
Abstract
In natural populations, allelic diversity of the major histocompatibility complex (MHC) is commonly interpreted as resulting from positive selection in varying spatiotemporal pathogenic landscapes. Composite pathogenic landscape data are, however, rarely available. We studied the spatial distribution of allelic diversity at two MHC class II loci (DQA, DQB) in hares, Lepus capensis, along a steep ecological gradient in North Africa and tested the role of climatic parameters for the spatial distribution of DQA and DQB proteins. Climatic parameters were considered to reflect to some extent pathogenic landscape variation. We investigated historical and contemporary forces that have shaped the variability at both genes, and tested for differential selective pressure across the ecological gradient by comparing allelic variation at MHC and neutral loci. We found positive selection on both MHC loci and significantly decreasing diversity from North to South Tunisia. Our multinomial linear models revealed significant effects of geographical positions that were correlated with mean annual temperature and precipitation on the occurrence of protein variants, but no effects of co-occurring DQA or DQB proteins, respectively. Diversifying selection, recombination, adaptation to local pathogenic landscapes (supposedly reflected by climate parameters) and neutral demographic processes have shaped the observed MHC diversity and differentiation patterns.
Similar content being viewed by others

Introduction
Genetic studies of natural populations based on presumably neutral markers such as mitochondrial d-loop DNA (mtDNA), microsatellites or SNPs are important to infer phylogeny, population history and gene flow1,2. However, in some cases the time span between the separation of populations might be too short or gene flow might be too strong to leave a signal of differentiation at neutral loci3. In such cases, differences between populations can only be detectable at functional genes under selection3. Moreover, studying molecular polymorphism at loci under natural selection will help to understand the genetics of adaptive processes and to increase our knowledge on the importance of adaptive genetic variability in free ranging animal populations1. Local adaptation at functional markers has mainly been addressed in populations structured at neutral markers and rarely in the case of seemingly panmictic populations4. A study of Tunisian hare populations indicated a strong population structure at mitochondrial OXPHOS genes, which are under positive selection at several codons, despite a high gene flow at neutral markers5. These findings are consistent with local adaptation according to climate variation5.
Adaptation to local or regional pathogenic landscapes is often addressed by analyses of major histocompatibility genes (MHC) which code for glycoproteins that recognize and bind antigens in order to present them to CD4+ and CD8+T-lymphocytes to initiate the adaptive immune response against pathogens. MHC class I and class II loci, the two main types of MHC loci involved in the adaptive immune system, have been shown to be highly polymorphic in a wide range of species6,7,8,9 and their adaptive significance in vertebrates has long been recognized10,11. Allelic diversity at these loci can be associated with variation across environmental and geographical scales as demonstrated from several studies12,13. The role of environmental characteristics in shaping selective pressures on MHC genes is suggested to be mediated by combined effects including recent and ancient demography as well as behavioural and ecological differences in pathogens exposure14,15,16. Moreover, host-pathogen interactions have been suggested to be affected by temperature, nutrient availability and environmental stress17. Whereas increases of ambient temperature can reinforce selection on immune genes, thereby accelerating host adaptation18,19, variation in nutrient levels can force physiological modification that could either impair or promote local adaptation of the host17.
Among all MHC molecules, class II genes have received the most attention, due to their role in studying mechanisms and significance of molecular adaptation in vertebrates20 and host parasite co-evolution2. MHCII molecules are heterodimers formed by an α and a β chain encoded by A and B genes, respectively21,22. These chains are characterized by several typical structures, such as N-linked glycosylation sites, connecting peptides, transmembrane regions, and cytoplasmic domains, as well as the α1/α2 domain and the β1/β2 domain, respectively23. The α1 and β1 domains constitute the peptide-binding region (PBR), involved with the recognition and binding to the antigens14,24,25. These domains are highly polymorphic and are encoded by the second exon of A and B MHCII genes. Given their role in presenting antigenic determinant peptides to the cell surface, these loci are typically expected to be subject to strong positive selection, since their greater allelic diversity should be associated with a response to a wider range of pathogens16. Genetic diversity in the MHC is also generated through several mechanisms, including point mutation, recombination, gene conversion, sexual selection and maternal–foetal interactions25,26,27,28. The variation generated by these processes is thought to be maintained in individuals and populations by a combination of balancing and diversifying selection, driven primarily by variation in selection pressure from pathogens and parasites across space and time3,28. Balancing selection encompasses a number of different evolutionary mechanisms that result in a large number of alleles being maintained in populations for longer than would be expected relative to neutral genetic variation24. Such balancing selection is generally explained by three mechanisms a) over-dominant selection or heterozygote advantage where heterozygous individuals have higher fitness than both corresponding homozygotes; b) frequency dependent selection or rare allele advantage where individuals with rare genotypes have higher fitness29,30; c) fluctuation in selection pressure where MHC polymorphism is maintained by changing pathogen composition through time and space. Conversely, under diversifying selection at MHC loci, the spread of an advantageous allele (positive selection) would be expected to lead to a loss of genetic variation. Similarly, selection against disadvantageous alleles (negative or purifying selection) would also be expected to reduce diversity31. Among the different suggested mechanisms of balancing selection, fluctuation in selection pressure have been reported for class II MHC genes among populations of the Asian cygomolgus macaque (Macaca fascicularis)12, and the Atlantic salmon32, among other studies. Patterns of MHC selection can be inferred by comparing MHC genetic differentiation to differentiation at presumably neutral markers28. Neutral loci reflect demographic factors such as migration and drift, whereas loci under selection usually show higher differentiation under different local selection regimes or lower differentiation under balancing selection28,33 or under negative/purifying selection33. Higher differentiation at MHC genes could therefore reflect local adaptation to different parasite faunas prevailing in the respective populations.
In Tunisia, hares (Lepus capensis) are distributed throughout the whole country in diverse environments and over short geographic distances, reaching from the Mediterranean climate in the north to the pronounced arid desert climate (Sahara) in the south. This situation provides a good precondition to study natural selection and adaptation on functional genes5. The likely differences in differential parasite pressure in these habitats could lead to different selection pressure at MHC loci among the local populations. Such selective pressure would shape diversity and differentiation profiles in interaction with neutral demographic/stochastic factors. In this study, we examined the MHC class II polymorphism for DQA and DQB genes in hares from Tunisia along a steep ecological gradient. We first investigated the variation of genetic diversity in the different studied populations distributed in heterogeneous ecoregions from North to South. If diversifying selection is the main evolutionary force acting on MHC genes, we expected significant genetic differentiation despite observed high gene flow in neutral markers. Second, we used different approaches to evaluate the effect of selective pressure in each codon of the obtained sequences. Moreover, we compared the levels of allelic diversity of the considered hare populations with those obtained for a set of neutral loci. We expect that MHC diversity varies significantly between the regional populations under study given that spatial changes in parasite communities and dynamics with environmental conditions might alter the selection pressures they exert on their host populations17. However, neutral demographic processes (i.e., gene flow, genetic drift…) can also have significant effects on the genetic diversity of natural populations4,34.
Given the steep ecological gradient along the relatively short geographical distance from where our samples were drawn and the absence of any obvious migration barrier across our study area, significant variation of MHC allele frequencies across the study area should reflect adaptation to local environments. Thus, we hypothesize that climatic factors associated with the geographical origin of the samples will lead to spatial variation in the frequencies of the most common alleles. Moreover, MHC alleles at different loci are expected to co-evolve. Thus, we further expect a significant level of linkage between the respective co-occurring allele or genotype at the second MHC locus studied on the occurrence of an MHC allele under consideration. Such allele effects of co-occurring MHC loci might have led to an “optimal” combination of alleles at the two loci studied independently of spatial or climatic effects.
Results
Allelic diversity and genetic differentiation
We identified 17 DQA alleles (accession numbers MH346126-MH346142) and 26 DQB alleles (accession numbers MH346143- MH346168) in 142 hares (L. capensis) from Tunisia with a total length of 228 and 222 bp, respectively. Among them, only three DQA alleles (Lecp-DQA*04, Lecp-DQA*05, Lecp-DQA*06 corresponding to Leeu-DQA*02, Leeu-DQA*03, Leeu-DQA*09 in L. europaeus, respectively) were previously described in L. europaeus. No insertions, deletions, or stop codons were detected in either DQA or DQB sequences. The nucleotide sequence of each allele in both loci translated into a different amino-acid sequence (Figs 1 and 2). Allele frequencies and basic diversity parameters for the two MHC class II exon 2 loci in the different populations as well as the overall values are displayed in Table 1. Hares were grouped into three populations according to climatic, geographic and phenotypic data (see methods section). These populations are northern Tunisia (NT), central Tunisia (CT) and southern Tunisia (ST).

Amino acid alignment of DQA exon 2. Antigen-binding sites (ABS) are indicated above with the sign “+”. Positively selected codons identified by the different methods are indicated with asterisks. Numbering of amino acid residues is according to Bondinas et al.89.

Amino acid alignment of DQB exon 2. Antigen-binding sites (ABS) are indicated above with the sign “+”. Positively selected codons identified by the different methods are indicated with asterisks. Numbering of amino acid residues is according to Bondinas et al.35.
The numbers of alleles per regional population were 14, 12, and 9 in NT, CT, and ST, respectively, for the DQA locus. For DQB, allele numbers were 18, 15, and 11 in NT, CT and ST, respectively (Table 1). In general, in all three populations, few alleles were relatively abundant, e.g. Lecp-DQA*07, Lecp-DQA*08, Lecp-DQB*01 and Lecp-DQB*09 were highly frequent, whereas most others were detected at very low frequencies.
A comparison of the obtained genotypes revealed alleles specific to each regional population. For DQA, seven alleles were shared between the three populations, whereas three alleles (Lecp-DQA*18, Lecp-DQA*19, Lecp-DQA*20) were specific to NT, one allele (Lecp-DQA*04) to CT and two (Lecp-DQA*13, Lecp-DQA*17) to ST. For DQB, only four alleles (Lecp-DQB*01, Lecp-DQB*02, Lecp-DQB*09, Lecp-DQB*12) were shared between the three regional populations. Nine alleles were specific to NT, two to CT, and two to ST. Frequencies of these specific alleles ranged between 0.0161 and 0.0968 for DQA and between 0.0137 and 0.0500 for DQB (Table 1). Finally, allelic richness (Rs), which measures the number of alleles independently of sample size, was significantly (p < 0.0001) higher for the two MHC loci than for the microsatellite loci in NT but not in CT and ST. Moreover, allelic richness varies significantly (p = 0.027) between NT and the two other populations (CT and ST). Significant (FIS for NT = 0.384, p < 0.001; FIS for CT = 0.447, p < 0.001; FIS for ST = 0.338, p < 0.001) deviation from Hardy-Weinberg Equilibrium in all three regional populations indicated that the MHC loci contained significantly fewer heterozygotes than expected. Finally, there was no significant linkage disequilibrium between genotypes at the DQA and DQB locus as indicated by the Linkage disequilibrium test (R = 0.16; p = 0.22).
Genetic differentiation between regional populations was revealed by pairwise comparisons of Dest and FST values (Table 2). Population differentiation as determined by mean Dest values across the two loci was high and ranged between 0.46 (NT vs CT) and 0.78 (NT vs ST). For the individual loci, Dest values for the DQA locus ranged between 0.29 (NT vs CT) and 0.59 (CT vs ST), whereas those for the DQB locus ranged between 0.73 (NT vs CT) and 0.96 (NT vs ST). Similarly, pairwise FST (Table 2) values revealed significant genetic divergence among regional populations ranging between 0.12 (NT vs CT) and 0.19 (NT vs ST). In contrast, pairwise values for the microsatellite loci were low ranging between 0.013 to 0.030 for FST values and between 0.043 and 0.112 for pairwise Dest values (Table 2). Finally, a hierarchical population structure (AMOVA) based on analyses of variance of gene frequencies was evident in the genetic differentiation across both MHC loci, with 17.13% of the genetic variance attributable to between regional population differences, 4.79% between sampling localities within regional populations, and 78.09% to within sampling localities (P < 0.001, P < 0.05, and P < 0.001, respectively). These estimations for the microsatellite loci were 0.60 (P > 0.05), 4.58 (P < 0.001) and 94.82 (P < 0.001) for the same parameters, respectively.
The first two axes of our PCA (Fig. 3) based on the DQA and DQB allele matrix explained 31.51% of the variance in our MHC genotypic data (18.19% F1, 13.32% F2). Both axes (F1 and F2) were structured by DQA alleles DQA*7, DQA*8, DQA*11, DQA*13 and DQB alleles DQA*1, DQA*3, DQA*9 (Fig. 3). Among these alleles, only DQA*13 was a rare allele.

Principal component analysis (PCA) of the allelic diversity in the different sampling regions of hares. Each dot represents one MHC allele; the DQA and DQB alleles are indicated as A-allele number and B-allele number, respectively.
Finally, bottleneck signatures were detected in CT (p = 0.0176) and ST (p = 0.0017) populations when using the Two Phase Mutation model (TPM). No tests detected a bottleneck in the northern Tunisian (NT) population. Different Ne estimates were obtained for each population depending on the used method (Table 3). The Heterozygote Excess based method gave an infinitely large estimate of Ne for all populations. The smallest estimates were observed in the ST population (317.9) by the Linkage disequilibrium method.
Selection and recombination analysis
Using the three PERMUTE statistics, no recombination was obtained for the DQA locus, whereas recombination evidence was obtained by one test (r2 = −0.163; p = 0.001) for the DQB locus (Table 4). One recombination event was detected for each locus when using GARD and one to three events were detected for DQB and DQA, respectively, using the RDP statistics (Tables S1 and S2). The estimates of population mutation (θ) and recombination (ρ) obtained with the LDhat approach showed that each of these processes has played a significant role in generating the diversity seen among the currently studied MHC beta genes (Table 4). To find out the relative contribution of recombination and point mutations in the evolution of these two currently studied genes in hares, we examined the ratio of average mutation (θ) and recombination rate (ρ). By this measure, mutation was of approximately hundred times more important in the evolution of DQA than recombination. In contrast, the recombination rate was about one and a half times greater than mutation in the evolution of the DQB gene (Table 4).
The codons found to be positively selected by all methods are shown in Figs 1 and 2. Five codons (8, 10, 16, 55 and 69) were considered to be under episodic diversifying selection for DQA as evidenced by more than one of the applied tests in DATAMONKEY. Six (8, 10, 50, 53, 55 and 76) and three (47, 55 and 69) codons were reported under positive selection for the same locus by PAML (see Table S3) and OMEGAMAP, respectively (Figs 1 and 2). Only site 55 was confirmed by all methods. Twelve codons were suggested to be under negative purifying selection in DQA as indicated by the different DATAMONKEY tests.
For DQB, five positions (amino acid sites 8, 26, 48, 60, 67) were under episodic diversifying selection in DATAMONKEY. Ten and eleven codons were detected with PAML (codons 13, 14, 26, 32, 37, 48, 57, 59, 60 and 67) and OMEGAMAP (13, 14, 26, 30, 37, 48, 57, 59, 60, 66 and 67), respectively. Four codons (26, 48, 60 and 67) were confirmed by all three methods to be under positive selection. Negative selection in the DQB locus was detected in five codons (15, 16, 45, 62 and 72) as suggested by DATAMONKEY tests.
Finally, the outlier test of Beaumont and Nichols35 indicated departure from expectation under a neutral evolution scenario for both the DQA and the DQB locus (Fig. 3), with higher values than the microsatellite loci, in accordance with the codon-based signals of positive selection.
Statistical Models of DQA and DQB protein occurrence
In three cases we found models (Table 5) with excellent support from the data (i.e. delta AICc > 10, Null-model insubstantial36), in two other cases models with strong support (delta AICc > 8) and in two cases models with some support (delta AICc > 4). Two models had a deviance explained of more than 40%, which is notable, because in multinomial models nominal data are compared with quantitative predictions.
In all models except for the “dqb1gt model” geographic variables (latitude or longitude) had a high RVI-value (> = 0.89) indicating a high probability for these variables being in the best model. Furthermore, the delta deviance value of one of the geographic variables was more than ten times higher than the respective co-occurring DQA or DQB protein variants combinations in four models and eight times higher in the “dqa11gt”-model (Table 5; see also Supplementary Table S4 for estimated coefficients, standard errors, and upper and lower bounds of the 95% confidence interval for all models). This indicates strong support for an effect of the geographic variables on the dependent variables in comparison to the respective co-occurring DQA or DQB protein variants combinations. In the “dqb1gt-model”, again the performance was very low. Overall, these results suggest a strong effect of geographic variables on the co-occurring DQA or DQB protein variants. In particular, the proteins of Lecp-DQA*07 and Lecp-DQA*08 did occur more often in homozygous state in Northern Tunisia, with lower mean annual temperatures and higher levels of precipitation, whereas proteins of Lecp DQA*11 were more common in South Tunisia, with higher mean annual temperatures and lower precipitation. For the DQB locus, proteins of Lecp-DQB*01 were more frequent, in homozygous state, in eastern parts of Tunisia, proteins of Lecp-DQB*01 in heterozygous state as well as proteins of Lecp-DQB*09 in homozygous and heterozygous state were more common in North Tunisia, whereas proteins of Lecp-DQB*03 in homozygous and heterozygous state were more frequent in South Tunisia. Moreover, proteins were generally more common, in heterozygous state, in North Tunisia.
Discussion
We assessed the MHC diversity and its distribution at two MHC class II genes, DQA and DQB, in hare populations (L. capensis) along a steep ecological gradient in North Africa. We found 17 DQA and 26 DQB alleles among 142 L. capensis individuals. We have analysed the molecular mechanisms of sequence evolution in both loci, i.e., substitution, recombination, and positive selection. We have also described the genetic differentiation and population structure and compared this across both MHC loci and populations. Apart from significant positive selection and recombination events, we found a significant decrease of allelic richness at the two MHC loci from the northern population under Mediterranean humid climate to the southern population in the Sahara region. In addition, we have found a high population substructuring at the two MHC loci compared to supposedly neutral microsatellite markers.
Our results show a high level of polymorphism for the DQA and DQB loci. Such wide allelic diversity in both allele numbers and pairwise nucleotide distances is characteristic for MHC genes37. Moreover, this is in accordance with the results of our previous analyses of the same individuals using mtDNA control region sequences38, partial transferrin sequences39, allozyme40, and microsatellites38, which confirms the generally high genetic variability of Tunisian hare populations. Notably, the detected levels of polymorphism of DQA and DQB were higher than those reported for brown hares (L. europaeus) from Austria and Belgium8,41,42,43. Klein44 and Takahata et al.45 suggested that such differences in polymorphism could be due to either a difference in the speed of allelic diversification or in time of their persistence in the gene pool. Both loci in our study have exhibited very similar substitution rates, but a greater recombination rate was recorded for DQB. This effect may be due to a longer persistence of DQA diversity in the populations, whereas DQB allele diversification may be more likely due to higher intra-locus recombination. Indeed, we found evidence for multiple (intra-allelic) recombination events creating novel DQA and DQB haplotypes. According to the different methods used (that vary in their conservative nature), breakpoints at position 120 and 221 were detected by only one test (Tables S1 and S2). For DQA, multiple breakpoint positions were obtained and confirmed by more than one of the methods. In contrast, the overall recombination rate (ρ = 16.771) of DQB was one and a half times more than the mean mutation rate (θ = 11.53) indicating that the generation of DQB diversity is driven by an accumulation of new recombinants relative to new mutations (ρ/θ = 1.455), which is similar to results obtained in other Lepus species43. For DQA, recombination seems to have a minor effect on its evolution compared to mutations (the ratio ρ/θ = 0.010). In contrast, Wegner46 showed that recombination rate and intensity of positive selection were positively correlated in MHC class I and class II sequences of nine different species of fish, suggesting that recombination and gene conversion played a significant role in the evolution of the studied MHC genes.
Genetic differentiation between the studied Tunisian hare populations was high in both MHC class II genes as suggested by high and significant FST and Dest values. But the same populations showed little genetic structure in the microsatellite loci, which is in accordance with mtDNA and nuclear sequences39,47, as well as microsatellite and allozyme genotypes38,40 studied earlier in these populations. Therefore, population genetic differentiation at MHC loci could result from either a divergence in qualitative and quantitative parasite pressure that would select for different types of alleles in the different ecoregions but may also be due to differences in modes of mutation and mutation rates between the two marker types. However, high gene flow was concordantly indicated by several markers38,39,40 (microsatellites, allozymes, mtDNA d-loop and transferrin sequences) characterized by different modes of mutation and mutation rates. Therefore, the observed pattern of differentiation only at MHC loci is likely the result of differential selection pressures across the studied populations which is consistent with theoretical expectations28.
Indeed, the few frequent alleles (those with allele frequency >10%; Lecp-DQA*07 and Lecp-DQB*09 for NT, Lecp-DQA*08 & Lecp-DQB*01 for CT, Lecp-DQA*11 and Lecp-DQB*03 for ST) vary greatly in each population for each locus. Those predominant alleles were suggested by the PCA to be responsible for the genetic differentiation observed at MHC loci between the three ecoregions. Such differences in allele frequencies between populations have been demonstrated to be mediated by pathogens in affected and unaffected chamois populations48. In contrast, cohorts of brown hares characterized by similar quantitative and qualitative pathogens have exhibited the same frequent allele as a sign of stabilizing selection41. Moreover, estimations of effective population size and bottleneck signature indicate that the reduced allelic richness of the ST population may be the result of a smaller and/or drifting population. Therefore, the lower MHC polymorphism, in terms of number of alleles, in the ST population may be due to demographic processes, independently from specific parasite pressure. The results indicate that the population structuring at MHC loci is linked to qualitative MHC polymorphism (allelic diversity: different alleles are frequent in different populations) likely shaped by variation in parasite pressure, but the quantitative MHC polymorphism (allelic richness: fewer alleles from North to South) may still be the results of demographic processes.
Both loci showed excess of non-synonymous to synonymous substitutions, a signal of positive selection49 that was stronger at the DQB locus than for DQA as indicated by a significantly higher number of positively selected sites. In contrast, negative purifying selection was stronger at the DQA than in DQB locus. In comparison with other studies of lagomorph species, Goüy de Bellocq et al.42 identified only one codon under positive selection (amino acid 56 in the current study) in thirteen DQA alleles of L. europaeus populations from Austria and Belgium and four codons (three were identified presently as well: 10, 56, and 70) among 19 Oryctolagus cuniculus alleles. For DQB, Smith et al.43 identified two codons (codons 26 and 60) under positive selection among eleven DQB alleles in L. europaeus, which are also under positive selection in the current study.
The observed heterozygosity values of MHC loci were rather low in all three regional populations showing a significant deficit. The low heterozygosity might fit to a pattern of recent expansion, as suggested by mitochondrial and nuclear sequences38,39 or be due to allelic dropout as different alleles might have similar conformation when using CE-SSCP techniques1,50. However, the distribution of MHC heterozygosity suggests that heterozygote advantage is unlikely given the observed positive selection in the studied populations. Indeed, the MHC FST values were clearly higher than those for microsatellite loci, suggesting an apparent effect of diversifying positive selection. This conclusion was also supported by the outliers test, where the DQA and DQB loci showed significantly higher mean FST than predicted from its mean heterozygosity (Fig. 4). Further support for our interpretation comes from our observation of regionally differentiated populations that also suggest diversifying selection maintained by changes of pathogen composition across geographic regions. However, other mechanisms, such as sexual selection and mating system, might drive the evolution of MHC genes28. Nevertheless, it is probable that only the effects of pathogens vary both over time and among populations8. Host-parasite interactions were suggested to be shaped by environmental conditions (i.e., temperature, nutrient availability; see Brunner and Eizaguirre17 for an overview) which can influence parasite species richness, as well as intensity of infestation of hosts. According to Klein and Horejsi51 an indirect assessment of parasite infestation can be seen via analysis of MHC class II molecules diversity in the host species, as these molecules primarily correspond to extracellular pathogens, such as bacteria, nematodes, cestodes. Indeed, associations between MHC variants and parasite load have been demonstrated in numerous cases28. Here we found significant effects of mean annual temperature and precipitation in the distribution of some MHC proteins/alleles which might indicate regional and climate effects. Concordantly, genetic differentiation and estimation of gene flow indicated regional distribution patterns for MHC loci compared to high genetic admixture in the neutral markers (i.e. microsatellites). Both results suggest that the MHC polymorphism across sampling populations might reflect an adaptation to variation in parasite pressure across the ecological gradient. We expect quantitative and qualitative changes of pathogens from the Mediterranean humid region in the north to the arid Sahara in the south. Indeed, Zvinorova et al.52 showed that the highest helminths and Eimeria infections in goats were observed in the wet season, whereas Pandey et al.53 suggested direct relationship between rainfall and intensity of infection with gastrointestinal nematodes in goats. Notably, diversity of the mammalian fauna that potentially constitutes a complex pathogen reservoir shows a reduction along the currently studied ecological gradient in Tunisia54, probably due to habitat changes and nutrient availability. Moreover, population density of hares from Tunisia seems to decrease from North to South Tunisia (personal observation during hunting trips). This may point towards a combined effect of various aspects, mainly driven by environmental conditions, resulting in the reduction of pathogens in the Sahara region and hence resulting in lower MHC diversity.

Estimated FST values from the 16 loci studied (14 microsatellites, DQA and DQB) plotted as a function of heterozygosity (He) using Lositan software. Gray shading indicates the area on the graph within the confidence limits. DQA and DQB were found to be under positive selection.
Estimation of effective population size showed that Ne estimates were not consistent across methods. This is expected as the accuracy and precision of the different methods to estimate Ne are still uncertain55. We followed Wang56 who showed that linkage disequilibrium (LD) methods perform dramatically better than the other methods. Ne estimates using this method indicated a similar effective population sizes in all three populations. Moreover, our current Ne estimate for the ST population was less than 400 which was suggested to be the minimum effective population size necessary to maintain genetic variation in the long term in a population57. This later population was suggested also by the bottleneck test on neutral microsatellite loci to be under genetic drift. Furthermore, Olivier et Piertney58 have demonstrated that selection counters drift to maintain polymorphism at a major histocompatibility complex (MHC) locus in water vole bottlenecked population from Coiresa Island. Therefore, we consider that demographic factors (e.g. bottlenecks or drift) were strong enough to shape the allelic richness in ST population and/or selective pressures were weak to counter the effect of genetic drift.
In conclusion, our present findings from hares along a steep ecological gradient in North Africa indicated that MHC polymorphism and differentiation were shaped by a combination of neutral demographic processes and climate-driven positive selection of MHC genes across relatively short geographical distances. How changes in pathogenic landscape diversity on the one hand, and how immunologic resistance or tolerance of infected hosts on the other hand may interact with local or regional climatic parameters in the evolution of MHC diversity and in the course of adaptation to environmental characteristics needs, however, further studies with particular inclusion of infection patterns of the hosts.
Methods
Samples
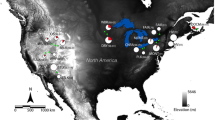
A total of 142 samples of cape hares (Lepus capensis, e.g.59,60,61) were collected during regulars hunts from various biogeographic zones within the major ecoregions in Tunisia. We never manipulated live animals; we got only hare samples during regular hunting season from hunters. All samples were collected with the agreement of the general forest department of Tunisia (authorization no. 2629DGF/DCF/CPN; Ministry of Agriculture, Water Resources and Fishing, Tunisia). Hares were grouped into three populations corresponding to the three main climatic zones in Tunisia (e.g. You et al.62). Acronyms of sampling localities and positions within northern (NT), central (CT), and southern (ST) Tunisia, as well as sample sizes are given in Fig. 5. The whole sampling area expanded from the Mediterranean humid and sub-humid bioclimatic zones in the north (NT), characterized by cork oak (Quercus suber) Mediterranean forest and scrubland (maquis: rosemary, carob tree, mastic tree, thyme, lavender), across the semi-arid central part of Tunisia (CT) to the arid and hyper-arid Sahara in the south (ST), with typical plants that have adapted to dry conditions such as Acacia tortilis subsp. raddiana, Vachellia spp., Stipagrostis pungens vegetation62,63. Our study area represents a remarkably steep climatic and ecological gradient across a straight distance of less than 500 km, with means of annual precipitation and temperature ranging between 916 mm and 16.3 °C in the north and 77 mm and 21.26 °C in the south.

Sampling regions of hares from North, Central and South Tunisia [The original map was created by using Google Maps, https://www.google.tn/maps/@34.6113892,8.7590835,6z?hl=fr and the map of Tunisia was drawn by using POWERPOINT 2007 part of the Microsoft Office Package (https://www.microsoft.com/fr-fr/software-download/office)]. Sample sizes appear in parentheses. Hares were grouped into three populations according to climatic, geographic and phenotypic data.
PCR amplification, CE-SSCP and Sequencing
Total genomic DNA was extracted using the GenElute Mammalian Genomic DNA Miniprep kit (Sigma-Aldrich) from diverse tissues (liver, skeletal muscles, ear cartilage, tongue) of each specimen5,39,47,60.
Amplification of exon 2 fragments for two MHC class II genes (DQA and DQB) were performed according to Goüy de Bellocq et al.42 and Smith et al.43. The amplified 258 bp and 249 bp (including primers) correspond to the total exon 2 of DQA and DQB genes, respectively. We used capillary electrophoresis single-strand conformation polymorphism (CE-SSCP) to screen allelic diversity in the above mentioned genes. Amplification was carried out using the Multiplex PCR kit (Qiagen) following the manufacturer’s instructions in a final reaction volume of 10 μl. Forward primers for both gene segments were 6’FAM-labelled and reverse primers were NED-labelled.
We selected individuals with diverse SSCP patterns, representing all identified alleles, to investigate sequence variation at both DQA and DQB loci, respectively. The genes were amplified using the same procedure as in Goüy de Bellocq et al.42 and Smith et al.43, but using non-labelled primers. As homozygous states were detected by SSCP analysis for all alleles in both gene segments, we directly sequenced the PCR product of the corresponding samples. Among all currently analysed samples, only eight were not screened with SCCP. These eight samples were directly sequenced using the same primers and the program phase64 was used to separate alleles in heterozygous samples.
For comparison between MHC and neutral genetic variation, genotype data of 14 microsatellite loci of 134 individuals out of the specimens analysed in the current study were used from a previous study38.
Data analysis
Population genetic parameters
Sequences were edited and aligned by eye using the BioEdit v.7.2.5 program©, 1997-201365. Allele frequencies, mean number of alleles (A), observed (Ho) and expected (He) heterozygosity were calculated for each locus and for each regional population (NT, CT, ST) with GENETIX66. Tests of deviation from Hardy–Weinberg equilibrium and linkage disequilibrium were calculated using GENEPOP 4.067 and GENETIX66, respectively. The allelic numbers for the cape hare sequences were assigned according to the guidelines of Klein et al.68 and begin at Lecp-DQA*04 and Lecp-DQB*01 for DQA and DQB, respectively. The FSTAT vers. 2.9.3.2. program69 was used to calculate locus-specific values of allelic richness (Rs) based on a rarefaction approach to account for different sample sizes. Allelic richness calculation was based on minimum sample size of 29 and 18 individuals for MHC and microsatellite loci, respectively.
Differentiation between regional populations was determined by calculation of standardized pairwise FST (10 000 permutations) in GENETIX66 and also Jost’s D (Dest)70 as a more robust measure than FST using GenAlEx 6.571. In addition, an analysis of molecular variance (AMOVA) from the MHC data was calculated for the three eco-regions (NT, CT, ST) using the Arlequin 3.5 program72. The same genetic parameters were also estimated for genotypes at 14 microsatellite loci obtained earlier for the same individuals38 for NT, CT, ST.
Furthermore, in order to identify the contribution of the different MHC alleles to the population structure, a principal components analysis (PCA) was performed in ADE-473, using the genotype data at both MHCII loci. Coordinates obtained for each allele were used for scatter plots using Past software74.
Finally, as demographic events might have affected the observed pattern of diversity and differentiation in MHC loci, we first use the “Wilcoxon signed-ranks test”75 implemented in the bottleneck software to test for bottleneck in the microsatellite data. As recommended by Piryet al.75, we used the Two Phase Mutation model (TPM) and the Stepwise Mutation Model (SMM).We then calculated effective population size using the neutral microsatellite data using Ne estimator76. Three different methods based on the information of linkage disequilibrium (LD), heterozygote excess (HE) and molecular coancestry (MC) were used.
Recombination analysis
The assumptions of several methods of sequence analysis are violated if recombination is present77. To check for the presence of recombination in the alignment of our sequences, we first used PERMUTE (included in the software package OMEGAMAP78), which computes three different statistics (r2, D, and G4) based on the correlation between physical distance of sites and their linkage disequilibrium. The RDP, GENECONV, MAXCHI, CHIMAERA and 3SEQ implemented in the RDP3 package79 were all employed to detect recombination breakpoint locations. In the latter analyses we used a significance level of 0.05. We also calculated the population scaled recombination rate ρ and mutation rate θ by using LDhat recombination rate scan implemented in theRDP3 package79. Finally, the GARD method80 was employed to search for possible recombination partitions.
Detecting positive selection
In order to infer evidence of positive selection, three different methods that compare the rates of synonymous and non-synonymous substitutions separately for every single codon were used. We chose to use HYPHY81 as implemented in the DATAMONKEY webserver (http://www.datamonkey.org/83; last accessed 30th January 2017), OMEGAMAP78 and CODEML (PAML 4 package82). The first two software packages have the important advantage of taking recombination into account whereas CODEML does not account for that.
The OmegaMap program is based on a Bayesian population genetics approximation to the coalescent theory and generates means and credible intervals for the selection parameter (dN/dS = ω) and recombination rate (ρ = 4Nr) for each codon (N and r represent the effective population size and the per codon rate of recombination, respectively). We used the same parameters as in Smith et al.43. Two Markov chain Monte Carlo runs of 250 000 iterations (25 000 iteration burn-in) for DQA and 400 000 iterations (40 000 iteration burn-in) for DQB on population allele frequencies at each locus, were compared for convergence. Codons are considered as positively selected with posterior probabilities greater than 95%.
In DATAMONKEY, we first identified recombination break points with genetic algorithm recombination detection (GARD80) that infers phylogenies for each putative non recombinant fragment. The output was used to run four different maximum likelihood methods for detection of selection: SLAC (single likelihood ancestral counting), FEL (fixed effects likelihood), REL (random effects likelihood), and Mixed Effects Model of Evolution (MEME)83. Significance levels of P < 0.25 in SLAC and FEL and P < 0.05 in MEME and Bayes factors >50 in REL were considered as indicating positively selected sites. We considered a codon to be positively selected only if it was identified by at least two of the methods implemented77,84.
Several studies42,85,86 that used CODEML to test for site specific positive selection in MHC data have shown its robustness although it does not account for recombination events. Two pairs of models were compared using this program: M1a (neutral model) was compared to M2a (adaptive model) and M7 (beta) was compared to M8 (beta plus omega). Pairwise comparisons on nested models were performed using the likelihood ratio test (LRT): twice the log-likelihood difference was compared with a χydistribution with degrees of freedom equal to the difference in the number of parameters between both models. When the LRT was significant, a Bayes Empirical Bayes (BEB) method was used to calculate the posterior probabilities of codon classes in models M2a and M8. Posterior probabilities of >0.95 were considered as supportive under the BEB method87. We considered a codon to be positively selected if it was identified by M2a or M8 or both.
In addition to the applied tests to infer molecular signature of positive selection, we used the model-based approach of Beaumont and Nichols35, based on MHC genotypes, implemented in Lositan88 to compare the observed FST values estimated at each locus to a null distribution of FST conditional on heterozygosity. DQA, DQB and fourteen microsatellite loci38 were tested for neutrality under 50000 simulations, estimated neutral mean FST, infinite alleles mutation model, 99% confidence interval and false discovery rate of 0.1%.
Testing for variation of genetic diversity between regions and for effects of climate variables on MHC alleles and heterozygosity
First, we used locus specific allelic richness of MHC and microsatellite loci in the three regional populations to run a linear mixed effects model (lmer) to check for significant variation of allelic richness from North to South Tunisia. Under a positive selection scenario for MHC loci we expected that allelic richness at both MHC loci varies between sampling regions in the case of different pathogen pressure. However, genetic diversity (allelic richness) for selectively neutral markers like microsatellites was not expected to vary between populations, as population genetic results indicated high gene flow across the whole study region38,39,40. Thus, with a hypothesized higher pathogenic diversity in the more climatically diverse NT population, MHC diversity at the two studied loci was expected to exceed the neutral microsatellite diversity particularly in NT. In the model we used locus specific allelic richness as obtained by FSTAT as dependent variable, regional population (NT, CT, ST) and locus type (MHC or microsatellite) as fixed factors and locus as random variable to account for potential locus-specific effects (particularly among the microsatellites) and specifically tested for a significant locus type by population interaction effect. The model syntax was as follows: lmer (random effect: ~ 1I locus; locus specific allelic richness ~ population * locus type). We also tested if allelic richness varies across the ecological gradient independently from microsatellite diversity using a general linear model of the syntax: MHC allelic richness ~ region; where MHC allelic richness means locus-specific allelic richness (for DQA and DQB) and region means NT or (CT & ST). CT and ST populations were combined because allelic richness values (see Table 1) were higher in NT but similarly lower in CT and ST.
Second, we used the statistical software package R 2.15.0 (R Development Core Team, 2011) to run multinomial log-linear models separately for the most frequent protein variants at the DQA (07, 08, 11) and the DQB (01, 03, 09) loci as response variables and geographical latitude, longitude, altitude as well as the protein combination at the respective co-occurring MHC locus as independent variables. However, mean annual temperature (r = −0.796) and annual precipitation (r = 0.831) correlated too closely with latitude to add them in the model, and latitude has the clearest effect of these three variables on the response variable (see Supplementary Table S5). Thus, hereafter latitude will be interpreted as surrogate for climate, acknowledging that other abiotic factors like soil or wetness also might be covered by this variable. Mean annual temperature, annual precipitation and altitude were obtained from WORLDCLIM data set for 2.5 min intervals (Version 1.4, http://www.worldclim.org/bioclim.htm) and were automatically extracted using DIVA-GIS ver. 7.5. We also run a multinomial log-linear model with individual heterozygosity in DQA and DQB loci as response variable and the same geographic variables as independent variables, expecting a higher level in NT (in parallel to allelic richness).
The model syntaxes were as follows (using the package nnet):
-
1)
Multinom (DQA protein class ~ latitude + longitude + altitude + as.factor (co-occurring DQB protein class)),
where the two co-occurring protein classes represented either any DQB protein combination of the most prevalent proteins 01, 03, 09, on the one hand and any other protein combination on the other,
-
2)
Multinom (DQB protein class ~ latitude * longitude + altitude + as.factor (co-occurring DQA protein class)),
where the two co-occurring protein classes represented either any DQA protein combination of the most prevalent proteins 07, 08, 11 on the one hand and any other protein combination on the other,
-
3)
Multinom (MHC heterozygosity ~ latitude * longitude + altitude).
We used an information-theoretic approach and techniques of model selection and multi-model inference36 (i.e., model ranking and model averaging) to obtain the best model explaining the data. Based on the global models (see syntaxes above), we run all possible models including the respective null-models and calculated the probability of each possible model being the best model using the Akaike weight based on AICc values (i.e., corrected for small sample sizes). Adding up the weight of all models in which a given variable is present gives the “Relative Variable Importance”, RVI, Table 5). RVI is an estimate of the absolute probability for a variable of being in the best model explaining this particular dataset. For each variable we also calculated the so-called delta deviance, i.e. the difference in deviance between the full model and the model after dropping this variable. In the cases of completely uncorrelated variables (which is almost the case here), delta deviance gives a relative estimate on the effect strength of one variable to the other, e.g. the effect of latitude on dqb3gt is 49.7/1.5 ≈ 32 times stronger than the effect of the respective co-occuring dqa genotype. Additionally we report the estimates, standard errors and the upper and lower 95% confidence interval (see Supp2), which indicates the precision of the estimate. We do not use these outputs to interpret the importance of the variables, because this would almost like using p-values in a Null-hypothesis testing framework, which should be avoided in an information-theoretic approach36. For each model we report delta AICc (difference in AICc between the best model and the intercept-only model) and the explained deviance as measure of goodness of fit (Table 5).
Accession codes
Sequence data from this article can be found in the GenBank under the accession numbers: MH346126-MH346168.
References
Sommer, S., Courtiol, A. & Mazzoni, C. J. MHC genotyping of non-model organisms using next‐generation sequencing: a new methodology to deal with artefacts and allelic dropout. BMC Genomics 14, 542–559 (2013).
Zhang, M. & He, H. Parasite-mediated selection of major histocompatibility complex variability in wild brandt’s voles (Lasiopodomys brandtii) from Inner Mongolia, China. BMC Evol. Biol. 13, 149 (2013).
Cohen, S. Strong positive selection and habitat-specific amino acid substitution patterns in MHC from an estuary fish under intense pollution stress. Mol. Biol. Evol. 19, 1870–1880 (2002).
Gillingham, M. A. F. et al. Very high MHC Class IIB diversity without spatial differentiation in the mediterranean population of greater Flamingos. BMC Evol. Biol. 17, 56 (2017).
Ben Slimen, H., Schaschl, H., Knauer, F. & Suchentrunk, F. Selection on the mitochondrial ATP synthase 6 and the NADH dehydrogenase 2 genes in hares (Lepus capensis L., 1758) from a steep ecological gradient in North Africa. BMC Evolutionary Biology 17, 46 (2017).
Klein, J. Natural History of the Major Histocompatability Complex. John Wiley and Sons: New York (1986).
Weber, D. S., Stewart, B. S., Schienman, J. & Lehman, N. Major histocompatibility complex variation at three class II loci in the northern elephant seal. Mol. Ecol. 13, 711–8 (2004).
Koutsogiannouli, E. A. et al. Major histocompatibility complex variation at class II DQA locus in the brown hare (Lepus europaeus). Mol. Ecol. 18, 4631–4649 (2009).
Balasubramaniam, S. et al. New data from basal Australian songbird lineages show that complex structure of MHC class II β genes has early evolutionary origins within passerines. BMC Evol. Biol. 16, 1–11 (2016).
Kohn, M. H., Murphy, W. J., Ostrander, E. A. & Wayne, R. K. Genomics and conservation genetics. Trends Ecol. Evol. 21, 629–637 (2006).
Bonin, A., Nicole, F., Pompanon, F., Miaud, C. & Taberlet, P. Population adaptive index: A new method to help measure intraspecific genetic diversity and prioritize populations for conservation. Conserv. Biol. 21, 697–708, https://doi.org/10.1111/j.1523-1739.2007.00685.x (2007).
Bonhomme, M., Blancher, A., Jalil, M. F. & Crouau-Roy, B. Factors shaping genetic variation in the MHC of natural non human primate populations. Tissue Antigens 70, 398–411 (2007).
Vassilakos, D., Natoli, A., Dahlheim, M. & Hoelzel, A. R. Balancing and directional selection at exon-2 of the MHC DQB1 locus among populations of odontocete cetaceans. Mol. Biol. Evol. 26, 681–689 (2009).
Baker, C. S. et al. Diversity and duplication of DQB and DRB-like genes of the MHC in baleen whales (suborder: Mysticeti). Immunogenetics 58, 283–296 (2006).
Xu, S. X. et al. Sequence polymorphism and evolution of three cetacean MHC genes. J. Mol. Evol. 69, 260–275 (2009).
Moreno-Santillán, D. D., Lacey, E. A., Gendron, D. & Ortega, J. Genetic variation and gene duplication at exon 2 of the MHC class II DQB locus in Blue Whale (Balaenoptera musculus) from the Gulf of California. PLoS ONE 11, e0141296 (2016).
Brunner, F. & Eizaguirre, C. Can environmental change affect host/parasite-mediated speciation? Zoology 119, 384–394 (2016).
Wegner, K. M., Kalbe, M., Milinski, M. & Reusch, T. B. H. Mortality selection during the 2003 European heat wave in three-spined sticklebacks: effect of parasites and MHC genotype. BMC. Evol. Biol. 8, 1–12 (2008).
Björklund, M., Aho, T. & Behrmann-Godel, J. Isolation over 35 years in a heated biotest basin causes selection on MHC class IIß genes in the European perch (Perca fluviatilis L.). Ecol. Evol. 5, 1440–55 (2015).
Klein, J. & Figueroa, F. The evolution of class I MHC genes. Immunol.Today 7, 41–44 (1986).
Hughes, A. L. & Nei, M. Evolutionary relationships of class II major-histocompatibility-complex genes in mammals. Mol. Biol. Evol. 7, 491–514 (1990).
Hughes, A. L. & Yeager, M. Natural selection at major histocompatibility complex loci of vertebrates. Annu. Rev. Genet. 32, 415–435 (1998).
Ottaviani, D. et al. Reconfiguration of genomic anchors upon transcriptional activation of the human major histocompatibility complex. Genome Res. 18, 1778–1786 (2008).
Sommer, S. The importance of immune gene variability (MHC) in evolutionary ecology and conservation. Front Zool. 2, 16 (2005).
Yeager, M. & Hughes, A. L. Evolution of the mammalian MHC: natural selection, recombination, and convergent evolution. Immunol. Rev. 167, 45–58 (1999).
Ohta, T. Gene conversion vs point mutation in generating variability at the antigen recognition site of major histocompatibility complex loci. J. Mol. Evol. 41, 115–119 (1995).
Miller, H. C. & Lambert, D. M. Gene duplication and gene conversion in class II MHC genes of New Zealand robins (Petroicidae). Immunogenetics 56, 178–191, https://doi.org/10.1007/s00251-004-0666-1 (2004).
Spurgin, L. & Richardson, D. How pathogens drive genetic diversity: MHC, mechanisms and misunderstandings. Proc R Soc B 277, 979–988 (2010).
Woolhouse, M. E. J., Webster, J. P., Domingo, E., Charlesworth, B. & Levin, B. R. Biological and biomedical implications of the co-evolution of pathogens and their hosts. Nat. Genet. 32, 569–577 (2002).
Gandon, S., Capowiez, Y., Dubois, Y., Michalakis, Y. & Olivieri, I. Local adaptation and gene-for-gene coevolution in a metapopulation model. Proc. R. Soc. B, Biol. Sci. 263, 1003–1009 (1996).
Teacher, A. G. F., Garner, T. W. J. & Nichols, R. A. Evidence for directional selection at a novel major histocompatibility class I marker in wild common frogs (Ranatemporaria) exposed to a viral pathogen (Ranavirus). PLoS ONE 4, e4616 (2009).
Landry, C. & Bernatcez, L. Comparative analysis of population structure across environments and geographical scales at major histocompatibility complex and microsatellite loci in Atlantic salmon (salmo salar). Mol. Ecol. 10, 2525–2539 (2001).
Schierup, M. H. The number of self-incompatibility alleles in a finite, subdivided population. Genetics 149, 1153–1162 (1998).
Knafler, G. J., Grueber, C. E., Sutton, J. T. & Jamieson, I. G. Differential patterns of diversity at microsatellite, MHC, and TLR loci in bottlenecked South Island saddleback populations. New Zealand Journal of Ecology 41, 98–106 (2017).
Beaumont, M. A. & Nichols, R. A. Evaluating loci for the use in the genetic analysis of population structure. Proc. R. Soc. B. 263, 1619–1636 (1996).
Burnham, K. P. & Anderson, D. R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach 2nd ed. Springer. NewYork (2002).
Klein, J., Satta, Y., O’hUigin, C. & Takahata, N. The molecular descent of the major histocompatibility complex. Annu. Rev. Immunol. 11, 269–295 (1993).
Ben Slimen, H. Phylogénie morphologique et moléculaire des lièvres d’Afrique du Nord du genre Lepus. PhD thesis, Faculty of Sciences of Tunis (2008).
Awadi, A., Suchentrunk, F., Makni, M. & Ben Slimen, H. Phylogenetic relationships and genetic diversity of Tunisian hares (Lepus sp. or spp., Lagomorpha) based on partial nuclear gene transferrin sequences. Genetica 144, 497–512 (2016).
Ben Slimen, H., Suchentrunk, F., Memmi, A. & Ben AmmarElgaaied, A. Biochemical genetic relationships among Tunisian hares (Lepus sp.), South African cape hares (L. capensis), and European brown hares (L. europaeus). Biochem.Genet. 43, 577–96 (2005).
Campos, J. L., Bellocq, J. Gd., Schaschl, H. & Suchentrunk, F. MHC class II DQA gene variation across cohorts of brown hares (Lepus europaeus) from eastern Austria: testing for different selection hypotheses. Mamm.Biol. 76, 251–257 (2011).
Goüy de Bellocq, J., Suchentrunk, F., Baird, S. & Schaschl, H. Evolutionary history of an MHC gene in two leporid species: characterisation of Mhc-DQA in the European brown hare and comparison with the European rabbit. Immunogenetics 61, 131–144 (2009).
Smith, S., Goüy de Bellocq, J., Suchentrunk, F. & Schaschl, H. Evolutionary genetics of MHC class II beta genes in the brown hare, Lepus europaeus. Immunogenetics 63, 743–751 (2011).
Klein, J. Origin of major histocompatibility complex polymorphism—The trans-species hypothesis. Hum. Immunol. 19, 155–162 (1987).
Takahata, N., Satta, Y. & Klein, J. Polymorphism and balancing selection at major histocompatibility complex loci. Genetics 130, 925–938 (1992).
Wegner, K. M. Historical and contemporary selection of teleost MHC genes: did we leave the past behind? Journal of Fish Biology 73, 2110–2132 (2008).
Ben Slimen, H., Suchentrunk, F., Shahin, A. B. & Ben Ammar Elgaaied, A. Phylogenetic analysis of mtCR-1 sequences of Tunisian and Egyptian hares (Lepus sp. or spp., Lagomorpha) with different coat colours. Mamm. Biol. 72, 224–239 (2007).
Schaschl, H. et al. Sex-specific selection for MHC variability in Alpine chamois. BMC Evolutionary Biology 12, 20 (2012).
Hill, R. E. & Hastie, N. D. Accelerated evolution in the reactive center regions of serine protease inhibitors. Nature 326, 96–99 (1987).
Promerová, M. et al. Evaluation of two approaches to genotyping major histocompatibility complex class I in a passerine-CE-SSCP and 454 pyrosequencing. Mol. Ecol. Res. 12(2), 285–92 (2012).
Klein, J. & Horejsi, V. Immunology. Blackwell Science Ltd. Oxford. UK (1997).
Zvinorova, P. I. et al. Prevalence and risk factors of gastrointestinal parasitic infections in goats in low-input low-output farming systems in Zimbabwe. Small Rumin.Res. 143, 75–83 (2016).
Pandey, V., Ndao, M. & Kumar, V. Seasonal prevalence of gastrointestinal nematodes in communal land goats from the Highveld of Zimbabwe. Vet. Parasitol. 51, 241–248 (1994).
Soberon, J. & Ceballos, G. Species Richness and Range Size of the Terrestrial Mammals of the World: Biological Signal within Mathematical Constraints. PLoS ONE 6, e19359 (2011).
Tallmon, D. A., Luikart, G. & Beaumont, M. A. Comparative evaluation of a new effective population size estimator based on approximate Bayesian computation. Genetics 167, 977–988 (2004).
Wang, J. A comparison of single-sample estimators of effective population sizes from genetic marker data. Molecular Ecology 25(19), 4692–4711 (2016).
Frankham, R., Ballou, J. D. & Briscoe, D. A. Introduction to Conservation Genetics. Cambridge University Press, Cambridge (2002).
Oliver, M. K. & Piertney, S. B. Selection maintains mhc diversity through a natural population bottleneck. Mol. Biol. Evol. 29, 1713–1720 (2012).
Flux, J. E. C. & Angermann, R. Hares and Jackrabbits inRabbits, Hares and Pikas. Status Survey and Conservation Action Plan (eds Chapman, J. A. & Flux, J. E. C.) 61–94. Information Press. Oxford. UK (1990).
Ben Slimen, H. et al. Population genetics of cape and brown hares (Lepus capensis and L. europaeus): a test of Petter´s hypothesis of conspecificity. Biochem.Syst. Ecol. 36, 22–39 (2008).
Hoffmann, R. S. & Smith, A. T. Order Lagomorpha in Mammal Species of the World 3 rd ed. (eds Wilson, D. E. & Reeder, D. M.) 185–211. The Johns Hopkins University Press. Baltimore (2005).
You, H. et al. Plant diversity in different bioclimatic zones in Tunisia. Journal of Asia-Pacific Biodiversity 9, 56–62 (2016).
Zahran, M. A. Climate - Vegetation: Afro-Asian Mediterranean and Red Sea Coastal Land. Springer Verlag. Dordrecht (2010).
Stephans, M. & Donelly, P. A. comparison of bayesian methods for haplotype reconstruction from population genotype data. Am. J. Hum. Genet. 73, 1162–1169 (2003).
Hall, T. A.BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids.Symp. Ser. 41, 95–98 (1999).
Belkhir, K., Borsa, P., Chikhi, L., Raufaste, N. & Bonhomme, F. GENETIX 4.05, logiciel sous Windows TM pour la génétique des populations. LaboratoireGénome, Populations, Interactions, CNRS UMR 5171.Université Montpellier 2. Monpellier. France (1996–2004).
Rousset, F. GENEPOP'007: A complete re-implementation of the GENEPOP software for Windows and Linux. Mol. Ecol. Resour. 8, 103–106 (2008).
Klein, J. et al. Nomenclature for the major histocompatibility complexes of different species: a proposal. Immunogenetics 31, 217–9 (1990).
Goudet, J. FSTAT, version 2.9. 3, A program to estimate and test gene diversities and fixation indices. Lausanne University, Lausanne, Switzerland (2001).
Jost, L. GST and its relatives do not measure differentiation. Mol. Ecol. 17, 4015–4026 (2008).
Peakall, R. & Smouse, P. E. GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 28, 2537–2539 (2012).
Excoffier, L. & Lischer, H. E. L. Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 10, 564–567 (2010).
Thioulouse, J., Chessel, D., Dolédec, S. & Olivier, J. Ade-4: a multivariate analysis and graphical display software. Statistics and Computing 7, 75–83 (1997).
Hammer, Ø., Harper, D. A. T. & Ryan, P. D. PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4(1), 9 (2001).
Piry, S., Luikart, G. & Cornuet, J. M. BOTTLENECK: a computer program for detecting recent reductions in the effective population size using allele frequency data. Journal of Heredity 90, 502–503 (1999).
Do, C. et al. Ne Estimator V2: reimplementation of software for the estimation of contemporary effective population size (Ne) from genetic data. Molecular Ecology Resources 14, 209–214 (2014).
Garrigan, D. & Hedrick, P. W. Perspective: detecting adaptive molecular polymorphism: lessons from the MHC. Evolution 57, 1707–1722 (2003).
Wilson, D. J. & McVean, G. Estimating diversifying selection and functional constraint in the presence of recombination. Genetics 172, 1411–1425 (2006).
Martin, D. P. et al. RDP3: A flexible and fast computer program for analyzing recombination. Bioinformatics 26, 2462–2463, https://doi.org/10.1093/bioinformatics/btq467 (2010).
Pond, S. L. K., Posada, D., Gravenor, M. B., Woelk, C. H. & Frost, S. D. W. Automated phylogenetic detection of recombination using a genetic algorithm. Mol. Biol. Evol. 23, 1891–1901 (2006).
Kosakovsky, P. S. L., Frost, S. D. W. & Muse, S. V. HyPhy: hypothesis testing using phylogenies. Bioinformatics 21, 676–9 (2005).
Yang, Z. PAML 4: phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 24, 1586–1591 (2007).
Murrell, B. et al. Detecting individual sites subject to episodic diversifying selection. PLoS Genet. 8, e1002764 (2012).
Kosakovsky, P. S. L. & Frost, S. D. W. Datamonkey: rapid detection of selective pressure on individual sites of codon alignments. Bioinformatics 21, 2531–3 (2005).
Consuegra, S. et al. Rapid Evolution of the MH Class I Locus Results in Different Allelic Compositions in Recently Diverged Populations of Atlantic Salmon. Mol. Biol.Evol. 22, 1095–106 (2005).
Kuduk, K. et al. Evolution of major histocompatibility complex class I and class II genes in the brown bear. BMC Evol. Biol. 12, 197–10; 1186/1471-2148-12-197 (2012).
Yang, Z., Nielsen, R., Goldman, N. & Pedersen, A. M. Codon-substitution models for heterogeneous selection pressure at amino acid sites. Genetics 155, 431–449 (2000).
Antao, T., Lopes, A., Lopes, R. J., Beja-Pereira, A. & Luikart, G. LOSITAN: a workbench to detect molecular adaptation based on a Fst-outlier method. BMC Bioinformatics 9, 323, https://doi.org/10.1186/1471-2105-9-323 (2008).
Bondinas, G., Moustakas, A. & Papadopoulos, G. The spectrum of HLA-DQ and HLA-DR alleles, 2006: a listing correlating sequence and structure with function. Immunogenetics 59, 539–553 (2007).
Acknowledgements
We thank A. Haiden for supporting laboratory work. We thank Dr. Mark Gillingham (University of Ulm) and an anonymous reviewer for their comments and suggestions, which helped us to improve the manuscript.
Author information
Authors and Affiliations
Contributions
F.S. and A.A. conceived the experiments. A.A. and S.S. conducted the experiments. F.S., A.A., S.S. and F.K. analyzed the results. A.A. and H.B.S. wrote the paper. F.S. and M.M. reviewed the manuscript. All authors approved the final manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Awadi, A., Ben Slimen, H., Smith, S. et al. Positive selection and climatic effects on MHC class II gene diversity in hares (Lepus capensis) from a steep ecological gradient. Sci Rep 8, 11514 (2018). https://doi.org/10.1038/s41598-018-29657-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-29657-3
This article is cited by
-
Patterns of evolution in MHC class II DQA and DQB exon 2 genes of Alpine mountain hares, Lepus timidus varronis, and sympatric and parapatric brown hares, L. europaeus, from Switzerland
Immunogenetics (2024)
-
Diversity and evolution of the MHC class II DRB gene in the Capra sibirica experienced a demographic fluctuation in China
Scientific Reports (2023)
-
Selection and demography drive range-wide patterns of MHC-DRB variation in mule deer
BMC Ecology and Evolution (2022)
-
Spatial diversity of MHC class II DRB exon2 sequences in North African cape hares (Lepus capensis): positive selection and climatic adaptation signals
Mammalian Biology (2022)
-
Positive selection on the MHC class II DLA-DQA1 gene in golden jackals (Canis aureus) from their recent expansion range in Europe and its effect on their body mass index
BMC Ecology and Evolution (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
