
Abstract
Upregulation of B-cell CLL/lymphoma (BCL)2 expression following lithium treatment is seemingly well established and has been related to the neuroprotective property of the drug. However, while demonstrated by some (but not all) studies based on low-throughput techniques (e.g. qPCR) this effect is not reflected in high-throughput studies, such as microarrays and RNAseq. This manuscript presents a systematic review of currently available reports of lithium’s effect on BCL2 expression. To our surprise, we found that the majority of the literature does not support the effect of lithium on BCL2 transcript or protein levels. Moreover, among the positive reports, several used therapeutically irrelevant lithium doses while others lack statistical power. We also noticed that numerous low-throughput studies normalized the signal using genes/proteins affected by lithium, imposing possible bias. Using wet bench experiments and reanalysis of publicly available microarray data, here we show that the reference gene chosen for normalization critically impacts the outcome of qPCR analyses of lithium’s effect on BCL2 expression. Our findings suggest that experimental results might be severely affected by the choice of normalizing genes, and emphasize the need to re-evaluate stability of these genes in the context of the specific experimental conditions.
Similar content being viewed by others

Introduction
Upregulation of B-cell CLL/lymphoma (BCL)2 transcript following chronic lithium treatment is considered to be well established1. Chen et al. and Chen and Chuang2,3 were the first to report lithium-induced increased gene and protein expression of BCL2 following lithium treatment. Due to the anti-apoptotic effect of BCL24 this upregulation has been interpreted as a possible mechanism underlying at least in part the neuroprotective property of the drug (reviewed in5).
However, BCL2’s function is not restricted to prevention of apoptosis. In the endoplasmic reticulum (ER) BCL2 is a potent inhibitor of autophagy6, a vital cellular process shown to be augmented by lithium7. Indeed, we have found that lithium-treated mice exhibit changes in protein levels of autophagy markers (upregulation of Beclin-1 and downregulation of p62) indicative of upregulation of this process8. Thus, the well accepted lithium-induced upregulation of BCL2, while corroborating other pro cell-survival effects of the drug, contradicts its pro-autophagy effect. Moreover, the lack of evidence for increased risk of cancer or poorer malignancy prognosis among lithium-treated subjects9,10 is counterintuitive with the anti-apoptotic consequence of increased BCL2 expression4,11 and with the positive correlation between malignancy and BCL2 expression12,13.
We have recently performed a real-time PCR quantification of the expression levels of several genes previously reported to be affected by lithium in hippocampi of chronically lithium-treated mice and of inositol monophosphotase-1 (IMPase-1, a lithium-inhibitable enzyme at therapeutically-relevant concentrations, encoded by IMPA1) homozygote knockout (KO) mice14. BCL2 was one of the genes examined in this study. Guided by a recent study demonstrating that stability of commonly used normalizing genes (e.g. GAPDH, ACTB) is affected by mood-stabilizer treatment, we used MAPK6 to normalize our data. MAPK6 was recommended as a brain-reference gene15, and we validated its stability using unpublished data from our microarrays study of similar lithium treatment conditions (i.e. regimen, tissue and mouse strain)16. Surprisingly, we observed significantly decreased rather than increased BCL2 mRNA levels, both following lithium treatment and in IMPA1 KO mice. In the search for a plausible explanation for our finding we carefully reviewed studies in which lithium’s effect on BCL2 transcript and protein levels was assessed. As summarized in Table 1, a laborious screening of previous studies revealed that upregulation of BCL2 by lithium is far from being robust. Rather, BCL2 upregulation was not observed in any of the microarrays studies we screened17,18,19,20,21,22. We have also noticed that in some of the studies supporting lithium-induced upregulation of BCL2 the reference genes used for normalization have previously been reported to be affected by lithium (e.g. ACTB, GAPDH, TUBB, Tables 1 and 2).
We therefore hypothesized that the discrepancy between microarrays and qPCR studies may, in part, result from the need to normalize qPCR expression using a chosen reference gene. To test our hypothesis we assessed the effect of different normalizing genes on lithium-induced BCL2 expression measured using qPCR. To further validate our results we also assessed the impact of normalizing genes on the expression of another gene known to be affected by lithium - Myristoylated Alanine Rich Protein Kinase C Substrate (MARCKS)17,23,24,25.
Results
The effect of lithium on BCL2 and MARCKS expression levels when normalized to ACTB
BCL2 levels normalized to β-Actin were higher in the lithium treatment group than in the regular food group [regular food vs. lithium treatment, median (interquartile): 0.75 (0.58, 1.17) vs. 1.01 (0.89, 1.48), location shift (95%CI): 0.28 (0.05 to 0.63), p = 0.023, Fig. 1A]. No significant difference was found in normalized MARCKS levels [regular food vs. lithium treatment: 0.9 (0.80, 1.05) vs. 1.14 (0.91, 1.59), location shift (95%CI): 0.23 (−0.01 to 0.65), p = 0.081, Fig. 1D].

The direction of the observed effect of lithium treatment on gene expression depends on the normalizing gene. Violin plots of hippocampal BCL2 (upper lane) and MARCKS (lower lane) mRNA levels of control mice and of chronic lithium-treated mice, normalized to different normalizing genes. When normalized to β-Actin (A,D), both BCL2 and MARCKS appear to be upregulated by lithium treatment. However, following normalization to MAPK6 (B,E), both transcripts appear to be downregulated by lithium treatment. Normalization to ANKRD11 (C,F) supports the results obtained following normalization to MAPK6. RF – regular food; Li – lithium treatment. Each point represents an individual normalized value, Boxplots show the median and the interquartile range. p-values were calculated using two-sided Wilcoxon rank sum test. *p < 0.05.
The effect of lithium on BCL2 and MARCKS expression levels when normalized to MAPK6
BCL2 levels normalized to MAPK6 were lower in the lithium treatment group than in the regular food group [regular food vs. lithium treatment: 0.98 (0.87, 1.30) vs. 0.82 (0.72, 1.00), location shift (95%CI): −0.21 (−0.44 to −0.02), p = 0.027, Fig. 1B]. Similarly, MARCKS levels normalized to MAPK6 were lower in the lithium treatment group than in the regular food group [regular food vs. lithium treatment: 1.17 (0.93, 1.24) vs. 1.01 (0.69, 1.18), location shift (95%CI): −0.18 (−0.44 to 0.007), p = 0.050, Fig. 1E].
Estimation of the expression stability of genes in mouse hippocampus
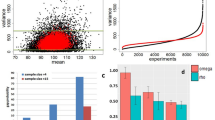
We utilized the RefGenes tool of the Genevestigator software that ranks genes based on the variance in their expression in a chosen set of samples from microarrays database. We searched for the most stable genes in the hippocampus of wildtype (WT) mice and estimated the stability of BCL2 and three reference genes: ACTB, GAPDH and MAPK6. As reflected by the standard deviation (SD) of the log2 transformed signals (Fig. 2) the expression stability of β-actin, GAPDH and MAPK6 in WT untreated mice is similar to that of BCL2 (SD: 0.4, 0.51, 0.5 and 0.48, respectively).

Expression stability of β-actin, GAPDH and MAPK6 in hippocampus of WT-untreated mice is similar to that of BCL2. A snapshot obtained using the RefGenes tool of the Genevestigator software. Based on 80 microarrays of hippocampal samples of WT untreated mice the signal variability of the commonly used normalizing genes is similar to that of BCL2, as can be noticed from the SD of the log2 transformed signals. The box-plots represent the ranges of the log2-transformed signals of the suggested reference genes (light blue), the commonly used reference genes (GAPDH, β-actin and MAPK6 - dark blue, red and green, respectively) and of BCL2 (orange). Asterisks indicate outliers’ expression values.
The effect of lithium on BCL2 and MARCKS expression levels when normalized to ANKRD11
We used the list of the most stable reference genes in mouse hippocampus obtained from the RefGenes tool (Fig. 2) to select an additional normalizing gene for real-time PCR analysis of BCL2 and MARCKS. We further evaluated the stability of the top suggested reference genes in our (unpublished) microarrays data of Toker et al.16. Among these genes ANKRD11 was the most stable gene that was not significantly affected by lithium treatment or IMPA1 KO in our microarray study (lithium treated WT mice from the SMIT1 colony: p > 0.8; lithium treated WT mice from the IMPA1 colony: p > 0.14). In agreement with the results obtained when normalized to MAPK6 (Fig. 1B,E), BCL2 and MARCKS levels normalized to ANKRD11 were lower in the lithium treatment group than is the regular food group: BCL2 - regular food vs. lithium treatment: 1.14 (1.00, 1.27) vs. 0.98 (0.85, 1.087), location shift (95%CI): −0.17 (−0.45 to 0.01), p = 0.067, Fig. 1C; MARCKS - regular food vs. lithium treatment: 1.19 (0.94, 1.38) vs. 0.9 (0.81, 1.06), location shift (95%CI): −27 (−0.57 to 0.11), p = 0.185, Fig. 1F.
Lithium’s effect on the expression of BCL2 and MARCKS based on publicly available microarrays datasets
Genes on microarray platforms are often represented by several probesets, targeting different parts of the gene (Table 3). None of the probesets representing the BCL2 gene was significantly affected by lithium treatment in the two analyzed datasets (non-adjusted p-value > 0.1, adjusted p-value > 0.5). The expression of MARCKS was significantly downregulated by lithium in the GSE35291 dataset (adjusted p-value = 5.5*10−3). None of the probesets corresponding to MARCKS in the GSE66277 dataset showed a significant effect of lithium after controlling for FDR (adjusted p-value > 0.2). However, in the hippocampal samples, in two out of the six probesets, MARCKS levels were downregulated (non-adjusted p-value < 0.025). Table 3 summarizes the results of differential analyses for all BCL2 and MARCKS probesets in the two datasets.
Discussion
In this study we show that conclusions of experiments assessing the effect of lithium treatment on BCL2 expression can noticeably be affected by different experimental set-ups. In qPCR experiments normalization with ACTB suggested significant upregulation of the gene, supporting the common notion of lithium’s treatment effect on this gene. Contrarily, normalization of the same samples with either MAPK6 or ANKRD11 suggested significant downregulation, contradicting the common notion. Alternatively, our analysis of publicly available microarray data from lithium treated rodents showed no apparent effect of the drug on BCL2 expression, in line with previous microarray studies of lithium’s effects17,18,19,20,21,22.
Our findings are surprising in light of the common notion that lithium treatment induces upregulation of BCL2 transcript and protein levels, interpreted to contribute to the neuroprotective effect of the drug. However, thorough examination of reported data reveals that the effect is not as robust as generally perceived. Thirty one out of 49 (63%) published experimental set-ups found no significant effect and only 15 supported significant upregulation of BCL2 transcript/protein. In four out of the latter 15 experimental set-ups the raw data was normalized to genes/proteins previously shown to be affected by lithium (Tables 1, 2). The remaining three experimental set-ups (out of the 49 ones) either did not provide information regarding the significance of the finding or reported downregulation of the gene.
There are several possible reasons for the discrepancies among the different studies, emphasizing the uncertainty in our knowledge of the truth regarding lithium’s effect on BCL2 transcript and/or protein levels.
Lithium treatment regime
The first to report lithium-induced BCL2 upregulation in rat brain were Chen and colleagues3. The effect was found following four weeks but not nine days of treatment with a high lithium dose − 4 meq/kg/day. The authors reported that this treatment resulted in therapeutically-relevant lithium blood levels. However, an earlier study using this dose26 reported dissimilar pharmacokinetics of the drug in plasma vs. brain during a prolonged treatment period (21 days); in plasma, after two weeks of treatment, the authors found a gradual decrease in the drug’s concentration. In contrast, in the brain, lithium levels increased continuously during the whole treatment period. This suggests that brain lithium levels in the Chen et al.’s study3 might have been higher than the therapeutic range of the drug. Lithium doses exceeding the range used for therapeutic purposes were used in several additional studies supporting lithium-induced upregulation of BCL2 transcript/protein27,28,29,30,31.
Possible misinterpretation of the findings
The possibility that the observed increase in BCL2 protein merely reflects lithium-induced cell proliferation32,33 cannot be ruled out. Based on their immunohistochemistry results, Chen et al.3 report “increased number of BCL2 positive cells” rather than increased BCL2 intensity. Interestingly, a similar treatment protocol was later used to study the effect of lithium on long-term potentiation (LTP) and neurogenesis30,31. The authors report 50% increase in newborn cells in the dentate gyrus, accompanied by ~40% increase in BCL2 protein levels in the same region. While a causative effect of BCL2 upregulation on cell proliferation/neurogenesis cannot be ruled out, Bernier and Parent34 have previously shown that BCL2 is a marker of immature neurons, which are likely to dominate in the dentate gyrus, where neurogenesis occurs.
An additional study frequently cited in support of lithium-induced upregulation of BCL2 examined the drug’s potency to protect chick cochlear nucleus neurons from deafferentiation-induced apoptosis35. In this model cessation of protein synthesis and upregulation of BCL2 transcript precede neuronal death36,37. As reported by Bush and Hyson35 chronic lithium treatment reduced cellular death and increased BCL2 protein levels in the deafferentiated cochlear nucleus. The authors speculated that increased BCL2 levels may have resulted from lithium-induced activation of specific transcription factors (TFs). Although generally plausible, regulation of TFs activity is unlikely to take place in a scenario of protein synthesis arrest. Rather, a more likely interpretation (in light of the increased BCL2 transcript in these cells prior treatment) is cellular recovery from cessation of protein synthesis as a result of lithium treatment. Indeed, it has been demonstrated that inhibition of protein synthesis in the deafferentiated cochlear nucleus results from phosphorylation of elongation factor 2 (eEF2)38,39 that can be reversed by lithium treatment40. It is thus conceivable that the observed lithium-induced upregulation of BCL2 in Bush and Hyson’s study35 does not reflect a specific upregulation of the BCL2 protein, but, rather, a general increase in protein synthesis.
The choice of the normalizing gene/protein
Among common normalizing genes MAPK6 has been suggested to be the most appropriate for brain qPCR analyses15,41. Other commonly used reference genes, for example GAPDH, ACTB, TUBA and TUBB have been repeatedly reported as altered in bipolar patients42,43,44, following lithium treatment17,18,45,46,47,48,49,50 or oxidative stress51. In addition, the tubulin proteins are involved in autophagy52,53, a cellular pathway known to be affected by lithium7. In our own microarrays and proteomics analyses (Table 2) mRNA and protein levels of GAPDH, β-Actin and α/β-Tubulin were affected in lithium-treated mice. Stability of different normalizing genes following acute and chronic treatment with mood stabilizers was recently evaluated by Powel et al.54. The authors show that neither of the genes shows stable expression following treatment with these drugs. Specifically, GAPDH and ACTB were among the least stable genes following chronic lithium treatment. Despite all of the above, in a substantial proportion of the reviewed studies data was normalized using genes/proteins shown to be affected by lithium treatment18,27,55,56,57,58.
Biased choice of authors which of the results to emphasize
A cautious analysis of Lowthert et al.’s report59 suggests that at least in some cases the authors only concentrate on favorable findings. Studying lithium-responsive and non-responsive bipolar patients the authors report upregulation of BCL2 in lithium-responders and downregulation in non-responders one month following treatment. In essence, they used microarrays to study peripheral blood gene expression in the patients over a two-month period. Blood samples were taken at baseline and every two weeks. Similarly to other microarrays studies of lithium’s effect17,18,19,20,21,22 the microarray data did not support BCL2 expression changes in any of the time points. Nevertheless, the authors performed qPCR analysis of the BCL2 gene family. No change was found in BCL2 expression neither in lithium-responders nor in non-responders as compared to untreated healthy subjects. The finding reported by the authors is based on an increased ratio of BCL2 transcript between lithium-responders and non-responders found at a single time point - one month after treatment initiation. This is regardless of the data showing that at the remainder of the time points (4 out of 5) BCL2 expression was similar or lower in the responders, both as compared to non-responders or to healthy subjects59.
Impact of normalizing genes/proteins in biological psychiatry
The discrepancy between microarray and qPCR findings and the contradicting results obtained by normalizing the same data using different ‘housekeeping’ genes raise the question regarding the impact of normalising genes on qPCR analyses in biological psychiatry. Several studies assessed the issue of “compatible” reference genes for qPCR, in various tissues, in general15,60,61, and for brain, in particular41,62. The take-home message of these and other tissue-specific studies is that there are no common reference genes. Rather, reference genes should be chosen de-novo for each species/tissue/condition studied. The present data obtained for WT untreated mouse hippocampus based on the RefGenes tool point out that the expression of reference genes commonly used in qPCR analyses are as unstable as the genes of interest. As discussed by Hugget et al.63 inter-group variability in normalizing gene expression is acceptable when the group effect on the expression of the gene of interest is substantially larger. In the field of biological psychiatry the effect sizes rarely exceed 1.5 fold64. According to the RefGenes tool, the lowest standard deviation (SD) of log2-transformed signals in mouse hippocampus is 0.12, and the SD of the commonly used housekeeping genes is ~0.45 (relative fold change of 1.18 and 1.8, respectively). Thus, changes in the expression level of genes of interest may be obscured by the variation in the expression of the normalizing gene. Moreover, our results suggest that the biological effect of the studied condition on the reference gene chosen may be larger than the effect on the gene of interest, resulting in significant but erroneous results. Similar findings in different tissues were reported by others65,66.
Stability of reference genes in control animals (namely, the SD of the genes’ expression in WT untreated animals) provides information regarding the within-group variability. Though ideally one should estimate the stability of the reference genes in both the control and the treatment groups, it is unlikely that microarrays data from a sufficient number of similar treatment samples is obtainable. Nevertheless, it is reasonable to assume that the stability of given reference genes in treated samples are similar to, or lower than that in control samples. Differences in the response to treatment of individual subjects/samples as well as potential differences in the actual treatment dose (e.g. injection volume deviations, variation in drug-containing food consumption) are additional sources of variation in gene expression.
To sum up, here we show that qPCR quantification of the effect of two weeks lithium treatment on BCL2 and MARCKS expression strongly depends on the normalizing gene chosen. Specifically, both genes exhibit downregulation when normalized to MAPK6 or ANKRD11, but upregulation when normalized to β-Actin. Inverse results obtained using different normalizing genes have previously been reported by others62. Our literature search as well as analysis of two publicly available datasets of lithium treatment revealed inconsistency among previous studies reporting an increase in BCL2 levels following lithium treatment. Moreover, the majority of studies supporting this increase either: (a) used lithium doses exceeding therapeutically-relevant ones; (b) normalized the signals to genes/proteins shown to be affected by lithium; or (c) used a small sample size. We also note that the increase in BCL2 expression was not supported by any of the numerous microarrays studies.
We therefore suggest that reports regarding lithium’s effect on BCL2 expression be considered with caution. While it cannot be ruled out that under some conditions lithium treatment increases BCL2 expression, this effect may not be a general and reproducible one57,67. According to our literature search this effect was primarily observed in rats at a four-week time point using treatment regimen that potentially results in higher than the therapeutically-relevant levels. Due to the complex nature of lithium’s mode of action and the diversity of BCL2’s functions, an effort should be invested in the experimental design, report, and interpretation of future studies. For example, rather than measuring BCL2 mRNA or protein levels in whole cell lysates, isolated subcellular organelles such as mitochondria, endoplasmic reticulum, nuclei, or cytosol might provide more consistent results. Furthermore, as lithium induces cell proliferation, and as BCL2 is also expressed in newly-born cells, BCL2 levels might better be normalized to cell count whenever possible. When normalization to specific genes or proteins is carried out, verification that they are not affected by the condition studied is mandatory. In addition, providing the non-normalized data as well as the results obtained post-normalization to each of the normalizing genes/proteins chosen may enable more reliable comparison among studies.
Perhaps the use of normalizing genes in psychiatry research should be revised. The following alternatives may be considered: (1) Utilizing methodologies to obtain higher precision of sample concentration. (2) Running duplicate/triplicate measurements of each sample, dismissing results exhibiting extreme standard deviation among the replicates relative to the expression level. (3) Comparing the expression ratios between two genes responding in opposite directions (e.g. the anti-apoptotic BCL2 and the pro-apoptotic Bax) rather than individual genes. Each of the above might improve our ability to reveal true biological effects.
Methods
All procedures involving animals were reviewed and approved by the Ben-Gurion University animal experimentation ethics committee. The methods were carried out in accordance with the approved guidelines.
Animals
Male, 10–12 weeks old wildtype mice from the IMPA1 colony68 were used. Animals were maintained on a 12 h/12 h light/dark cycle (lights on between 8:00 a.m and 8:00 p.m.) with ad libitum access to food and water. Sample collection was performed during the light phase of the cycle between 9:00 am and 7:00 pm. All experiments were performed in accordance with the Ben-Gurion University animal experimentation ethics committee guidelines and regulations.
Chronic lithium treatment
WT mice were divided into two groups (control and Li-treatment) and subjected to lithium-supplemented food or regular food for two weeks, as previously described69. At the end of the treatment, blood was extracted using cardiac puncture. Lithium plasma levels were measured in an ion-selective electrode apparatus ISE (AVL 9180 Electrolyte Analyzer, Hoffmann-La Roche, Basel, Switzerland). The measured lithium levels were in the range of 0.52–0.91 mM.
RNA extraction
Total RNA was extracted from hippocampi specimens using the TRI reagent (Sigma-Aldrich, St. Louis, MO) followed by purification using the RNeasy kit (Qiagen, Germantown, MD). RNA concentration was determined spectrophotometrically (NanoDrop 2000, Thermo Fisher Scientific, Waltham, MA).
Real-time PCR
RNA was reverse transcribed using Verso cDNA (Thermo Fisher Scientific, Waltham, MA). Real-time PCR was performed using ABsolute™ Blue Syber mix (ABgene, Lithuania) and Eco qPCR system (Illumina, San Diego, CA). The thermal cycler program was as follows: hold on 95 °C for 15 min, followed by 40 cycles of: 10 sec at 95 °C, 15 sec at 60 °C, 40 sec at 72 °C. The relative expression of each gene was calculated using the Pfaffl method70 implemented in the Eco qPCR system software. Samples were run in duplicates. Only samples with standard deviation (SD) < 0.05 between the duplicates were used in the analysis. Each qPCR plate contained equal number of samples from each of the groups. Samples with normalized expression values > |1.96| SD from the mean were removed from the analysis. Wilcoxon rank sum test was used to determine the significance of the results.
The expression of BCL2 and MARCKS was evaluated separately with each of the three normalizing genes ACTB, MAPK6 or ANKRD11. We chose these genes for the following reasons: (1) MAPK6 was recommended as a brain-reference gene15. Previous study from Padmos et al.71 reported no effect of lithium treatment on MAPK6 expression in human monocyte. Unpublished data from our microarrays study16 confirmed that brain MAPK6 expression is not affected by lithium treatment (SMIT1 colony: p > 0.23; IMPA1 colony: p > 0.29, raw data is available upon request). (2) ACTB is the most commonly used normalizing gene in brain qPCR studies. (3) Based on the RefGenes tool of the Genevestigator software60 ANKRD11 is the most stable gene in mouse hippocampus that was not affected by lithium treatment in our previous microarray study16 (SMIT1 colony: p > 0.8; IMPA1 colony: p > 0.14), as discussed in detail under the Results section. Supplementary Table S1 lists the primers’ sequences for the genes examined and the efficiencies of their reactions. The corresponding r2 values of the standard curves calculated for each of the genes were > 0.99.
Estimation of gene expression stability
The RefGenes tool of the Genevestigator software platform was used according to the online tutorial (http://www.refgenes.org/rg/doc/ tutorial.jsp) in order to evaluate gene expression stability. Sample sets included in the analysis fulfilled the following criteria: Mouse 430_2: 40 k array/only WT genetic background/hippocampal samples. This filter resulted in a total of 80 arrays which were used to calculate the median and the standard deviation (SD) of BCL2, ACTB, MAPK6 as well as GAPDH, another commonly used normalizing gene72. Calculation of the expected maximal fold of change relative to the mean expression signal for each of these genes was carried out using the inversed log2 of 1.96*SD73.
Analysis of publicly available microarray datasets of the effect of lithium treatment
We performed differential expression analysis of two publicly available gene expression datasets – GSE66277 and GSE35291. GSE66277 contains expression profiles from three rat brain regions (cortex, hippocampus and striatum) following chronic treatment with different antipsychotics and mood-stabilizers (including lithium). GSE35291 contains expression profiles of hematopoietic progenitors following a one-week treatment with lithium or valproic acid. For GSE66277 signal intensities were quantile-normalized and log2-transformed using the RMA function from the R Bioconductor ‘affy’ package. For GSE35291 we used the log2 quantile-normalized intensities available on the Gene Expression Omnibus (GEO) site. We next mapped each probeset to a corresponding gene using Gemma annotation files74. Differential analyses were performed using the eBayes function from ‘limma’ R Bioconductor package. Benjamini–Hochberg procedure was used to control the false discovery rate (FDR).
References
Machado-Vieira, R., Manji, H. K. & Zarate, C. A. The role of lithium in the treatment of bipolar disorder: convergent evidence for neurotrophic effects as a unifying hypothesis. Bipolar Disord 11(Suppl 2), 92–109 (2009).
Chen, R. W. & Chuang, D. M. Long term lithium treatment suppresses p53 and Bax expression but increases Bcl-2 expression. A prominent role in neuroprotection against excitotoxicity. J Biol Chem 274, 6039–42 (1999).
Chen, G. et al. The mood-stabilizing agents lithium and valproate robustly increase the levels of the neuroprotective protein bcl-2 in the CNS. J Neurochem 72, 879–82 (1999).
Hockenbery, D., Nunez, G., Milliman, C., Schreiber, R. D. & Korsmeyer, S. J. Bcl-2 is an inner mitochondrial membrane protein that blocks programmed cell death. Nature 348, 334–6 (1990).
Chiu, C. T. & Chuang, D. M. Molecular actions and therapeutic potential of lithium in preclinical and clinical studies of CNS disorders. Pharmacol Ther 128, 281–304 (2010).
Levine, B., Sinha, S. & Kroemer, G. Bcl-2 family members: dual regulators of apoptosis and autophagy. Autophagy 4, 600–6 (2008).
Sarkar, S. et al. Lithium induces autophagy by inhibiting inositol monophosphatase. J Cell Biol 170, 1101–11 (2005).
Sade, Y. et al. IP3 accumulation and/or inositol depletion: two downstream lithium’s effects that may mediate its behavioral and cellular changes. Transl. Psychiatry 6, e968 (2016).
Cohen, Y., Chetrit, A., Sirota, P. & Modan, B. Cancer morbidity in psychiatric patients: influence of lithium carbonate treatment. Med Oncol 15, 32–6 (1998).
Hallas, J. et al. Cancer risk in long-term users of valproate: a population-based case-control study. Cancer Epidemiol Biomark. Prev 18, 1714–9 (2009).
Liu, J. J., Lin, M., Yu, J. Y., Liu, B. & Bao, J. K. Targeting apoptotic and autophagic pathways for cancer therapeutics. Cancer Lett 300, 105–14 (2010).
Del Bufalo, D., Biroccio, A., Leonetti, C. & Zupi, G. Bcl-2 overexpression enhances the metastatic potential of a human breast cancer line. FASEB J 11, 947–53 (1997).
Placzek, W. J. et al. A survey of the anti-apoptotic Bcl-2 subfamily expression in cancer types provides a platform to predict the efficacy of Bcl-2 antagonists in cancer therapy. Cell Death Dis 1, e40 (2010).
Damri, O. et al. Molecular effects of lithium are partially mimicked by inositol-monophosphatase (IMPA)1 knockout mice in a brain region-dependent manner. Eur. Neuropsychopharmacol. J. Eur. Coll. Neuropsychopharmacol. 25, 425–434 (2015).
Cai, J. H. et al. Validation of rat reference genes for improved quantitative gene expression analysis using low density arrays. Biotechniques 42, 503–12 (2007).
Toker, L. et al. Inositol-related gene knockouts mimic lithium’s effect on mitochondrial function. Neuropsychopharmacology 39, 319–28 (2013).
McQuillin, A., Rizig, M. & Gurling, H. M. D. A microarray gene expression study of the molecular pharmacology of lithium carbonate on mouse brain mRNA to understand the neurobiology of mood stabilization and treatment of bipolar affective disorder. Pharmacogenet. Genomics 17, 605–617 (2007).
Chetcuti, A., Adams, L. J., Mitchell, P. B. & Schofield, P. R. Microarray gene expression profiling of mouse brain mRNA in a model of lithium treatment. Psychiatr Genet 18, 64–72 (2008).
Seelan, R. S., Khalyfa, A., Lakshmanan, J., Casanova, M. F. & Parthasarathy, R. N. Deciphering the lithium transcriptome: microarray profiling of lithium-modulated gene expression in human neuronal cells. Neuroscience 151, 1184–97 (2008).
Fatemi, S. H., Reutiman, T. J. & Folsom, T. D. The role of lithium in modulation of brain genes: relevance for aetiology and treatment of bipolar disorder. Biochem Soc Trans 37, 1090–5 (2009).
Walasek, M. A. et al. The combination of valproic acid and lithium delays hematopoietic stem/progenitor cell differentiation. Blood 119, 3050–3059 (2012).
Linares, G. R. et al. Preconditioning mesenchymal stem cells with the mood stabilizers lithium and valproic acid enhances therapeutic efficacy in a mouse model of Huntington’s disease. Exp. Neurol. 281, 81–92 (2016).
Lenox, R. H., Watson, D. G., Patel, J. & Ellis, J. Chronic lithium administration alters a prominent PKC substrate in rat hippocampus. Brain Res 570, 333–40 (1992).
Watson, D. G. & Lenox, R. H. Chronic lithium-induced down-regulation of MARCKS in immortalized hippocampal cells: potentiation by muscarinic receptor activation. J Neurochem 67, 767–77 (1996).
Lenox, R. H. & Wang, L. Molecular basis of lithium action: integration of lithium-responsive signaling and gene expression networks. Mol Psychiatry 8, 135–44 (2003).
Ghoshdastidar, D., Dutta, R. N. & Poddar, M. K. In vivo distribution of lithium in plasma and brain. Indian J Exp Biol 27, 950–4 (1989).
Alural, B., Ozerdem, A., Allmer, J., Genc, K. & Genc, S. Lithium protects against paraquat neurotoxicity by NRF2 activation and miR-34a inhibition in SH-SY5Y cells. Front. Cell. Neurosci. 9, 209 (2015).
Hiroi, T., Wei, H., Hough, C., Leeds, P. & Chuang, D.-M. Protracted lithium treatment protects against the ER stress elicited by thapsigargin in rat PC12 cells: roles of intracellular calcium, GRP78 and Bcl-2. Pharmacogenomics J. 5, 102–111 (2005).
Plant, K. E. et al. The neuroprotective action of the mood stabilizing drugs lithium chloride and sodium valproate is mediated through the up-regulation of the homeodomain protein Six1. Toxicol. Appl. Pharmacol. 235, 124–134 (2009).
Son, H. et al. Lithium enhances long-term potentiation independently of hippocampal neurogenesis in the rat dentate gyrus. J. Neurochem. 85, 872–881 (2003).
Yu, I. T., Kim, J. S., Lee, S.-H., Lee, Y.-S. & Son, H. Chronic lithium enhances hippocampal long-term potentiation, but not neurogenesis, in the aged rat dentate gyrus. Biochem. Biophys. Res. Commun. 303, 1193–1198 (2003).
Chen, G., Rajkowska, G., Du, F., Seraji-Bozorgzad, N. & Manji, H. K. Enhancement of hippocampal neurogenesis by lithium. J Neurochem 75, 1729–34 (2000).
Hanson, N. D., Nemeroff, C. B. & Owens, M. J. Lithium, but not fluoxetine or the corticotropin-releasing factor receptor 1 receptor antagonist R121919, increases cell proliferation in the adult dentate gyrus. J Pharmacol Exp Ther 337, 180–6 (2011).
Bernier, P. J. & Parent, A. Bcl-2 Protein as a Marker of Neuronal Immaturity in Postnatal Primate Brain. J. Neurosci. 18, 2486–2497 (1998).
Bush, A. L. & Hyson, R. L. Lithium increases bcl-2 expression in chick cochlear nucleus and protects against deafferentation-induced cell death. Neuroscience 138, 1341–1349 (2006).
Steward, O. & Rubel, E. W. Afferent influences on brain stem auditory nuclei of the chicken: Cessation of amino acid incorporation as an antecedent to age-dependent transneuronal degeneration. J. Comp. Neurol. 231, 385–395 (1985).
Wilkinson, B. L., Sadler, K. A. & Hyson, R. L. Rapid deafferentation-induced upregulation of bcl-2 mRNA in the chick cochlear nucleus. Brain Res. Mol. Brain Res. 99, 67–74 (2002).
McBride, E. G., Rubel, E. W. & Wang, Y. Afferent Regulation of Chicken Auditory Brainstem Neurons: Rapid Changes in Phosphorylation of Elongation Factor 2. J. Comp. Neurol. 521, 1165–1183 (2013).
Hizli, A. A. et al. Phosphorylation of Eukaryotic Elongation Factor 2 (eEF2) by Cyclin A–Cyclin-Dependent Kinase 2 Regulates Its Inhibition by eEF2 Kinase. Mol. Cell. Biol. 33, 596–604 (2013).
Karyo, R. et al. Identification of eukaryotic elongation factor-2 as a novel cellular target of lithium and glycogen synthase kinase-3. Mol. Cell. Neurosci. 45, 449–455 (2010).
Bangaru, M. L. Y., Park, F., Hudmon, A., McCallum, J. B. & Hogan, Q. H. Quantification of gene expression after painful nerve injury: validation of optimal reference genes. J. Mol. Neurosci. MN 46, 497–504 (2012).
Beasley, C. L. et al. Proteomic analysis of the anterior cingulate cortex in the major psychiatric disorders: Evidence for disease-associated changes. Proteomics 6, 3414–25 (2006).
Behan, A. T., Byrne, C., Dunn, M. J., Cagney, G. & Cotter, D. R. Proteomic analysis of membrane microdomain-associated proteins in the dorsolateral prefrontal cortex in schizophrenia and bipolar disorder reveals alterations in LAMP, STXBP1 and BASP1 protein expression. Mol Psychiatry 14, 601–13 (2009).
Focking, M. et al. Common proteomic changes in the hippocampus in schizophrenia and bipolar disorder and particular evidence for involvement of cornu ammonis regions 2 and 3. Arch Gen Psychiatry 68, 477–88 (2011).
Bennett, G. S., Hollander, B. A., Laskowska, D. & DiLullo, C. Rapid degradation of newly synthesized tubulin in lithium-treated sensory neurons. J Neurochem 57, 130–9 (1991).
Bosetti, F. et al. Analysis of gene expression with cDNA microarrays in rat brain after 7 and 42 days of oral lithium administration. Brain Res Bull 57, 205–9 (2002).
Cannell, G. R., Bailey, M. J. & Dickinson, R. G. Inhibition of tubulin assembly and covalent binding to microtubular protein by valproic acid glucuronide in vitro. Life Sci 71, 2633–43 (2002).
Corena-McLeod Mdel, P. et al. Paliperidone as a mood stabilizer: a pre-frontal cortex synaptoneurosomal proteomics comparison with lithium and valproic acid after chronic treatment reveals similarities in protein expression. Brain Res 1233, 8–19 (2008).
Nielsen, J. et al. Proteomic analysis of lithium-induced nephrogenic diabetes insipidus: mechanisms for aquaporin 2 down-regulation and cellular proliferation. Proc Natl Acad Sci U A 105, 3634–9 (2008).
Lakshmanan, J. et al. Proteomic analysis of rat prefrontal cortex after chronic lithium treatment. Proteomics Bioinforma. 5, 140–146 (2012).
Costain, W. J., Haqqani, A. S., Rasquinha, I., Giguere, M. S. & Slinn, J. Cerebral Ischemia Induced Proteomic Alterations: Consequences for the Synapse and Organelles. (In Tech, 2012).
He, C. & Klionsky, D. J. Regulation mechanisms and signaling pathways of autophagy. Annu Rev Genet 43, 67–93 (2009).
Dengjel, J. et al. Identification of autophagosome-associated proteins and regulators by quantitative proteomic analysis and genetic screens. Mol Cell Proteomics 11(M111), 014035 (2012).
Powell, T. R. et al. Mood-stabilizers differentially affect housekeeping gene expression in human cells. Int. J. Methods Psychiatr. Res. 23, 279–288 (2014).
Huang, X., Wu, D.-Y., Chen, G., Manji, H. & Chen, D. F. Support of retinal ganglion cell survival and axon regeneration by lithium through a Bcl-2-dependent mechanism. Invest. Ophthalmol. Vis. Sci. 44, 347–354 (2003).
Keshavarz, M., Emamghoreishi, M., Nekooeian, A. A., J Warsh, J. & Zare, H. R. Increased bcl-2 Protein Levels in Rat Primary Astrocyte Culture Following Chronic Lithium Treatment. Iran. J. Med. Sci. 38, 255–262 (2013).
Lai, J. S., Zhao, C., Warsh, J. J. & Li, P. P. Cytoprotection by lithium and valproate varies between cell types and cellular stresses. Eur. J. Pharmacol. 539, 18–26 (2006).
Valvassori, S. S. et al. Effects of mood stabilizers on oxidative stress-induced cell death signaling pathways in the brains of rats subjected to the ouabain-induced animal model of mania: [Mood stabilizers exert protective effects against ouabain-induced activation of the cell death pathway]. J. Psychiatr. Res. 65, 63–70 (2015).
Lowthert, L. et al. Increased ratio of anti-apoptotic to pro-apoptotic Bcl2 gene-family members in lithium-responders one month after treatment initiation. Biol Mood Anxiety Disord 2, 15 (2012).
Hruz, T. et al. RefGenes: identification of reliable and condition specific reference genes for RT-qPCR data normalization. BMC Genomics 12, 156 (2011).
Radonić, A. et al. Guideline to reference gene selection for quantitative real-time PCR. Biochem. Biophys. Res. Commun. 313, 856–862 (2004).
Durrenberger, P. F. et al. Selection of novel reference genes for use in the human central nervous system: a BrainNet Europe Study. Acta Neuropathol 124, 893–903 (2012).
Huggett, J., Dheda, K., Bustin, S. & Zumla, A. Real-time RT-PCR normalisation; strategies and considerations. Genes Immun. 6, 279–284 (2005).
Beech, R. D. et al. Gene-expression differences in peripheral blood between lithium responders and non-responders in the Lithium Treatment-Moderate dose Use Study (LiTMUS). Pharmacogenomics J, https://doi.org/10.1038/tpj.2013.16 (2013).
Bas, A., Forsberg, G., Hammarström, S. & Hammarström, M.-L. Utility of the Housekeeping Genes 18S rRNA, β-Actin and Glyceraldehyde-3-Phosphate-Dehydrogenase for Normalization in Real-Time Quantitative Reverse Transcriptase-Polymerase Chain Reaction Analysis of Gene Expression in Human T Lymphocytes. Scand. J. Immunol. 59, 566–573 (2004).
Tricarico, C. et al. Quantitative real-time reverse transcription polymerase chain reaction: normalization to rRNA or single housekeeping genes is inappropriate for human tissue biopsies. Anal. Biochem. 309, 293–300 (2002).
Corson, T. W., Woo, K. K., Li, P. P. & Warsh, J. J. Cell-type specific regulation of calreticulin and Bcl-2 expression by mood stabilizer drugs. Eur. Neuropsychopharmacol. 14, 143–150 (2004).
Cryns, K. et al. IMPA1 is essential for embryonic development and lithium-like pilocarpine sensitivity. Neuropsychopharmacology 33, 674–84 (2008).
O’Brien, W. T. et al. Glycogen synthase kinase-3beta haploinsufficiency mimics the behavioral and molecular effects of lithium. J. Neurosci. Off. J. Soc. Neurosci. 24, 6791–6798 (2004).
Pfaffl, M. W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res 29, e45 (2001).
Padmos, R. C. et al. A discriminating messenger RNA signature for bipolar disorder formed by an aberrant expression of inflammatory genes in monocytes. Arch. Gen. Psychiatry 65, 395–407 (2008).
Kozera, B. & Rapacz, M. Reference genes in real-time PCR. J. Appl. Genet. 54, 391–406 (2013).
Quackenbush, J. Microarray data normalization and transformation. Nat Genet 32(Suppl), 496–501 (2002).
Zoubarev, A. et al. Gemma: a resource for the reuse, sharing and meta-analysis of expression profiling data. Bioinformatics 28, 2272–2273 (2012).
Schuettauf, F. et al. Citicoline and lithium rescue retinal ganglion cells following partial optic nerve crush in the rat. Exp. Eye Res. 83, 1128–1134 (2006).
Hammonds, M. D., Shim, S. S., Feng, P. & Calabrese, J. R. Effects of subchronic lithium treatment on levels of BDNF, Bcl-2 and phospho-CREB in the rat hippocampus. Basic Clin. Pharmacol. Toxicol. 100, 356–359 (2007).
Hammonds, M. D. & Shim, S. S. Effects of 4-week Treatment with Lithium and Olanzapine on Levels of Brain-derived Neurotrophic Factor, B-Cell CLL/Lymphoma 2 and Phosphorylated Cyclic Adenosine Monophosphate Response Element-binding Protein in the Sub-regions of the Hippocampus. Basic Clin. Pharmacol. Toxicol. 105, 113–119 (2009).
Liechti, F. D. et al. The Mood-Stabilizer Lithium Prevents Hippocampal Apoptosis and Improves Spatial Memory in Experimental Meningitis. PLoS ONE 9, e113607 (2014).
Chen, W.-T., Huang, T.-L. & Tsai, M.-C. Bcl-2 associated with severity of manic symptoms in bipolar patients in a manic phase. Psychiatry Res. 225, 305–308 (2015).
Song, N. et al. Mood stabilizers commonly restore staurosporine-induced increase of p53 expression and following decrease of Bcl-2 expression in SH-SY5Y cells. Prog. Neuropsychopharmacol. Biol. Psychiatry 38, 183–189 (2012).
McColl, G. et al. Pharmacogenetic Analysis of Lithium-induced Delayed Aging in Caenorhabditis elegans. J. Biol. Chem. 283, 350–357 (2008).
Acknowledgements
We thank Dr. Paul Pavlidis, Department of Psychiatry, University of British Columbia, Vancouver, BC, Canada and Prof. Haim Einat, School of Behavioral Sciences, Academic College of Tel Aviv-Yafo, Israel for their thoughtful remarks and suggestions.
Author information
Authors and Affiliations
Contributions
G.A. and L.T. designed the study and the experiments. O.D. performed the experiments and carried out the preliminary statistical analyses, L.T. analyzed the data, G.A. and L.T. wrote the manuscript. All authors reviewed the final version of the manuscript.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Odeya, D., Galila, A. & Lilah, T. The observed alteration in BCL2 expression following lithium treatment is influenced by the choice of normalization method. Sci Rep 8, 6399 (2018). https://doi.org/10.1038/s41598-018-24546-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-24546-1
This article is cited by
-
Mitochondrial function parameters as a tool for tailored drug treatment of an individual with psychosis: a proof of concept study
Scientific Reports (2020)
-
Biological Targets Underlying the Antisuicidal Effects of Lithium
Current Behavioral Neuroscience Reports (2020)
-
Utilization of lithium incorporated mesoporous silica for preventing necrosis and increase apoptosis in different cancer cells
BMC Chemistry (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
