
Abstract
GPR101 is an orphan G protein-coupled receptor actively participating in energy homeostasis. Here we report the cryo-electron microscopy structure of GPR101 constitutively coupled to Gs heterotrimer, which reveals unique features of GPR101, including the interaction of extracellular loop 2 within the 7TM bundle, a hydrophobic chain packing-mediated activation mechanism and the structural basis of disease-related mutants. Importantly, a side pocket is identified in GPR101 that facilitates in silico screening to identify four small-molecule agonists, including AA-14. The structure of AA-14–GPR101–Gs provides direct evidence of the AA-14 binding at the side pocket. Functionally, AA-14 partially restores the functions of GH/IGF-1 axis and exhibits several rejuvenating effects in wild-type mice, which are abrogated in Gpr101-deficient mice. In summary, we provide a structural basis for the constitutive activity of GPR101. The structure-facilitated identification of GPR101 agonists and functional analysis suggest that targeting this orphan receptor has rejuvenating potential.

This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others
Data availability
All data generated in this study are included in the main text or Supplementary Information. The cryo-EM density map and the atomic coordinate have been deposited in Electron Microscopy Data Bank (EMDB) and Protein Data Bank (PDB) databases under accession codes EMD-37356 and 8W8Q for apo-GPR101–Gs complex; EMD-37357 and 8W8R for AA-14–GPR101–Gs complex; EMD-37358 and 8W8S for locally refined AA-14–GPR101 complex, respectively. RNA-sequencing data have been deposited into Gene Expression Ominibus under accession number GSE241719. All other data are available from the corresponding authors. Source data are provided with this paper.
References
Meye, F. J., Ramakers, G. M. & Adan, R. A. The vital role of constitutive GPCR activity in the mesolimbic dopamine system. Transl. Psychiatry 4, e361 (2014).
Sando, R. & Sudhof, T. C. Latrophilin GPCR signaling mediates synapse formation. eLife https://doi.org/10.7554/eLife.65717 (2021).
Xiao, X. et al. A new understanding of GHSR1a—independent of ghrelin activation. Ageing Res Rev. 64, 101187 (2020).
Tao, Y. X. Constitutive activation of G protein-coupled receptors and diseases: insights into mechanisms of activation and therapeutics. Pharmacol. Ther. 120, 129–148 (2008).
Bond, R. A. & Ijzerman, A. P. Recent developments in constitutive receptor activity and inverse agonism, and their potential for GPCR drug discovery. Trends Pharmacol. Sci. 27, 92–96 (2006).
Zhang, D. L. et al. Gq activity- and β-arrestin-1 scaffolding-mediated ADGRG2/CFTR coupling are required for male fertility. eLife https://doi.org/10.7554/eLife.33432 (2018).
Yao, Y. et al. A striatal-enriched intronic GPCR modulates huntingtin levels and toxicity. eLife https://doi.org/10.7554/eLife.05449 (2015).
Audo, I. et al. Whole-exome sequencing identifies mutations in GPR179 leading to autosomal-recessive complete congenital stationary night blindness. Am. J. Hum. Genet 90, 321–330 (2012).
Watkins, L. R. & Orlandi, C. In vitro profiling of orphan G protein coupled receptor (GPCR) constitutive activity. Br. J. Pharmacol. https://doi.org/10.1111/bph.15468 (2021).
Wacker, D., Stevens, R. C. & Roth, B. L. How ligands illuminate GPCR molecular pharmacology. Cell 170, 414–427 (2017).
Abboud, D. et al. GPR101 drives growth hormone hypersecretion and gigantism in mice via constitutive activation of Gs and Gq/11. Nat. Commun. 11, 4752 (2020).
Bates, B. et al. Characterization of Gpr101 expression and G-protein coupling selectivity. Brain Res. 1087, 1–14 (2006).
Nilaweera, K. N. et al. G protein-coupled receptor 101 mRNA expression in the mouse brain: altered expression in the posterior hypothalamus and amygdala by energetic challenges. J. Neuroendocrinol. 19, 34–45 (2007).
Trivellin, G., Faucz, F. R., Daly, A. F., Beckers, A. & Stratakis, C. A. Hereditary endocrine tumours: current state-of-the-art and research opportunities: GPR101, an orphan GPCR with roles in growth and pituitary tumorigenesis. Endocr. Relat. Cancer 27, T87–T97 (2020).
Hou, Z. S. & Tao, Y. X. Mutations in GPR101 as a potential cause of X-linked acrogigantism and acromegaly. Prog. Mol. Biol. Transl. Sci. 161, 47–67 (2019).
Trivellin, G. et al. Gigantism and acromegaly due to Xq26 microduplications and GPR101 mutation. N. Engl. J. Med. 371, 2363–2374 (2014).
Melmed, S. Pathogenesis and diagnosis of growth hormone deficiency in adults. N. Engl. J. Med. 380, 2551–2562 (2019).
Trivellin, G. et al. The X-linked acrogigantism-associated gene gpr101 is a regulator of early embryonic development and growth in zebrafish. Mol. Cell. Endocrinol. 520, 111091 (2021).
Yao, X. et al. Coupling ligand structure to specific conformational switches in the β2-adrenoceptor. Nat. Chem. Biol. 2, 417–422 (2006).
Yang, D. et al. Allosteric modulation of the catalytic VYD loop in Slingshot by its N-terminal domain underlies both Slingshot auto-inhibition and activation. J. Biol. Chem. 293, 16226–16241 (2018).
Suomivuori, C. M. et al. Molecular mechanism of biased signaling in a prototypical G protein-coupled receptor. Science 367, 881–887 (2020).
Wingler, L. M. et al. Angiotensin and biased analogs induce structurally distinct active conformations within a GPCR. Science 367, 888 (2020).
Veldhuis, J. D. Changes in pituitary function with ageing and implications for patient care. Nat. Rev. Endocrinol. 9, 205–215 (2013).
van den Beld, A. W. et al. The physiology of endocrine systems with ageing. Lancet Diabetes Endocrinol. 6, 647–658 (2018).
Savino, W., Mendes-da-Cruz, D. A., Lepletier, A. & Dardenne, M. Hormonal control of T-cell development in health and disease. Nat. Rev. Endocrinol. 12, 77–89 (2016).
Savino, W. & Dardenne, M. Pleiotropic modulation of thymic functions by growth hormone: from physiology to therapy. Curr. Opin. Pharmacol. 10, 434–442 (2010).
Yang, F. et al. Structural basis of GPBAR activation and bile acid recognition. Nature 587, 499–504 (2020).
Lu, J. et al. Structural basis for the cooperative allosteric activation of the free fatty acid receptor GPR40. Nat. Struct. Mol. Biol. 24, 570–577 (2017).
Giordano, R., Bonelli, L., Marinazzo, E., Ghigo, E. & Arvat, E. Growth hormone treatment in human ageing: benefits and risks. Hormones 7, 133–139 (2008).
Qu, C. et al. Ligand recognition, unconventional activation, and G protein coupling of the prostaglandin E2 receptor EP2 subtype. Sci. Adv. https://doi.org/10.1126/sciadv.abf1268 (2021).
Zheng, S. Q. et al. MotionCor2: anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nat. Methods 14, 331–332 (2017).
Mindell, J. A. & Grigorieff, N. Accurate determination of local defocus and specimen tilt in electron microscopy. J. Struct. Biol. 142, 334–347 (2003).
Zivanov, J. et al. New tools for automated high-resolution cryo-EM structure determination in RELION-3. eLife https://doi.org/10.7554/eLife.42166 (2018).
Kucukelbir, A., Sigworth, F. J. & Tagare, H. D. Quantifying the local resolution of cryo-EM density maps. Nat. Methods 11, 63–65 (2014).
Scheres, S. H. & Chen, S. Prevention of overfitting in cryo-EM structure determination. Nat. Methods 9, 853–854 (2012).
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. D 66, 486–501 (2010).
Adams, P. D. et al. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D 66, 213–221 (2010).
Yang, F. et al. Structure, function and pharmacology of human itch receptor complexes. Nature 600, 164–169 (2021).
Cheng, J. et al. Autonomous sensing of the insulin peptide by an olfactory G protein-coupled receptor modulates glucose metabolism. Cell Metab. 34, 240–255 e210 (2022).
Li, A. et al. Discovery of novel FFA4 (GPR120) receptor agonists with β-arrestin2-biased characteristics. Future Med Chem. 7, 2429–2437 (2015).
Du, Y. Q. et al. Endogenous lipid-GPR120 signaling modulates pancreatic islet homeostasis to different extents. Diabetes 71, 1454–1471 (2022).
Huang, J. et al. CHARMM36m: an improved force field for folded and intrinsically disordered proteins. Nat. Methods 14, 71–73 (2017).
Lee, J. et al. CHARMM-GUI input generator for NAMD, GROMACS, AMBER, OpenMM, and CHARMM/OpenMM simulations using the CHARMM36 additive force field. J. Chem. Theory Comput. 12, 405–413 (2016).
Vanommeslaeghe, K. et al. CHARMM general force field: a force field for drug-like molecules compatible with the CHARMM all-atom additive biological force fields. J. Comput. Chem. 31, 671–690 (2010).
Metropolis, N. & Ulam, S. The Monte Carlo method. J. Am. Stat. Assoc. 44, 335–341 (1949).
Humphrey, W., Dalke, A. & Schulten, K. VMD: visual molecular dynamics. J. Mol. Graph 14, 33–38 (1996). 27-38.
Kumari, R., Kumar, R., Open Source Drug Discovery, C. & Lynn, A. g_mmpbsa—a GROMACS tool for high-throughput MM-PBSA calculations. J. Chem. Inf. Model. 54, 1951–1962 (2014).
Lin, H. et al. Structures of the ADGRG2–Gs complex in apo and ligand-bound forms. Nat. Chem. Biol. 18, 1196–1203 (2022).
Ping, Y. Q. et al. Structures of the glucocorticoid-bound adhesion receptor GPR97–Go complex. Nature 589, 620–626 (2021).
Acknowledgements
We thank the Translational Medicine Core Facility of Advanced Medical Research Institute at Shandong University for their technical assistance with the Multimode Plate Reader. We thank S.-Y. Yu and Y. Li for their help with stereotactic injection. The scientific calculations in this paper have been done on the HPC Cloud Platform of Shandong University. We acknowledge support from the National Key R&D Program of China (2019YFA0904200 to J.-P.S., 2019YFA0508904 to N.G. and 2022YFC2702600 to F. Yang). the National Science Fund for Distinguished Young Scholars Grant (81825022 to J.-P.S. and 82225011 to X.Y.), National Science Fund for Excellent Young Scholars (82122070 to F. Yang), the National Natural Science Foundation of China Grant (32130055 to J.-P.S., 92057121 to X.Y., 31725007 to N.G., 31701230 to Z.Y. and 32201065 to Y. Lu), the Major Fundamental Research Program of the Natural Science Foundation of Shandong Province, China (ZR2020ZD39 to J.-P.S. and ZR2021ZD18 to X.Y.), the Key Research Project of the Natural Science Foundation of Beijing, China (Z20J00129 to J.-P.S.), the China Postdoctoral Science Foundation (2022M711916 to Y.Lu and 2022M711911 to S.-L.N.), Natural Science Foundation of Shandong (ZR2023MH196 to S.-L.N.), the Instrument Improvement Funds of Shandong University Public Technology Platform (ts20220103 to F. Yang), the Cutting Edge Development Fund of Advanced Medical Research Institute (to Z.Y., F. Yang, K.-K.Z., P.X., J.-P.S.).
Author information
Authors and Affiliations
Contributions
J.-P.S., N.G. and X.Y. conceived and supervised the overall project. J.-Y.W. generated the GPR101 insect expression constructs. X.Y. and Z.Y. initiated the functional study of GPR101 and its mutants. F. Yang, J.-Y.W. and Y.G. purified the protein complex and prepared the samples for cryo-EM. G.-P.W. and J.-Y.W. collected cryo-EM data and performed cryo-EM map calculation, model building and the structure refinement under the supervision of N.G. and J.-P.S. J.-P.S., N.G., Z.Y., F. Yang, P.X. and J.-Y.W. analyzed the complex structures and designed structure–function experiments. J.-P.S. designed all the mutations for AA-14 ligand binding pocket and GPR101–Gs interface. Z.Y. and J.-Y.W. performed all the cAMP assays, β-arrestin recruitment assay, GRK recruitment assay, G protein dissociation assay and cell surface ELISA assay. Y.G. performed the FlAsH BRET assay. Z.Y., J.-Y.W., F. Yi and P.X. analyzed the data for cellular assays. J.-P.S. and Z.Y. designed the Bimane fluorescence quenching assay. J.-Y.W., Z.Y. and F. Yang performed the Bimane fluorescence quenching assay. J.-P.S. and F.X. designed the GPR52-related experiments. K.-K.Z. and H.Z. performed a hierarchical virtual screen. C. Zhang performed the side pocket analysis and GPR101 modeling. Z.Y., J.-Y.W. and M.-W.W. designed the AAV strategy and performed transcriptome analysis. Y. Lu, Y. Li and Y.G. performed MS-based ligand binding experiment and relevant analysis. Z.Y., J.-Y.W., M.-W.W., C. Zhang and P.X. performed animal experiments. C. Zheng., P.X. and B.-H.L. participated in data analysis of animal experiments. Y.Z. analyzed the pituitary single-cell RNA sequencing data. S.-H.Z. and X.-W.W. measured the hormone levels and inositol monophosphate levels. S.-L.N. performed the immunofluorescence and western blotting under the supervision of P.-J.Z. J.-Y.W. prepared Nb35 for the formation of the complex. Z.Y., J.-Y.W., F. Yang, Y.G. and G.-P.W. prepared the figures. J.-P.S., N.G. and X.Y. wrote the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Chemical Biology thanks John McCorvy, Constantine Stratakis and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
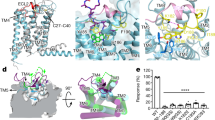
Extended Data Fig. 1 Constitutive G protein and β-arrestin activities of GPR101 SNPs.
a, Association of GPR101 SNPs with human endocrine diseases. The c.924 G > C (p. E308D), c.365 T > Y (p. I122T) and c.589 G > T (p. V197L) were identified in patients with acromegaly, Cushing’s disease and congenital GH deficiency, respectively. ND, not determined. b, Summary of the Gs and β-arrestin2 activities of GPR101 mutants. Gs or β-arrestin2 activity was defined by the maximal cAMP level in GPR101 (or mutant)-transfected HEK293 cells or the BRET signal between Rluc-β-arrestin2 and GPR101 (or mutant)-YFP, respectively. Data were normalized to the saturating cAMP level or BRET signal of WT GPR101 (n = 3). c, Effects of GPR101 SNPs on GH secretion in GH3 cells. Cells transfected with pcDNA3.1 were used as the negative control. E308D mutant induced higher GH secretion than WT GPR101, whereas I122T or V197L showed the opposite effect. Data were from three independent experiments (n = 3). d, Effects of GPR101 mutations on ACTH secretion in AtT-20 cells. AtT-20 cells transfected with pcDNA3.1 were used as the negative control. I122T significantly increased ACTH secretion compared with WT GPR101, whereas V197L led to a subtle decrease in ACTH secretion. Data were from three independent experiments (n = 3). For c-d, data are presented as mean ± SEM. P values were calculated using one-way ANOVA with Dunnett’s post hoc test and are indicated in the graphs.
Extended Data Fig. 2 Cryo-EM images of apo-GPR101-Gs complex and data processing.
a-b, Representative Cryo-EM micrograph (Scale bar: 50 nm) (a) and 2D class averages (b) of the GPR101-Gs complex. Representative Cryo-EM micrographs from 6,161 movies and representative 2D class averages determined using approximately 1.82 million particles from 2D classification were shown. c, Flow chart of cryo-EM data processing of GPR101-Gs complex leading to the construction of electron density maps with an overall resolution of 2.89 Å. d, 3D density map of the GPR101-Gs complex colored according to local resolution. e, Angular distribution of the particles used for the final reconstruction of the GPR101-Gs complex. f, Gold-standard Fourier shell correlation (FSC) curves for GPR101-Gs complex.
Extended Data Fig. 3 Binding of ECL2 into the orthosteric pocket of GPR101.
a, EM densities of the residues of GPR101 ECL2 (orange) and its interacting residues (slate) in the orthosteric binding pocket. Three hydrophobic residues of the downstream β2-strand, L181ECL2, M184ECL2 and W186ECL2, sit in a hydrophobic groove composed of V1083.29, H1123.33, P1664.60, Y1925.38, F2005.46, Y4156.51 and I4407.39. b, The distances between the ECL2 (measured at W186 in GPR101 and the equivalent H186 in GPR52) and the traditional toggle switch residue (L6.48 in GPR101 and W6.48 GPR52). Compared with GPR52, ECL2 of GPR101 binds shallower within the orthosteric site (9.2 Å vs. 4.9 Å) and does not directly reach the toggle switch residue.
Extended Data Fig. 4 GPR101 activation induced by hydrophobic chain packing.
a, Structural comparison of TM3 and TM6 of GPR101 (slate) with the BI-167107-bound active (PDB ID: 3SN6, cyan) and ICI-118551-bound inactive (PDB ID: 3NY8, gray) structures of β2AR. The distance between the ends of TM3 and TM6 of GPR101 and activated β2AR is 13.3 Å and 13.5 Å, respectively, when measured between the Cα carbons of I1333.54 and A3976.33. b, Schematic representation of the hydrophobic packing of GPR101 transmitting the activation signal induced by ECL2 binding. Y4156.51, which contacts W186ECL2 and H1123.33 via hydrophobic interactions and hydrogen bonds, respectively, packs with F2005.46 and F4437.42, which then transduce the packing to L4126.48 and contribute to the stabilization of the P5.50-V3.40-Y6.44 motif. c, Effects of mutations of key residues of the hydrophobic chain packing, including W186 ECL2, Y4156.51 and L4126.48, on the constitutive Gs activity of GPR101. Data were normalized to the basal cAMP level in HEK293 cells transfected with WT GPR101. Data are from three independent experiments (n = 3). WT GPR101 and the corresponding mutants were expressed at similar levels. Data are presented as mean ± SEM values. P values were calculated using one-way ANOVA with Dunnett’s post hoc test and are indicated in the graphs.
Extended Data Fig. 5 Coupling of GPR101 to Gs.
a, The binding interface between GPR101 and Gs with the α5 helix highlighted in yellow. b-c, Superposition of the structures of the GPR101-Gs, β2AR-Gs (PDB ID: 3SN6) and GPR52-Gs (PDB ID: 6LI3) complexes revealing a similar direction of the α5 helix (b) and less separation between TM3 and TM7 of GPR101 (c). GPR101 is shown in slate, β2AR in yellow, and GPR52 in green. d-f, Detailed interactions of residues at TM7-helix 8 (d), ICL1 (e), and ICL2 (f) of GPR101 (slate) with Gα (orange) and Gβ (green). The polar bonds are presented as red dashed lines.
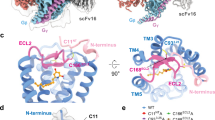
Extended Data Fig. 6 The side binding pocket of GPR101.
a, The side pocket volumes of GPR101 (slate) and GPR52 (green). The PyVOL, which is a python library packaged into a PyMOL GUI, was used to calculate the side pocket volumes with the probe radius set at 1.4 Å. b-c, Sequence alignment of c17-binding side pocket residues at the N-terminus (b) and TM bundles or ECL2 (c) in GPR52 with the equivalent residues in GPR101. The conserved residues are shown in blue, while the distinct residues are shown in red. d, Effects of substitution of key residues in the side pocket of GPR101 with the equivalent residues of GPR52 on the responsiveness of GPR101 to c17 stimulation. Data were normalized to the basal cAMP level in HEK293 cells transfected with WT GPR101 and are from three independent experiments (n = 3). The WT GPR101 and corresponding mutants were expressed at similar levels. e, Cell surface ELISA assay showing similar expression levels of WT GPR101, WT GPR52 and GPR101 mutants with key residues in side pocket substituted with the equivalent residues of GPR52. Data were normalized to the expression level of WT GPR101 in HEK293 cells. Data were from three independent experiments (n = 3). For d-e, all data are presented as mean ± SEM values. P values were calculated using two-way ANOVA (d) or one-way ANOVA (e) with Dunnett’s post hoc test and are indicated in the graphs.
Extended Data Fig. 7 Screening and signal profiling of GPR101 agonists.
a, An overview of the algorithm for in silico GPR101 ligands screening and validation. b, The docking scores of the 137 candidate compounds revealed by DOCK4.025. c, Summarized EC50 and Emax values of different ligands in activating Gs, Gq or β-arrestin2 pathways downstream of GPR101. AA-1 was used as the negative control. Values represent the mean ± SEM from three independent experiments (n = 3) performed in triplicate. ND, not detectable. d, Summarized biased factors and bias properties of the GPR101 agonists. β values (bias factor) of the selective ligands towards individual pathway were calculated using AA-14-induced Gq activation as the reference. Data are from three independent experiments (n = 3) performed in triplicates.
Extended Data Fig. 8 Interaction mode of AA-14 with GPR101.
a, The binding energy contribution of side pocket residues in the AA-14-GPR101-Gs complex calculated by the molecular mechanics Poisson–Boltzmann surface area (MM-PBSA) method. Data are from three independent MD simulations (n = 3). b-c, Cryo-EM density maps of AA-14 and its interacting residues at the side binding pocket (b) and the second binding pocket (c) of GPR101. The density maps were shown with the same counter level set in Chimera software (0.0110 V). d, Schematic representation of the FlAsH-BRET assay design. NanoLuc (Nluc) was fused to N-terminus of GPR101, and the FlAsH motif (CCPGCC) was incorporated at the designated positions of the extracellular loops (ECLs) of the receptor. The WT GPR101 and corresponding mutants were expressed at similar levels. e, The maximal response of GPR101 FlAsH-BRET sensors in response to AA-14 stimulation. Values are from three independent experiments (n = 3). ND, not detectable. f, Effects of mutations in the second binding pocket on AA-14-induced ECL1 conformational changes of GPR101 measured by FlAsH-BRET. Values are from three independent experiments (n = 3). The WT GPR101 and corresponding mutants were expressed at similar levels. For a, e and f, data are presented as mean ± SEM values. P values were calculated using one-way ANOVA with Dunnett’s post hoc test and are indicated in the graphs.
Extended Data Fig. 9 AA-14 promotes hormone secretion and regulates metabolism through activating GPR101.
a, Growth hormone secretion from GH3 cells transfected with siGpr101 or control siRNA in response to stimulation with 100 μM AA-14. Data are from three independent experiments (n = 3). b-c, Plasma growth hormone (b) and IGF-1 (c) levels in Gpr101f l/f l or Pomc-cre+/−Gpr101f l/f l mice treated with AA-14 (0.1 mg/kg/d) or control vehicle (n = 6 mice per group). d, Energy expenditure of the 8-week-old or 18-month-old mice treated with AA-14 (0.1 mg/kg/d) or control vehicle for 2 months (n = 6 mice per group). e, mRNA levels of Glb1, p21, Il6 and Il1α in the skin of mice treated with AA-14 or control vehicle for 2 months (n = 3 mice per group). For a-e, all data are presented as mean ± SEM values. P values were calculated using unpaired two-sided Student’s t-test (a) or two-way ANOVA with Dunnett’s post hoc test (b-e), and are indicated in the graphs.
Supplementary information
Supplementary Information
Supplementary Tables 1–8, Figs. 1–22 and uncropped scans of all blots and gels.
Supplementary Data
Statistical source data of Supplementary Information.
Source data
Source Data Fig. 3
Statistical source data.
Source Data Fig. 4
Statistical source data.
Source Data Fig. 5
Statistical source data.
Source Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 1
Statistical source data.
Source Data Extended Data Fig. 4
Statistical source data.
Source Data Extended Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 7
Statistical source data.
Source Data Extended Data Fig. 8
Statistical source data.
Source Data Extended Data Fig. 9
Statistical source data.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Yang, Z., Wang, JY., Yang, F. et al. Structure of GPR101–Gs enables identification of ligands with rejuvenating potential. Nat Chem Biol 20, 484–492 (2024). https://doi.org/10.1038/s41589-023-01456-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41589-023-01456-6
This article is cited by
-
G protein-coupled receptors (GPCRs): advances in structures, mechanisms, and drug discovery
Signal Transduction and Targeted Therapy (2024)
