
Abstract
Contact-dependent growth inhibition (CDI) is a wide-spread mechanism of inter-bacterial competition. CDI+ bacteria deliver CdiA-CT toxins into neighboring bacteria and produce specific immunity proteins that protect against self-intoxication. The CdiA-CT toxin from uropathogenic Escherichia coli 536 is a latent tRNase that is only active when bound to the cysteine biosynthetic enzyme CysK. Remarkably, the CysK:CdiA-CT binding interaction mimics the ‘cysteine synthase’ complex of CysK:CysE. The C-terminal tails of CysE and CdiA-CT each insert into the CysK active-site cleft to anchor the respective complexes. The dissociation constant for CysK:CdiA-CT (K d ~ 11 nM) is comparable to that of the E. coli cysteine synthase complex (K d ~ 6 nM), and both complexes bind through a two-step mechanism with a slow isomerization phase after the initial encounter. However, the second-order rate constant for CysK:CdiA-CT binding is two orders of magnitude slower than that of the cysteine synthase complex, suggesting that CysE should outcompete the toxin for CysK occupancy. However, we find that CdiA-CT can effectively displace CysE from pre-formed cysteine synthase complexes, enabling toxin activation even in the presence of excess competing CysE. This adventitious binding, coupled with the very slow rate of CysK:CdiA-CT dissociation, ensures robust nuclease activity in target bacteria.
Similar content being viewed by others

Introduction
Though long considered to be isolated and independent unicellular organisms, bacteria engage in a multitude of cooperative and competitive behaviors. Many bacteria secrete soluble antibiotics and bacteriocins1,2,3,4, which diffuse through the environment and kill competing bacteria at a distance. More recently, proximity-dependent inter-bacterial competition systems have been characterized5,6,7,8. This phenomenon was first described in Escherichia coli isolate EC93, which inhibits the growth of other E. coli strains in a contact-dependent manner5. Contact-dependent growth inhibition (CDI) is mediated by CdiB/CdiA two-partner secretion proteins, which transfer protein toxins between Gram-negative bacteria9, 10. CdiB is an outer-membrane transport protein that exports CdiA onto the cell surface. CdiA forms a long β-helical filament that extends from the inhibitor cell to bind specific receptors on neighboring bacteria. Upon binding receptor, CdiA delivers its C-terminal toxin domain (CdiA-CT) into the target cell to inhibit growth. CDI+ bacteria also express CdiI immunity proteins, which bind to the CdiA-CT domain and neutralize toxin activity to prevent self-intoxication. Analysis of CdiA from many species has revealed that the family carries a wide variety of C-terminal toxin domains, each with a distinct activity11,12,13,14,15. Thus, a given CdiI immunity protein only protects against its cognate toxin and not the toxins deployed by other bacteria. Together, these observations suggest that CDI systems mediate inter-bacterial competition for growth niches and other environmental resources.
We recently discovered that the CDI toxin deployed by uropathogenic E. coli 536 is a latent tRNase that is only active when bound to the biosynthetic enzyme CysK16. CysK is a pyridoxal 5′-phosphate (PLP)-dependent O-acetyl-L-Ser sulfhydrylase that catalyzes the last step of cysteine biosynthesis in eubacteria, plants and some archaea17,18,19. E. coli and many other bacteria encode an additional isozyme, CysM. Both sulfhydrylases are coordinately regulated with the enzymes responsible for sulfate reduction to bisulfide17, although the functional role of CysM is less well characterized. The structure and catalytic properties of CysK from Gram-negative bacteria and plants have been characterized thoroughly20,21,22,23,24,25,26,27,28,29,30,31,32,33. CysK has long been known to form a high-affinity “cysteine synthase” (CS) complex with CysE, which is a serine O-acetyltransferase responsible for the penultimate reaction in cysteine biosynthesis34,35,36,37. The three-dimensional structure of the cysteine synthase complex is unknown, but biochemical studies indicate that each CysE hexamer binds to two CysK homodimers38. Moreover, it is well established that the flexible C-terminal tail of CysE inserts into the CysK active site to anchor the complex36, 37, 39. The C-terminal Ile residue of CysE is particularly critical, and deletion or mutation of this conserved residue consistently interferes with cysteine synthase assembly39, 40. The Ile side-chain mimics substrate to bind the CysK active site, and consequently the cysteine synthase complex is dissociated with micromolar concentrations of O-acetyl-L-Ser34, 41. Remarkably, the E. coli 536 CdiA-CT toxin mimics CysE and uses its C-terminal peptide motif to bind the CysK active site16, 42. In fact, a number of proteins engage in so-called “moonlighting” binding interactions with CysK homologs43. One intriguing example is the EGL-9 prolyl hydroxylase from Caenorhabditis elegans, which uses its C-terminal Ile residue to interact with CYSL-1, a CysK homolog that has lost biosynthetic activity but retains the bisulfide-binding site. Under hypoxic conditions, bisulfide accumulates and promotes EGL-9:CYSL-1 binding. The sequestered EGL-9 is no longer able to hydroxylate HIF-1, which stabilizes the transcription factor and increases the expression of genes required to respond to hypoxia44. Thus, CysK homologs have been co-opted to regulate diverse biological activities.
Here, we explore the thermodynamics and kinetics of the CdiA-CT interaction with E. coli CysK (EcCysK) to gain insight into toxin activation. We find that the dissociation constant for the EcCysK:CdiA-CT complex is comparable to that of the CS complex, suggesting that EcCysE could attenuate toxicity by competing with CdiA-CT for access to EcCysK. In addition, the second-order rate constant for cysteine synthase complex formation is ~200-fold greater than the rate constant for EcCysK:CdiA-CT binding. Though cysteine synthase assembly is kinetically favored, CdiA-CT toxin is still activated in the presence of competing EcCysE. Robust toxin activation reflects the ability of CdiA-CT to displace EcCysE from pre-formed cysteine synthase complexes. This property, coupled with the very slow rate of EcCysK:CdiA-CT dissociation, ensures toxin activation upon entry into target bacteria. Finally, we show that CysK homologs from different bacterial species support CdiA-CT toxin activity to varying degrees. Although CDI-mediated toxin delivery only occurs between closely related bacteria45, 46, CdiA-CT toxin homologs are found in several species and therefore must interact with different CysK variants. Because CDI genes are acquired through horizontal gene transfer47, 48, we propose that the toxin domain evolved to bind a highly conserved partner that is ubiquitous in bacteria.
Results
Two CdiA-CT toxin domains bind each CysK dimer
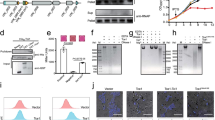
Our previous work has shown that the CdiA-CT toxin domain forms a stable complex with EcCysK and that this interaction is required for toxic tRNase activity in vivo and in vitro 16. The initial study suggested that the C-terminal tail of CdiA-CT inserts into the EcCysK active site, and this conclusion was recently confirmed by crystal structures of the EcCysK:CdiA-CT complex42. Therefore, we reasoned that EcCysK:CdiA-CT complex formation could be monitored by measuring changes in pyridoxal 5′-phosphate (PLP) fluorescence. This spectroscopic approach has been used extensively to track cysteine synthase complex formation38 as well as to identify CysK inhibitors49,50,51,52,53,54. Indeed, PLP fluorescence increased about five-fold when EcCysK was titrated with increasing concentrations of CdiA-CT (Fig. 1A). This effect is very similar to that observed for the cysteine synthase complexes of Haemophilus influenzae 38 and E. coli 55. Moreover, the fluorescence spectrum of EcCysK:CdiA-CT exhibited a blue-shift in the emission maximum from 505 to 498 nm compared to free EcCysK (Fig. 1A). This latter change indicates that the fluorophore is in a less polar environment, consistent with insertion of the toxin’s C-terminal Ile residue into the EcCysK active site. Stoichiometric titrations determined a molar ratio of 1.1:1 (Fig. 1B), in agreement with crystal structures showing two CdiA-CT domains bound to each EcCysK dimer42.

EcCysK:CdiA-CT complex formation. (A) Fluorescence emission spectra of EcCysK excited at 412 nm. Spectra were collected with EcCysK (1.15 µM) in the presence of the indicated concentrations of CdiA-CT. (B) Stoichiometry of the EcCysK:CdiA-CT complex. 1 μM EcCysK was titrated with CdiA-CT to saturation. The intersection of the lines corresponds to a CdiA-CT:EcCysK ratio of 1.1:1 (dashed line). (C) Determination of the EcCysK:CdiA-CT dissociation constant. EcCysK (80 nM) was titrated with increasing concentrations of CdiA-CT, and fluorescence emission at 500 nm monitored at the indicated times. The solid line indicates the Eq. 2 fit to the 20 min data set, with K d = 10 ± 11 nM and [CysK] = 134 ± 33 nM. (D) CdiA-CT inhibits EcCysK sulfhydrylase activity. EcCysK (6 nM) was titrated with CdiA-CT and sulfhydrylase activity measured as described in the Methods. Eq. 3 was fitted to the dependence of vi/v0 on CdiA-CT concentration, yielding an apparent IC50 that was used to calculate a K i of 11.0 ± 0.4 nM (closed circles). A K i of 6.4 ± 0.5 nM was calculated under the same conditions in the presence of 2.4 µM CdiI (open squares).
CdiA-CT and CysE bind to CysK with comparable affinity
The affinity of the EcCysK:CdiA-CT complex can be estimated through titrations of dilute EcCysK with toxin. We observed the same spectroscopic changes upon CS complex formation56, but the emission spectra evolved over time, stabilizing after about 20 min at the lowest toxin concentrations (Fig. 1C). Fitting of Eq. 2 to binding data collected after 20 min yields an estimated K d of about 10 nM (Fig. 1C). The protein concentrations required for fluorescence-based titrations limit the measurable dissociation constants to about 5 nM. Therefore, we used an orthologous assay to measure binding interactions more accurately. Because CdiA-CT occludes the EcCysK active site, complex formation can be monitored by measuring fractional sulfhydrylase activity as a function of toxin concentration (Fig. 1D). The half maximal inhibitory concentration (IC50) of 15.4 ± 0.6 nM was obtained by fitting Eq. 3 to these data. Accounting for substrate concentration and the K M, the IC50 was converted to an inhibition constant (K i) of 11.0 ± 0.4 nM using Eq. 4. This value is in agreement with the EcCysK:CdiA-CT binding constant calculated from surface plasmon resonance data57. Moreover, EcCysE inhibits EcCysK activity with a K i of 6.2 ± 0.7 nM55, indicating that the toxin and EcCysE bind to EcCysK with similar affinities.
CdiI immunity protein binds specifically to CdiA-CT toxin, but also forms a ternary complex with CdiA-CT and EcCysK16, 42. A recent report has suggested that the CdiA-CT:CdiI complex has a six-fold higher affinity for EcCysK than CdiA-CT toxin alone57. However, the crystal structure of the EcCysK:CdiA-CT:CdiI ternary complex shows that the immunity protein makes no direct contacts with EcCysK42. To monitor the influence of CdiI on EcCysK:CdiA-CT complex formation, we measured EcCysK sulfhydrylase activity in the presence of CdiA-CT and excess CdiI (Fig. 1D). The calculated K i in the presence of CdiA-CT and CdiI was 6.4 ± 0.5 nM, indicating a similar affinity for EcCysK under these conditions.
The slow binding of EcCysK and CdiA-CT prompted an examination of complex formation under pre-steady state conditions to calculate microscopic rate constants56, 58. We used stopped-flow spectroscopy to measure PLP fluorescence emission as a probe of CdiA-CT:EcCysK complex formation. Under the buffer and temperature conditions used for equilibrium binding, we varied EcCysK or CdiA-CT concentrations under pseudo-first order conditions, keeping the concentration of the binding partner constant (Fig. 2A,B). A single exponential equation (Eq. 5) was sufficient to fit all the kinetic traces. The kobs values calculated from Eq. 5 show a linear dependence on protein concentration with slopes of about 0.02 µM−1·s−1 (Fig. 2C). This linear relationship may indicate a single-step binding mechanism, but could also be obtained from a two-step process under conditions that do not allow saturation of the effect59. Because the dependence was linear independent of temperature between 5 °C and 37 °C (Fig. 2D), we were unable to ascertain whether EcCysK:CdiA-CT complex formation is limited by a conformational change. However, given the unusually slow binding kinetics, rapid formation of an encounter complex followed by a slow conformational rearrangement is likely (Fig. 2E). As previously observed for the cysteine synthase (CS) complex56 the initial fast step was not associated with changes in the fluorescence emission, and therefore only the slow rate-limiting process can be measured by this technique. According to this model, the dependence of kobs on protein concentration represents the linear portion of the hyperbole and is equal to the second-order rate constant (k3/K d). Fitting Eq. 6 to the dependence yields k3/K d of 2·104 M−1·s−1. By contrast, the second-order rate constant for the H. influenzae cysteine synthase complex is about 107 M−1·s−1 56 and 4·106 M−1·s−1 for the EcCysE:EcCysK complex (Fig. S1). Thus, the EcCysK:CdiA-CT complex forms ~200-fold more slowly than the E. coli cysteine synthase complex.

Pre-steady state kinetics of EcCysK:CdiA-CT assembly. (A,B) Representative time courses for the interaction of CdiA-CT (1 µM and 10 µM) with EcCysK (200 nM) (panel A) and EcCysK (1.7 µM and 16 µM) with CdiA-CT (270 nM) (panel B) as monitored by fluorescence emission intensity upon excitation at 412 nm. Individual traces are presented in grey, and the dashed black lines represent Eq. 5 fits to the time-course binding data. (C) Dependence of the observed kinetic constant (kobs) on EcCysK and CdiA-CT concentrations. The lines represent linear equation fits with slopes of 0.019 ± 0.007 µM−1·s−1 and 0.017 ± 0.007 µM−1·s−1. (D) Observed kinetic constant as a function of CysK concentration and temperature. Solid lines represent linear equation fits with slopes of 0.099 ± 0.019 µM−1·s−1 (37 °C), 0.017 ± 0.007 µM−1·s−1 (20 °C) and 0.005 ± 0.0003 µM−1·s−1 (5 °C). (E) Two-step model for EcCysK:CdiA-CT complex formation including a slow conformational change. CysK:CdiA-CT is the encounter complex, and CysK:CdiA-CT* corresponds to the isomerized, nucleolytic complex.
CdiA-CT competes with CysE for binding to CysK
The comparable affinities of the cysteine synthase and EcCysK:CdiA-CT complexes suggest that the toxin competes with EcCysE for access to EcCysK. We first used an indirect approach to test whether CdiA-CT interferes with the assembly of cysteine synthase complexes. Because EcCysE activity is stimulated when bound to EcCysK55, we titrated the CS complex with CdiA-CT and measured serine acetyltransferase activity. The maximal rate of serine acetylation was obtained with 28 nM EcCysE and 19 nM EcCysK, conditions in which the two proteins are at stoichiometric amounts based on the 3:2 CysE:CysK stoichiometry of the cysteine synthase complex. Pre-incubation of EcCysK with increasing concentrations of CdiA-CT reduced the stimulatory effect, decreasing acetyltransferase activity down to a plateau equivalent to that of free EcCysE (Fig. 3A).

EcCysE and CdiA-CT compete for binding to CysK. (A) CdiA-CT blocks formation of the cysteine synthase complex. Increasing concentrations of CdiA-CT were pre-incubated with EcCysK (19 nM) for 20 min prior to the addition of L-Ser (20 mM) and EcCysE (28 nM). Reactions were then initiated by addition of 0.3 mM acetyl-CoA and acetyltransferase activity measured by monitoring the decrease in acetyl-CoA absorption at 232 nm as described in the Methods. The activities of isolated CysE and CysE in the CS complex are shown by horizontal reference lines. (B) EcCysE inhibits CdiA-CT toxin activation in vivo. Inhibitor and target cells were co-cultured as described in the Methods. Total RNA and protein were isolated upon initial mixing and after 1 h, and analyzed by Northern blot (top panel) and SDS-PAGE (bottom panel). Where indicated, target cells over-produced EcCysE (+) or truncated EcCysE lacking the C-terminal tail (no tail). Target cells carried a deletion of the cysK gene (∆) where indicated. Asterisks (*) in the bottom panel indicate over-produced EcCysE proteins. (C) EcCysE inhibits CdiA-CT toxin activation in vitro. EcCysK and EcCysE were pre-incubated at the indicated concentrations (µM) prior to addition of CdiA-CT and RNA substrate as described in the Methods. Reactions were quenched after 10 min at 37 °C, then run on 8 M urea-polyacrylamide gels and visualized by ethidium bromide staining. The migration positions of 5 S rRNA and tRNA are indicated.
We next examined the influence of EcCysE on the EcCysK:CdiA-CT complex, first testing whether excess EcCysE blocks toxin activation during CDI. We incubated target bacteria with inhibitor cells that deploy CdiA-CT, then isolated RNA from the mixed culture to detect toxic tRNase activity by Northern blot hybridization60, 61. To facilitate this analysis, we over-expressed tRNACCU Arg in the target-cell population. Because this substrate is present at very low levels in wild-type E. coli 62, essentially all of the tRNACCU Arg detected by Northern blot is derived from target bacteria. Most of the tRNACCU Arg substrate was cleaved within 1 h of co-culture (Fig. 3B, compare lanes 1 & 5); and this nuclease activity was dependent on EcCysK, because substrate was not degraded when ∆cysK mutants were used as target bacteria (Fig. 3B, lane 8). We then over-produced EcCysE in target cells and examined the effect on toxin activity. Notably, EcCysE was readily detected by SDS-PAGE analysis of crude lysates prepared from the co-culture (Fig. 3B, bottom panel), indicating that target cells likely contained enough EcCysE to saturate endogenous EcCysK. As predicted, over-produced EcCysE suppressed toxin activity, but substantial tRNA degradation was still detected in target cells (Fig. 3B, lane 6). By contrast, an EcCysE variant lacking 11 residues from the C-terminus was less effective in blocking toxin activity (Fig. 3B, lane 7), consistent with the importance of these residues in CS complex stability. We obtained similar results with in vitro tRNase assays. As reported previously16, CdiA-CT has no appreciable nuclease activity in vitro, but efficiently cleaves tRNA when reactions are supplemented with EcCysK (Fig. 3C, compare lanes 2 & 3). To examine the effect of EcCysE on nuclease activity, we pre-incubated EcCysK with EcCysE for 20 min to assemble cysteine synthase complexes. CdiA-CT was then added, and the protein mixture incubated for an additional 30 min prior to the addition of tRNA substrate. Even when used in eight-fold excess over EcCysK (with respect to cysteine synthase stoichiometry), EcCysE did not block tRNase activity to the same extent as CdiI immunity protein (Fig. 3C, compare lanes 6 & 7). Together, these results demonstrate that CdiA-CT toxin is activated efficiently even in the presence of EcCysE.
The homodimeric structure of EcCysK provides a possible explanation for robust CdiA-CT activation in the presence of competing EcCysE. We reasoned that if only one EcCysK active site per dimer is occupied by EcCysE in the cysteine synthase complex, then the other active site should be available to bind toxin. This model predicts that EcCysK can bind EcCysE and CdiA-CT simultaneously. To explore this hypothesis, we sought to isolate EcCysE:EcCysK:CdiA-CT ternary complexes. We equilibrated His6-tagged CdiA-CT with untagged EcCysK and EcCysE for 1 h, then subjected the mixture to Ni2+-affinity chromatography to purify the toxin and associated proteins. EcCysK clearly interacted with His6-CdiA-CT under these conditions, but none of the EcCysE co-purified with the His6-CdiA-CT:EcCysK complex, even at concentrations up to 15 µM (Fig. 4A). Instead, there appeared to be competition for EcCysK occupancy, with much of the EcCysK remaining in the “free” fraction due to its association with EcCysE (Fig. 4A). These results show that high-affinity ternary complexes of EcCysE:EcCysK:CdiA-CT do not form, indicating that the binding of toxin and EcCysE to EcCysK is mutually exclusive.

EcCysE and CdiA-CT compete for binding to CysK. (A) Affinity purification of EcCysK:CdiA-CT complexes. EcCysK, EcCysE and His6-tagged CdiA-CT were mixed at the indicated concentrations and subjected to Ni2+-affinity chromatography. The initial protein mixture (input), the column flow-through fraction (free) and the elution fraction (bound) were analyzed by SDS-PAGE. (B,C) Native PAGE analysis of EcCysK:EcCysE and EcCysK:CdiA-CT complexes. Proteins were mixed and run on non-denaturing polyacrylamide gels as described in the Methods. Proteins within parentheses were pre-incubated for 10, 20 or 40 min (as indicated by subscript) prior to native PAGE analysis. The migration positions of EcCysE, EcCysK and CdiA-CT are indicated on the left of panel B, and the migration positions of the cysteine synthase (CS) and EcCysK:CdiA-CT (CTox) complexes are indicated on the right.
We then developed a native PAGE approach to monitor the relative proportions of EcCysK:EcCysE and EcCysK:CdiA-CT in complex mixtures. Electrophoresis conditions were optimized to allow unambiguous identification of each complex based on its gel mobility (Fig. 4B, compare lanes 2 & 4). Simultaneous mixing of EcCysE, EcCysK and CdiA-CT resulted in the formation of both complexes, with lower levels (39%) of EcCysK:CdiA-CT with respect to CS (61%) (Fig. 4B, lane 3, Fig. 4C, lane 1). A similar result was obtained when EcCysE was pre-incubated with EcCysK (Fig. 4C, lane 5). By contrast, the proportion of CdiA-CT:EcCysK complex increased significantly (75%) when toxin and EcCysK were pre-incubated before the EcCysE addition (Fig. 4C, lane 6). This latter observation suggests that the CdiA-CT:EcCysK complex reaches equilibrium more slowly than CS. The finding is further supported by a time-driven experiment where EcCysE, EcCysK and CdiA-CT are mixed simultaneously and monitored over time. The proportion of EcCysK:CdiA-CT complex increased from about 40% to 60% after 40 min of incubation (Fig. 4C, lanes 1–4 and Supplemental Fig. S2). This result confirms that equilibrium conditions are reached slowly when EcCysE and CdiA-CT compete for EcCysK occupancy, and further suggests that CdiA-CT might displace EcCysE from the CS complex.
CdiA-CT toxin is activated by CysK from diverse bacterial species
CdiA-CT is a member of the Ntox28 RNase family and closely related toxin domains are found in CdiA proteins from Enterobacter cloacae, Yersinia enterocolitica and Pseudomonas syringae 14, 15, 42, 43. These observations suggest that CdiA-CT interacts with other CysK enzymes to ensure activation in different bacterial species. We tested this prediction using a previously described plasmid-transformation assay16, 60. In this assay, separate plasmids that express CdiA-CT or CysK are simultaneously introduced into E. coli ∆cysK cells, and transformants are selected on antibiotic-supplemented media. Because CdiA-CT is toxic when bound to EcCysK, cells that take up both plasmids are unable to grow, and therefore stable transformants are not obtained even when toxin expression is repressed with D-glucose in the media (Fig. 5A). To control for transformation efficiency, we introduced a catalytically inactive CdiA-CT construct carrying the His178Ala mutation and obtained several transformants (Fig. 5A)16. We then tested plasmids encoding heterologous enzymes that share between 50% and 96% sequence identity with EcCysK. These CysK homologs share virtually identical active sites, and 8 of the 13 residues that make direct contact with the toxin domain are conserved (Fig. S3). As expected, closely related enzymes from Enterobacter cloacae (ECLCysK, 96% identity) and Dickeya dadantii (DdCysK, 91% identity) promoted CdiA-CT toxicity in the transformation assay (Fig. 5A). More distantly related CysK proteins from Haemophilus influenzae (HiCysK, 68% overall identity, 92% identity in toxin-binding residues) and Bacillus subtilis (BsCysK, 50% overall identity, 92% identity in toxin-binding residues) also activated the toxin in vivo (Fig. 5A). However, NlCysK from Neisseria lactamica (53% identity, 69% identity in toxin-binding residues) only supported toxicity when its expression was fully induced with L-arabinose (Fig. 5A, compare glucose and arabinose plates). This latter result suggests that CdiA-CT has significantly lower affinity for NlCysK. Similar results were obtained when we tested the function of heterologous CysK in CDI competition co-cultures. We provided E. coli ∆cysK target cells with cysK-his 6 expression plasmids and incubated the resulting strains with inhibitor bacteria that deploy CdiA-CT. Growth inhibition was assessed by enumerating viable target bacteria after three hours of co-culture. Target cells lacking CysK were resistant to growth inhibition, and viable cell counts increased during the incubation (Fig. 5B). By contrast, target cells expressing EcCysK, DdCysK and HiCysK were inhibited, with each population showing ~100-fold losses in viability (Fig. 5B). The other CysK enzymes were less effective at promoting toxicity, particularly NlCysK, which showed less than a ten-fold decrease in viable cell counts (Fig. 5B). To ascertain the levels of heterologous CysK in target cells, we performed immunoblot analysis using antibodies to the His6 epitope appended to the C-terminus of each enzyme. This analysis revealed lower levels of BsCysK and NlCysK (Fig. 5C), perhaps accounting for the resistance of NlCysK expressing cells to growth inhibition.

Heterologous CysK promotes CdiA-CT toxicity. (A) Activation of internally expressed CdiA-CT toxin. Plasmids encoding CdiA-CT and CysK proteins were introduced into E. coli ∆cysK cells, and transformants isolated on selective media supplemented with D-glucose or L-arabinose as indicated. To control for transformation efficiency, a plasmid encoding catalytically inactive CdiA-CT(H178A) was also tested. (B) CDI competition co-cultures. Inhibitor cells that deploy CdiA-CT were incubated with E. coli ∆cysK target bacteria that express CysK-His6 from the indicated bacterial species. Viable target bacteria were quantified as colony forming units per mL upon mixing and after 3 h of co-culture. Presented data are averages ± standard errors for four independent experiments. (C) Immunoblot analysis of heterologous CysK-His6. Total protein was isolated from the target-cell strains in panel B and analyzed by immunoblotting using antibodies to the His6 epitope. 10 μg of total protein was loaded in each lane.
Finally, we examined toxin binding and activation by heterologous CysK in vitro. We first used affinity co-purification to screen interactions between CdiA-CT and His6-tagged CysK proteins. This approach showed that DdCysK, ECLCysK and BsCysK all form high-affinity complexes with CdiA-CT (Fig. 6A). Because the toxin failed to co-purify with HiCysK and NlCysK (Fig. 6A), we quantified the binding interactions using fluorimetric titrations and determined dissociation constants of 3.3 ± 0.3 µM for the HiCysK:CdiA-CT complex and 6.4 ± 0.6 µM for NlCysK:CdiA-CT (Fig. 6B). Thus, CdiA-CT binds these latter enzymes with ~1,000-fold lower affinity than EcCysK. Consistent with this low affinity, high concentrations of HiCysK and NlCysK were required to activate the CdiA-CT nuclease in vitro (Fig. 6C). As we found in the in vivo analyses, NlCysK was the least effective at promoting toxin activity. In fact, tRNase reactions supplemented with NlCysK up to 10 µM did not go to completion after 1 h incubation. Together, these results show that CdiA-CT toxin can be activated by a variety of CysK enzymes, but the binding constants span several orders of magnitude.

In vitro activation of CdiA-CT by heterologous CysK. (A) Affinity purification of CysK:CdiA-CT complexes. His6-tagged CysK proteins from the indicated species were incubated with untagged CdiA-CT and the mixture subjected to Ni2+-affinity chromatography. The initial protein mixture (input), the column flow-through fraction (free) and the elution fraction (bound) were analyzed by SDS-PAGE. (B) Determination of the CysK:CdiA-CT dissociation constants. HiCysK (300 nM) and NlCysK (860 nM) were titrated with increasing concentrations of CdiA-CT, and fluorescence emission at 500 nm monitored after 20 min. Eq. 1 was fitted to the binding data to obtain K d = 3.3 ± 0.3 µM for HiCysK and K d = 6.4 ± 0.6 µM for NlCysK. (C) Heterologous CysK promotes CdiA-CT nuclease activity. Proteins were incubated at the indicated concentrations (µM) for 20 min prior to the addition of RNA substrate. Reactions were quenched after 1 h at 37 °C, then run on 8 M urea-polyacrylamide gels and visualized by ethidium bromide staining. The migration position of full-length tRNA is indicated.
Discussion
Here, we show that EcCysK and CdiA-CT form a high-affinity complex with two toxin domains bound per EcCysK homodimer. These results are broadly consistent with a prior thermodynamic study by Kaundal et al.57, though our data indicate that CdiA-CT could displace EcCysK from pre-formed CS complexes and suggest that CdiI has less of an effect on binding affinity. However, both studies show that complex formation is remarkably slow. Kaundal et al. used surface plasmon resonance to measure a kon of 6.2·103 M−1·s−1 57, and here we calculate a second-order rate constant of 2·104 M−1·s−1 for EcCysK:CdiA-CT binding, consistent with slow conformational rearrangements following the formation of an encounter complex. The cysteine synthase complex also exhibits a two-step binding mechanism56, which is perhaps not surprising given that the CdiA-CT toxin mimics CysE by inserting its C-terminus into the CysK active site, anchoring the interaction to allow further conformational changes. For the CS complex, the slow conformational changes entail closure of the CysK active site38, 56; but allosteric changes in CysE are also likely because its O-acetyltransferase activity is stimulated in the E. coli complex (see Fig. 3A). We note that EcCysK adopts an open active-site conformation in the EcCysK:CdiA-CT crystal structure42, indicating that the toxin does not induce significant structural changes in EcCysK. Together, these observations suggest that conformational changes in CdiA-CT are responsible for the slow phase of complex formation. This model also accounts for toxin activation, whereby EcCysK-induced structural changes organize the nuclease active site for catalysis. CdiA-CT is probably delivered in a partially unfolded state (vide infra) and folding to the final active conformation likely represents the slow, rate limiting step in complex formation. Reversal of this isomerization, which is described by the k4 rate constant is even slower (see Fig. 2E). Direct determination of k4 is hindered by the intrinsically high error in the calculation of the y-axis intercept. However, Eq. 8 can be used to estimate the rate constant for the reversal of isomerization at 2.2·10−4 s−1, which is two orders of magnitude slower than the corresponding value of 0.024 s−1 for the CS complex56 and in good agreement with the overall koff calculated by SPR57. Thus, the activated toxin complex dissociates exceptionally slowly, prolonging nuclease activity in target cells. Such long residence times have been observed for antigen-antibody and protease-inhibitor interactions and are important in open biological systems, where ligand concentrations vary over time63. For many protein-ligand interactions, binding efficacy can be explained entirely by the koff value rather than dissociation constant alone64.
EcCysE and CdiA-CT bind with comparable affinities to the same site on EcCysK, indicating that the toxin must compete with EcCysE in order to be activated. Further, the toxin is presumably at a disadvantage with respect to endogenous EcCysE, because only a few CdiA-CT domains are delivered into target cells during CDI65. This is compounded by the fact that the second-order rate constant for CS complex formation is ~200-fold greater than that of the EcCysK:CdiA-CT complex. However, early studies in Salmonella Typhimurium suggested that StCysK levels exceed those of StCysE, with only 5–25% of StCysK found in the CS complex66, 67. On the other hand, cysE and cysK are regulated by different transcription factors, raising the possibility that their relative proportions are modulated in response to changing growth conditions. For example, high cysteine levels inhibit CysE activity68, 69, reducing the production of O-acetyl-L-Ser and N-acetyl-L-Ser, which are required as co-activators to induce CysB-dependent transcription of the cys regulon17. Because cysE transcription is not regulated by CysB, it is possible that CysE becomes more abundant than CysK when the cell is replete with cysteine. Moreover, recent transcriptomic data show that cysK and cysE transcript levels are comparable in S. Typhimurium cells grown in rich media and other conditions70. Thus, CdiA-CT activity and CDI could be modulated by environmental conditions, though we have found that target bacteria are still inhibited in cysteine supplemented media (C.M.B. & C.S.H., unpublished data). These observations indicate that EcCysE levels are no impediment to toxin activation. Moreover, the data presented here show that even supra-physiological EcCysE concentrations are insufficient to block toxin activation. Thus, CdiA-CT competes effectively with EcCysE, and may even displace EcCysK from pre-formed CS complexes. There are no structures available for the CS complex, but biochemical studies indicate that each CysE hexamer engages two CysK dimers (Fig. 7). Further, molecular modeling shows that distance and geometrical constraints prevent CysE from engaging both CysK active sites simultaneously71. Therefore, only one active site per CysK dimer is engaged with CysE in the CS complex (Fig. 7). This architecture provides opportunities for CdiA-CT to bind the unoccupied CysK active site. However, stable EcCysE:EcCysK:CdiA-CT ternary complexes cannot be isolated, suggesting that the binding of CdiA-CT and EcCysE to EcCysK is mutually exclusive. Several studies indicate that the CysK active site undergoes allosteric closure in the CS complex38, 56, whereas CdiA-CT binds to EcCysK with an open active-site conformation (Fig. 7)42. Thus, differential affinities for the open and closed states could account for the observed binding behavior. This model may also explain how CdiA-CT disrupts the CS complex, though we note that because the two complexes have similar affinities, their proportions at equilibrium should reflect the relative concentrations of toxin and EcCysE.

Formation of CS and CysK:CdiA-CT complexes. CysK exists in two conformations: an open, inactive conformation and a closed one that was isolated as a complex with substrate analog31. Structural data42 demonstrate that CdiA-CT binds to the open conformation of CysK. Binding is slow and is limited by a large conformational change, which likely corresponds to CdiA-CT folding to its active conformation. Here, we propose that CysE preferentially binds to CysK in the closed active-site conformation.
The CdiA-CT toxin from E. coli 536 has evolved a dependence on CysK, but most other CDI toxins do not require additional factors to promote toxicity12, 13, 72,73,74. Moreover, the benefit of extrinsic activation is not clear in the context of inter-bacterial conflict, because target bacteria can readily acquire resistance through cysK mutations16, 60. One explanation invokes possible physical constraints on CDI toxin delivery, which entails CdiA-CT translocation across the outer and inner membranes of target bacteria. Though the mechanistic basis of CDI toxin transfer is not completely understood, the analogous import of colicins into E. coli requires the unfolding of toxin domains75, 76. If CDI toxins must also unfold during delivery, then there should be a selective pressure for domains with low global stability. This in turn could provide the impetus to evolve binding interactions that compensate for intrinsic instability. Consistent with this hypothesis, CdiA-CT has relatively low thermostability and is significantly stabilized when bound to EcCysK42, 57. Thus, EcCysK-binding could ensure that the toxin regains its native fold after delivery into the target-cell cytoplasm. In principle, the CdiA-CT toxin could have evolved binding interactions with any number of cytosolic proteins, but it appears that CysK was selected due to its conservation throughout bacteria. Although uropathogenic E. coli are unable to deliver the CdiA-CT toxin into other bacterial species45, 46, there is still a selective pressure for activation in diverse bacteria because CDI systems are encoded on mobile genetic elements and are spread by horizontal gene transfer47, 48. Database searches reveal closely related toxin domains (>60% sequence identity) in CdiA proteins from various Enterobacteriaceae and Pseudomonads. By targeting the conserved active-site cleft of CysK, the toxin is likely to be activated in the cytosol of any given species. Finally, we note that this phenomenon appears to be widespread, because unrelated CDI toxins from E. coli isolates EC869, NC101 and 96.154 have recently been shown to interact functionally with the highly conserved translation factors EF-Tu and EF-Ts77.
Methods
Bacterial strains and plasmid constructions
Bacterial strains and plasmids are listed in Table 1. Bacteria were grown in lysogeny broth (LB) or on LB agar unless otherwise noted. Where indicated, media were supplemented with antibiotics at the following concentrations: ampicillin, 150 µg mL−1; kanamycin, 50 µg mL−1; rifampicin, 200 µg mL−1; and tetracycline, 12.5 µg mL−1. The ∆cysK::kan disruption was obtained from the Keio collection78 and transduced into E. coli strains MG1655 (DE3) and CH10013. Kanamycin-resistance cassettes were subsequently removed with FLP recombinase expressed from plasmid pCP2079. Bacterial cysK open-reading frames were amplified by PCR using the following primer pairs: CH2095/CH2102 for D. dadantii 3937, CH2101/CH2099 for E. cloacae ATCC 13047, CH3466/CH3467 for H. influenzae Rd, CH3345/CH3346 for N. lactamica ATCC 23970, and CH2096/CH2094 for B. subtilis 168 (Table S1). The resulting products were digested with NcoI/SpeI, then ligated to plasmid pCH6505 to generate T7 over-expression constructs, and to plasmid pCH6478 for complementation of E. coli ∆cysK mutants. The N. lactamica cysK fragment was ligated using NcoI/XhoI restriction sites. E. coli cysE was amplified with primers CH3642/CDI235 and ligated to pET21P with KpnI/XhoI restiction sites to generate pCH12028 for the gratuitous over-production of native EcCysE in target cells. This fragment was also ligated to pCH1006880 to generate plasmid pCH9764 for the purification of untagged EcCysE. Primers CH3642/CH4125 were used to generate plasmid pCH13299, which over-produces EcCysE lacking 11 residues from the C-terminus. The E. coli cysK gene was amplified with primers CH3865/CH2797 and ligated to pCH10068 for the purification of untagged EcCysK.
Protein expression and purification
Proteins were over-produced in E. coli BL21(DE3) TunerTM or CH2016 cells grown in LB media supplemented with 1 mM isopropyl β-D-1-thiogalactopyranoside (IPTG). Cells were resuspended in buffer A [20 mM sodium phosphate (pH 7.0), 85 mM sodium chloride, 10 mM 2-mercaptoethanol, 2 mM EDTA] and broken by sonication or French pressure cell. His6-tagged proteins were purified by Ni2+- or Co2+-affinity chromatography according to85 with minor modifications. The His6 epitope tag was removed from HiCysK using thrombin36. CysK concentrations were determined by PLP absorbance, calculated by the alkali denaturation method86. Extinction coefficients at 412 nm are 7,600 M−1·cm−1 for HiCysK, 9,370 M−1·cm−1 for EcCysK, and 8,280 M−1·cm−1 for NlCysK. Purity was assessed by SDS-PAGE and demonstrated to be greater than 97% (Fig. S4A). All enzymes showed the typical absorption spectrum of fold-type II PLP-dependent enzymes with peaks at 278 and 412 nm and a specific activity of 0.025 U/mg, 0.013 U/mg and 0.016 U/mg for EcCysK, HiCysK and NlCysK (respectively) in agreement with previously reported kinetic data23.
EcCysE was over-produced as a fusion with His6-thioredoxin (His6-TrxA) linked by a TEV protease recognition sequence. Affinity resins were washed with buffer containing 10 mM O-acetyl-L-Ser to dissociate contaminating EcCysK. His6-TrxA-EcCysE was eluted with 1 M imidazole and dialyzed against 20 mM Tris-HCl (pH 7.5), 50 mM NaCl, 1% glycerol, 1 mM dithiothreitol, 1 mM EDTA. The fusion was digested with His6-tagged TEV protease for 4 h at 25 °C, and the His6-TrxA and His6-TEV proteins were removed by metal-affinity chromatography. EcCysE was loaded on a FPLC column packed with Ultrogel AcA44 resin (exclusion limit 200 kDa, operating range 17–175 kDa, column volume 63 mL and void volume 20.4 mL) and run at 0.2 mL/min in buffer A. EcCysE eluted at 28 mL with an apparent molecular mass of 167,200 Da, consistent with the expected hexameric quaternary structure. Protein concentration was calculated using an extinction coefficient at 278 nm of 26,900 M−1 cm−1. Purified EcCysE was ~96% pure (Fig. S4A), with a specific activity of 290 U/mg in agreement with previously published reports87, 88.
The CdiA-CT:CdiI-His6 complex was expressed in either E. coli BL21(DE3) TunerTM or CH2016 as described11. CdiA-CT and CdiI-His6 were separated by metal-affinity chromatography in 8 M urea and the proteins refolded by dialysis into buffer A. The isolated proteins were greater than 95% pure (Fig. S4A), and circular dichroism spectroscopy showed that each protein regained native structure under these conditions (Fig. S4B). CdiA-CT was further purified using size-exclusion chromatography as described for EcCysE above. Protein concentration was estimated using an extinction coefficient at 278 nm of 13,300 M−1·cm−1 and 8,480 M−1·cm−1 for CdiA-CT and CdiI, respectively.
Spectroscopy
Absorption spectra were collected at 20.0 ± 0.5 °C using a Varian CARY400 spectrophotometer. All spectra were corrected for buffer contributions. Circular dichroism measurements were carried out using a JASCO J-715 spectropolarimeter. Each spectrum is the average of three measurements and is subtracted of the buffer contribution. EcCysE/CdiA-CT binding to CysK was monitored by measuring PLP fluorescence emission at 500 nm following excitation at 412 nm38, 56. CysK emission spectra were collected using a FluoroMax-3 fluorometer (HORIBA) at 20 ± 0.5 °C. Unless otherwise specified, titration samples were equilibrated for 5 min prior to spectra acquisition. All spectra were corrected for buffer contribution, and the slit width set to optimize the signal to noise ratio. For equilibrium binding experiments, the dependence of emission intensity on ligand concentration was determined using the binding isotherm:
or a quadratic equation that describes tight binding:
where I is the fluorescence intensity at 500 nm, I 0 is a horizontal offset, I max is the maximum change in fluorescence at saturating [CdiA-CT] and K d is the dissociation constant for the CysK:CdiA-CT complex.
Enzyme activity assays
CysK specific activities were quantified by a continuous spectrophotometric assay using 2-thio-5-nitrobenzoate (TNB) as a nucleophilic substrate23. EcCysE specific activity was determined indirectly with 5,5′-dithio-bis(2-nitrobenzoic acid) (DTNB) as described89. An extinction coefficient of 14,150 M−1·cm−1 at 412 nm was used to quantify TNB in both enzyme assays90. EcCysE steady-state kinetics were measured by an adaptation of a published method91 in buffer A without 2-mercaptoethanol at 20 °C. Briefly, O-acetylation of 20 mM L-Ser was monitored by measuring the absorption at 232 nm of the thioester bond (ε232 = 4,440 M−1 cm−1), while varying acetyl-CoA concentrations. At a fixed 0.25 mM acetyl-CoA concentration, EcCysE activity increases as a function of EcCysK concentration55. Displacement of EcCysE from the cysteine synthase complex by CdiA-CT was monitored using 28 nM CysE (4.7 nM hexamer) in the presence of 19 nM CysK (9.5 nM dimer). EcCysK and CdiA-CT were incubated for 20 min at 20 °C, then EcCysE and L-Ser were added, and the reaction was initiated with the addition of acetyl-CoA.
CysK steady-state kinetics were measured by quantifying L-Cys using the discontinuous method of Gaitonde in a 96-well plate format92. The sulfhydrylase reaction was initiated by addition of 0.6 mM Na2S to a solution containing 6 nM EcCysK, 60 nM bovine serum albumin and variable concentrations of O-acetyl-L-Ser in buffer A. Aliquots (60 µL) were removed at intervals and quenched with 60 µL of acetic acid in a PCR tube strip. Ninhydrin (60 µL) was added with a multichannel pipette and the mixture heated at 100 °C for 10 min in a thermal cycler. The solution was cooled down and 46 µL were added to the wells of a 96-well plate containing 154 µL of cold ethanol. The absorbance at 550 nm was measured using a plate reader and blanks subtracted. The amount of L-Cys produced at each time point was calculated from a calibration curve and a linear equation was fitted to the data to determine initial rate (v i ) of production. All kinetic data sets were collected from at least two independent experiments. The kinetic parameters were calculated as follows: kcat = 241 ± 5 s−1, KM,OAS = 5.1 ± 0.3 mM and KM,HS − = 0.006 ± 0.003 mM. The dependence of the initial velocity on either EcCysE or CdiA-CT concentration was measured in buffer A containing 2 mM O-acetyl-L-Ser. Morrison’s equation (3) was used to calculate IC 50 and hence the K i for tight-binding inhibitors63:
where [E]T is the total enzyme concentration, and [I]T is the total inhibitor concentration (EcCysE or CdiA-CT). For competitive inhibitors of a ping-pong reaction63, 93, 94:
Pre-steady state binding kinetics
Pre-steady state kinetic traces were collected under similar conditions to those reported in ref. 56. Experiments were carried out in buffer A under pseudo-first order conditions with 200 nM EcCysK, 270 nM CdiA-CT or 400 nM EcCysE. The temperature of the loading syringes and the stopped-flow cell compartment was maintained constant with a circulating water bath. Kinetic traces were collected upon direct excitation of PLP at 412 nm using an SX-18MV apparatus (Applied Photophysics) equipped with a 75-watt xenon lamp as a light source and a photomultiplier as a detector. The emission signal was collected at 90° with respect to the excitation source and filtered below 440 nm by a cut-off filter. A single exponential equation
was fitted to data averaged from three to five kinetic traces. I t and I 0 are the emission values at a given time and at zero time, respectively; I is the total fluorescence change, and τ is the relaxation time, such that k obs is 1/τ. The dependence of kobs on protein concentrations was obtained from the linear equation:
to account for a two-step mechanism with a slow conformational change where the plateau cannot be attained under the experimental conditions56, 59.The K d in Eq. 6 accounts for the first step (i.e. encounter complex formation) of a two-step binding reaction. The dissociation constant as measured under equilibrium conditions accounts for the contributions of both binding and isomerization steps and is usually indicated as \({K}_{d}^{overall}\), indeed for a slow binding mechanism, where k4<<k3 56, 59:
If k3/K d and \({K}_{d}^{{overall}}\) are known, then k4 can be calculated as follows:
Complex co-purification and native gel electrophoresis
Purified EcCysK (5 µM) and His6-tagged CdiA-CT (5 µM) were incubated with EcCysE (5 or 15 µM) in 20 mM sodium phosphate (pH 7.5), 140 mM NaCl for one h at room temperature. A sample of the mixture was removed (for subsequent SDS-PAGE analysis) and the remainder subjected to Ni2+-affinity chromatography as described16. Samples of the original mixture (input), the column void (free) and imidazole elution (bound) were analyzed by SDS-PAGE and proteins detected with Coomassie blue stain. The same procedure was used to screen for stable interactions between bacterial CysK-His6 proteins and untagged CdiA-CT. Native gel electrophoresis was used to detect cysteine synthase and activated toxin complexes in mixtures. Purified EcCysK (12 µM monomer), EcCysE (18 µM monomer) and CdiA-CT (12 µM) were mixed in various combinations and the resulting complexes resolved on 8% polyacrylamide gels run at 10 mA constant current and 4 °C. The gel running buffer was 5 mM sodium phosphate (pH 7.0) and proteins were detected with Coomassie blue stain. Native-PAGE gels were analyzed using Image Lab™ software (version 5.2.1, Bio-Rad). Software auto analysis procedure was applied to detect lanes and bands with manual adjustments. The exposure time was set to 0.074 s. The relative intensity of each band was calculated using the band % parameter, which calculates band volume as percentage of the total band volume for each sample lane.
Competition co-cultures and in vivo toxin activity
E. coli EPI100 cells that deploy CdiA-CT from plasmid pCH10673 were used as inhibitors in experiments to determine the effect of EcCysE over-production on toxin activation in target bacteria. Inhibitors were mixed at a 1:1 ratio with E. coli CH7076 (cysK +) or CH13316 (∆cysK) target cells that overexpress tRNACCU Arg. Target cells also harbored plasmids pET21P, pCH12028 or pCH13299 (where indicated), and were induced with 1 mM IPTG to allow EcCysE accumulation for 30 min prior to mixing with inhibitor cells. Samples were harvested into an equal volume of ice-cold methanol upon initial cell mixing and after 1 h of co-culture. Cells were collected by centrifugation at 4 °C and frozen at −80 °C. RNA was extracted from frozen cell pellets with guanidinium isothiocyanate-phenol as described previously81. RNA was resolved on 50% urea – 10% polyacrylamide gels and electro-blotted to nylon membrane and hybridized to 5′-radiolabeled oligonucleotide (5′ – CCT GCA ATT AGC CCT TAG G)83. Protein was isolated from co-culture samples with two freeze-thaw cycles in urea lysis buffer [8 M urea, 50 mM Tris-HCl (pH 8.0), 150 mM NaCl]. Urea-soluble protein was quantified by Bradford assay and 10 µg resolved on SDS-polyacrylamide gels. Proteins were detected with Coomassie blue stain.
Plasmid co-transformation was used to assess CdiA-CT toxicity in combination with heterologous CysK enzymes. Arabinose-inducible CdiA-CT expression plasmids (100 ng) were introduced into E. coli ∆cysK cells together with plasmids pTrc99A (no CysK), pCH9280 (EcCysK), pCH11860 (NlCysK), pCH12113 (HiCysK), pCH12286 (DdCysK), pCH12287 (ECLCysK) or pCH12288 (BsCysK). After recovery for 1 h at 37 °C in LB media supplemented with 0.4% D-glucose, cells were plated onto LB-agar supplemented with Tet, Amp and 0.4% D-glucose or L-arabinose to select for transformants carrying both plasmids.
E. coli EPI100 inhibitors that express the cdiBAI EC536 gene cluster from pDAL866 were used in competition co-cultures to test complementation with heterologous cysK. E. coli CH10801 (∆cysK) target cells harboring the various cysK expression plasmids were grown to mid-log phase in LB media supplemented with ampicillin, then mixed at a 1:10 ratio with inhibitor cells in LB medium supplemented with 0.2% L-arabinose and incubated for 3 h at 37 °C with vigorous shaking in baffled flasks. Viable target-cell counts were enumerated as colony forming units (cfu) mL−1 on LB-agar supplemented with rifampicin. Data are presented as averages ± standard errors for four independent experiments. Heterologous CysK levels were monitored by immunoblot analysis. Total protein was isolated from target-cell strains using urea lysis as described above. Proteins were resolved by SDS-PAGE, electro-blotted onto nitrocellulose, and CysK detected with polyclonal antibodies to the C-terminal His6 epitope. Immunoblots were visualized using IRDye® 680 (LI-COR) labeled anti-rabbit secondary antibodies and an Odyssey® infrared imager as described previously95.
In vitro nuclease assays
CdiA-CT tRNase activity was assayed in 20 mM Tris-HCl (pH 7.5), 150 mM NaCl, 0.5 mM MgCl2 at 37 °C. To determine the influence of EcCysE on nuclease activity, EcCysK (0.5 µM) was pre-incubated with EcCysE (1.5 to 6 µM) for 25 min prior to the addition of CdiA-CT (0.5 µM). After further incubation for 25 min, reactions were initiated by addition of total E. coli RNA to a final concentration of 2 µg µL−1. Reactions were quenched with SDS-formamide gel-loading buffer after 10 min. CysK (0.1 to 10 µM) enzymes from other bacteria species were assayed in the same manner, except that CdiA-CT was used at 1 µM final concentration and the reactions were quenched after 1 h. All reactions were run on 8 M urea, Tris-borate-EDTA polyacrylamide gels, and RNA visualized by ethidium bromide staining.
References
Waksman, S. The role of antibiotics in nature. Perspect Biol Med 4, 271–286 (1961).
Cascales, E. et al. Colicin biology. Microbiol Mol Biol Rev 71, 158–229 (2007).
Severinov, K., Semenova, E., Kazakov, A., Kazakov, T. & Gelfand, M. S. Low-molecular-weight post-translationally modified microcins. Mol Microbiol 65, 1380–94 (2007).
Gabrielsen, C., Brede, D. A., Nes, I. F. & Diep, D. B. Circular bacteriocins: biosynthesis and mode of action. Appl Environ Microbiol 80, 6854–62 (2014).
Aoki, S. K. et al. Contact-dependent inhibition of growth in Escherichia coli. Science 309, 1245–8 (2005).
Hood, R. D. et al. A type VI secretion system of Pseudomonas aeruginosa targets a toxin to bacteria. Cell Host Microbe 7, 25–37 (2010).
MacIntyre, D. L., Miyata, S. T., Kitaoka, M. & Pukatzki, S. The Vibrio cholerae type VI secretion system displays antimicrobial properties. Proc Natl Acad Sci USA 107, 19520–4 (2010).
Souza, D. P. et al. Bacterial killing via a type IV secretion system. Nat Commun 6, 6453 (2015).
Ruhe, Z. C., Low, D. A. & Hayes, C. S. Bacterial contact-dependent growth inhibition. Trends Microbiol 21, 230–7 (2013).
Willett, J. L., Ruhe, Z. C., Goulding, C. W., Low, D. A. & Hayes, C. S. Contact-Dependent Growth Inhibition (CDI) and CdiB/CdiA Two-Partner Secretion Proteins. J Mol Biol 427, 3754–65 (2015).
Aoki, S. K. et al. A widespread family of polymorphic contact-dependent toxin delivery systems in bacteria. Nature 468, 439–42 (2010).
Morse, R. P. et al. Structural basis of toxicity and immunity in contact-dependent growth inhibition (CDI) systems. Proc Natl Acad Sci USA 109, 21480–21485 (2012).
Nikolakakis, K. et al. The toxin/immunity network of Burkholderia pseudomallei contact-dependent growth inhibition (CDI) systems. Mol Microbiol 84, 516–29 (2012).
Zhang, D., de Souza, R. F., Anantharaman, V., Iyer, L. M. & Aravind, L. Polymorphic toxin systems: Comprehensive characterization of trafficking modes, processing, mechanisms of action, immunity and ecology using comparative genomics. Biol Direct 7, 18 (2012).
Zhang, D., Iyer, L. M. & Aravind, L. A novel immunity system for bacterial nucleic acid degrading toxins and its recruitment in various eukaryotic and DNA viral systems. Nucleic Acids Res 39, 4532–52 (2011).
Diner, E. J., Beck, C. M., Webb, J. S., Low, D. A. & Hayes, C. S. Identification of a target cell permissive factor required for contact-dependent growth inhibition (CDI). Genes Dev 26, 515–25 (2012).
Kredich, N. M. Biosynthesis of cysteine. In Escherichia coli and Salmonella (ed. Neidhardt, F.C.) 514–527 (ASM Press, Washington, 1996).
Sekowska, A., Kung, H. F. & Danchin, A. Sulfur metabolism in Escherichia coli and related bacteria: facts and fiction. J Mol Microbiol Biotechnol 2, 145–77 (2000).
Schelle, M. W. & Bertozzi, C. R. Sulfate metabolism in mycobacteria. ChemBioChem 7, 1516–24 (2006).
Cook, P. F. & Wedding, R. T. A reaction mechanism from steady state kinetic studies for O-acetylserine sulfhydrylase from Salmonella typhimurium LT-2. J Biol Chem 251, 2023–9 (1976).
Cook, P. F. & Wedding, R. T. Initial kinetic characterization of the multienzyme complex, cysteine synthetase. Arch Biochem Biophys 178, 293–302 (1977).
Cook, P. F. & Wedding, R. T. Overall mechanism and rate equation for O-acetylserine sulfhydrylase. J Biol Chem 252, 3459 (1977).
Tai, C. H., Nalabolu, S. R., Jacobson, T. M., Minter, D. E. & Cook, P. F. Kinetic mechanisms of the A and B isozymes of O-acetylserine sulfhydrylase from Salmonella typhimurium LT-2 using the natural and alternative reactants. Biochemistry 32, 6433–42 (1993).
Schnackerz, K. D. et al. Identification and spectral characterization of the external aldimine of the O-acetylserine sulfhydrylase reaction. Biochemistry 34, 12152–60 (1995).
Woehl, E. U., Tai, C. H., Dunn, M. F. & Cook, P. F. Formation of the alpha-aminoacrylate immediate limits the overall reaction catalyzed by O-acetylserine sulfhydrylase. Biochemistry 35, 4776–83 (1996).
Tai, C. H. & Cook, P. F. O-acetylserine sulfhydrylase. Adv Enzymol Relat Areas Mol Biol 74, 185–234 (2000).
Tai, C. H. et al. Characterization of the allosteric anion-binding site of O-acetylserine sulfhydrylase. Biochemistry 40, 7446–52 (2001).
Mozzarelli, A. et al. The multifaceted pyridoxal 5′-phosphate-dependent O-acetylserine sulfhydrylase. Biochim Biophys Acta 1814, 1497–510 (2011).
Burkhard, P. et al. Three-dimensional structure of O-acetylserine sulfhydrylase from Salmonella typhimurium. J Mol Biol 283, 121–33 (1998).
Mozzarelli, A., Bettati, S., Pucci, A. M., Burkhard, P. & Cook, P. F. Catalytic competence of O-acetylserine sulfhydrylase in the crystal probed by polarized absorption microspectrophotometry. J Mol Biol 283, 135–46 (1998).
Burkhard, P., Tai, C. H., Ristroph, C. M., Cook, P. F. & Jansonius, J. N. Ligand binding induces a large conformational change in O-acetylserine sulfhydrylase from Salmonella typhimurium. J Mol Biol 291, 941–953 (1999).
Burkhard, P., Tai, C. H., Jansonius, J. N. & Cook, P. F. Identification of an allosteric anion-binding site on O-acetylserine sulfhydrylase: Structure of the enzyme with chloride bound. J Mol Biol 303, 279–286 (2000).
Guan, R., Roderick, S. L., Huang, B. & Cook, P. F. Roles of histidines 154 and 189 and aspartate 139 in the active site of serine acetyltransferase from Haemophilus influenzae. Biochemistry 47, 6322–8 (2008).
Kredich, N. M., Becker, M. A. & Tomkins, G. M. Purification and characterization of cysteine synthetase, a bifunctional protein complex, from Salmonella typhimurium. J Biol Chem 244, 2428–39 (1969).
Droux, M., Ruffet, M. L., Douce, R. & Job, D. Interactions between serine acetyltransferase and O-acetylserine (thiol) lyase in higher plants–structural and kinetic properties of the free and bound enzymes. Eur J Biochem 255, 235–45 (1998).
Huang, B., Vetting, M. W. & Roderick, S. L. The active site of O-acetylserine sulfhydrylase is the anchor point for bienzyme complex formation with serine acetyltransferase. J Bacteriol 187, 3201–5 (2005).
Mino, K. et al. Effects of bienzyme complex formation of cysteine synthetase from Escherichia coli on some properties and kinetics. Biosci Biotechnol Biochem 64, 1628–40 (2000).
Campanini, B. et al. Interaction of serine acetyltransferase with O-acetylserine sulfhydrylase active site: evidence from fluorescence spectroscopy. Protein Sci 14, 2115–24 (2005).
Mino, K. et al. Characteristics of serine acetyltransferase from Escherichia coli deleting different lengths of amino acid residues from the C-terminus. Biosci Biotechnol Biochem 64, 1874–80 (2000).
Zhao, C. et al. On the interaction site of serine acetyltransferase in the cysteine synthase complex from Escherichia coli. Biochem Biophys Res Commun 341, 911–6 (2006).
Berkowitz, O., Wirtz, M., Wolf, A., Kuhlmann, J. & Hell, R. Use of biomolecular interaction analysis to elucidate the regulatory mechanism of the cysteine synthase complex from Arabidopsis thaliana. J Biol Chem 277, 30629–34 (2002).
Johnson, P. M. et al. Unraveling the essential role of CysK in CDI toxin activation. Proc Natl Acad Sci USA 113, 9792–7 (2016).
Campanini, B. et al. Moonlighting O-acetylserine sulfhydrylase: New functions for an old protein. Biochimica et Biophysica Acta 1854, 1184–93 (2015).
Ma, D. K., Vozdek, R., Bhatla, N. & Horvitz, H. R. CYSL-1 interacts with the O2-sensing hydroxylase EGL-9 to promote H2S-modulated hypoxia-induced behavioral plasticity in C. elegans. Neuron 73, 925–40 (2012).
Beck, C. M. et al. CdiA effectors from uropathogenic Escherichia coli use heterotrimeric osmoporins as receptors to recognize target bacteria. PLoS Pathog 12, e1005925 (2016).
Ruhe, Z. C., Wallace, A. B., Low, D. A. & Hayes, C. S. Receptor polymorphism restricts contact-dependent growth inhibition to members of the same species. MBio 4, e00480–13 (2013).
Poole, S. J. et al. Identification of functional toxin/immunity genes linked to contact-dependent growth inhibition (CDI) and rearrangement hotspot (Rhs) systems. PLoS Genet 7, e1002217 (2011).
Ruhe, Z. C. et al. CDI systems are stably maintained by a cell-contact mediated surveillance mechanism. PLoS Genet 12, e1006145 (2016).
Amori, L. et al. Design and synthesis of trans-2-substituted-cyclopropane-1-carboxylic acids as the first non-natural small molecule inhibitors of O-acetylserine sulfhydrylase. MedChemComm 3, 1111–1116 (2012).
Salsi, E. et al. Design of O-acetylserine sulfhydrylase inhibitors by mimicking nature. J Med Chem 53, 345–56 (2010).
Spyrakis, F. et al. Fine tuning of the active site modulates specificity in the interaction of O-acetylserine sulfhydrylase isozymes with serine acetyltransferase. Biochim Biophys Acta Proteins Proteom 1834, 169–81 (2013).
Spyrakis, F. et al. Isozyme-specific ligands for O-acetylserine sulfhydrylase, a novel antibiotic target. PLoS One 8, e77558 (2013).
Pieroni, M. et al. Rational design, synthesis, and preliminary structure-activity relationships of alpha-substituted-2-phenylcyclopropane carboxylic acids as inhibitors of Salmonella typhimurium O-acetylserine sulfhydrylase. J Med Chem 59, 2567–78 (2016).
Annunziato, G. et al. Cyclopropane-1,2-dicarboxylic acids as new tools for the biophysical investigation of O-acetylserine sulfhydrylases by fluorimetric methods and saturation transfer difference (STD) NMR. J Enzyme Inhib Med Chem 31, 78–87 (2016).
Benoni, R. et al. Modulation of Escherichia coli serine acetyltransferase catalytic activity in the cysteine synthase complex. FEBS Lett 591, 1212–1224 (2017).
Salsi, E. et al. A two-step process controls the formation of the bienzyme cysteine synthase complex. J Biol Chem 285, 12813–22 (2010).
Kaundal, S., Uttam, M. & Thakur, K. G. Dual role of a biosynthetic enzyme, CysK, in contact dependent growth inhibition in bacteria. PLoS One 11, e0159844 (2016).
Wang, T. & Leyh, T. S. Three-stage assembly of the cysteine synthase complex from Escherichia coli. J Biol Chem 287, 4360–7 (2012).
Bernasconi, C. F. Relaxation Kinetics. (Academic Press Inc., New York, 1976).
Beck, C. M., Diner, E. J., Kim, J. J., Low, D. A. & Hayes, C. S. The F pilus mediates a novel pathway of CDI toxin import. Mol Microbiol 93, 276–90 (2014).
Ruhe, Z. C., Nguyen, J. Y., Beck, C. M., Low, D. A. & Hayes, C. S. The proton-motive force is required for translocation of CDI toxins across the inner membrane of target bacteria. Mol Microbiol 94, 466–81 (2014).
Dong, H., Nilsson, L. & Kurland, C. G. Co-variation of tRNA abundance and codon usage in Escherichia coli at different growth rates. J Mol Biol 260, 649–63 (1996).
Copeland, R. A. Evaluation of Enzyme Inhibitors in Drug Discovery - A guide for medicinal chemists and pharmacologists. (John Wiley and Sons, Hoboken, 2013).
Copeland, R. A. The drug-target residence time model: a 10-year retrospective. Nat Rev Drug Discov 15, 87–95 (2016).
Webb, J. S. et al. Delivery of CdiA nuclease toxins into target cells during contact-dependent growth inhibition. PLoS One 8, e57609 (2013).
Becker, M. A., Kredich, N. M. & Tomkins, G. M. The purification and characterization of O-acetylserine sulfhydrylase-A from Salmonella typhimurium. J Biol Chem 244, 2418–27 (1969).
Hulanicka, M. D., Hallquist, S. G., Kredich, N. M. & Mojica, A. T. Regulation of O-acetylserine sulfhydrylase B by L-cysteine in Salmonella typhimurium. J Bacteriol 140, 141–6 (1979).
Hindson, V. J. Serine acetyltransferase of Escherichia coli: substrate specificity and feedback control by cysteine. Biochem J 375, 745–52 (2003).
Olsen, L. R., Huang, B., Vetting, M. W. & Roderick, S. L. Structure of serine acetyltransferase in complexes with CoA and its cysteine feedback inhibitor. Biochemistry 43, 6013–9 (2004).
Kroger, C. et al. An infection-relevant transcriptomic compendium for Salmonella enterica Serovar Typhimurium. Cell Host Microbe 14, 683–95 (2013).
Feldman-Salit, A., Wirtz, M., Hell, R. & Wade, R. C. A mechanistic model of the cysteine synthase complex. J Mol Biol 386, 37–59 (2009).
Beck, C. M. et al. CdiA from Enterobacter cloacae delivers a toxic ribosomal RNase into target bacteria. Structure 22, 707–18 (2014).
Johnson, P. M. et al. Functional diversity of cytotoxic tRNase/immunity protein complexes from Burkholderia pseudomallei. J Biol Chem 291, 19387–400 (2016).
Morse, R. P. et al. Diversification of beta-augmentation interactions between CDI toxin/immunity proteins. J Mol Biol 427, 3766–84 (2015).
Penfold, C. N. et al. Flexibility in the receptor-binding domain of the enzymatic colicin E9 is required for toxicity against Escherichia coli cells. J Bacteriol 186, 4520–7 (2004).
Vankemmelbeke, M., Housden, N. G., James, R., Kleanthous, C. & Penfold, C. N. Immunity protein release from a cell-bound nuclease colicin complex requires global conformational rearrangement. Microbiologyopen 2, 853–61 (2013).
Jones, A. M., Garza-Sanchez, F., So, J., Hayes, C. S. & Low, D. A. Activation of contact-dependent antibacterial tRNase toxins by translation elongation factors. Proc Natl Acad Sci USA 114, E1951–E1957 (2017).
Baba, T. et al. Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: the Keio collection. Mol Syst Biol 2, 2006 0008 (2006).
Cherepanov, P. P. & Wackernagel, W. Gene disruption in Escherichia coli: TcR and KmR cassettes with the option of Flp-catalyzed excision of the antibiotic-resistance determinant. Gene 158, 9–14 (1995).
Koskiniemi, S. et al. Selection of orphan Rhs toxin expression in evolved Salmonella enterica serovar Typhimurium. PLoS Genet 10, e1004255 (2014).
Garza-Sanchez, F., Janssen, B. D. & Hayes, C. S. Prolyl-tRNA(Pro) in the A-site of SecM-arrested ribosomes inhibits the recruitment of transfer-messenger RNA. J Biol Chem 281, 34258–68 (2006).
Hayes, C. S. & Sauer, R. T. Cleavage of the A site mRNA codon during ribosome pausing provides a mechanism for translational quality control. Mol Cell 12, 903–11 (2003).
Garza-Sanchez, F., Gin, J. G. & Hayes, C. S. Amino acid starvation and colicin D treatment induce A-site mRNA cleavage in Escherichia coli. J Mol Biol 378, 505–19 (2008).
Willett, J. L., Gucinski, G. C., Fatherree, J. P., Low, D. A. & Hayes, C. S. Contact-dependent growth inhibition toxins exploit multiple independent cell-entry pathways. Proc Natl Acad Sci USA 112, 11341–6 (2015).
Tian, H. et al. Identification of the structural determinants for the stability of substrate and aminoacrylate external Schiff bases in O-acetylserine sulfhydrylase-A. Biochemistry 49, 6093–103 (2010).
Peterson, E. A. & Sober, H. A. Preparation of crystalline phosphorylated derivatives of vitamin B6. J Am Chem Soc 76, 169–75 (1954).
Baecker, P. A. & Wedding, R. T. Purification of serine acetyltransferase, a component of a multienzyme complex, by immunoadsorption and selective dissociation of the complex. Anal Biochem 102, 16–21 (1980).
Hindson, V. J., Moody, P. C., Rowe, A. J. & Shaw, W. V. Serine acetyltransferase from Escherichia coli is a dimer of trimers. J Biol Chem 275, 461–6 (2000).
Hindson, V. J. & Shaw, W. V. Random-order ternary complex reaction mechanism of serine acetyltransferase from Escherichia coli. Biochemistry 42, 3113–9 (2003).
Eyer, P. et al. Molar absorption coefficients for the reduced Ellman reagent: reassessment. Anal Biochem 312, 224–7 (2003).
Hama, H., Kayahara, T., Ogawa, W., Tsuda, M. & Tsuchiya, T. Enhancement of serine-sensitivity by a gene encoding rhodanese-like protein in Escherichia coli. J Biochem 115, 1135–40 (1994).
Gaitonde, M. K. A spectrophotometric method for the direct determination of cysteine in the presence of other naturally occurring amino acids. Biochem J 104, 627–33 (1967).
Bevans, C. G. et al. Determination of the warfarin inhibition constant Ki for vitamin K 2,3-epoxide reductase complex subunit-1 (VKORC1) using an in vitro DTT-driven assay. Biochim Biophys Acta Gen Subjects 1830, 4202–10 (2013).
Cheng, Y. & Prusoff, W. H. Relationship between the inhibition constant (K1) and the concentration of inhibitor which causes 50 per cent inhibition (I50) of an enzymatic reaction. Biochem Pharmacol 22, 3099–108 (1973).
Janssen, B. D. & Hayes, C. S. Kinetics of paused ribosome recycling in Escherichia coli. J Mol Biol 394, 251–67 (2009).
Acknowledgements
Circular dichroism experiments were carried out at Centro Interdipartimentale Misure of the University of Parma. We thank Gianluca Paredi and Omar De Bei for assistance with protein electrophoresis and Elie J. Diner for generating plasmid constructs. This work was supported in part by grant GM117373 (to C.S.H.) from the National Institutes of Health and by grants from the University of Parma (prot. FIL2014 to SB, BC and AM).
Author information
Authors and Affiliations
Contributions
R.B.: investigation; C.M.B.: investigation; F.G.S.: investigation; S.B.: original idea and planning, funding acquisition, supervision and manuscript reviewing and editing; A.M.: original idea and planning, funding acquisition and manuscript reviewing and editing; C.S.H.: original idea and planning, funding acquisition, supervision, original draft preparation and manuscript reviewing and editing; B.C.: original idea and planning, funding acquisition, supervision, formal analysis, original draft preparation and manuscript reviewing and editing.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Benoni, R., Beck, C.M., Garza-Sánchez, F. et al. Activation of an anti-bacterial toxin by the biosynthetic enzyme CysK: mechanism of binding, interaction specificity and competition with cysteine synthase. Sci Rep 7, 8817 (2017). https://doi.org/10.1038/s41598-017-09022-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-09022-6
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
