
Abstract
Children with anxiety disorders (ADs) experience persistent fear and worries that are highly debilitating, conferring risk for lifelong psychopathology. Anticipatory anxiety is a core clinical feature of childhood ADs, often leading to avoidance of uncertain and novel situations. Extensive studies in non-human animals implicate amygdala dysfunction as a critical substrate for early life anxiety. To test specific amygdala-focused hypotheses in preadolescent children with ADs, we used fMRI to characterize amygdala activation during uncertain anticipation and in response to unexpected stimuli. Forty preadolescent (age 8–12 years) children, 20 unmedicated AD patients and 20 matched controls completed an anticipation task during an fMRI scan. In the task, symbolic cues preceded fear or neutral faces, such that ‘certain’ cues always predicted the presentation of fear or neutral faces, whereas ‘uncertain’ cues were equally likely to be followed by fear or neutral faces. Both AD children and controls showed robust amygdala response to faces. In response to the uncertain cues, AD children had increased amygdala activation relative to controls. Moreover, in the AD children, faces preceded by an ‘uncertain’ cue elicited increased amygdala activation, as compared with the same faces following a ‘certain’ cue. Children with ADs experience distress both in anticipation of and during novel and surprising events. Our findings suggest that increased amygdala activation may have an important role in the generation of uncertainty-related anxiety. These findings may guide the development of neuroscientifically informed treatments aimed at relieving the suffering and preventing the lifelong disability associated with pediatric ADs.
Similar content being viewed by others


INTRODUCTION
Anxiety disorders (ADs) are the most prevalent psychiatric ailments in children and adolescents, affecting up to 20% of youth (Costello et al, 2005). In addition to causing considerable distress and disability, extreme childhood anxiety is a significant risk factor for later psychopathology (Kessler et al, 2008; Pine et al, 1998). ADs are typically characterized by persistent and excessive worry, often accompanied by physiological and cognitive symptoms, which can lead to functional impairment (Connolly and Bernstein, 2007). As anticipation and uncertainty, in relation to potentially negative situations, commonly provoke anxiety, it has been hypothesized that AD patients have particular difficulty tolerating uncertain conditions (Grupe and Nitschke, 2013a; Krain et al, 2008; Krain et al, 2006). Despite the fact that most ADs begin in childhood, few studies have investigated the neural basis of uncertainty in relation to anxiety in preadolescent children with ADs. Understanding the pathophysiology of childhood ADs will provide a neuroscientific basis for novel treatment development targeting neural correlates of ADs manifesting early in life, with the potential to reduce the long-term suffering associated with the childhood onset of ADs.
Animal studies have defined the neural circuit that underlies adaptive anxiety and fear responses (Aggleton and Passingham, 1981; LeDoux, 2007). The amygdala is a core component of this circuit, and numerous imaging studies link alterations in amygdala function to various forms of stress-related psychopathology (eg, Etkin and Wager, 2007; Phan et al, 2013; Stein, 2009). Relevant to ADs, uncertain conditions engage the amygdala (Davis and Whalen, 2001; Rosen and Donley, 2006), which functions to direct attention to potentially negative stimuli (Whalen, 2007). Studies from our laboratory imaging large numbers of young anxious rhesus monkeys demonstrate trait-like increased metabolic activity in the amygdala (Oler et al, 2010). Other evidence from our primate studies, based on specific dorsal amygdala lesions, implicate a causal role for the amygdala in the early life presentation of extreme anxiety (Kalin et al, 2004). Because of the well-documented role of the amygdala in adaptive and maladaptive anxiety, as well as in processing uncertainty, it is a prime candidate for further study to understand the pathogenesis of childhood ADs.
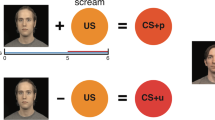
To assess whether preadolescent children with ADs have altered amygdala function, we performed fMRI to characterize the anticipation of, and response to, ethologically relevant fear stimuli. Fear faces are ideal stimuli, because they reliably elicit amygdala reactivity (Phillips et al, 1998; Whalen et al, 2001), and are highly relevant cues that are important to negotiating uncertain social situations. To test the hypothesis that preadolescent children with ADs have increased amygdala responses to uncertainty, we used a cued anticipation task (Figure 1) (Nitschke et al, 2009; Sarinopoulos et al, 2010). In the real world, an individual’s experience of negative events is influenced by their expectations and past experiences. The cued anticipation paradigm allowed us to explore alterations in the temporal dynamics of amygdala function during uncertain anticipation and the subsequent experience of unexpected events.

Anticipation task—negative and neutral cues (X, N) were always followed by fear or neutral faces, respectively. The uncertain cue (?) was followed half of the time by fear faces and half of the time by neutral faces.
MATERIALS AND METHODS
Participants
We studied 20 unmedicated, treatment-seeking, symptomatic children with ADs and 20 demographically matched control participants between the ages of 8–12 years (mean age 9.83 years). Children with anxiety were recruited from outpatient psychiatric clinics in the Madison area via clinician referral and through advertisements. Healthy control children were recruited via newspaper ads. Study procedures were approved by the University of Wisconsin–Madison Institutional Review Board, and informed consent was collected from all participants. All participants completed the Kiddie-Schedule for Affective Disorders and Schizophrenia Present and Lifetime versions (KSADS-PL) (Birmaher et al, 2009), administered by a PhD clinical psychologist trained in KSADS administration (GMR). Recordings of KSADs interviews were reviewed and checked for reliability by a psychiatrist (DSP), and diagnoses were confirmed with a second in-person interview with the patient and a parent by a second psychiatrist (NHK). Preadolescent anxiety often presents with an admixture of symptoms associated with generalized, separation, and social ADs (Kendall et al, 2010). As such, we enrolled children with ADs suffering from any of these three conditions, an approach similar to that used in large treatment studies of childhood ADs (Walkup et al, 2001; Compton et al, 2010) and in most prior imaging studies of ADs in children and adolescents (eg, Beesdo et al, 2009; Guyer et al, 2012). Likewise, these treatment and imaging studies have excluded patients with obsessive-compulsive disorder, post-traumatic stress disorder, autism spectrum disorders, bipolar disorder, schizophrenia, and/or IQ<80; we have done so similarly. We also collected parent and child ratings of anxiety symptoms using the Screen for Child Anxiety and Related Emotional Disorders (SCARED) (Birmaher et al, 1999), child reported depression using the Children’s Depression Inventory (Kovacs, 1985), and parent-rated externalizing behavior using the revised Conners’ Parents Rating Scale (Conners et al, 1998). Child IQ was evaluated using the Weschler Abbreviated Scale of Intelligence (WASI-II, 2011, Full Scale IQ-2), and pubertal stage was rated using the Tanner scale (Morris and Udry, 1980). Unrelated neuroimaging data from a subset of these participants (8 ADs, 6 controls) has been previously published (Birn et al, 2014). Children with ADs and controls did not differ in age, sex, Tanner stage, or IQ (all p’s>0.5), and the anxiety group scored higher on all clinical scales (all p’s<0.05). For demographics and diagnoses, see Table 1.
Study Visits and Mock Scan
Because of challenges related to collecting high-quality MRI data from pediatric participants, in particular children with anxiety, we were extremely deliberate in creating our study design, scanning parameters, and experimental procedures, to optimize the comfort of participants and maximize compliance with the protocol. The protocol included three study visits, to provide participants with a gradual introduction to the MRI procedures and environment. Study visit 1 involved the KSADs interview and a mock MRI scan session, which allowed children to acclimate to the scanner environment, receive feedback from study staff to minimize motion, and to practice with a modified version of the anticipation task (Figure 1) using happy faces. The second visit included the MRI scan, divided into two 25-min segments, which allowed children to take a break, to minimize the amount of time they were required to stay still. Task runs were kept as brief as possible (6 min) and were all collected in the first half of the scan session. Average time between study visit 1 and 2 was 14 days (range 1–25 days). At the third study visit we collected behavioral data, which are not reported here.
Anticipation Task
As shown in Figure 1, on each trial participants viewed an anticipatory cue presented for 2 s, followed by a blank screen to produce a jittered inter-stimulus interval (2–5 s), and a set of five faces presented for 1 s each. Negative cues (red ‘X’) were always followed by fear faces, neutral cues (blue ‘N’) were always followed by neutral faces, and uncertain cues (yellow ‘?’) were equally likely to be followed by fear or neutral faces. Children were told that the red X will be followed by scared faces, the blue N will be followed by neutral faces, and the question mark will be followed by either scared or neutral faces. Participants completed three 6-min runs, each of which included six negative cue–fear face, six neutral cue–Nnutral face, three uncertain cue–fear face, and three uncertain cue–nNeutral face trials. Faces were taken from two standardized sets of facial affect (Ekman and Friesen, 2006; Tottenham et al, 2009), and participants viewed both the fear and neutral expressions for each face. To promote attention to task trials, five blocks of a target detection task were presented throughout each run. In the target detection task, a cartoon figure appeared at different screen locations and participants pressed a button to indicate the position of the figure. For data to be included in final analyses, participants were required to detect one of the first two targets on at least 80% of target blocks (one run excluded for two control participants and one run excluded for one anxiety participant).
MRI/fMRI Parameters
MRI data were collected on a General Electric (GE) 3.0 T Discovery MR750 using a GE 8-channel head coil (GE, Milwaukee, WI). Anatomical scans were obtained with a 3D T1-weighted, inversion-recovery, fast gradient echo prescription: (TI/TR/TE/Flip/NEX/FOV/Matrix/Bandwidth: 450 ms/8.16 ms/3.18 ms/12°/1/256 mm/256 × 256/31.25 kHz) with whole-brain coverage (156 slices over 156 mm). Functional scans were obtained using a 2D T2*-weighted EPI prescription (TR/TE/Flip/FOV/Matrix: 2000 ms/25 ms/60°/240/64 × 64; 40 × 4.0-mm sagittal slices; gap: 0 mm; 180 volumes).
Analysis
Demographic and clinical variables were compared between groups using two-tailed independent samples t-tests χ2-tests.
For fMRI data, native-space T1 images were nonlinearly registered to the MNI probabilistic template (MNI152_T1_1mm_brain; http://fsl.fmrib.ox.ac.uk) using FLIRT (http://fsl.fmrib.ox.ac.uk/fsl/flirt) and FNIRT (http://www.fmrib.ox.ac.uk/fsl/fnirt). All fMRI data processing used standard methods in AFNI (Cox, 1996). Data processing included reconstruction using a Fermi filter, slice timing correction, six-parameter rigid-body volume registration, correction for field inhomogeneities, and removal of the skull and gross artifacts. Single-subject EPI data were spatially normalized, combining flirt alignment of EPI to anatomy matrices with the anatomical transformation matrices, and re-sampled to 2-mm isotropic voxels. To ensure high-quality EPI data, all data were visually inspected. EPI volumes with >2 mm of volume-to-volume motion were censored from data analyses. The percentage of included data was high and did not differ between children with ADs (M=97.8%, SD=3.7) and controls (M=96.5%, SD=5.0) (p=0.38).
Single-subject time series were analyzed using a general linear model with separate regressors for the anticipation and face periods, formed by convolving stimulus functions (2 s cue, 5 s face) with a canonical hemodynamic response function. A total of eight parameters were included in the model (three cue types, four face types, and one target detection task). Regressors were compared with an implicitly modeled baseline that included fixation periods between trials and at the beginning and end of each run (15 and 25 s, respectively). Resultant parameter estimates were converted to percentage signal change. The normalized percent signal change maps were spatially smoothed using a 4-mm FWHM Gaussian kernel. We performed voxel-wise t-tests between groups to examine hypothesized differences in percent signal change during the cue and face periods, including age and sex as covariates. To ensure that differences observed during the face period were not due to carry-over from the anticipation period, all face period analyses included percent signal change during the anticipation period as a voxelwise covariate.
We first performed a whole-brain analysis to identify face-responsive regions in preadolescent children (fear and neutral faces relative to baseline) (Figure 2). Because of our a priori hypotheses related to the amygdala and uncertainly, further analyses were restricted to face-responsive voxels within the amygdala (Harvard Oxford atlas (http://www.fmrib.ox.ac.uk/fsl/data/atlast-descriptions.html). Taking a similar approach using the whole, anatomical amygdala yields similar findings. We accounted for multiple comparisons using a small volume correction within this targeted region. Correction for multiple comparisons was applied at the cluster level following Monte Carlo simulations conducted in the 3dClustSim program within AFNI (http://afni.nimh.nih.gov/pub/dist/doc/program_help/3dClustSim.html) (Telzer et al, 2013). Results of the 3dClustSim indicated a voxel-wise threshold of p<0.025 combined with a minimum cluster size of 15 voxels for the bilateral amygdala corresponding to a cluster-corrected p<0.05 (similar to Tottenham et al, 2014). One sample t-tests were performed on extracted percent signal change from clusters that significantly differed between AD children and controls to further characterize the pattern of group differences. Exploratory whole-brain analyses for all contrasts were corrected with a voxel-wise threshold of p<0.005, within a sample-specific gray matter (GM) mask that required a minimum cluster size of 77 voxels for a whole-brain-corrected p<0.05. To create the GM mask, individual anatomical scans were segmented into GM, white matter, and cerebrospinal fluid probability maps using FAST (http://www.fmrib.ox.ac.uk/fsl/fast4). GM masks were averaged across all subjects and thresholded at 0.5 to include voxels labeled as GM in at least 50% of subjects.

Preadolescent children showed significant activation to faces in a network of brain regions including (a) the amygdala and (b) the orbital frontal cortex, hippocampus/paraphippocampal gyrus, fusiform gyrus, cerebellum, and the inferior occipital gyrus. Within the amygdala, there was no interaction with group, indicating that children with anxiety disorders (ADs) and control children did not differ in amygdala response to faces.
RESULTS
Amygdala
Amygdala response to neutral and fear faces in preadolescent children
As the neural response to emotional faces varies across development and few studies have examined neural responses to faces in preadolescent children (eg, Guyer et al, 2008; Thomas et al, 2001), we first characterized face-responsive regions in our entire 8- to 12-year-old sample. Among other regions, we found robust amygdala responses to faces (both fear and neutral) while controlling for the group (Figure 2, Supplementary Table S1; p<0.05, corrected). AD children and controls did not differ in their amygdala response to faces (p=0.98). Similar to studies of adults, we also found greater amygdala activation to fear relative to neutral faces (Supplementary Figure S1), as well as greater activation within regions of the fusiform gyrus and middle temporal gyrus (Supplementary Table S2; p<0.05, corrected). Amygdala activation to fear vs neutral faces did not differ between AD children and controls (p=0.39).
Increased amygdala activation in children with ADs in response to uncertainty
To test the hypothesis that children with ADs have increased amygdala activation in response to uncertainty, we compared AD children and controls for their difference in activation between the uncertain and negative cues within the face-responsive amygdala region. As reviewed above, the uncertain and negative cues are most likely to be experienced as aversive, and therefore this contrast isolates the effects of uncertainty while controlling for neural response associated with viewing a potentially aversive visual cue, as well as general effects of face anticipation. Results demonstrated that children with ADs had differential amygdalar responses to uncertainty (p<0.05, corrected). As shown in Figure 3, this effect was driven by differences in the uncertain cue condition, as groups did not differ in their response to negative cues (p=0.94). Within the uncertain cue condition, control children demonstrated significant deactivation (p<0.005) that was absent in the AD group (p=0.10). A control analysis comparing the uncertain cues with the neutral cues yields a similar pattern of results, further demonstrating the specific effect of uncertainty (Supplementary Figure S2; p<0.05, corrected).

During uncertain anticipation, relative to certain anticipation of fear faces, children with anxiety disorders (ADs) exhibited greater activation in the left amygdala than control children (32 voxels; peak x=−24, y=−4, z=−26). Graphs display percent signal change extracted from the cluster, and show that the group difference is driven by higher amygdala response to the Uncertain cue in children with ADs. Error bars represent SEM. Region of interest (face-responsive voxels within the amygdala) outlined in green.
Uncertain anticipation modulates amygdala response to faces
As individuals with ADs are particularly sensitive to uncertainty and potentially threatening conditions, we hypothesized that AD children would show increased amygdala reactivity to this combination of stimuli, specifically fear faces preceded by a cue that signals uncertainty. Therefore, within the face-responsive amygdala region, we compared AD with control children for the difference in activation between fear faces that were preceded by uncertain cues vs the same face stimuli preceded by certain, negative cues. To remove any possible influences of anticipation-related BOLD response on neural activation to the faces, analysis of face responses controlled for voxel-level activation during the cue period by including cue period activation maps as a covariate. Children with ADs exhibited greater amygdala activation than controls in response to the fear faces that were preceded by the uncertain cue (Figure 4; p<0.05, corrected). When viewing fear faces preceded by the uncertain cue, AD children had significant activation within the amygdala (p<0.005) that was absent in the control group (p=0.96). To examine the specificity of this effect, we performed the same analysis on neutral faces. Similar to the finding for fear faces relative to controls, AD children demonstrated enhanced amygdala reactivity to neutral faces that were preceded by an uncertain cue (Supplementary Figure S3).

When viewing fear faces preceded by the uncertain cue, children with anxiety exhibited greater activation in the right amygdala (20 voxels; peak voxel x=24, y=−2, z=−22). Graphs display percent signal change extracted from the cluster. Note that the group difference is driven by higher amygdala response to fear faces presented after the uncertain cue in children with anxiety disorders (ADs). Error bars represent SEM. Region of interest (face-responsive voxels within the amygdala) outlined in green.
Exploratory Whole-Brain Analyses
Exploratory whole-brain analyses within the anticipation task uncovered additional regions that differed between AD and control children during uncertain anticipation, including portions of the left insula, cerebellum, and the visual cortex (Supplementary Figure S4, Supplementary Table S3). Whole-brain analyses of responses to faces preceded by uncertain cues did not reveal any additional responses between group differences.
Correlational Analyses
To examine the relationship between uncertainty-related activation within regions that demonstrate a main effect of group and anxiety symptoms, we performed additional analyses. Within clusters that showed a main effect of group, there were no significant correlations between brain responses to uncertainty and anxiety severity as determined by Parent and Child SCARED-total scores (tested within anxiety group only and across full sample with group as a nuisance covariate) (all t’s <0.93, all p’s>0.36). Similarly, within the same clusters, participant age was not significantly correlated with response to uncertainty across groups (all t’s<1.26, all p’s<0.21), nor was there any significant group by age interactions.
DISCUSSION
Recent animal and human studies highlight the importance of neurodevelopmental brain changes that likely influence the amygdala response to threat (eg, Gabard-Durnam et al, 2014; Giedd, 2004; Qin et al, 2012). Developmental brain-imaging studies demonstrate that amygdala response to fear faces varies across childhood and adolescence (eg, Guyer et al, 2008; Hare et al, 2008; Thomas et al, 2001). Although most studies of adults find increased amygdala reactivity to fear compared with neutral faces (for review, see Fusar-Poli et al, 2009), data from studies of children are mixed, with some finding similar valence effects as those in adult studies (eg, Guyer et al, 2008), while others, particularly in younger children, do not (Pagliaccio et al, 2013). Our study with preadolescent children found adult-like increased amygdala reactivity to fear compared with neutral faces (Supplementary Figure S1). These findings complement previous work examining the developmental trajectory of amygdala function, and provide the foundation for our examination of aberrant amygdala activation in childhood ADs. In contrast to some previous studies that have found increased amygdala activation to negative stimuli in AD patients relative to control participants (eg, Beesdo et al, 2009; Hattingh et al, 2012; McClure et al, 2007), we did not find a group by valance interaction during face viewing within the amygdala. Further studies are needed to determine whether this is a true developmental difference observed in our preadolescent sample, or whether our findings are better explained by variations in diagnosis, task, and/or analysis approaches from previous studies that demonstrate amygdala hyperactivity to threat in patients with ADs.
In the present study, 8- to 12-year-old children with ADs had differential amygdala activation during uncertain anticipation. This finding suggests that the increased emotional reactivity experienced by children with ADs during uncertain or ambiguous situations could result from altered amygdala function. Although not the primary focus of this study, our exploratory whole-brain analysis also found differential uncertainty-related insula activation in AD children, a finding consistent with studies of uncertainty processing in adults and adolescents (Dunsmoor et al, 2007; Krain et al, 2006). The differential amygdala and insula reactivity during uncertain anticipation was driven by a significant deactivation in the control group that was absent in AD children (Figure 3). Similar insula deactivation has been observed in healthy adults during anticipation of negative and neutral images, and it has been suggested that this pattern is consistent with active regulation of activity in these regions (Grupe et al, 2013b). Thus, the lack of deactivation in the AD group may represent the failure of normative regulation of the amygdala and insula during uncertain anticipation.
Although unexpected events often follow a period of uncertainty, our experimental design and analytic strategy allowed us to disambiguate neural activation related to uncertain anticipation from that associated with the presentation of unexpectedly valenced faces. We found that when AD children could predict whether the faces they viewed were fearful or neutral, their amygdala response did not differ from controls. In contrast, these same faces elicited increased amygdala activation in AD children when the valence of the faces could not be predicted. This finding is consistent with the increased reactivity of AD children when confronted with surprising or unexpected events. These findings suggest that increased amygdala activation in part underlies the heightened sensitivity of children with ADs to these commonly associated, but separable, anxiety-provoking circumstances.
Although anticipatory anxiety and worry are core features of ADs, it is noteworthy that very few studies have examined the neural underpinnings of uncertainty, as it relates to psychopathology. Consistent with our findings in preadolescent AD children, studies that have examined uncertainty processing in adolescents (Krain et al, 2008) and adults (Yassa et al, 2012) with ADs also find altered amygdala function. A common coping strategy for AD children is to attempt to anticipate and control all potential outcomes related to future events. In part, this may be in response to the intense distress that AD children experience when uncertain about the future. Importantly, we investigated not only neural activation in response to uncertain conditions, but also how uncertainty affects brain responses to subsequent stimuli. These data have important implications for understanding why children with ADs are driven to seek control of their environment and are compelled to avoid new situations. The differential amygdala activation observed in this study may help to explain why AD children have such difficulty coping and why they experience negative affect both before and during novel and surprising events. It will be important in the future to perform studies with larger sample sizes that will allow for the detection of possible relationships between uncertainty processing, specific anxiety symptoms, and/or diagnostic categories. We also note that the mean IQ of participants in the current study was in the high average range, which could limit the generalizability of our findings.
Previous studies in adults suggest that uncertainty enhances behavioral, physiological, and neural responses to aversion (Dunsmoor et al, 2007; Sarinopoulos et al, 2010). It is important to emphasize that in children with ADs, heightened anticipatory anxiety can result in pronounced disability, eg, school avoidance, social isolation, and erosion of self-esteem. Repeated requests for reassurance, inflexible behaviors, and emotional outbursts are consequences that parents of children with ADs must negotiate. Our data highlight a potential role for the amygdala in the generation of anticipatory anxiety and its consequences. Future studies focused on the early onset and pathophysiology of childhood ADs would benefit from understanding the neurodevelopmental antecedents that mediate the development of heightened uncertainty-related amygdala responsivity. An important complement to further studies in children will be translational studies in young animals aimed at understanding mechanisms mediating the influences of uncertainty early in life. Together, these studies will help change treatment approaches for children suffering from early life anxiety.
FUNDING AND DISCLOSURE
Dr Kalin has served on the scientific advisory board of Corcept Therapeutics. Dr Kalin has stock options in Corcept Therapeutics and has received honoraria from CME Outfitters and Elsevier. All other authors declare no potential conflict of interest.
References
Aggleton JP, Passingham RE (1981). Syndrome produced by lesions of the amygdala in monkeys (Macaca mulatta). J Comp Physiol Psychol 95: 961–977.
Beesdo K, Lau JY, Guyer AE, McClure-Tone EB, Monk CS, Nelson EE et al (2009). Common and distinct amygdala-function perturbations in depressed vs anxious adolescents. Arch Gen Psychiatry 66: 275–285.
Birmaher B, Brent DA, Chiappetta L, Bridge J, Monga S, Baugher M (1999). Psychometric properties of the Screen for Child Anxiety Related Emotional Disorders (SCARED): a replication study. J Am Acad Child Adolesc Psychiatry 38: 1230–1236.
Birmaher B, Ehmann M, Axelson DA, Goldstein BI, Monk K, Kalas C et al (2009). Schedule for affective disorders and schizophrenia for school-age children (K-SADS-PL) for the assessment of preschool children—a preliminary psychometric study. J Psychiatr Res 43: 680–686.
Birn RM, Shackman AJ, Oler JA, Williams LE, McFarlin DR, Rogers GM et al (2014). Evolutionarily conserved prefrontal-amygdalar dysfunction in early-life anxiety. Mol Psychiatry 19: 915–922.
Compton SN, Walkup JT, Albano AM, Piacentini JC, Birmaher B, Sherrill JT et al (2010). Child/Adolescent Anxiety Multimodal Study (CAMS): rationale, design, and methods. Child Adolesc Psychiatry Ment Health 4: 1.
Conners CK, Sitarenios G, Parker JD, Epstein JN (1998). The revised Conners' Parent Rating Scale (CPRS-R): factor structure, reliability, and criterion validity. J Abnorm Child Psychol 26: 257–268.
Connolly SD, Bernstein GA (2007). Practice parameter for the assessment and treatment of children and adolescents with anxiety disorders. J Am Acad Child Adolesc Psychiatry 46: 267–283.
Costello EJ, Egger HL, Angold A (2005). The developmental epidemiology of anxiety disorders: phenomenology, prevalence, and comorbidity. Child Adolesc Psychiatr Clin N Am 14: 631–648 vii.
Cox RW (1996). AFNI: software for analysis and visualization of functional magnetic resonance neuroimages. Comput Biomed Res 29: 162–173.
Davis M, Whalen PJ (2001). The amygdala: vigilance and emotion. Mol Psychiatry 6: 13–34.
Dunsmoor JE, Bandettini PA, Knight DC (2007). Impact of continuous versus intermittent CS-UCS pairing on human brain activation during Pavlovian fear conditioning. Behav Neurosci 121: 635–642.
Ekman P, Friesen W (2006) Pictures of Facial Affect. Consulting Psychologists Press: Palo Alto.
Etkin A, Wager TD (2007). Functional neuroimaging of anxiety: a meta-analysis of emotional processing in PTSD, social anxiety disorder, and specific phobia. Am J Psychiatry 164: 1476–1488.
Fusar-Poli P, Placentino A, Carletti F, Landi P, Allen P, Surguladze S et al (2009). Functional atlas of emotional faces processing: a voxel-based meta-analysis of 105 functional magnetic resonance imaging studies. J Psychiatry Neurosc 34: 418–432.
Gabard-Durnam LJ, Flannery J, Goff B, Gee DG, Humphreys KL, Telzer E et al (2014). The development of human amygdala functional connectivity at rest from 4 to 23years: a cross-sectional study. NeuroImage 95C: 193–207.
Giedd JN (2004). Structural magnetic resonance imaging of the adolescent brain. Annals NY Acad Sci 1021: 77–85.
Grupe DW, Nitschke JB (2013a). Uncertainty and anticipation in anxiety: an integrated neurobiological and psychological perspective. Nat Rev Neurosci 14: 488–501.
Grupe DW, Oathes DJ, Nitschke JB (2013b). Dissecting the anticipation of aversion reveals dissociable neural networks. Cerebral Cortex 23: 1874–1883.
Guyer AE, Choate VR, Detloff A, Benson B, Nelson EE, Perez-Edgar K et al (2012). Striatal functional alteration during incentive anticipation in pediatric anxiety disorders. Am J Psychiatry 169: 205–212.
Guyer AE, Monk CS, McClure-Tone EB, Nelson EE, Roberson-Nay R, Adler AD et al (2008). A developmental examination of amygdala response to facial expressions. J Cogn Neurosci 20: 1565–1582.
Hare TA, Tottenham N, Galvan A, Voss HU, Glover GH, Casey BJ (2008). Biological substrates of emotional reactivity and regulation in adolescence during an emotional go-nogo task. Biol Psychiatry 63: 927–934.
Hattingh CJ, Ipser J, Tromp SA, Syal S, Lochner C, Brooks SJ et al (2012). Functional magnetic resonance imaging during emotion recognition in social anxiety disorder: an activation likelihood meta-analysis. Front Hum Neurosci 6: 347.
Kalin NH, Shelton SE, Davidson RJ (2004). The role of the central nucleus of the amygdala in mediating fear and anxiety in the primate. J Neurosci 24: 5506–5515.
Kendall PC, Compton SN, Walkup JT, Birmaher B, Albano AM, Sherrill J et al (2010). Clinical characteristics of anxiety disordered youth. J Anxiety Disord 24: 360–365.
Kessler RC, Gruber M, Hettema JM, Hwang I, Sampson N, Yonkers KA (2008). Co-morbid major depression and generalized anxiety disorders in the National Comorbidity Survey follow-up. Psychol Med 38: 365–374.
Kovacs M (1985). The Children’s Depression Inventory (CDI). Psychopharmacol Bull 21: 995–998.
Krain AL, Gotimer K, Hefton S, Ernst M, Castellanos FX, Pine DS et al (2008). A functional magnetic resonance imaging investigation of uncertainty in adolescents with anxiety disorders. Biol Psychiatry 63: 563–568.
Krain AL, Hefton S, Pine DS, Ernst M, Castellanos FX, Klein RG et al (2006). An fMRI examination of developmental differences in the neural correlates of uncertainty and decision-making. J Child Psychol Psychiatry 47: 1023–1030.
LeDoux J (2007). The amygdala. Curr Biol 17: R868–R874.
McClure EB, Monk CS, Nelson EE, Parrish JM, Adler A, Blair RJ et al (2007). Abnormal attention modulation of fear circuit function in pediatric generalized anxiety disorder. Arch Gen Psychiatry 64: 97–106.
Morris NM, Udry JR (1980). Validation of a self-administered instrument to assess stage of adolescent development. J Youth Adolesc 9: 271–280.
Nitschke JB, Sarinopoulos I, Oathes DJ, Johnstone T, Whalen PJ, Davidson RJ et al (2009). Anticipatory activation in the amygdala and anterior cingulate in generalized anxiety disorder and prediction of treatment response. Am J Psychiatry 166: 302–310.
Oler JA, Fox AS, Shelton SE, Rogers J, Dyer TD, Davidson RJ et al (2010). Amygdalar and hippocampal substrates of anxious temperament differ in their heritability. Nature 466: 864–868.
Pagliaccio D, Luby JL, Gaffrey MS, Belden AC, Botteron KN, Harms MP et al (2013). Functional brain activation to emotional and nonemotional faces in healthy children: evidence for developmentally undifferentiated amygdala function during the school-age period. Cogn Affect Behav Neurosci 13: 771–789.
Phan KL, Coccaro EF, Angstadt M, Kreger KJ, Mayberg HS, Liberzon I et al (2013). Corticolimbic brain reactivity to social signals of threat before and after sertraline treatment in generalized social phobia. Biol Psychiatry 73: 329–336.
Phillips ML, Young AW, Scott SK, Calder AJ, Andrew C, Giampietro V et al (1998). Neural responses to facial and vocal expressions of fear and disgust. Proc Biol Sci 265: 1809–1817.
Pine DS, Cohen P, Gurley D, Brook J, Ma Y (1998). The risk for early-adulthood anxiety and depressive disorders in adolescents with anxiety and depressive disorders. Arch Gen Psychiatry 55: 56–64.
Qin S, Young CB, Supekar K, Uddin LQ, Menon V (2012). Immature integration and segregation of emotion-related brain circuitry in young children. Proc Natl Acad Sci USA 109: 7941–7946.
Rosen JB, Donley MP (2006). Animal studies of amygdala function in fear and uncertainty: relevance to human research. Biol Psychol 73: 49–60.
Sarinopoulos I, Grupe DW, Mackiewicz KL, Herrington JD, Lor M, Steege EE et al (2010). Uncertainty during anticipation modulates neural responses to aversion in human insula and amygdala. Cerebral Cortex 20: 929–940.
Stein MB (2009). Neurobiology of generalized anxiety disorder. J Clin Psychiatry 70 Suppl 2: 15–19.
Telzer EH, Flannery J, Shapiro M, Humphreys KL, Goff B, Gabard-Durman L et al (2013). Early experience shapes amygdala sensitivity to race: an international adoption design. J Neurosci 33: 13484–13488.
Thomas KM, Drevets WC, Whalen PJ, Eccard CH, Dahl RE, Ryan ND et al (2001). Amygdala response to facial expressions in children and adults. Biol Psychiatry 49: 309–316.
Tottenham N, Hertzig ME, Gillespie-Lynch K, Gilhooly T, Millner AJ, Casey BJ (2014). Elevated amygdala response to faces and gaze aversion in autism spectrum disorder. Soc Cogn Affect Neurosci 9: 106–117.
Tottenham N, Tanaka JW, Leon AC, McCarry T, Nurse M, Hare TA et al (2009). The NimStim set of facial expressions: judgments from untrained research participants. Psychiatry Res 168: 242–249.
Walkup JT, Labellarte MJ, Riddle MA, Pine DS, Greenhill L, Klein R et al, for the Research Unit on Pediatric Psychopharmacology Anxiety Study Group (2001). Fluvoxamine for the treatment of anxiety disorders in children and adolescents. N Engl J Med 344: 1279–1285.
Whalen PJ (2007). The uncertainty of it all. Trends Cogn Sci 11: 499–500.
Whalen PJ, Shin LM, McInerney SC, Fischer H, Wright CI, Rauch SL (2001). A functional MRI study of human amygdala responses to facial expressions of fear versus anger. Emotion 1: 70–83.
Yassa MA, Hazlett RL, Stark CE, Hoehn-Saric R (2012). Functional MRI of the amygdala and bed nucleus of the stria terminalis during conditions of uncertainty in generalized anxiety disorder. J Psychiatr Res 46: 1045–1052.
Acknowledgements
We acknowledge the assistance of Anna White and Travis Doran, and the staffs of the HealthEmotions Research Institute (HERI) and Lane Neuroimaging Laboratory. This work was supported by the National Institutes of Health grant R21 MH092581 (NHK and JAO).
Author information
Authors and Affiliations
Corresponding author
Additional information
Supplementary Information accompanies the paper on the Neuropsychopharmacology website
Supplementary information
Rights and permissions
About this article

Cite this article
Williams, L., Oler, J., Fox, A. et al. Fear of the Unknown: Uncertain Anticipation Reveals Amygdala Alterations in Childhood Anxiety Disorders. Neuropsychopharmacol 40, 1428–1435 (2015). https://doi.org/10.1038/npp.2014.328
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/npp.2014.328
This article is cited by
-
A dynamic relation between whole-brain white matter microstructural integrity and anxiety symptoms in preadolescent females with pathological anxiety
Translational Psychiatry (2022)
-
Acute neurofunctional effects of escitalopram during emotional processing in pediatric anxiety: a double-blind, placebo-controlled trial
Neuropsychopharmacology (2022)
-
Young patients’ self-reported fear compared to professionals’ assessments during invasive and non-invasive dental visits: a prospective, longitudinal study
European Archives of Paediatric Dentistry (2022)
-
Psychophysiological mechanisms underlying the failure to speak: a comparison between children with selective mutism and social anxiety disorder on autonomic arousal
Child and Adolescent Psychiatry and Mental Health (2021)
-
The electrophysiology correlation of the cognitive bias in anxiety under uncertainty
Scientific Reports (2020)
