated versus single open-field testing on welfare in C57BL/6J mice. Behav Brain Res 336, 261–268, https://doi.org/10.1016/j.bbr.2017.08.029 (2018).
Jirkof, P. Burrowing and nest building behavior as indicators of well-being in mice. J Neurosci Methods 234, 139–146, https://doi.org/10.1016/j.jneumeth.2014.02.001 (2014).
Deacon, R. Assessing burrowing, nest construction, and hoarding in mice. J Vis Exp 59, e2607, https://doi.org/10.3791/2607 (2012).
Pain and distress in laboratory rodents and lagomorphs. Report of the Federation of European Laboratory Animal Science Associations (FELASA) Working Group on Pain and Distress accepted by the FELASA Board of Management November 1992. Lab Anim 28, 97–112, https://doi.org/10.1258/002367794780745308 (1994).
Touma, C., Palme, R. & Sachser, N. Analyzing corticosterone metabolites in fecal samples of mice: a noninvasive technique to monitor stress hormones. Horm Behav 45, 10–22 (2004).
Hohlbaum, K. et al. Severity classification of repeated isoflurane anesthesia in C57BL/6JRj mice-Assessing the degree of distress. PloS One 12, e0179588, https://doi.org/10.1371/journal.pone.0179588 (2017).
Hohlbaum, K. et al. Systematic Assessment of Well-Being in Mice for Procedures Using General Anesthesia. J Vis Exp 133, e57046, https://doi.org/10.3791/57046 (2018).
Benedetti, M. et al. Plasma corticosterone levels in mouse models of pain. Eur J Pain 16, 803–815, https://doi.org/10.1002/j.1532-2149.2011.00066.x (2012).
Rice, D. A. et al. Persistent postoperative pain after total knee arthroplasty: a prospective cohort study of potential risk factors. Br J Anaesth 121, 804–812, https://doi.org/10.1016/j.bja.2018.05.070 (2018).
Mazzara, S. et al. CombiROC: an interactive web tool for selecting accurate marker combinations of omics data. Sci Rep 7, 45477, https://doi.org/10.1038/srep45477 (2017).
Schrage, B. et al. Neuron-specific-enolase as a predictor of the neurologic outcome after cardiopulmonary resuscitation in patients on ECMO. Resuscitation 136, 14–20, https://doi.org/10.1016/j.resuscitation.2019.01.011 (2019).
Matias Júnior, I. et al. Effective Parameters for Gait Analysis in Experimental Models for Evaluating Peripheral Nerve Injuries in Rats. Neurospine 2, 305–316, https://doi.org/10.14245/ns.1836080.040 (2019).
Jacobsen, P. B. et al. Screening for psychologic distress in ambulatory cancer patients. Cancer 103, 1494–1502, https://doi.org/10.1002/cncr.20940 (2005).
Recklitis, C. J., Blackmon, J. E. & Chang, G. Screening young adult cancer survivors for distress with the Distress Thermometer: Comparisons with a structured clinical diagnostic interview. Cancer 122, 296–303, https://doi.org/10.1002/cncr.29736 (2016).
Zhou, Q. et al. Transforming growth factor-β in pancreatic diseases: Mechanisms and therapeutic potential. Pharmacol Res 142, 58–69, https://doi.org/10.1016/j.phrs.2019.01.038 (2019).
Poulsen, J. L., Olesen, S. S., Malver, L. P., Frøkjær, J. B. & Drewes, A. M. Pain and chronic pancreatitis: a complex interplay of multiple mechanisms. World J Gastroenterol 19, 7282–7291, https://doi.org/10.3748/wjg.v19.i42.7282 (2013).
Keller, C. E., Wilcox, C. M., Gudleski, G. D., Branham, S. & Lackner, J. M. Beyond Abdominal Pain: Pain Beliefs, Pain Affect, and Distress as Determinants of Quality of Life in Patients With Chronic Pancreatitis. J Clin Gastroenterol 52, 563–568, https://doi.org/10.1097/MCG.0000000000000922 (2018).
Zechner, D. et al. Diabetes increases pancreatic fibrosis during chronic inflammation. Exp Biol Med (Maywood) 239, 670–676, https://doi.org/10.1177/1535370214527890 (2014).
Zhan, X., Wang, F., Bi, Y. & Ji, B. Animal models of gastrointestinal and liver diseases. Animal models of acute and chronic pancreatitis. Am J Physiol Gastrointest Liver Physiol 311, G343–55, https://doi.org/10.1152/ajpgi.00372.2015 (2016).
Drewes, A. M. et al. Guidelines for the understanding and management of pain in chronic pancreatitis. Pancreatology 17, 720–731, https://doi.org/10.1016/j.pan.2017.07.006 (2017).
Anderson, M. A. et al. Mechanism, assessment and management of pain in chronic pancreatitis: Recommendations of a multidisciplinary study group. Pancreatology 16, 83–94, https://doi.org/10.1016/j.pan.2015.10.015 (2016).
Niederberger, E., Resch, E., Parnham, M. J. & Geisslinger, G. Drugging the pain epigenome. Nat Rev Neurol 13, 434–447, https://doi.org/10.1038/nrneurol.2017.68 (2017).
Zhang, Z.-J. et al. TLR8 and its endogenous ligand miR-21 contribute to neuropathic pain in murine DRG. J Exp Med 215, 3019–3037, https://doi.org/10.1084/jem.20180800 (2018).
Hori, N. et al. Changes in the expression of IL-6-Mediated MicroRNAs in the dorsal root ganglion under neuropathic pain in mice. Synapse 70, 317–324, https://doi.org/10.1002/syn.21902 (2016).
Drewes, A. M. et al. Pain in chronic pancreatitis: the role of neuropathic pain mechanisms. Gut 57, 1616–1627, https://doi.org/10.1136/gut.2007.146621 (2008).
Ceyhan, G. O., Michalski, C. W., Demir, I. E., Müller, M. W. & Friess, H. Pancreatic pain. Best Pract Res Clin Gastroenterol 22, 31–44, https://doi.org/10.1016/j.bpg.2007.10.016 (2008).
Resch, M., Neels, T., Tichy, A., Palme, R. & Rülicke, T. Impact assessment of tail-vein injection in mice using a modified anaesthesia induction chamber versus a common restrainer without anaesthesia. Lab Anim (UK) 53, 190–201, https://doi.org/10.1177/0023677218786982 (2019).
Jirkof, P. et al. Burrowing is a sensitive behavioural assay for monitoring general wellbeing during dextran sulfate sodium colitis in laboratory mice. Lab Anim 47, 274–283, https://doi.org/10.1177/0023677213493409 (2013).
Obafemi, T. F. et al. Comparable Responses in Male and Female Mice to Cerulein-Induced Chronic Pancreatic Injury and Recovery. JOP 19, 236–243 (2018).
Jirkof, P. et al. Burrowing behavior as an indicator of post-laparotomy pain in mice. Front Behav Neurosci 4, 165, https://doi.org/10.3389/fnbeh.2010.00165 (2010).
Pfeiffenberger, U. et al. Assessment and refinement of intra-bone marrow transplantation in mice. Lab Anim 49, 121–131, https://doi.org/10.1177/0023677214559627 (2015).
Palme, R. Non-invasive measurement of glucocorticoids: Advances and problems. Physiol Behav 199, 229–243, https://doi.org/10.1016/j.physbeh.2018.11.021 (2019).
Nedaeinia, R. et al. Inhibition of microRNA-21 via locked nucleic acid-anti-miR suppressed metastatic features of colorectal cancer cells through modulation of programmed cell death 4. Tumour Biol 39, 1010428317692261, https://doi.org/10.1177/1010428317692261 (2017).
Thum, T. et al. MicroRNA-21 contributes to myocardial disease by stimulating MAP kinase signalling in fibroblasts. Nature 456, 980–984, https://doi.org/10.1038/nature07511 (2008).
Wang, H., Xu, W., Shao, Q. & Ding, Q. miR-21 silencing ameliorates experimental autoimmune encephalomyelitis by promoting the differentiation of IL-10-producing B cells. Oncotarget 8, 94069–94079, https://doi.org/10.18632/oncotarget.21578 (2017).
Zhao, Q., Manohar, M., Wei, Y., Pandol, S. J. & Habtezion, A. STING signalling protects against chronic pancreatitis by modulating Th17 response. Gut, https://doi.org/10.1136/gutjnl-2018-317098 (2019).
Zhang, L. et al. Decreased immunoglobulin G in brain regions of elder female APOE4-TR mice accompany with Aβ accumulation. Immun Ageing 16, 2, https://doi.org/10.1186/s12979-018-0142-7 (2019).
Kurstjens, S. et al. Diabetes-induced hypomagnesemia is not modulated by metformin treatment in mice. Sci Rep 9, 1770, https://doi.org/10.1038/s41598-018-38351-3 (2019).
Dimitroulas, T., Duarte, R. V., Behura, A., Kitas, G. D. & Raphael, J. H. Neuropathic pain in osteoarthritis: a review of pathophysiological mechanisms and implications for treatment. Semin Arthritis Rheum 44, 145–154, https://doi.org/10.1016/j.semarthrit.2014.05.011 (2014).
Chen, Q.-Y. et al. Mechanism of persistent hyperalgesia in neuropathic pain caused by chronic constriction injury. Neural Regen Res 14, 1091–1098, https://doi.org/10.4103/1673-5374.250631 (2019).
Vallès, A., Martí, O., Harbuz, M. S. & Armario, A. A single lipopolysaccharide administration is sufficient to induce a long-term desensitization of the hypothalamic-pituitary-adrenal axis. Neuroscience 112, 383–389, https://doi.org/10.1016/s0306-4522(02)00047-7 (2002).
Martí, O. & Armario, A. Anterior pituitary response to stress: time-related changes and adaptation. Int J Dev Neurosci 16, 241–260 (1998).
Ryabinin, A. E., Wang, Y. M. & Finn, D. A. Different levels of Fos immunoreactivity after repeated handling and injection stress in two inbred strains of mice. Pharmacol Biochem Behav 63, 143–151, https://doi.org/10.1016/s0091-3057(98)00239-1 (1999).
Stamp, J. & Herbert, J. Corticosterone modulates autonomic responses and adaptation of central immediate-early gene expression to repeated restraint stress. Neuroscience 107, 465–479, https://doi.org/10.1016/s0306-4522(01)00364-5 (2001).
Zechner, D. et al. Intratumoral heterogeneity of the therapeutical response to gemcitabine and metformin. Oncotarget 7, 56395–56407, https://doi.org/10.18632/oncotarget.10892 (2016).
Zhang, X. et al. Blocking Autophagy in Cancer-Associated Fibroblasts Supports Chemotherapy of Pancreatic Cancer Cells. Front Oncol 8, 590, https://doi.org/10.3389/fonc.2018.00590 (2018).
Touma, C., Sachser, N., Möstl, E. & Palme, R. Effects of sex and time of day on metabolism and excretion of corticosterone in urine and feces of mice. Gen Comp Endocrinol 130, 267–278 (2003).
Dalm, S. et al. Age-related changes in hypothalamic-pituitary-adrenal axis activity of male C57BL/6J mice. Neuroendocrinology 81, 372–380, https://doi.org/10.1159/000089555 (2005).
Gong, S. et al. Dynamics and correlation of serum cortisol and corticosterone under different physiological or stressful conditions in mice. PloS One 10, e0117503, https://doi.org/10.1371/journal.pone.0117503 (2015).
Kumstel, S. et al. Grading Distress of Different Animal Models for Gastrointestinal Diseases Based on Plasma Corticosterone Kinetics. Animals (Basel). 9, https://doi.org/10.3390/ani9040145 (2019).
Vahl, T. P. et al. Comparative analysis of ACTH and corticosterone sampling methods in rats. Am J Physiol Endocrinol Metab. 289, E823–8, https://doi.org/10.1152/ajpendo.00122.2005 (2005).
Kugler, J., Lange, K. W. & Kalveram, K. T. Influence of bleeding order on plasma corticosterone concentration in the mouse. Exp Clin Endocrinol. 91, 241–243, https://doi.org/10.1055/s-0029-1210754 (1988).
van Herck, H. et al. Blood sampling from the retro-orbital plexus, the saphenous vein and the tail vein in rats: comparative effects on selected behavioural and blood variables. Lab Anim. 35, 131–139, https://doi.org/10.1258/0023677011911499 (2001).
Acknowledgements
This study was supported by the Deutsche Forschungsgemeinschaft (DFG research group FOR 2591, project number: 321137804, ZE 712/1–1 and VO 450/15–1). The authors are grateful for the perfect technical assistance from Berit Blendow, Dorothea Frenz and Edith Klobetz-Rassam.
Author information
Authors and Affiliations
Contributions
Individual author contributions were as follows: D.Z. and B.V. acquired the funding. D.Z., T.T. and B.V. developed the study concept. D.Z., X.Z., M.L., R.P. and B.V. developed the concept for experiments or data evaluation. A.A., S.K., E.H.U.W., J.E., D.Z. and R.P. carried out the experiments. A.A., D.Z., S.K., E.H.U.W., J.E. analyzed data. A.A. and D.Z. wrote the manuscript. All authors read and approved the final version of the manuscript.
Corresponding authors
Ethics declarations
Competing Interests
T.T. filed and licensed patents about the use of miR-21 modulators for cardiovascular disease and fibrosis treatment. T.T. is founder and shareholder of Cardior Pharmaceuticals GmbH. All other authors declare no potential conflicts of interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Abdelrahman, A., Kumstel, S., Zhang, X. et al. A novel multi-parametric analysis of non-invasive methods to assess animal distress during chronic pancreatitis. Sci Rep 9, 14084 (2019). https://doi.org/10.1038/s41598-019-50682-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-50682-3
This article is cited by
-
Robustness of a multivariate composite score when evaluating distress of animal models for gastrointestinal diseases
Scientific Reports (2023)
-
The impact of handling technique and handling frequency on laboratory mouse welfare is sex-specific
Scientific Reports (2020)
-
MicroRNAs as systemic biomarkers to assess distress in animal models for gastrointestinal diseases
Scientific Reports (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
