
Abstract
Cytoplasmic incompatibility (CI) is a common form of reproductive sabotage caused by maternally inherited bacterial symbionts of arthropods. CI is a two-step manipulation: first, the symbiont modifies sperm in male hosts which results in the death of fertilized, uninfected embryos. Second, when females are infected with a compatible strain, the symbiont reverses sperm modification in the fertilized egg, allowing offspring of infected females to survive and spread the symbiont to high frequencies in a population. Although CI plays a role in arthropod evolution, the mechanism of CI is unknown for many symbionts. Cardinium hertigii is a common CI-inducing symbiont of arthropods, including parasitoid wasps like Encarsia partenopea. This wasp harbors two Cardinium strains, cEina2 and cEina3, and exhibits strong CI. The strains infect wasps at different densities, with the cEina3 present at a lower density than cEina2, and it was previously not known which strain caused CI. By differentially curing wasps of cEina3, we found that this low-density symbiont is responsible for CI and modifies males during their pupal stage. cEina2 does not modify host reproduction and may spread by ‘hitchhiking’ with cEina3 CI or by conferring an unknown benefit. The cEina3 strain also shows a unique localization pattern in male reproductive tissues. Instead of infecting sperm like other CI-inducing symbionts, cEina3 cells are found in somatic cells at the testis base and around the seminal vesicle. This may allow the low-density cEina3 to efficiently modify host males and suggests that cEina3 uses a different modification strategy than sperm-infecting CI symbionts.
Similar content being viewed by others


Introduction
Most terrestrial arthropods harbor intracellular bacterial symbionts that are transmitted vertically from infected females to their offspring (Weinert et al. 2015; Zchori‐Fein and Perlman 2004; Zug and Hammerstein 2012). Due to their reliance on maternal transmission, many symbionts increase the likelihood of their transmission by providing benefits to their host, or by manipulating host reproduction to favor production of infected female progeny (Doremus and Hunter 2020; Oliver and Martinez 2014). One common manipulation is a symbiont-induced crossing incompatibility called cytoplasmic incompatibility (CI). Symbiont-induced CI is a two-step reproductive sabotage that is best described by a “modification” and “rescue” model: first, the symbiont modifies male hosts in a manner that will result in embryonic mortality of his offspring when he mates with uninfected females. The next step occurs within infected egg cytoplasm following fertilization by infected male sperm, where the symbiont reverses this modification and “rescues” the embryonic offspring. The modification and rescue steps of CI work in tandem to facilitate the spread of the CI symbiont through the host population by increasing infected female relative fitness at the expense of males and uninfected females (O’Neill et al. 1997; Shropshire et al. 2020; Turelli 1994).
Symbiont-induced CI has received considerable attention for its potential role in arthropod evolution and speciation (Shoemaker et al. 1999; Bordenstein et al. 2001; Gebiola et al. 2016a) as well as its application in driving desirable traits, like suppression of RNA viruses or mating incompatibility, through arthropod pest populations (Moreira et al. 2009; Ryan et al. 2019; Zheng et al. 2019; Li et al. 2020). One widespread symbiont lineage, Wolbachia, has received the bulk of attention, which has culminated recently in the identification of the Wolbachia effectors responsible for CI, cytoplasmic incompatibility factor A (cifA) and cifB (Beckmann et al. 2017; Lepage et al. 2017; Shropshire et al. 2018). Wolbachia has also been directly applied to control the spread of arthropod-vectored diseases like the Dengue virus via its associated mating incompatibility and its ability to suppress RNA viruses harbored by the host mosquito, as well as its gene drive capabilities via CI (Ryan et al. 2019; Zheng et al. 2019). Yet beyond the Wolbachia system, the CI caused by other common symbionts remains largely uncharacterized (Hunter et al. 2003; Takano et al. 2017; Konig et al. 2019; Rosenwald et al. 2020; Takano et al. 2021).
Cardinium hertigii is a common heritable symbiont that infects ~7–10% of arthropod species (Weinert et al. 2015; Zchori‐Fein and Perlman 2004) and causes CI in a range of hosts including mites (Gotoh et al. 2007; Wu and Hoy 2012), planthoppers (Nakamura et al. 2012), thrips (Nguyen et al. 2017), and parasitic Encarsia wasps (Gebiola et al. 2016a; Hunter et al. 2003). Unfortunately, many of the species that host Cardinium are minute, non-model arthropods that make the study of Cardinium-induced CI difficult. Most of what is known about Cardinium CI stems from research on a single symbiosis between the Cardinium strain cEper1 and its minute wasp host, Encarsia suzannae (Hunter et al. 2003). Sequencing and analysis of the cEper1 genome revealed little homology in eukaryotic-associated genes to Wolbachia (Penz et al. 2012). Notably, Cardinium lacks the genetic factors (i.e., cifA and cifB) responsible for Wolbachia CI, indicating that Cardinium independently evolved CI (Beckmann et al. 2017; LePage et al. 2017; Lindsey et al. 2018; Mann et al. 2017; Penz et al. 2012; Shropshire et al. 2018). Despite the different origins of CI in Wolbachia and Cardinium, cytological analysis of CI-affected embryos revealed that both symbionts induce similar lethal cellular defects during cellular replication in early embryogenesis (Gebiola et al. 2017). Likewise, cEper1 shows a similar localization pattern in host testes to many Wolbachia strains, in which the symbiont infects developing sperm cells but is lost during spermiogenesis, the final stage of sperm maturation and elongation (Doremus et al. 2020). During spermiogenesis the sperm DNA also becomes hyper-condensed (Raja and Renkawitz-Pohl 2005; Balhorn 2007; Ferree et al. 2019) and the nucleus begins to elongate as the cytoplasmic content of sperm cells is removed, the sperm tail grows, and the sperm cells individualize (Fuller 1993; Ferree et al. 2019; Doremus et al. 2020).
Several lines of evidence suggest that cEper1 modifies host sperm cells to deliver the fatal CI modification into the egg cell and that cEper1 performs these CI modifications during the E. suzannae pupal stage. First, prolonging the duration of the pupal stage increases the severity of CI (Doremus et al. 2019, 2020). Second, E. suzannae produces most of its lifetime sperm in the pupal stage and Cardinium is not present in seminal vesicles, the male storage organ where mature sperm accumulates (Doremus et al. 2020). Third, reduction of Cardinium titer in adult wasps has no effect on CI strength (Doremus et al. 2020), suggesting that modification is complete prior to the adult stage. Whether this modification window and localization pattern is unique to cEper1 or is a common feature of Cardinium CI symbioses is not known, as other Cardinium CI symbioses remain largely uncharacterized.
Cardinium-induced CI has been described in another parasitic wasp, Encarsia partenopea (Gebiola et al. 2016b). The name E. partenopea was earlier considered a synonym of E. inaron (Polaszek et al. 1992). Later studies showed E. partenopea was genetically and biologically distinct from E. inaron (Gebiola et al. 2016b) so we will use the original name here. Like all Hymenoptera, E. partenopea is haplodiploid; males develop from unfertilized eggs and females develop from fertilized eggs. Encarsia are typically autoparasitic with male and female wasps developing in different hosts; however, in E. partenopea both males and females develop as primary parasitoids of whiteflies. Cardinium CI specifically kills fertilized female E. partenopea embyros (Gebiola et al. 2016b).
The lethality of CI in this second Encarsia species is greater than cEper1-induced CI in E. suzannae, with CI crosses causing nearly complete female offspring mortality (~90–100% mortality) (Gebiola et al. 2016b). Recently, attempts at sequencing the genome of the Cardinium symbiont in E. partenopea revealed a cryptic infection with a second Cardinium strain (Stouthamer 2018). It was not known if one strain is responsible for CI in this system or if both strains contributed to this manipulative phenotype. The coinfecting Cardinium strains are found at different infection densities in adult wasps, with one strain, cEina2, found at a much higher density than the cryptic second strain, cEina3 (Stouthamer 2018; Stouthamer et al. 2018). Phylogenetic analysis of both strains using a Multi-Locus Sequence Typing (MLST) system revealed that the higher density strain, cEina2, is most closely related to asymptomatic Cardinium strains infecting the sweet potato whitefly, Bemisia tabaci (cBtQ1) and US populations of E. inaron (cEina1) (Fang et al. 2014; Stouthamer et al. 2019; White et al. 2009).
Here we investigated Cardinium-induced CI in E. partenopea. By exposing wasps to low levels of antibiotics (White et al. 2009), we were able to establish wasp lines that were coinfected, singly infected with cEina2, and uninfected. We then used reciprocal crosses among all three lines to test whether cEina2, cEina3, or both strains were responsible for CI in E. partenopea. We next used qPCR to estimate infection dynamics of both strains across host development. To determine whether CI modification occurs prior to the adult stage in E. partenopea, we tested the relative strength of CI in infected adult male wasps that had severely reduced Cardinium titer from antibiotic feeding. Finally, we explored the localization of Cardinium in testes across host development using fluorescence in-situ hybridization (FISH) with a strain-specific probe. We found that the low-density cEina3 strain is responsible for CI in E. partenopea. While this strain, like cEper1 in E. suzannae, also modifies male hosts during pupal development, cEina3 appears to use a unique localization strategy to efficiently modify males despite its low abundance in male reproductive tissues. These results suggest that some Cardinium strains use different strategies to modify male hosts, and that insights into one Cardinium CI symbiosis may not apply universally to all Cardinium.
Methods
Insect and symbiont cultures
Encarsia partenopea (Hymenoptera: Aphelinidae) is a minute parasitoid of whiteflies, collected in Portici, Southern Italy in 2005 (Gebiola et al. 2016b) and established in culture at the University of Arizona in Tucson, AZ USA. Parasitoid wasps were cultured on sweet potato whitefly (Bemisia tabaci) infesting cowpea (Vigna unguiculata) plants at 27 °C.
Previously, an E. partenopea line lacking Cardinium was established via feeding female wasps rifampicin in honey (50 mg ml−1) (Gebiola et al. 2016b; Hunter et al. 2003). An additional wasp line singly infected with the cEina2 strain was created by feeding doubly infected female wasps a low dose of rifampicin (0.1 mg ml−1) for 48 h to destabilize symbiont transmission prior to egg laying (White et al. 2009). Following antibiotic treatment, the progeny of individual females was evaluated by diagnostic PCR, and when confirmed to contain just cEina2, multiple families were pooled for a cEina2-only line. Unfortunately, this treatment did not yield cEina3-only progeny.
Strain-specific symbiont infection status was confirmed using primers for the single copy ydhE multi-drug efflux gene (also used for quantitative PCR) or bioH, a gene in the biotin synthesis pathway, specific to either cEina2 or cEina3. Primer and PCR cycle information is included in Supplementary Table S1. All wasp lines were maintained for at least four generations prior to use in experiments and underwent periodic diagnostic screening for symbiont infection.
CI crossing assays
Using the doubly, singly, and uninfected wasp lines, we performed reciprocal CI (infected male X uninfected female) and rescue (infected male X infected female) crosses to reveal whether one or both symbiont strains (cEina2 or cEina3) were responsible for CI. Male and female wasps from each of the lines were isolated from parasitized whiteflies as pupae in 1.2 mL vials. After adult eclosion, wasps were provided with a small amount of honey for two days and males were allowed to mate with either an uninfected female, a cEina2-infected female, or a doubly infected female (n = 9–10 mating pairs per cross). Unlike many Encarsia species, E. partenopea do not readily mate unless in the presence of whiteflies. To allow ample time for mating to occur and provide female wasps time to lay eggs, males and females were placed together in parasitism arenas for 48 h. Arenas consisted of a 35 mm petri dish with a vented lid containing cowpea leaves infested with 30–50 2nd instar B. tabaci nymphs on 1% agar. After 48 h, male and female wasps were removed from the parasitism arena and stored at −80 °C for mating confirmation. Arenas were then monitored for 14 days post-parasitism, when offspring pupae were collected into cotton enclosed 1.2 mL vials. In haplodiploid organisms like Encarsia, in which only female eggs are fertilized, CI specifically targets female offspring. Therefore, offspring of CI crosses exhibit heavily male-biased sex ratios (Breeuwer 1997; Breeuwer and Werren 1990; Mouton et al. 2005; Vavre et al. 2000). To calculate the severity of CI, offspring sex was recorded as adults emerged. Compatible crosses yielded equal numbers of female and male offspring, indicating that mating occurred readily once in the parasitism arena. In this haplodiploid system, all-male broods can be caused either by CI or lack of mating. For crosses that produced only male offspring, female spermathecae (the female sperm storage organ) were dissected, cleared with a lactophenol solution (1 part carbolic acid, 1 part lactic acid, 2 parts glycerin and 1 part distilled water) and examined at 200–400 × magnification with a compound microscope for the presence of sperm for mating confirmation (White et al. 2009). The crosses with females determined to be unmated were removed from analysis (5 out of 90 crosses in total were removed). Male and female offspring production in potential CI and rescue crosses were analyzed using ANOVA, and sex ratios (expressed as the proportion of male offspring) were analyzed using logistic regression with a quasibinomial distribution in the R 4.0.4 statistical platform (R Core Team 2021).
Cardinium within-host densities across host development
To characterize the changes in symbiont infection density across male host development, we collected male E. partenopea at five different life stages: 3rd instar larvae, white pupae (~2 days post pupation), mottled black and white pupae (~4 days post pupation), black pupae (~6 days post pupation), and adults (2 days after emergence) (n = 5 per samples per stage). Wasps were stored at −80 °C until DNA extraction. To extract DNA, wasps were homogenized in 3 μL of 20 mg ml −1 proteinase k, followed by adding the homogenate to 50 μL of 5–10% w/v Chelex (White et al. 2009). Samples were incubated at 37 °C for 1 h with periodic vortexing, followed by incubation at 97 °C for 8 min and storage at −20 °C. As in our previous work (Stouthamer et al. 2018; Doremus et al. 2019, 2020), we estimated Cardinium density relative to host cells by performing quantitative PCR (qPCR) using Maxima SYBR Green/ROX qPCR Master Mix (2×) (ThermoFisher Scientific) with primers specific for the cEina2 or cEina3 versions of the single-copy Cardinium ydhE gene and the host Ef1a gene on a Bio-Rad CFX Connect Real-Time cycler (Stouthamer et al. 2018). We also created standards for each primer set (host-ef1α, cEina2-ydhE, cEina3-ydhE) via serial dilutions of PCR products. PCR products were initially diluted to a concentration of 1.0 ng/μL DNA, which was confirmed with a Qubit 4.0 fluorometer prior to serial dilution. Samples were run in triplicate and each qPCR plate included standards for all primer sets to correct for between-plate differences in reaction efficiency. Raw Cq values were averaged and corrected before conversion to relative density (Perlman et al. 2014). Relative Cardinium densities (cEina2 or cEina3) were compared across host life stage and between strains (cEina2 vs. cEina3) at each life stage using a linear model with developmental stage as an ordered factor, and strain and interaction as additional factors. We performed Tukey’s HSD post-hoc test with Benjamini–Hochberg corrected p values for multiple comparisons in R v4.0.4 (R Core Team 2021).
Antibiotic CI trials
To explore whether the timing of CI modification in E. partenopea occurs during development (larval and/or pupal stages) or the adult stage, we used a modified version of the antibiotic trials used to confirm the CI modification window in another Cardinium-infected wasp, E. suzannae (Doremus et al. 2020). Doubly-infected male E. partenopea were fed either rifampicin in honey (50 mg mL−1) or pure honey for seven days after adult emergence. This level of rifampicin consistently causes a large loss of Cardinium in E. suzannae (Doremus et al. 2020). Males were then allowed to mate either with an uninfected female (the CI cross) or doubly infected female (the rescue cross) as described above (n = 10–15 mating pairs per cross type and treatment). Here, the rescue cross serves as a control for potential direct effects of rifampicin on male reproductive viability, even though previous rifampicin experiments on E. suzannae did not show any adverse effects of an antibiotic diet (Doremus et al. 2020). A subset of five male wasps per treatment were saved at −80 °C for DNA extraction and confirmation of Cardinium loss in rifampicin-treated males (Doremus et al. 2020). The details of the crossing experiments and analysis of offspring production and sex ratio proceeded as described above. Mating status was confirmed by visualizing the presence of sperm in the spermatheca of females that produced all male broods. In total, 4 of the 52 crosses were removed from the analysis because of mating failure.
Cardinium localization using fluorescence in-situ Hybridization
We characterized Cardinium localization in E. partenopea testes relative to developing sperm cells using fluorescence in-situ hybridization (FISH) on doubly infected E. partenopea testes with a double-labeled cy3 probe specific to a region on the 16S rRNA of cEina3 (cEina3_1416 5’- (Cy3)-ACAGCTCCTATTACGGCCACTGTC-(Cy3)-3’), while a general Cardinium 16S rRNA probe also labeled with cy3 (Ch1162 5′- (Cy3)-TTGACCTCATCCTCTCCT (Cy3-Q)−3′) were used to visualize both cEina2 and cEina3 (Supplementary Fig. 2). Male wasps raised at 27 °C were collected at different developmental stages based on the progression of melanization in the pupal sheath covering developing pupae. Stages included the mid-stage pupa (mottled black and white, 2–4 days post pupation), late-stage pupa (black, 4–6 days post pupation), pharate adults (adult wasps that have not yet emerged from the whitefly host, ~8 days post pupation), and adult wasps (two days post adult emergence). Males were stored at −80 °C until dissection.
Using a FISH protocol modified from Daims et al. (2005) and previously used to visualize Cardinium in Encarsia suzannae testes (Doremus et al. 2020), testes were dissected from males in 5 µL of 1X PBS on a coverslip and fixed in 20 µL of 4% paraformaldehyde for 20 min at room temperature. Fixative was then removed and samples were rinsed twice with ddH2O. Samples were dehydrated in an ethanol series with 3-minute rinses in 50, 80, and 96% ethanol. Prior to hybridization, dried samples were attached to the coverslip by covering them with a thin layer of 0.5% agarose (pulse field gel electrophoresis grade). Samples were then incubated with 10 µL hybridization buffer (25% formamide, 0.9 M NaCl, 20 mM Tris HCl (pH 8), 0.01% SDS) and 1 μL cy3 probes (30 ng/μL) in the dark at 46 °C for ∼2 h. After hybridization, samples were washed in washing buffer (0.15 M NaCl, 20 mM Tris HCl (pH 8), 5 mM EDTA (pH 8)) for ∼10 min in a 48 °C water bath. Optimal formamide concentration was determined via a formamide series with 0–35% formamide (Supplementary Fig. 1). Samples were then placed in ice cold ddH2O for 2-3 sec and dried with compressed air. They were then immediately stained with 10 μL DAPI (10 μg/mL in Tris-buffered saline with Tween 20 (TBST)) for 7 min and then rinsed 2-3 times with ddH2O. Coverslips with attached specimens were placed on glass slides (VWR International) with mounting media (80% glycerol, 20% TBST with 2% n-propyl-gallate (Sigma)). To confirm that the cEina3 probe specifically bound to the low-density Cardinium strain, we performed FISH on E. partenopea ovaries from doubly infected, cEina2 singly infected, and uninfected wasps using the cEina3_1416 probe (Supplementary Fig. 2). Cardinium infection (cEina2 strain) in singly infected testes was confirmed using the Cardinium specific Ch1162 probe in a separate FISH preparation (Supplementary Fig. 2).
Images were collected as z-stacks using a Zeiss LSM 880 inverted confocal microscope using a 63× lens with oil immersion. DAPI and cy3 fluorescence were captured using a 405 and 561 nm excitation beam, respectively. Here we present representative images that capture the center depth of the testis. Testes from at least five males of each stage were prepared and imaged. Images were processed using Zen Blue (Zeiss) or ImageJ imaging software.
Results
The low-density cEina3 Cardinium strain is required for CI induction and rescue in E. partenopea
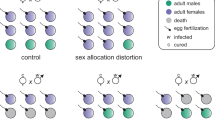
To determine whether one or both of the Cardinium strains infecting E. partenopea was responsible for causing CI, we crossed cEina2-singly infected, doubly infected (cEina2 and cEina3) and uninfected wasps in all combinations. Female progeny production varied significantly across infection- and cross-types (Fig. 1; Supplementary Table S2; ANOVA F = 10.79, p ≤ 0.0001). As expected, crosses between doubly infected males and uninfected females (CI cross) produced significantly fewer female offspring than crosses between uninfected male and female wasps (Fig. 1; p ≤ 0.0001). Also as expected, female progeny production was restored in rescue crosses between doubly infected male and female wasps and did not differ significantly from control (uninfected X uninfected) crosses, indicative of CI rescue (Fig. 1; p = 0.999). Crosses between cEina2 singly infected males and uninfected females did not produce significantly fewer female offspring (Fig. 1; p = 0.995), indicating that cEina2 alone does not induce CI. Crosses between doubly infected males and cEina2 singly infected females also produced significantly fewer female offspring than all other compatible crosses (Fig. 1; p ≤ 0.0001), indicating that cEina2 also does not rescue CI caused by cEina3. Female offspring production did not vary significantly for all other cross types, indicating that Cardinium infection did not alter reproduction in compatible crosses (Fig. 1; p = 0.99 for all crosses). The production of male progeny also did not differ across infection or cross-type, indicating that CI in incompatible crosses did not arise from increased male production, but rather female-specific mortality (Fig. 1; ANOVA F = 0.5, p = 0.849; Vavre et al. 2000; Gebiola et al. 2016b). CI-induced female offspring mortality caused the offspring sex ratio of CI crosses to be extremely male-biased relative to compatible crosses, with most CI crosses producing entirely male offspring broods (Fig. 1B; Supplementary Table S3; Logistic regression Funinfected-CI = 60.9, Puninfected-CI < 0.0001; FcEina2-infectedCI = 62.3, PcEina2-infectedCI < 0.0001) Together, these data indicate that the low-density cEina3 strain, present in doubly infected wasps, is responsible for CI and rescue of CI in E. partenopea.

A Number of female (cyan boxes) and male (purple boxes) offspring produced in crosses, analyzed using ANOVA with Tukey’s HSD post-hoc test. B Resulting sex ratio from crosses, analyzed with logistic regression. Significant differences are represented with asterisks. ***p value of <0.0001, n = 9–10 for all crosses.
Cardinium shows dynamic changes in infection density across male E. partenopea development
To estimate infection frequencies of each of the two Cardinium strains present in E. partenopea during male development, we performed qPCR on males from different life stages (3rd instar larvae, white pupa, mottled pupa, black pupa, adult). We found that both Cardinium strains (cEina2 and cEina3) exhibited similar infection density in 3rd instar larvae (Fig. 2; Supplementary Table S4; Linear model p = 0.698). However, the titers of the symbionts significantly diverged as the wasp hosts progressed through the pupal development (Fig. 2; LM p < 0.01 for comparisons between strains in the black and adult stages). This titer difference was driven by the change in cEina2 titer, which increased significantly at the onset of pupation (Fig. 2; LM plarva-white = 0.05), midway through pupation (Fig. 2; LM pmottled-black = 0.02) and again at the end of pupation (Fig. 2; LM pblack-adult = 0.05). Unlike cEina2, the titer of the cEina3 strain did not change significantly across pupal development (Fig. 2; LM p > 0.391 for all pupal stage comparisons) and decreased significantly in titer between the pupal and adult stage (Fig. 2; LM pblack-adult = 0.01).

Cardinium density was measured relative to host cells using the single copy ydhE gene (Cardinium) and ef1a gene (host). Cyan boxes represent cEina2 and purple boxes represent cEina3. Samples were performed with three technical replicates, n = 5. Densities were analyzed using a linear model followed by Tukey’s HSD with Benjamini–Hochberg corrected p values. Significant differences between the densities of the strains at different life stages are denoted by asterisks (*p value < 0.05). For simplicity, only significant differences between cEina2 and cEina3 at each life stage are shown.
Antibiotic exposure during the male wasp adult stage does not change severity of CI
To test whether CI modification occurs prior to the E. partenopea adult stage, as in the cEper1 Cardinium CI symbiosis with E. suzannae, we fed doubly infected adult male E. partenopea antibiotics for seven days prior to performing CI and rescue crosses (Doremus et al. 2020). The antibiotic treatment significantly reduced the density of both strains of Cardinium in males, compared with honey-fed Cardinium-infected control wasps (Fig. 3A; MWU-test p value < 0.001 for all comparisons). Despite the successful reduction of Cardinium abundance, CI crosses still resulted in significantly fewer female progeny than in compatible rescue crosses, regardless of diet treatment (Fig. 3B; Supplementary Table S2; ANOVA F = 11.62, p < 0.0001; PHoney-CI = 0.035, PRifampicin-CI < 0.0001). Female offspring production from CI crosses also did not differ between honey and rifampicin fed treatments (Fig. 3B; P = 0.99). The number of male offspring produced was generally consistent in all treatments, except in the antibiotic-fed CI cross, which produced more male offspring than did honey-fed rescue crosses (Fig. 3B; p = 0.007). The progeny of the CI crosses involving both honey- and rifampicin-fed males exhibited strongly male-biased offspring sex ratios consistent with the female-mortality type of CI (Fig. 3C; Supplementary Table S3; FHoney = 12.4, PHoney = 0.001; FRifampicin = 80.8, PRifampicin < 0.0001). The resulting sex ratios of CI crosses did not differ between the honey and antibiotic treatments (Fig. 3C; F = 2.3, P = 0.143).

A Cardinium densities (both cEina2 + cEina3) in co-infected males fed honey or rifampicin + honey for seven days. Cyan boxes refer to cEina2 and purple boxes refer to cEina3 density. Densities were estimated using qPCR with primers for the single copy ydhE symbiont genes and host ef1-alpha gene and analyzed using Mann–Whitney U tests. Significantly different densities after diet treatment are indicated with asterisks (**p value < 0.001), n = 5 with three technical replicates. B Number of male and female offspring produced in crosses with honey-fed control or rifampicin-fed males. Rescue crosses between infected male and females were included to control for effects of antibiotic diet on male viability. Parental infection status is indicated by either filled (infected) or open (uninfected) male and female symbols. Offspring counts were analyzed using ANOVA with Tukey’s HSD post-hoc test. Significant differences are denoted by different uppercase (female offspring) and lowercase (male offspring) letters. C Resulting sex ratio from crosses in B. The proportions of adults that were males were analyzed using logistic regression. Significant differences in sex ratios are indicated with asterisks (***p value < 0.0001), n = 10–15 for all crosses in B and C.
Cardinium localization within developing E. partenopea testes
Like E. suzannae, E. partenopea sperm begin entering spermiogenesis, the final elongation stage of sperm development, approximately halfway through pupal development when the wasp cuticle begins to darken (“mottled” stage; Fig. 4A, B; Doremus et al. 2020). At this point in E. partenopea development, sperm cells that have begun spermiogenesis have small, hypercondensed nuclei that are aligned in parallel “cup” formations and, in some cases, nuclei and sperm tails have also begun to elongate (Fig. 4A, B). In E. suzannae, Cardinium cells infect developing sperm and symbiont cells are also observed trailing elongating nuclei as sperm jettison the bulk of their cytoplasmic content during spermiogenesis (Ferree et al. 2019; Doremus et al. 2020). Unlike the E. suzannae system, cEina3 does not appear to infect developing sperm cells, as symbiont cells are not seen associating with spermatogonia nuclei nor do they trail elongating spermatid nuclei (Fig. 4A, B). Instead, cEina3 cells largely localize at the testis base and the downstream reproductive tract during pupal development (Fig. 4B, C). The symbiont cells in the testis base appear to associate with somatic cell nuclei, as these nuclei are larger than sperm nuclei, do not elongate, and are not grouped into closely associating cysts with sperm cells (Fig. 4C). The cEina3 symbiont is most prevalent in the downstream reproductive tract, where symbiont cells associate with somatic nuclei of the vas deferens and the seminal vesicle, a sperm storage organ that does not yet hold mature sperm in the mottled pupal stage (Fig. 4C).

DAPI-stained host nuclei are colored cyan and cEina3 tagged with the cEina3-1416 probe double-labeled with cy3 are colored purple in all images. Testes and downstream reproductive tract are oriented with the apical tip of the testis at the top of each image, the testis base and downstream structures are at the bottom of images (when shown). A Full image of the testis from a mottled 2–4 day old pupa, with a white box representing the region shown in image B. B Midsection of testis showing multiple sperm cysts at various stages of development. The solid arrowhead points to sperm cysts undergoing nuclear and cellular elongation, with sperm nuclei aligned and trailed by cytoplasmic voids (unfilled arrowheads). No cEina3 cells are present. C 2–4 day old pupal seminal vesicle (brackets), vas deferens (white arrowhead), and testis base (unfilled arrowhead) showing somatic cells infected with cEina3. D Full image of the testis from a black 4–6 day old pupa with the white box representing the region shown in image E. E Midsection and base of testis showing multiple sperm cysts at various stages of development. The unfilled arrowhead points to extending cytoplasm of developing sperm tails (shown as a dark void in the image) trailing elongating spermatid nuclei. The white arrowhead shows cEina3 cells associating with larger nuclei of somatic cells. F Black-stage seminal vesicles (circled) and downstream reproductive tract of a black stage pupa. cEina3 cells are associating with somatic nuclei of the seminal vesicle and several mature sperm (thread-like nuclei) have migrated into the vesicle at this stage. G Another testis of a black-stage pupa, showing cEina3 cells associating with large nuclei (white arrowheads), possibly belonging to somatic cyst cells that associate with developing sperm cells (unfilled arrowheads). H Lower reproductive tract of pharate adult testes, ~6–8 days post pupation, with boxes representing regions magnified in I and J. I Seminal vesicle of pharate adult wasps, now filled with mature sperm cells. cEina3 cells are present, infecting epithelial cells forming the distal portion of the seminal vesicle (arrowhead). J Downstream reproductive tract of pharate adult wasps. cEina3 are present infecting the junction of the reproductive tract (bottom of image, white arrowhead) leading to the aedeagus. K Full adult testis, 2 days post emergence. The testis is outlined in white for clarity, with tip of the testis on the right corner of the image. The testis at this stage is reduced in size and content (~60 µm long compared to ~150 µm long for previous stages), although some mature sperm and developmentally arrested spermatogonia (unfilled arrowhead) are present, with cEina3 cells nearby. cEina3 cells can also be found associating with larger somatic cell nuclei (white arrowhead). Mature sperm now fill the seminal vesicle, which has become enlarged relative to earlier stages.
As E. partenopea progresses through the pupal stage, its pupal case darkens completely (“black stage”) and a majority of its sperm cells progress into spermiogenesis (Fig. 4D). The elongated nuclei of these cells are trailed by dark voids in the testis representing sperm tails; these cells now take up most of the inner space of the testis (Fig. 4D). cEina3 cells continue to associate primarily with larger nuclei at the testis base and with somatic cells of the downstream reproductive tract and the seminal vesicle, which now continues several mature sperm cells (Fig. 4E, F). Cardinium cells also associate with large nuclei of cells that proceed elongating spermatid cysts (Fig. 4G). These nuclei may belong to somatic cyst cells, which enclose sperm cells during development and possibly provision them with resources (Fuller 1993). Some symbiont cells also infect host cells at the apical tip of the testes, possibly infecting the germ cell hub, although this localization pattern is not uniform across all testes (Fig. 4D). Approximately 24–48 h later, pupal E. partenopea molt into a pharate adult stage, in which the adult wasp remains in the whitefly cuticle unmoving for ~24 h. Most sperm cells complete development and migrate into the seminal vesicles during the black pupal and pharate adult stages (Fig. 4D, G, H). cEina3 continues to localize primarily in the epithelial cells of the seminal vesicle and vas deferens during the pharate adult stage (Fig. 4I, J).
The testes of emerged adult E. partenopea are reduced in size and number of sperm cells compared to earlier life stages and contain seminal vesicles full of mature sperm indicating that, like E. suzannae, E. partenopea is prospermatogenic and produces most of its lifetime sperm load during the pupal stage (Fig. 4A, D, G, K; Doremus et al. 2020). Several cysts of elongate sperm remain in the adult testes, possibly to serve as a reservoir in the unlikely event of sperm depletion. (Fig. 4K; Doremus et al. 2020). The cEina3 symbiont can be found throughout the adult testis associating with a variety of nuclei that may belong to somatic cells, mature sperm that have yet to migrate into the seminal vesicle, and developmentally arrested cysts of spermatogonia (Fig. 4K).
Discussion
We found that the low-density Cardinium strain cEina3 is responsible for a severe CI phenotype in the coinfected parasitoid wasp E. partenopea (Fig. 1). The second Cardinium strain coinfecting this host, cEina2, does not manipulate host reproduction, much like closely related asymptomatic Cardinium strains in whiteflies (Fang et al. 2014) and in E. inaron (White et al. 2009, Stouthamer et al. 2019). Like the Cardinium-E. suzannae CI symbiosis, cEina3 fatally modifies male E. partenopea during the pupal stage when cEina3 reaches its highest density (Figs. 2 and 3; Doremus et al. 2020). There is also indirect evidence suggesting that another CI symbiont, Wolbachia, induces CI during the pupal stage of the parasitoid, Nasonia vitripennis (Bordenstein and Bordenstein 2011); it is possible that pupal modification is a common trend shared amongst CI symbionts infecting parasitoid wasps. This shared timing is possibly the result of host sperm development, as many parasitic wasps in the Chalcidoidea, including both Encarsia species (Fig. 4; Doremus et al. 2020) and N. vitripennis (Ferree et al. 2019), produce the bulk of their sperm during pupation (Boivin et al. 2005). Using FISH microscopy, we also found that, unlike most CI Wolbachia symbionts and the one other CI Cardinium studied (Clark et al. 2002; Clark et al. 2008; Doremus et al. 2020), cEina3 does not infect sperm cells and instead localizes in the epithelial tissue of the seminal vesicle (sperm storage organ), downstream reproductive tract, and somatic cyst cells within the testis (Fig. 4). Symbiont localization can give important clues for mechanistic function (Clark et al. 2002, 2003, 2008) and this localization pattern appears to involve relatively few cells and is downstream of most sperm maturation, perhaps indicating that modification targets mature sperm.
Antibiotic treatment of adult males did not influence CI strength, demonstrating that modification is finished by adult eclosion. This was also found in a second Cardinium-Encarsia CI interaction (Doremus et al. 2020), but the timing of modification has received little explicit attention in Wolbachia CI symbioses (Snook et al. 2000; Clark et al. 2008; Bordenstein and Bordenstein 2011). It remains to be seen if this is a general trend across Cardinum infecting other arthropod groups (Gotoh et al. 2007; Nakamura et al. 2012; Nguyen et al. 2017). This seems unlikely, however, as the importance of the Encarsia pupal stage for CI modification is ultimately due to the prospermatogenic development of these wasps (Doremus et al. 2020). In general, antibiotic trials may be useful for determining the timing of CI induction for other CI systems.
The near complete lethality of cEina3-induced CI is especially striking in light of how few cEina3 cells infect the male reproductive system of E. partenopea. This is in contrast with the cEper1 Cardinium strain of E. suzannae, which causes milder and more variable CI than cEina3 despite occurring at a higher infection density and directly infecting sperm cells (Doremus et al. 2019, 2020). This stark difference in CI severity could be a result of one or more factors. First, the duration of the pupal stage is important for determining CI severity in E. suzannae (Doremus et al. 2019, 2020), and a slower host developmental rate could explain the stronger CI displayed by E. partenopea. Encarsia suzannae has both a shorter overall pupal stage (~5–6 days) compared to E. partenopea (~6–8 days), and a faster rate of sperm development (Doremus et al. 2020). In E. suzannae, sperm cells fill the seminal vesicle during the pupal stage, while in E. partenopea this occurs primarily during the transition between the pupal and adult stages (i.e., as a pharate adult), prolonging the time available for modification.
Alternatively, it is possible that cEina3 is more transcriptionally active than cEper1 to compensate for its reduced abundance. Another possibility lies in variation of the CI factors themselves, as a recent study of a weak CI Wolbachia strain infecting Drosophila yakuba showed that a single point mutation in cifB was responsible for reduced CI penetrance (Beckmann et al. 2021).
The localization of cEina3 represents a third distinct localization pattern observed in CI symbioses, whereby the symbiont localizes in somatic cells at the testis base and surrounding the seminal vesicle (Clark et al. 2002, 2008; Doremus et al. 2020). In Wolbachia symbioses with drosophilid flies and the parasitoid Nasonia, and in the Cardinium- E. suzannae symbiosis, the CI symbiont infects developing sperm cells to varying degrees but is lost during spermiogenesis, the sperm elongation phase of development (Clark et al. 2002, 2008). A second pattern is found in the beetle host Chelymorpha alterans, which is co-infected with two CI-inducing Wolbachia strains, with neither Wolbachia strain present in developing sperm cells and instead both infect the outer somatic sheath cells that encapsulate the testicular follicles (Clark et al. 2008), presumably allowing secretion of a modification factor into cysts of developing sperm.
These different localization patterns may reflect variation in the CI modification factors, with some CI factor types directly altering sperm during development prior to the removal of the symbiont from the cell, while others are packaged internally into the sperm or coat the sperm surface to be delivered directly into the egg cytoplasm (Shropshire et al. 2020; Doremus et al. 2020). In the case of Wolbachia CI, there is substantial variation in the cif genes responsible for CI across different Wolbachia strains (Lindsey et al. 2018), although how these different cif types interact with male sperm remains a mystery. In the case of Cardinium, the factors responsible for CI are unknown. The different localization patterns exhibited by the CI strains in Encarsia wasps suggest there may similarly be a variety of CI factors used by different Cardinium strains (Doremus et al. 2020). While the cEper1 strain that localizes within sperm cysts may modify each sperm directly and internally (Doremus et al. 2020), the cEina3 symbiont may instead introduce the modification factor into the duct leading from the testis to the seminal vesicle or directly into the seminal vesicle lumen, enriching this region with the modification factor (Fig. 5). The factor would then either diffuse into mature sperm as they enter the seminal vesicle or bind to the outside surface of the sperm. Once bound, the factor could be released upon sperm entry into an egg cell and the dissociation of the male nucleus from the sperm tail (Karr 1991; Lassy and Karr 1996).

In model 1, Cardinium residing in somatic cyst cells modifies developing sperm cells during spermiogenesis. In model 2, Cardinium residing in somatic cells at the testis base modify matured sperm as they migrate out of the testis. In model 3, Cardinium residing in somatic cells forming the seminal vesicle modify mature sperm as they collect inside the seminal vesicle lumen. Cells with white cytoplasm are germ/sperm cells and gray cytoplasm are somatic cells, host nuclei are cyan, Cardinium cells are purple, and the modification factor is represented by yellow diamonds.
The cEina3 symbiont could also utilize a similar strategy in the testis base to modify sperm within the testis itself, either by using somatic cyst cells to distribute the CI factor among developing sperm cells, which share cytoplasm during development, or by releasing a diffusible CI factor into the extracellular space of the testis (Fig. 5). This strategy is similar to one previously suggested for CI Wolbachia in Nasonia, in which the Wolbachia symbiont may induce nearly complete CI despite infecting a low percentage of developing sperm cells by using a diffusible modification factor (Clark et al. 2008). Some cEina3 cells also infect somatic cyst cells that closely associate and interact with developing sperm. The symbiont could use these somatic cysts cells to deliver its modification factor into maturing sperm cells during spermiogenesis, although cEina3 does not appear to consistently infect each somatic cyst cell (Fig. 5). While it is unlikely that cEina3 infects sperm cells during development based on the localization of the symbiont relative to sperm cell nuclei, we are unable to confirm cell boundaries with the FISH imaging used in this study. Confirmation of cEina3 infection in somatic cells will require higher magnification imaging in a future study.
While cEina3 is responsible for CI in E. partenopea, it remains unclear what, if anything, cEina2 provides for its parasitoid host. The cEina2 strain is closely related to two other, apparently asymptomatic Cardinium strains (cBtQ1 in whiteflies and cEina1 in E. inaron) (White et al. 2009; Fang et al. 2014; Stouthamer et al. 2019). All three of these asymptomatic strains infect either whiteflies or parasitoids that attack whiteflies, and all three exist as coinfections with additional symbionts (White et al. 2009; Guo et al. 2014; Fujiwara et al. 2015). In the case of cEina1 and cEina2, both asymptomatic strains coinfect Encarsia hosts with another CI-inducing symbiont (Wolbachia in E. inaron (White et al. 2009) and cEina3 in E. partenopea in the current study). Given that cEina2 densely infects both the testis and seminal vesicle (Supplementary Fig. 3), it is possible that cEina2 somehow amplifies the CI phenotype in E. partenopea, perhaps by making sperm cells more amenable to modification. Unfortunately, as we were unable to establish cEina3 as a single infection, we were not able to test the strength of CI caused by cEina3 alone.
Despite this possibility, the simplest scenario is that cEina2 does not contribute to CI, and instead cEina2 and the other asymptomatic Cardinium strains have spread through host populations by conferring a host benefit under specific conditions. A whitefly-associated Cardinium strain was recently found to increase host thermal tolerance, suggesting that some Cardinum strains may offer additional benefits to their host beyond manipulating host reproduction (Yang et al. 2021). Alternatively, cEina2 and other asymptomatic Cardinium strains may spread by hitchhiking off the benefits (in whiteflies) or reproductive manipulations (in Encarsia species) conferred by their coinfecting symbiont partners (White et al. 2009; Smith et al. 2015; Doremus and Oliver 2017). Similar hitchhiking effects have been reported for other maternally inherited elements like mitochondria and coinfecting symbionts, and may be a more widespread trend given the frequency of coinfection symbioses in nature (Jiggins 2003; Hurst and Jiggins 2005; Doremus and Oliver 2017; Jaenike 2012; Mathé-Hubert et al. 2019; Smith et al. 2015).
In summary, we found that a cryptic, low-density Cardinium strain (cEina3) is responsible for CI in coinfected E. partenopea. The high-density strain (cEina2) is apparently asymptomatic and may act as a hitchhiker, spreading as a result of the effects of cEina3-induced CI. We also found that while cEina3 appears to modify sperm during the pupal stage like previously studied Cardinium strains, this strain shows a markedly different localization pattern in male hosts and does not infect developing sperm cells. Instead, cEina3 infects the base of the testis and the seminal vesicle, where it may modify newly matured sperm cells as they await transfer to females. It remains to be determined whether the variation in localization pattern in these two related symbioses is indicative of different modification strategies and CI factors.
Data availability
Confocal images are available upon request and data for CI crosses and symbiont density estimates will be archived in Dryad https://doi.org/10.5061/dryad.6wwpzgn0j.
References
Balhorn R (2007) The protamine family of sperm nuclear proteins. Genome Biol 8(9):227. https://doi.org/10.1186/gb-2007-8-9-227
Beckmann JF, Ronau JA, Hochstrasser M (2017) A Wolbachia deubiquitylating enzyme induces cytoplasmic incompatibility. Nat Microbiol 2:5
Beckmann JF, Van Vaerenberghe K, Akwa DE, Cooper BS (2021) A single mutation weakens symbiont-induced reproductive manipulation through reductions in deubiquitylation efficiency. Proc Nat Acad Sci 118(39):e2113271118
Boivin G, Jacob S, Damiens D (2005) Spermatogeny as a life-history index in parasitoid wasps. Oecologia 143(2):198–202
Bordenstein SR, O’Hara FP, Werren JH (2001) Wolbachia-induced incompatibility precedes other hybrid incompatibilities in Nasonia. Nature 409(6821):707–710
Bordenstein SR, Bordenstein SR (2011) Temperature affects the tripartite interactions between bacteriophage WO, Wolbachia, and cytoplasmic incompatibility. PLoS One 6(12):e29106
Breeuwer JA (1997) Wolbachia and cytoplasmic incompatibility in the spider mites Tetranychus urticae and T. turkestani. Heredity 79(1):41–47
Breeuwer JA, Werren JH (1990) Microorganisms associated with chromosome destruction and reproductive isolation between two insect species. Nature 346(6284):558–560
Clark M, Bailey-Jourdain C, Ferree P, England S, Sullivan W, Windsor DM et al. (2008) Wolbachia modification of sperm does not always require residence within developing sperm. Heredity 101(5):420–428
Clark ME, Veneti Z, Bourtzis K, Karr TL (2002) The distribution and proliferation of the intracellular bacteria Wolbachia during spermatogenesis in Drosophila. Mech Dev 111(1-2):3–15
Clark ME, Veneti Z, Bourtzis K, Karr TL (2003) Wolbachia distribution and cytoplasmic incompatibility during sperm development: the cyst as the basic cellular unit of CI expression. Mech Dev 120(2):185–198
Daims H, Stoecker K, Wagner M (2005) Fluorescent in situ hybridization for the detection of prokaryotes. In Molecular Microbial Ecology, ed. Osborn AM, Smith CJ, pp. 208-228 Taylor & Francis Group, New York, NY.
Doremus MR, Hunter MS (2020) The saboteur’s tools: Common mechanistic themes across manipulative symbioses. Adv Insect Physiol 58:317–353
Doremus MR, Kelly SE, Hunter MS (2019) Exposure to opposing temperature extremes causes comparable effects on Cardinium density but contrasting effects on Cardinium-induced cytoplasmic incompatibility. PLoS Pathog 15(8):e1008022. https://doi.org/10.1371/journal.ppat.1008022
Doremus MR, Oliver KM (2017) Aphid heritable symbiont exploits defensive mutualism. Appl Environ Microbiol 83(8):e03276–16. https://doi.org/10.1128/AEM.03276-16
Doremus MR, Stouthamer CM, Kelly SE, Schmitz-Esser S, Hunter MS (2020) Cardinium localization during its parasitoid wasp host’s development provides insights into cytoplasmic incompatibility. Front Microbiol 11(3153):606399
Fang Y-W, Liu L-Y, Zhang H-L, Jiang D-F, Chu D (2014) Competitive ability and fitness differences between two introduced populations of the invasive whitefly Bemisia tabaci Q in China. PLoS ONE 9(6):e100423
Ferree PM, Aldrich JC, Jing XYA, Norwood CT, Van Schaick MR, Cheema MS et al. (2019) Spermatogenesis in haploid males of the jewel wasp Nasonia vitripennis. Sci Rep 9:12194. https://doi.org/10.1038/s41598-019-48332-9
Fujiwara A, Maekawa K, Tsuchida T (2015) Genetic groups and endosymbiotic microbiota of the Bemisia tabaci species complex in Japanese agricultural sites. J Appl Entomol 139(1-2):55–66
Fuller MT (1993) Spermatogenesis. In The Development of Drosophila, ed. Bate M, Martinez-Arias A, pp. 71–147. Cold Spring Harbor, NY: Cold Spring Harbor Lab. Press
Gebiola M, Giorgini M, Kelly SE, Doremus MR, Ferree PM, Hunter MS (2017) Cytological analysis of cytoplasmic incompatibility induced by Cardinium suggests convergent evolution with its distant cousin Wolbachia. Proc R Soc B Biol Sci 284(1862):20171433. https://doi.org/10.1098/rspb.2017.1433
Gebiola M, Kelly SE, Hammerstein P, Giorgini M, Hunter MS (2016a) “Darwin’s corollary” and cytoplasmic incompatibility induced by Cardinium may contribute to speciation in Encarsia wasps (Hymenoptera: Aphelinidae). Evolution 70(11):2447–2458
Gebiola M, White JA, Cass BN, Kozuch A, Harris LR, Kelly SE et al. (2016b) Cryptic diversity, reproductive isolation and cytoplasmic incompatibility in a classic biological control success story. Biol J Linn Soc 117(2):217–230
Gotoh T, Noda H, Ito S (2007) Cardinium symbionts cause cytoplasmic incompatibility in spider mites. Heredity 98(1):13–20
Guo H, Qu Y, Liu X, Zhong W, Fang J (2014) Female-biased symbionts and Tomato Yellow Leaf Curl Virus infections in Bemisia tabaci. PLOS ONE 9:e84538
Hunter MS, Perlman SJ, Kelly SE (2003) A bacterial symbiont in the Bacteroidetes induces cytoplasmic incompatibility in the parasitoid wasp Encarsia pergandiella. Proc R Soc B Biol Sci 270(1529):2185–2190
Hurst GD, Jiggins FM (2005) Problems with mitochondrial DNA as a marker in population, phylogeographic and phylogenetic studies: the effects of inherited symbionts. Proc R Soc B Biol Sci 272(1572):1525–1534
Jaenike J (2012) Population genetics of beneficial heritable symbionts. Trends Ecol Evol 27(4):226–232
Jiggins FM (2003) Male-killing Wolbachia and mitochondrial DNA: selective sweeps, hybrid introgression and parasite population dynamics. Genetics 164(1):5–12
Karr TL (1991) Intracellular sperm/egg interactions in Drosophila: a three-dimensional structural analysis of a paternal product in the developing egg. Mech Dev 34(2):101–111
Konig K, Zundel P, Krimmer E, Konig C, Pollmann M, Gottlieb Y et al. (2019) Reproductive isolation due to prezygotic isolation and postzygotic cytoplasmic incompatibility in parasitoid wasps. Ecol Evol 9(18):10694–10706
Lassy CW, Karr TL (1996) Cytological analysis of fertilization and early embryonic development in incompatible crosses of Drosophila simulans. Mech Dev 57(1):47–58
LePage DP, Metcalf JA, Bordenstein SR, On JM, Perlmutter JI, Shropshire JD et al. (2017) Prophage WO genes recapitulate and enhance Wolbachia-induced cytoplasmic incompatibility. Nature 543(7644):243–247
Li T-P, Zhou C-Y, Zha S-S, Gong J-T, Xi Z, Hoffmann AA et al. (2020) Stable establishment of Cardinium spp. in the brown planthopper Nilaparvata lugens despite decreased host fitness. Appl Environ Microbiol 86(4):e02509–02519
Lindsey ARI, Rice DW, Bordenstein SR, Brooks AW, Bordenstein SR, Newton ILG (2018) Evolutionary genetics of cytoplasmic incompatibility genes cifA and cifB in prophage WO of Wolbachia. Gen Biol Evol 10(2):434–451
Mann E, Stouthamer CM, Kelly SE, Dzieciol M, Hunter MS, Schmitz-Esser S (2017) Transcriptome sequencing reveals novel candidate genes for Cardinium hertigii- caused cytoplasmic incompatibility and host-cell interaction. mSystems 2:6
Mathé-Hubert H, Kaech H, Hertaeg C, Jaenike J, Vorburger C (2019) Nonrandom associations of maternally transmitted symbionts in insects: the roles of drift versus biased cotransmission and selection. Mol Ecol 28(24):5330–5346
Moreira LA, Iturbe-Ormaetxe I, Jeffery JA, Lu G, Pyke AT, Hedges LM et al. (2009) A Wolbachia symbiont in Aedes aegypti limits infection with dengue, Chikungunya, and Plasmodium. Cell 139(7):1268–1278
Mouton L, Henri H, Boulétreau M, Vavre F (2005) Multiple infections and diversity of cytoplasmic incompatibility in a haplodiploid species. Heredity 94(2):187–192
Nakamura Y, Yukuhiro F, Matsumura M, Noda H (2012) Cytoplasmic incompatibility involving Cardinium and Wolbachia in the white-backed planthopper Sogatella furcifera (Hemiptera: Delphacidae). Appl Entomol Zool 47(3):273–283
Nguyen DT, Morrow JL, Spooner-Hart RN, Riegler M (2017) Independent cytoplasmic incompatibility induced by Cardinium and Wolbachia maintains endosymbiont coinfections in haplodiploid thrips populations. Evolution 71(4):995–1008
O’Neill SL, Hoffman A, Werren JH (1997). Influential passengers: inherited microorganisms and arthropod reproduction. Oxford University Press.
Oliver KM, Martinez AJ (2014) How resident microbes modulate ecologically-important traits of insects. Curr Opin Insect Sci 4:1–7
Penz T, Schmitz-Esser S, Kelly SE, Cass BN, Muller A, Woyke T et al. (2012) Comparative genomics suggests an independent origin of cytoplasmic incompatibility in Cardinium hertigii. PLoS Genet 8(10):e1003012. https://doi.org/10.1371/journal.pgen.1003012
Perlman SJ, Dowdy NJ, Harris LR, Khalid M, Kelly SE, Hunter MS (2014) Factors affecting the strength of Cardinium-induced cytoplasmic incompatibility in the parasitic wasp Encarsia pergandiella (Hymenoptera: Aphelinidae). Micro Ecol 67(3):671–678
Polaszek A, Evans G, Bennett F (1992) Encarsia parasitoids of Bemisia tabaci (Hymenoptera: Aphelinidae, Homoptera: Aleyrodidae): a preliminary guide to identification. Bull Entomol Res 82(3):375–392
R Core Team (2021). R foundation for statistical computing: Vienna, Austria.
Raja SJ, Renkawitz-Pohl R (2005) Replacement by Drosophila melanogaster protamines and Mst77F of histones during chromatin condensation in late spermatids and role of sesame in the removal of these proteins from the male pronucleus. Mol Cell Biol 25(14):6165–6177
Rosenwald LC, Sitvarin MI, White JA (2020) Endosymbiotic Rickettsiella causes cytoplasmic incompatibility in a spider host. Proc R Soc B Biol Sci 287(1930):20201107
Ryan PA, Turley AP, Wilson G, Hurst TP, Retzki K, Brown-Kenyon J et al. (2019) Establishment of wMel Wolbachia in Aedes aegypti mosquitoes and reduction of local dengue transmission in Cairns and surrounding locations in northern Queensland, Australia. Gates Open Res 3:1547. https://doi.org/10.12688/gatesopenres.13061.2
Shoemaker DD, Katju V, Jaenike J (1999) Wolbachia and the evolution of reproductive isolation between Drosophila recens and Drosophila subquinaria. Evolution 53(4):1157–1164
Shropshire JD, Leigh B, Bordenstein SR (2020) Symbiont-mediated cytoplasmic incompatibility: what have we learned in 50 years? eLife 9:e61989
Shropshire JD, On J, Layton EM, Zhou H, Bordenstein SR (2018) One prophage WO gene rescues cytoplasmic incompatibility in Drosophila melanogaster. Proc Natl Acad Sci USA 115(19):4987–4991
Smith AH, Łukasik P, O’Connor MP, Lee A, Mayo G, Drott MT et al. (2015) Patterns, causes and consequences of defensive microbiome dynamics across multiple scales. Mol Ecol 24(5):1135–1149
Snook RR, Cleland SY, Wolfner MF, Karr TL (2000) Offsetting effects of Wolbachia infection and heat shock on sperm production in Drosophila simulans: analyses of fecundity, fertility and accessory gland proteins. Genetics 155(1):167–178
Stouthamer CM (2018). The evolution and comparative genomics of the reproductive manipulator Cardinium hertigii. Doctoral dissertation, The University of Arizona.
Stouthamer CM, Kelly S, Hunter MS (2018) Enrichment of low-density symbiont DNA from minute insects. J Microbiol Met 151:16–19
Stouthamer CM, Kelly SE, Mann E, Schmitz-Esser S, Hunter MS (2019) Development of a multi-locus sequence typing system helps reveal the evolution of Cardinium hertigii, a reproductive manipulator symbiont of insects. BMC Microbiol 19(1):1–13
Takano SI, Tuda M, Takasu K, Furuya N, Imamura Y, Kim S et al. (2017) Unique clade of alphaproteobacterial endosymbionts induces complete cytoplasmic incompatibility in the coconut beetle. Proc Natl Acad Sci USA 114(23):6110–6115
Takano S-i, Gotoh Y, Hayashi T (2021) “Candidatus Mesenet longicola”: novel endosymbionts of Brontispa longissima that induce cytoplasmic incompatibility. Micro Ecol 82:512–522
Turelli M (1994) Evolution of incompatibility‐inducing microbes and their hosts. Evolution 48(5):1500–1513
Vavre F, Fleury F, Varaldi J, Fouillet P, Bouleatreau M (2000) Evidence for female mortality in Wolbachia‐mediated cytoplasmic incompatibility in haplodiploid insects: epidemiologic and evolutionary consequences. Evolution 54(1):191–200
Weinert LA, Araujo-Jnr EV, Ahmed MZ, Welch JJ (2015) The incidence of bacterial endosymbionts in terrestrial arthropods. Proc R Soc B Biol Sci 282:1807
White JA, Kelly SE, Perlman SJ, Hunter MS (2009) Cytoplasmic incompatibility in the parasitic wasp Encarsia inaron: disentangling the roles of Cardinium and Wolbachia symbionts. Heredity 102(5):483–489
Wu K, Hoy MA (2012) Cardinium is associated with reproductive incompatibility in the predatory mite Metaseiulus occidentalis (Acari: Phytoseiidae). J Invert Pathol 110(3):359–365
Yang K, Yuan MY, Liu Y, Guo CL, Liu TX, Zhang YJ et al (2021). First evidence for thermal tolerance benefits of the bacterial symbiont Cardinium in an invasive whitefly, Bemisia tabaci. Pest Man Sci https://doi.org/10.1002/ps.6543
Zchori‐Fein E, Perlman SJ (2004) Distribution of the bacterial symbiont Cardinium in arthropods. Mol Ecol 13(7):2009–2016
Zheng X, Zhang D, Li Y, Yang C, Wu Y, Liang X et al. (2019) Incompatible and sterile insect techniques combined eliminate mosquitoes. Nature 572(7767):56–61
Zug R, Hammerstein P (2012) Still a host of hosts for Wolbachia: analysis of recent data suggests that 40% of terrestrial arthropod species are infected. PloS One 7(6):e38544
Acknowledgements
Many thanks go to Massimo Giorgini, who sent us this wasp, and without whom we never would have known to study it. Thanks also to Patty Jansma at the Marley Light Microscopy Facility (University of Arizona) for help with the confocal imaging.
Author information
Authors and Affiliations
Contributions
MRD designed, performed, and analyzed all experiments. CMS and SEK helped with initial antibiotic curing, CI assays, and developing the FISH protocol. SSE helped design fluorescent probes and helped develop the FISH protocol. MSH helped design and analyze experiments. All authors helped write and edit the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Associate editor: Louise Johnson.
Supplementary information
Rights and permissions
About this article

Cite this article
Doremus, M.R., Stouthamer, C.M., Kelly, S.E. et al. Quality over quantity: unraveling the contributions to cytoplasmic incompatibility caused by two coinfecting Cardinium symbionts. Heredity 128, 187–195 (2022). https://doi.org/10.1038/s41437-022-00507-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41437-022-00507-3
