
Abstract
The US swine industry has been impaired over the last 25 years by the far-reaching financial losses caused by the porcine reproductive and respiratory syndrome (PRRS). Here, we explored the relations between the spatial risk of PRRS outbreaks and its phylodynamic history in the U.S during 1998–2016 using ORF5 sequences collected from swine farms in the Midwest region. We used maximum entropy and Bayesian phylodynamic models to generate risk maps for PRRS outbreaks and reconstructed the evolutionary history of three selected phylogenetic clades (A, B and C). High-risk areas for PRRS were best-predicted by pig density and climate seasonality and included Minnesota, Iowa and South Dakota. Phylodynamic models demonstrated that the geographical spread of the three clades followed a heterogeneous spatial diffusion process. Furthermore, PRRS viruses were characterized by typical seasonality in their population size. However, endemic strains were characterized by a substantially slower population growth and evolutionary rates, as well as smaller spatial dispersal rates when compared to emerging strains. We demonstrated the prospects of combining inferences derived from two unique analytical methods to inform decisions related to risk-based interventions of an important pathogen affecting one of the largest food animal industries in the world.
Similar content being viewed by others

Introduction
The United States (U.S.) is the third largest pork-producer country and the second major exporter globally1. Porcine reproductive and respiratory syndrome (PRRS) first appeared in the southeastern region of the U.S. in 1989, causing reproductive failure in sows and respiratory disorders in pigs of all ages2. Subsequently, the disease spread throughout the North American swine population and became endemic. Although the disease was first discovered in the U.S., earliest serological evidence suggests that the virus first emerged in eastern Canada3. In 2011, the annual losses associated with PRRS virus (PRRSv) infections on the national economy of the U.S. reached up to USD 664 million4. Thus, arguably, the disease has become one of the most import swine diseases and has been considered a national threat to food security5. PRRSv is an enveloped single-stranded RNA virus and belongs to the Arteriviridae family3. The virus has seven structural and 14 non-structural proteins encoded by a genome that comprises nine open reading frames (ORF)6. Envelope surface glycoprotein (GP5) is encoded by gene segment ORF5, which is characterized by the highest genetic diversity compared to other ORF segments, and thus, became popular for most molecular epidemiology studies7,8,9.
PRRSV infections have been reported worldwide, except for Australia and Antarctica, and have been classified into two main groups; North American and European strains10, 11. The European strain is referred to as Type I genotype and has relatively low prevalence in the U.S. The North American strain is referred to as Type II genotype and is currently causing most PRRSv infections in the country11. PRRSv can be transmitted rapidly through direct and indirect contact12, which include between-farm transmission through animal movement (direct contact between infected and susceptible animals), fomites and airborne transmission13, 14. Endemic PRRSv strains are commonly characterized by annual seasonal increases in the number of observed outbreaks, with incidence of cases being low during spring and summer and high during fall and winter15. However, the emergence of new virulent viruses is also a common characteristic of PRRSv (referred to as emerging strains), with “spreader events” being recognized for different United States regions16; those emerging strains are of considerable concern to the swine industry as they cause severe economic losses. It is currently unknown if alternative transmission routes are equally important for emerging and endemic PRRSv strains.
Currently, PRRSv control and prevention activities are unregulated in the U.S., which, combined with the varied and rapid biological properties of this RNA virus and the limited effectiveness of available vaccines, jeopardize the effectiveness of those mitigating measures17. Hence, risk-based interventions against emerging strains are required to minimize their impact on the industry5. Current swine production systems are commonly not separated by well-defined geographical boundaries and are characterized by highly frequent, often long-distance movement of pigs and supplies. Indeed, geographical distribution and dynamics of PRRSv spread are influenced by system-level decisions and needs, as well as environmental risk factors including, mainly, pig density and climate18,19,20.
Species distribution models (SDM) are spatially explicit analytical methods that offer the possibility to build predictive risk maps using disease presence and environmental data21. SDM are capable of extracting associations between the presence of disease cases (or outbreaks) and environmental factors to characterize environmental requirements for pathogen occurrence, which subsequently provide predictions on suitable geographical locations for virus circulation and spread over non-sampled areas21. SDMs have recently become popular for risk mapping of both animal and human diseases22,23,24,25 at local, regional and global scales. Such methods have been proven useful for modeling spatial distribution of diseases over large geographical areas, and thus, can provide a supportive platform for targeted sampling schemes of PRRSv within and between swine production regions in the U.S.
Molecular characterization of PRRSv constitutes a substantial portion of the pathogen’s surveillance efforts in the U.S., where a large number of sequences became available due to the growing accessibility to affordable molecular tests26. However, the increase in the size of PRRSv’s genomic data led to new challenges regarding the interpretation of results, especially considering the rapid mutation and recombination events of the virus. One of those challenges relates to the need for differentiating endemic from emerging strains. Past PRRSv evolutionary epidemiology studies either focused on establishing associations between phylogenetics and outbreak characteristics in different geographical levels7, 27,28,29, or discriminating between endemic and emerging strains to infer about their spread and maintenance within affected swine populations7, 10, 30,31,32. Those studies relied on traditional phylogenetic methods to either genotype new viruses using the restriction fragment length polymorphism (RFLP) patterns, or assess correlations between the similarities of nucleotide sequences and spatio-temporal outbreak dynamics33. Such methods ignore important evolutionary parameters, uncertainties associated with phylogenetic relationships, and spatio-temporal factors that shape the evolutionary history of rapidly evolving pathogens like PRRSv34, 35. Thus, tools provided by the field of phylodynamics have become a necessity to effectively characterize the joint evolutionary and epidemiological patterns of rapidly evolving pathogens36. These methods heavily rely on Bayesian statistical frameworks, which can provide methods that are able to account for uncertainties in the evolutionary parameters of the phylogeny, and subsequently can provide estimates on population dynamics, divergence times, and history of geographic spread37.
In the past decade, a few studies attempted to reconstruct the evolutionary history of PRRSv using Bayesian phylodynamic models14, 33, 38,39,40. Such studies answered important a long-standing hypothesis on the evolutionary epidemiology of PRRSv, and subsequently encouraged the notion for such methods to be routinely applied in surveillance of field data with the ultimate goal of supporting risk-based interventions. However, following the continuous rapid growth in both size and complexity of PRRSv data, routine implementation of such analytical methods is quite challenging. To date, no analytical pipelines were formulated to rigorously distinguish between endemic and emerging PRRSv strains. Instead, the distinction between such strains is naively made, for example, through the incidence of outbreaks and traditional phylogenetic methods. The application of more robust analytical pipelines that specializes in characterizing spatiotemporal dynamics of PRRSv strains can be of great value for the swine industry by improving surveillance, and subsequently control and prevention measures.
In this study, we illustrate the prospect of combining inferences derived from modern spatial explicit and phylodynamic disciplines for routine surveillance of PRRSv, and, potentially, other food animal pathogens, with the goal of supporting risk-based interventions in near-real time. Specifically, the objectives of this study were to identify environmental requirements for the circulation and spread of endemic PRRSv strains, and to characterize the evolutionary features of endemic and emerging PRRSv strains in a complex geographical setting. Results here will ultimately help support prevention and control of the most devastating disease affecting one of the largest food animal industries worldwide.
Results
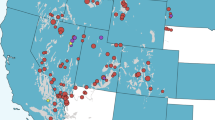
All PRRSv ORF5 sequences (n = 3,582) were field isolates obtained from different type of farms from one swine production system, including farrow to wean, farrow to feeder, and growing pig farms. The complete ORF5 nucleotide sequences were collected between 1st January 1998 and 21st April 2016 from PRRSv-infected swine farms located in the Midwest region of the U.S. Here, we defined the Midwest region to include Minnesota, Iowa, South Dakota and Nebraska. Sequencing was performed in Midwest-based veterinary diagnostic laboratories or private laboratories on a fee-for-service basis and according to the procedures in use at the time of virus detection. The information contained in the dataset obtained from the system included RFLP genotype classification, geographical location (latitude/longitude) of the farm from which the virus was isolated, and date of sequencing. The data were shared under a strict confidentiality agreement, in which identity of the production system and location of the participant farms cannot be revealed. Therefore, we generated and plotted a kernel density function of 5 km2 spatial resolution to represent the locations of infected farms (Fig. 1A) using ArcGIS version 10.441. Sequence dataset used here is currently under preparation to be deposited in the Genbank and will be available soon to the readers with accessions xx-yy. Furthermore, full alignment of sequences has been provided on a FASTA file with the supplementary materials (File S1).

Geographical locations of the PRRSv sequences and predicted spatial probability of PRRSV outbreaks in swine farms located in the Midwest region of the U.S., collected from January 1998 to April 2016. (A) Smoothed kernel density function for the geographical locations of the farms where sequences were collected and geographical exntension of the selected clades. (B) Spatial probability distribution of PRRSV outbreaks predicted by the final presence-only maximum entropy ecological niche model. Figure’s maps were generated using ArcGIS version 10.4.
We used presence-only maximum entropy, an SDM technique (Maxent)42 to predict the geographical range of PRRSv high-risk areas in the Midwest region. Geographical locations of all sequences and a set of selected environmental predictors (Table S1) were used for the subsequent Maxent analyses. Furthermore, we used Bayesian Phylodynamic methods implemented in the BEAST43 software package to model the demographic and phylogeographic history of 3 selected viral clades, from the sequences described above. We decided to select three clades as cases studies, because analyzing 3,582 sequences at one run using complex and advanced phylodynamic methods is computationally demanding and time consuming, which was neither feasible for us nor the swine producers. The selected three clades were genetically distinct, in which two of them were dominated by strains known as endemic (based on the RFLP classification), whereas the third clade compromised emerging strains. Resulting risk maps and posterior parameters of the spatiotemporal diffusion history, from the Maxent and phylodynamic analyses, respectively, of the selected clades were combined visually to qualitatively infer some epidemiological insights about the characteristics of the selected PRRSv clades in the Midwest.
The set of selected environmental variables (Table S1) used in the Maxent analyses, included: (1) pig density; (2) climate; and (3) land cover. The density of pig farms in the region (referred to as pig density) was represented using a kernel density function with a spatial resolution of 5 km2 and was derived from locations of all pig farms across the US in 2012, retrieved from the United States Department of Agriculture Census of Agriculture website44. We obtained climate data (referred to as Bioclime) from the WorldClim in the form of 5 km2 resolution rasters45. We used 15 bioclimatic variables, out of the 19, and excluded the rest due to known spatial artifact in those four variables, as suggested elsewhere46. Finally, we retrieved MODIS-based global land cover climatology from the United States Geographical Survey (USGS) webpage, with a spatial resolution of 0.5 km2, to provide an estimate of the geographical distribution of 16 different land cover features in the US47. We processed the 17 environmental variables using the ‘Raster’ package48 implemented in R statistical software version 3.2.249. All environmental layers were converted into a common projection, spatial resolution, and map extent, and each raster was cropped so that the geographical extent of the spatial analyses covered the Midwest swine production region. Raster data were aggregated and resampled to create a uniform grid size, which resulted in a scale of approximately 5 km2. Finally, we visually inspected collinearity between each pair of environmental predictors using scatter-plots.
Species distribution modeling (SDM) of PRRSv spatial risk in the Midwest
Only four environmental predictors contributed to the geographical range of high-risk areas for PRRSv outbreaks in the Midwest, with AUC values greater than 0.7 (Table 1). Pig density was, by far, the most important environmental predictor, followed by temperature seasonality, precipitation seasonality, and land cover in the Midwest (Table 1). Areas with high pig densities, temperature changes around 11 to 13 °C, standard deviation and precipitation variability between 40% and 60% over the course of the year, and located nearby permanent wetlands were found mostly suitable for PRRSv outbreaks in the Midwest. Most PRRSv outbreaks (>25%) were observed in southwestern Minnesota and eastern South Dakota (Fig. 1A). However, the predicted spatial range of high-risk areas (Probability >0.8) in the Midwest included southern and western Minnesota, northwestern Iowa, and eastern and central of South Dakota as well (Fig. 1B).
Demographic and Phylogeographic history of PRRSv in the Midwest
Results of the ORF5 Maximum likelihood (ML) tree yielded over 20 distinct PRRSV clades (Fig. 2). We selected three clades (A, B and C) for further analyses, ensuring that each selected clade’s spatial and temporal distribution represented the geographical extent and period of the study (Figs 1A and 2). Clades A and B ORF5 sequence data favored the expansion and the exponential coalescent tree models, respectively (BF > 20; Table S2). That finding suggests that Clade A viral population grew at an increasingly exponential rate, whereas Clade B population grew at a fixed exponential rate. However, while both clades favored different coalescent tree models, inferred posterior growth rate parameter was similar and equal to 0.23 year−1 (95% HPD:0.07, 0.38) and 0.21 year−1 (95% HPD:0.08, 0.36) for clades A and B, respectively. Clade C ORF5 sequence data favored the logistic coalescent tree model (BF > 34; Table S2), which assumes that population size of the virus grew at a decreasing rate over time (Table S2). However, Clade’s C inferred posterior growth rate parameter was substantially larger than that of Clades’ A and B and was equal to 2.63 year−1 (95% HPD:1.28, 4.20). Furthermore, estimates of the clades’ relative genetic diversity through time suggest that all the three clades exhibited typical annual seasonal increases in population size (Fig. 3).

Maximum Likelihood (ML) phylogeny of PRRSV ORF-5 sequences collected between January 1998 and April 2016 in swine farms in the Midwest of the U.S. The ML tree was inferred from the GTR + Γ model of evolution. Support given at nodes based on through bootstrap search using 10 runs, with 100 ML replicates in each run, implemented in RAxML version 8. The scale bar indicates the number of substitutions per site. Selected clades are highlighted with green (Clade ‘A’), purple (Clade ‘B’), and Blue (‘F’), with their corresponding RFLP-types isolates listed and collection dates (Month-Year).

Temporal variation in the effective population size of PRRSv selected clades in the in the Midwest isolated between 2009 and 2016. This ‘Bayesian skygrid’ plot summarizes the inferred effective population size (N e T)—which summarize genetic variation in terms of effective population size trajectories of the sampled ORF-5 gene sequences—through time in the Midwest. The mean estimate is indicated by the dark green line; the shaded light green regions correspond to the 95% HPD. Red bars represents the number of sequences collected over the period of the study and correspond to each selected clade. Vertical doted lines represent corresponds to the estimated time at which each clade transitioned from slow to fast population growth. Each figure (A–C) corresponds to Clades (A–C), respectively.
Continuous diffusion phylogeographic models indicated that geographical spread of PRRSv outbreaks for the three clades followed a heterogeneous, rather than homogenous, spatial diffusion process (Table 2). Both A and B clades favored the gamma RRW model with initial dispersal rates of 16.3 and 13.1 km/year, respectively (Table 2). However, clade C favored the Cauchy RRW model with a dispersal rate that was substantially higher and reached up to 73.1 km/year (Table 2). Mean evolutionary rates of clades A and B were closely similar (0.0074 and 0.0089 substitution/site/year, respectively) and substantially smaller than clade C (0.0114; Table 2). However, viruses from clade C have evolved more recently than those from clades A and B (Table 2; Fig. 4). Tree topologies inferred by the ML method (Figure S1) for the three clades were substantially different from the topologies inferred by the Bayesian diffusion phylogeographic models (Fig. 4). For all the three clades, highest among-branch spatial diffusion rate variation was inferred at most of the tips of the MCC trees, which is a typical reflection of the heterogeneous diffusion process (Fig. 4).

Maximum clade credibility (MCC) phylogeny of ORF-5 gene of PRRS clades ‘A’, ‘B’ and ‘C’ viruses in the Midwest estimated under the best-fitting spatial diffusion model (Table 2). The color of the branches represents the among-branch spatial diffusion rate variation and corresponds to the color gradient legend on the middle right. Well-supported posterior probabilities (P > 0.85) of branching events are indicated by black circles at the nodes.
Finally, the inferred geographical origins of the posterior phylogeny and dispersal routes with a high rate of spatial diffusion for clades A and B were mostly encompassed by the identified high-risk areas predicted by the final Maxent model (Fig. 5). Indeed, most of the MCC tree branches of clades A and B had accumulated at the predicted high-risk areas (Probability <0.8), with narrow dispersal patterns (Fig. 5). However, even though clade C’s point of geographical origin was encompassed by the predicted high-risk areas in the borders between Minnesota and Iowa (Fig. 5), the MCC branches exhibited a substantially wider dispersal pattern compared to clades A and B, and did not accumulate in the predicted high-risk areas (Fig. 5). Instead, the MCC branches of clade C predominately spread throughout the study region as demonstrated in Fig. 5.

Clades’ ‘A’, ‘B’ and ‘C’ MCC tree spread and ecology in the Midwest. The Google Earth Pro snapshots of Clades MCC trees are superimposed on the predicted spatial risk of PRRSV outbreaks by the final Maxent model. The gradient color legend on the upper right represents the scale of the spatial probability distribution. The lines represent the continuous diffusion phylogeographic history of clade viruses, and their gradient colors represent their age (light pink = older, dark pink = younger). The figure is composed of satellite images captured as snapshots from Google Earth Pro (https://www.google.com/earth/). Figure’s associated three-dimensional movies corresponding to each clade are provided in the supplementary materials (File S2–4).
Discussion
This study represents the first attempt to explore the potential of combining novel analytical methods including maximum entropy and Bayesian phylodynamic modeling to provide insights into the spatial and evolutionary epidemiology of viral diseases, using PRRSv in the U.S. a working example. We estimated, in quantitative and qualitative terms, the evolutionary history and extent of the association between environmental and demographic factors and geographical distribution of the PRRSv in endemic settings. We demonstrated that pig density and climate seasonality were not only important factors for maintaining endemic PRRSv strains, but also, they likely shaped genetic diversity over time as well as spatiotemporal diffusion patterns. Simultaneously, we revealed the wide and rapid spatiotemporal diffusion characteristics in the evolution of emerging strains. We demonstrated how the combined use of the analytical tools here was not only able to identify the geographical risk of PRRSv outbreaks but also were able to distinguish between endemic and emerging strains.
Although the highest incidence of PRRSv outbreaks (>%25) was observed in southwestern Minnesota and eastern South Dakota between 1998 and 2016 (Fig. 1A), our final Maxent model identified that the geographical risk of PRRSV outbreaks also included northwestern Iowa (Fig. 1B). The AUC values of the final Maxent model suggested that the selected environmental variables were adequate predictors for PRRSv outbreaks between 1998 and 2016 (Table 1). Pig density accounted for most of the background spatial risk in the Midwest (Table 1), as suggested elsewhere5, 18, 19. However, climate seasonality and land cover features had a relatively large role (approximately 40% relative contribution; Table 1) in predicting the risk of PRRSv outbreaks. Indeed, the combination of rainy and windy seasons, the large temperature variation over the course of the year, and proximity to permanent wetland, a dominant demographic characteristic of the Midwest, led to seasonal increases in the number of outbreaks, and contributed in maintaining the endemic state of PRRSv50. Despite these findings; it is important to point out that farms in the Midwest are mainly closed facilities where pigs are contained in a closed environment to minimize the impact of climate. Thus, the effect of climate and land cover may be due, at least in part, to climate- related anthropogenic activities, such as management practices, pig or semen movements and nearby farming activities, which occurs on seasonal patterns, as suggested elsewhere5, 19.
Results of the ORF5 gene BSg reconstruction revealed similar seasonal increases in the relative population size of the three PRRSv clades, which agrees with results from the final Maxent model described above and highlights the role of climate seasonality and climate-anthropogenic related activities in shaping the evolution of endemic viral strains. This finding, that area spread of PRRSv in high pig-dense regions (such as the Midwest) during certain environmental conditions (e.g. seasonal winds), has been previously suggested38. However, results from the spatial diffusion models (Table 2) for clades A and B showed that the transmission of the virus was heterogeneous rather than homogenous, suggesting that long-distance pig movements, climate- related anthropogenic activities, or other unexplored indirect networks (e.g. service providers) might play a larger role compared to aerosol transmission per se in dispersal of the virus across the Midwest51, 52. Our results are in agreement with past studies, which suggested that the shape of pig transportation networks within and between production regions have a significant role in shaping the phylogeny of the virus and mirrors the spatial diffusion of endemic strains14, 33, 53.
However, results from other phylodynamic analyses revealed distinct evolutionary characteristics of PRRSv isolated in the Midwest and could distinguish between endemic and emerging viral strains. For example, while all selected clades showed typical seasonality in the population size overtime, viruses of clades A and B favoured the expansion model (which assumes that population growth increases over time54), and exponential model (which assumes that population growth is fixed overtime54) (Table S2), viruses of clade C significantly favoured the logistic model (Table S2), which assumes that the population growth of the virus decreases over time54. In addition, the inferred growth rate for clades A and B was similar, but was 90% slower than that of Clade C. This result suggests that clade C was an emerging strain, which rapidly spread across the Midwest and then reached a certain equilibrium or died out. Indeed, the distinct fast growth rate (Fig. 3C) of emerging strains, such as those observed in clade C, accelerates their peak genetic diversity, which subsequently lead to population decline. Ecologically speaking, such observation may suggest that environmental conditions in the Midwest may accelerate the spread of emerging strains, ultimately leading to virus population decline due to the rapid decrease in the size of the susceptible population. Furthermore, clade C contained viruses that belonged to RFLP type 1-7-4 and some of its close relatives, which has been reported previously33 as an emerging virulent strain which caused severe outbreaks between 2014 and 2015 across the U.S. Moreover, clade C demonstrated a substantially larger mean evolutionary and dispersal rates than clades A and B (Table 2), suggesting a distinct evolutionary behaviour of an emerging PRRSv strain. Finally, Fig. 5 demonstrates the rapid and widespread of the MCC tree branches of clade C all other the Midwestern swine production system, and did not accumulate in the highest-risk areas predicted by the Maxent model, like clades A and B. This result suggests that clade C evolution and spread is not maintained by the environmental characteristics of the Midwest region.
Both clades A and B had similar inferred posterior dispersal rates (16.3 and 13.1 km/year, respectively; Table 2), suggesting that both clades share similar spatiotemporal evolutionary patterns. Additionally, our inferred MCC trees for both clades demonstrated initial low among-branch spatial diffusion rate variation at the early stages of the epidemic, followed by a substantial increase in the rate variation at the branches’ tips (Fig. 4). This observation is also supported by Fig. 5, where dispersal of the MCC trees’ branches was substantially more intense in the later years than earlier years, suggesting that the dispersal rate of endemic PRRSV strains might continuously increase onward, in parallel to the expanding pig population densities in the Midwest region over time. Figure 5 also demonstrates how most of the MCC trees’ branches are concentrated in, and encompassed by the highest risk areas (probability >0.7) predicted by the Maxent model, suggesting that those geographical areas provide sufficient environmental requirements for the circulation and maintenance of PRRSv endemic strains.
Similar to clades A and B, results from the spatial diffusion model suggest that clade C followed a heterogeneous spatial diffusion process (Table 2), indicating that most PRRSvs, circulating or emerging in the Midwest region, are most likely transmitted or maintained by geographically heterogenous activities. This conclusion is important, because airborne spread of this particular clade would have likely resulted on homogeneous dispersal. In contrast, the heterogeneous dispersal found here suggests that the clade spread by mechanisms other than airborne spread only. Furthermore, our inferred MCC trees for clade C demonstrated, similarly to the other clades assessed here, an initial low among-branch spatial diffusion rate variation, followed by a substantial increase in the rate variation at the branches’ tips at the later stages of the virus dispersal (Fig. 4). However, both mean evolutionary and dispersal rates were substantially larger compared to those inferred for clades A and B (Table 2). Such behaviour is consistent with the emerging nature of clade C, because herds with a population that is naïve against emerging strains would accelerate virus mutation within the farm and, the virus geographical dispersal rate due to a subsequent increase in the between-farm transmission rates, associated with animal transportation activities.
This study represents the first attempt of using continuous spatial diffusion models for inferring posterior phylogeographic dispersal rates for PRRSv endemic strains (Table 2, Fig. 4). Inferring phylogeographic dispersal rates can be a useful measure for modeling the unobserved patterns of virus evolution in space and time44, and hence, shed deeper insights onto the degree of severity of virus transmission and spread within a swine production system over time. We expect that the analytical framework presented here will set the scene for improved surveillance aimed for early detection of new virulent emerging PRRSv strains, and subsequently guidance for risk-based intervention strategies. Our analytical approach can be used in the future for viruses that may emerge and re-emerge within the swine and other food animal industries, such as circovirus55.
Although Bayesian phylodynamic methods became well established for rapidly evolving RNA viruses like PRRS56, 57, past PRRSv studies continued using traditional phylogenetic methods (e.g. ML trees) to characterize new emerging strains, without accounting for their related evolutionary parameters, spatial or temporal information58,59,60,61,62. Furthermore, they continued to use the RFLP patterns to genotype the newly detected strains and made unrealistic and unsupported conclusions about their origins, transmission and evolution. A comparison between the MCC trees of the three selected clades (Fig. 4) and the ML tree (Figure S1), shows substantial discrepancies between their topologies estimated by the Bayesian phylodynamic and ML methods, respectively. Furthermore, the MCC tree (Figure S2) demonstrates how the past RFLP pattern classification method based on the ML tree for PRRSvs was redundant and doesn’t correctly place newly detected viral strains in their truly related viral clade or cluster. In contrast, by accounting for important evolutionary and epidemiological information in our Bayesian phylodynamic models, we were able to estimate and quantify many inferences (Table 2) that distinguish between endemic and emerging strains and, therefore, we made realistic and data-supported conclusions about their evolutionary, spatial and temporal characters. Our analytical approach offers a realistic and robust framework for spatial inference from genetic data, when information about the specific geographical locations of the outbreaks (latitude/longitude) is available. Past studies14, 33 attempted to infer the phylogeographic history of the virus using discrete traits (i.e. countries or regions, instead of actual geographical locations), which doesn’t provide a realistic representation of the spatial diffusion process of the virus within a geographical region. Indeed, here we could not only provide a realistic representation of the spatiotemporal diffusion process of the virus within the Midwest, but also we were able to quantify the spatial dispersal rate of this diffusion process and distinguish between the phylogeographic evolutionary characteristics of endemic and emerging strains. Furthermore, we could, indirectly, establish relationships between the geographical suitability for endemic strains (which has been predicted spatial risk by Maxent model) and their evolutionary history. To our knowledge there is no valid quantitative method that can statistically link result outputs from both analytical methods. That said, we must acknowledge that the direct inclusion of the Maxent environmental predictors into the phylodynamic analysis might improve our posterior inferences about the evolutionary dynamics of the virus in the Midwest. Lemey et al., (2014) developed an extension for his novel Bayesian discrete phylogeographic inference37, that can include and exclude environmental variables into the phylodynamic model56, but the method is currently not optimized for continuous space phylogeography, as is the case of the study here. Optimizing the GLM method for continuous space phylogeography would be of great value for future studies as well as for molecular surveillance, as it yields more realistic inferences about the spatial and evolutionary epidemiology of rapidly evolving pathogens like PRRSv. Therefore, our study represents a motivation to pursue future studies related to establishing a full quantitative pipeline for combining the inferences derived from both analytical methods.
Another limitation of the present study was attributed to the lack of information related to spatiotemporal patterns of pig movements. Such information would substantially improve the inferences of the methods presented here. With the continuous advancements in the phylodynamic methods, described above, the inclusion of pig movement into future phylogeographic models of PRRSv will soon be possible as it would, indeed, yield a more realistic inference about the spatiotemporal evolutionary patterns of PRRSv endemic and emerging strains. Finally, data represented only a sample from the region, and data from other farms and systems may have helped to improve our predictions; noteworthy, however, the quantity of data available to us is unprecedented.
In conclusion, PRRS viruses are characterized by typical seasonality in their population size. However, endemic strains are characterized by a substantially slower population growth and evolutionary rates, as well as smaller spatial dispersal rates when compared to emerging strains. Thus, study here demonstrates an analytical framework for inferring the evolutionary history of strains that coincidently circulate in a region. The framework provided valuable inferences with application to disease control, including, for example, predictions on the likelihood that specific clades or strains will continue to spread, or not, and the most likely mechanism of spread, as suggested by the homogeneity of the dispersal pattern. While we were unable to provide a direct quantitative link between the analytical methods used in this study, the results presented here will help to inform decisions on disease surveillance and control for, arguably, one of the most important non-regulated diseases affecting one of the largest food animal industries in the world.
Materials and Methods
Species distribution modeling
The Maxent method is implemented as a function in the ‘Dismo’ package in R63. This method has recently become popular for predicting spatial distribution of infectious diseases of both public health and veterinary significance and has been described extensively elsewhere64, 65. We used the default logistic model, convergence threshold, regularization, and number of iterations. We used a Jackknife test to calculate the contribution of each environmental variable to the final Maxent model, and evaluated the performance of the candidate Maxent models by partitioning the data into training and testing sets and using the threshold independent method (TIm)66. We set the TIm k-fold partitioning scheme to create 5 partitions and randomly sample each partition with replacement, where our candidate Maxent model was tested five times (k = 5) against 10,000 randomly generated background points (pseudo-absences). Subsequently, we calculated the area under the curve (AUC) through a receiver operator characteristic (ROC) plot of the sensitivity vs. 1 - specificity over the whole range of threshold values between 0 and 1. We used training AUC for model building, whereas the testing sets to evaluate model accuracy using the average value of the AUC calculated for each partitioned set. AUC values >0.75 for both training and testing data were considered reliably discriminating models and indicated that the selected environmental layers were adequate predictors67, 68. Finally, because the geographic extent of the study area was fairly large, we used a calibrated AUC (cAUC) final Maxent model to evaluate the presence of the spatial sorting bias (SSB) as suggested elsewhere69. If the cAUC value was close to 1, then we concluded the absence of SSB (i.e. locations within the Midwest region with the highest number of observed outbreaks have a small impact on the resulted Maxent model), whereas if the value was close to zero, we concluded the opposite. We then plotted the spatial probability distribution of PRRSV outbreaks predicted by the final Maxent model (Fig. 1B) using ArcGIS version 10.441.
Preliminary phylogenetic analysis
We converted the collection date for each sequence into fractional years (decimal days) to estimate divergence times. Then, we aligned the ORF5 sequences using MUSCLE70 and confirmed the reading frame by examining the amino-acid translation using AliView (Lorenz et al., 2011). Finally, we found no homologous recombinant sequences using the Recombination Detection Program71. Next, we used PartitionFinder72 to define the partitioning scheme of our alignment corresponding to the codon positions of the ORF5 protein-coding gene and we selected the most realistic partition schemes for the set of substitution models implemented in BEAST 1.8.443 for the subsequent analyses based on the value of Bayesian Information Criterion (BIC). Then, we estimated the maximum-likelihood (ML) of the phylogeny for all ORF5 sequences, using RAxML73, and examined the tree topology under the GTR + Γ substitution model, with 100 non-parametric bootstrap replicate searches. Finally, we selected three well-supported phylogenetic clades, referred to as clades A, B and C, and ensured that each selected clade’s spatial and temporal distribution well represented the geographical extent and period of the study (Fig. 2). Clade A compromised 191 non-identical sequences collected between May 2009 and April 2016, clade B compromised 214 non-identical sequences collected between August 2006 and February 2016, and clade C compromised 187 non-identical sequences, collected between December 2008 and April 2016. Again, we performed RAxML analyses on each selected clade to compare the topology of the ML tree to the corresponding estimated topology of the posterior phylogeny in the subsequent Bayesian analyses (Figure S1).
Model selection and estimation of virus evolutionary demographics
We inferred posterior phylogenetic relationships and demographic for each of the three PRRSv clades from the alignment of the sampled ORF5 sequences within a Bayesian statistical framework using relaxed-clock models implemented in BEAST 1.8.443. We used a 3-codon position mixed GTR + Γ substitution model to realistically represent the sequence data for all the three clades (as indicated by the value of the BIC using ParitionFinder). Next, we evaluated five tree prior models for each of the three clades under a separate analytical setting, including (1) the constant population size (CP)74; (2) the exponential growth (EG)45; (3) the expansion growth (EGx)54; (4) the logistic growth (LG)54 parametric coalescent models; and (5) the nonparametric, piecewise-constant Bayesian Skygrid (BSg) coalescent model, which employ a Gassian Markov Random Field (GMRF) prior to smoothing the trajactories of the past population dynamics (BSg-GMRF)75. Furthermore, for each tree prior model, we evaluated two branch-rate models using: (1) the uncorrelated lognormal (UCLN)76 branch-rate prior model, and; (2) the uncorrelated exponential (UCED)76 branch-rate prior model. We estimated parameters of the branch-rate prior distributions using the continuous-time Markov chain (CTMC)-rate reference 77 as a hyperprior for the mean of the lognormal and exponential distributions, and an exponential hyperprior for the standard deviation of the lognormal distribution. Isolation dates of the sequences were used to calibrate tree-height (divergence times) estimates. We estimated the marginal likelihood of each candidate relaxed-clock models (ten candidate models for each of the three clades) using path-sampling (PS)77, 78 and stepping-stone sampling (SS)79,80,81 estimators. We estimated the posterior evolutionary parameters, phylogeny and population demographics under each candidate relaxed model using two replicate MCMC simulations for 200 million cycles and sampled every 20,000th state, to assess the stability of the marginal-likelihood estimates. We used Tracer v. 1.682 to assess the reliability of the MCMC simulations by examining the convergence of each MCMC simulation to the stationary distribution and calculating effective sample size (ESS)83 for every parameter. We assessed mixing of each chain over the stationary distribution by monitoring the acceptance rates for all parameters. We then evaluated the fit of each candidate relaxed-clock models based on the resulting marginal-likelihood estimates using the Bayes factor (BF) comparison approach (Table S2). Finally, we used the best-fitting branch-rate prior (UCED v.s. UCLN) with the BSg-GMRF coalescent model (Table S2) to generate a BSg plot of the effective population size trajectories between 1998 and 2016 for each PRRSv clade.
Relaxed random-walk phylogeographic analyses
We extended the above BEAST analyses to infer the spatio-temporal evolutionary history of each clade using the continuous phylogeographic diffusion models, suggested by Lemey et al.84. This approach models movement of the virus in two dimensions as a scaled-mixture generalization of a Brownian motion process83, 84. Briefly, the method implements a diffusion rate variation by rescaling the diffusion process along each branch of the inferred posterior phylogeny83, 84. This procedure is achieved through scalers drawn from a specific distribution, which generates a scaled mixture of a wide range of relaxed-random walks. We used the priors of the best-fitting demographic models (coalescent tree prior/branch rate combinations), from the above analyses (Table S2), and extended the analyses using additional, independent runs to assess for the best continuous trait model that accommodate among-branch diffusion rate variation for each selected viral clade. This procedure included additional simulations (in duplicates to assess the stability of the resulting marginal-likelihoods) to evaluate the four continuous trait models implemented in BEAST, which included the simple naïve homologous Brownian model, and three heterogeneous relaxed random walks (RRW) models (Cauchy, gamma and lognormal probability distribution). We ran the subsequent MCMC simulations for 500 million states and sampled every 50,000 states. We used the resulting marginal-likelihoods, estimated by the PS and SS methods, to selected the best-fitting continuous trait model for each viral clade by performing additional BF comparisons.
We used TreeAnnotator to summarize the posterior distribution of the phylogenies for the best-fitting spatio-temporal diffusion model of each clade, in the form of a maximum clade credibility (MCC) trees. We then used FigTree85 to plot the resulting MCC trees colored by the inferred among-branch spatial diffusion rate variation for each clade. Finally, we used spreaD386 to generate keyhole markup language (KML) files to visualize clades’ MCC trees, and then superimposed them on the resulting risk map of PRRSv outbreaks predicted by the final Maxent model using Google Earth Pro.
References
Pork Checkoff. Top 10 Pork-Producing Countries Accessed December 2016. Available online: http://www.pork.org/pork-quick-facts/home/stats/u-s-pork-exports/top-10-pork-producing-countries/(2016).
Quaife, T. Scramble is on to solve mystery disease. Swine Practice 5, 10 (1989).
Benfield, D. A. et al. Characterization of swine infertility and respiratory syndrome (SIRS) virus (isolate ATCC VR-2332). J Vet Diagn Invest 4, 127–133 (1992).
Holtkamp, D. J. et al. Assessment of the economic impact of porcine reproductive and respiratory syndrome virus on United States pork producers. Journal of Swine Health and Production 21, 72–84 (2013).
Perez, A. M. et al. Lessons learned and knowledge gaps about the epidemiology and control of porcine reproductive and respiratory syndrome virus in North America. J Am Vet Med Assoc 246, 1304–1317, doi:10.2460/javma.246.12.1304 (2015).
Dokland, T. The structural biology of PRRSV. Virus Res 154, 86–97, doi:10.1016/j.virusres.2010.07.029 (2010).
Nilubol, D., Tripipat, T., Hoonsuwan, T., Tipsombatboon, P. & Piriyapongsa, J. Genetic diversity of the ORF5 gene of porcine reproductive and respiratory syndrome virus (PRRSV) genotypes I and II in Thailand. Arch Virol 158, 943–953, doi:10.1007/s00705-012-1573-7 (2013).
Thuy, N. T. et al. Genetic analysis of ORF5 porcine reproductive and respiratory syndrome virus isolated in Vietnam. Microbiol Immunol 57, 518–526, doi:10.1111/1348-0421.12067 (2013).
Chen, N., Trible, B. R., Kerrigan, M. A., Tian, K. & Rowland, R. R. ORF5 of porcine reproductive and respiratory syndrome virus (PRRSV) is a target of diversifying selection as infection progresses from acute infection to virus rebound. Infect Genet Evol 40, 167–175, doi:10.1016/j.meegid.2016.03.002 (2016).
Ropp, S. L. et al. Characterization of emerging European-like porcine reproductive and respiratory syndrome virus isolates in the United States. J Virol 78, 3684–3703 (2004).
Rowland, R. R. & Morrison, R. B. Challenges and opportunities for the control and elimination of porcine reproductive and respiratory syndrome virus. Transbound Emerg Dis 59(Suppl 1), 55–59, doi:10.1111/j.1865-1682.2011.01306.x (2012).
Dea, S., Gagnon, C. A., Mardassi, H., Pirzadeh, B. & Rogan, D. Current knowledge on the structural proteins of porcine reproductive and respiratory syndrome (PRRS) virus: comparison of the North American and European isolates. Arch Virol 145, 659–688 (2000).
Shi, M. et al. Phylogeny-based evolutionary, demographical, and geographical dissection of North American type 2 porcine reproductive and respiratory syndrome viruses. J Virol 84, 8700–8711, doi:10.1128/JVI.02551-09 (2010).
Shi, M. et al. The spread of type 2 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) in North America: a phylogeographic approach. Virology 447, 146–154, doi:10.1016/j.virol.2013.08.028 (2013).
Tousignant, S. J., Perez, A. M., Lowe, J. F., Yeske, P. E. & Morrison, R. B. Temporal and spatial dynamics of porcine reproductive and respiratory syndrome virus infection in the United States. Am J Vet Res 76, 70–76, doi:10.2460/ajvr.76.1.70 (2015).
Arruda, A. G. et al. Estimation of Time-Dependent Reproduction Numbers for Porcine Reproductive and Respiratory Syndrome across Different Regions and Production Systems of the US. Frontiers in Veterinary Science 4: 46, doi:10.3389/fvets.2017.00046 (2017).
Corzo, C. A. et al. Control and elimination of porcine reproductive and respiratory syndrome virus. Virus Res 154, 185–192, doi:10.1016/j.virusres.2010.08.016 (2010).
Holtkamp, D. J., Yeske, P. E., Polson, D. D., Melody, J. L. & Philips, R. C. A prospective study evaluating duration of swine breeding herd PRRS virus-free status and its relationship with measured risk. Prev Vet Med 96, 186–193, doi:10.1016/j.prevetmed.2010.06.016 (2010).
Holtkamp, D. J., Lin, H., Wang, C. & O’Connor, A. M. Identifying questions in the American Association of Swine Veterinarian’s PRRS risk assessment survey that are important for retrospectively classifying swine herds according to whether they reported clinical PRRS outbreaks in the previous 3 years. Prev Vet Med 106, 42–52, doi:10.1016/j.prevetmed.2012.03.003 (2012).
Truong, V. M. & Gummow, B. Risk factors for porcine reproductive and respiratory syndrome outbreaks in Vietnamese small stock farms. N Z Vet J 62, 199–207, doi:10.1080/00480169.2014.888640 (2014).
Pearson, R. G. Species’ Distribution Modeling for Conservation Educators and Practitioners. Synthesis, Accessed Jan 2016. Available: http://www.amnh.org (2007).
Scholte, R. G., Carvalho, O. S., Malone, J. B., Utzinger, J. & Vounatsou, P. Spatial distribution of Biomphalaria spp., the intermediate host snails of Schistosoma mansoni, in Brazil. Geospat Health 6, S95–S101 (2012).
Alkhamis, M. A. & VanderWaal, K. Spatial and Temporal Epidemiology of Lumpy Skin Disease in the Middle East, 2012–2015. Front Vet Sci 3, 19, doi:10.3389/fvets.2016.00019 (2016).
Belkhiria, J., Alkhamis, M. A. & Martínez-López, B. Application of Species Distribution Modeling for Avian Influenza surveillance in the United States considering the North America Migratory Flyways. Scientific Reports 6, 33161, doi:10.1038/srep33161.
Fuller, T. L. et al. Mapping the risk of avian influenza in wild birds in the US. BMC infectious diseases 10, 187, doi:10.1186/1471-2334-10-187 (2010).
Alvarez, J. et al. Novel analytic tools for the study of porcine reproductive and respiratory syndrome virus (PRRSv) in endemic settings: lessons learned in the U.S. Porcine Health Management 2, 1–9, doi:10.1186/s40813-016-0019-0 (2016).
Goldberg, T. L., Hahn, E. C., Weigel, R. M. & Scherba, G. Genetic, geographical and temporal variation of porcine reproductive and respiratory syndrome virus in Illinois. J Gen Virol 81, 171–179 (2000).
Yoon, S. H. et al. Complete genome sequences of porcine reproductive and respiratory syndrome viruses: perspectives on their temporal and spatial dynamics. Mol Biol Rep, doi:10.1007/s11033-013-2802-1 (2013).
Rosendal, T. et al. Spatial and temporal patterns of porcine reproductive and respiratory syndrome virus (PRRSV) genotypes in Ontario, Canada, 2004–2007. BMC Vet Res 10, 83, doi:10.1186/1746-6148-10-83 (2014).
Larochelle, R., D’Allaire, S. & Magar, R. Molecular epidemiology of porcine reproductive and respiratory syndrome virus (PRRSV) in Quebec. Virus Res 96, 3–14 (2003).
Alonso, C., Murtaugh, M. P., Dee, S. A. & Davies, P. R. Epidemiological study of air filtration systems for preventing PRRSV infection in large sow herds. Prev Vet Med 112, 109–117, doi:10.1016/j.prevetmed.2013.06.001 (2013).
Chen, J. Z. et al. Characterization of two novel porcine reproductive and respiratory syndrome virus isolates with deletions in the GP2 gene. Vet Microbiol 176, 344–351, doi:10.1016/j.vetmic.2015.01.018 (2015).
Alkhamis, M. A., Perez, A. M., Murtaugh, M. P., Wang, X. & Morrison, R. B. Applications of Bayesian Phylodynamic Methods in a Recent U.S. Porcine Reproductive and Respiratory Syndrome Virus Outbreak. Front Microbiol 7, 67, doi:10.3389/fmicb.2016.00067 (2016).
Suchard, M. A., Weiss, R. E. & Sinsheimer, J. S. Bayesian selection of continuous-time Markov chain evolutionary models. Molecular biology and evolution 18, 1001–1013 (2001).
Drummond, A. J., Nicholls, G. K., Rodrigo, A. G. & Solomon, W. Estimating mutation parameters, population history and genealogy simultaneously from temporally spaced sequence data. Genetics 161, 1307–1320 (2002).
Grenfell, B. T. et al. Unifying the epidemiological and evolutionary dynamics of pathogens. Science 303, 327–332, doi:10.1126/science.1090727 (2004).
Lemey, P., Rambaut, A., Drummond, A. J. & Suchard, M. A. Bayesian phylogeography finds its roots. PLoS Comput Biol 5, e1000520, doi:10.1371/journal.pcbi.1000520 (2009).
Brito, B., Dee, S., Wayne, S., Alvarez, J. & Perez, A. Genetic diversity of PRRS virus collected from air samples in four different regions of concentrated swine production during a high incidence season. Viruses 6, 4424–4436, doi:10.3390/v6114424 (2014).
Nguyen, V. G. et al. A Bayesian phylogeographical analysis of type 1 porcine reproductive and respiratory syndrome virus (PRRSV). Transbound Emerg Dis 61, 537–545, doi:10.1111/tbed.12058 (2014).
Chaikhumwang, P. et al. Dynamics and evolution of highly pathogenic porcine reproductive and respiratory syndrome virus following its introduction into a herd concurrently infected with both types 1 and 2. Infect Genet Evol 30, 164–174, doi:10.1016/j.meegid.2014.12.025 (2015).
ESRI. ArcGIS Desktop: Release 10.4 Redlands, CA: Environmental Systems Research Institute (2016).
Phillips, S. J., Anderson, R. P. & Schapire, R. E. Maximum entropy modeling of species geographic distributions. Ecological Modelling 190, 231–259 (2006).
Drummond, A. J., Suchard, M. A., Xie, D. & Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Molecular biology and evolution 29, 1969–1973, doi:10.1093/molbev/mss075 (2012).
USDA. United States department of Agriculture Census of Agriculture Accessed Jan 2016. Availabe: https://www.agcensus.usda.gov/(2012).
Hijmans, R. J., Cameron, S. E., Parra, J. L., Jones, P. G. & Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology 25, 1965–1978 (2005).
Reeves, T., Samy, A. M. & Peterson, A. T. MERS-CoV geography and ecology in the Middle East: analyses of reported camel exposures and a preliminary risk map. BMC research notes 8, 801, doi:10.1186/s13104-015-1789-1 (2015).
USGS. United States geographical survey MODIS-based global land cover climatology Accessed March 2016. Available: http://www.landcover.usgs.gov/(2016).
Hijmans, R. J. R: Geographic Data Analysis and Modeling Accessed April 2016. Available: http://CRAN.R-project.org/package=raster (2015).
R Core Team R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/(2013).
Hermann, J. et al. Effect of temperature and relative humidity on the stability of infectious porcine reproductive and respiratory syndrome virus in aerosols. Vet Res 38, 81–93, doi:10.1051/vetres:2006044 (2007).
Arruda, A. G. et al. Investigation of the Occurrence of Porcine Reproductive and Respiratory Virus in Swine Herds Participating in an Area Regional Control and Elimination Project in Ontario, Canada. Transboundary and Emerging Diseases n/a–n/a, doi:10.1111/tbed.12343 (2015).
Arruda, A. G., Friendship, R., Carpenter, J., Hand, K. & Poljak, Z. Network, cluster and risk factor analyses for porcine reproductive and respiratory syndrome using data from swine sites participating in a disease control program. Prev Vet Med 128, 41–50, doi:10.1016/j.prevetmed.2016.03.010 (2016).
Brar, M. S., Shi, M., Murtaugh, M. P. & Leung, F. C. Evolutionary Diversification of Type 2 Porcine Reproductive and Respiratory Syndrome Virus. J Gen Virol, doi:10.1099/vir.0.000104 (2015).
Griffiths, R. C. & Tavare, S. Ancestral Inference in Population Genetics. 307–319, doi:10.1214/ss/1177010378 (1994).
Palinski, R. et al. A Novel Porcine Circovirus Distantly Related to Known Circoviruses Is Associated with Porcine Dermatitis and Nephropathy Syndrome and Reproductive Failure. J Virol 91, doi:10.1128/jvi.01879-16 (2017).
Pybus, O. G., Fraser, C. & Rambaut, A. Evolutionary epidemiology: preparing for an age of genomic plenty. Philos Trans R Soc Lond B Biol Sci 368, 20120193, doi:10.1098/rstb.2012.0193 (2013).
Lemey, P. et al. Unifying viral genetics and human transportation data to predict the global transmission dynamics of human influenza H3N2. PLoS Pathog 10, e1003932, doi:10.1371/journal.ppat.1003932 (2014).
Sinn, L. J. et al. Characterization of two Austrian porcine reproductive and respiratory syndrome virus (PRRSV) field isolates reveals relationship to East Asian strains. Vet Res 47, 17, doi:10.1186/s13567-015-0293-x (2016).
Wilkinson, J. M. et al. Genome-wide analysis of the transcriptional response to porcine reproductive and respiratory syndrome virus infection at the maternal/fetal interface and in the fetus. BMC Genomics 17, 383, doi:10.1186/s12864-016-2720-4 (2016).
Workman, A. M., Smith, T. P., Osorio, F. A. & Vu, H. L. Complete Genome Sequence of Highly Virulent Porcine Reproductive and Respiratory Syndrome Virus Variants That Recently Emerged in the United States. Genome Announc 4, doi:10.1128/genomeA.00772-16 (2016).
Yang, T. et al. A genome-wide association study of fetal response to type 2 porcine reproductive and respiratory syndrome virus challenge. Sci Rep 6, 20305, doi:10.1038/srep20305 (2016).
Zhang, Q. et al. Emerging of two new subgenotypes of porcine reproductive and respiratory syndrome viruses in Southeast China. Microb Pathog 97, 27–33, doi:10.1016/j.micpath.2016.05.011 (2016).
Hijmans, R.J, Phillips, S., Leathwick, J. and Elith, J. Package ‘dismo’ Accessed March 2016. Available: http://cran.r-project.org/web/packages/dismo/index.html (2011).
Alkhamis, M. A. & VanderWaal, K. L. Spatial and Temporal Epidemiology of Lumpy Skin Disease in the Middle East, 2012–2015. Frontiers in Veterinary Science 3, doi:10.3389/fvets.2016.00019 (2016).
Alkhamis, M., Hijmans, R. J., Al-Enezi, A., Martinez-Lopez, B. & Perea, A. M. The Use of Spatial and Spatiotemporal Modeling for Surveillance of H5N1 Highly Pathogenic Avian Influenza in Poultry in the Middle East. Avian Dis 60, 146–155, doi:10.1637/11106-042115-Reg (2016).
Elith, J. & Leathwick, J. R. Species Distribution Models: Ecological Explanation and Prediction Across Space and Time. Annu Rev Ecol Evol S 40, 677–697, doi:10.1146/annurev.ecolsys.110308.120159 (2009).
Soberón, J. Interpretation of models of fundamental ecological niches and species’ distributional areas, http://hdl.handle.net/1808/6566 (2005).
Elith, J. et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 29, 129–151, doi:10.1111/j.2006.0906-7590.04596.x (2006).
Hijmans, R. J. Cross-validation of species distribution models: removing spatial sorting bias and calibration with a null model. Ecology 93, 679–688 (2012).
Edgar, R. C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res 32, 1792–1797, doi:10.1093/nar/gkh340 (2004).
Martin, D. P. et al. RDP3: a flexible and fast computer program for analyzing recombination. Bioinformatics 26, 2462–2463, doi:10.1093/bioinformatics/btq467 (2010).
Lanfear, R., Calcott, B., Ho, S. Y. & Guindon, S. Partitionfinder: combined selection of partitioning schemes and substitution models for phylogenetic analyses. Molecular biology and evolution 29, 1695–1701, doi:10.1093/molbev/mss020 (2012).
Stamatakis, A. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics, doi:10.1093/bioinformatics/btu033 (2014).
Kingman, J. F. C. On the Genealogy of Large Populations. Journal of Applied Probability 19, 27–43, doi:10.2307/3213548 (1982).
Gill, M. S. et al. Improving Bayesian population dynamics inference: a coalescent-based model for multiple loci. Molecular biology and evolution 30, 713–724, doi:10.1093/molbev/mss265 (2013).
Drummond, A. J., Ho, S. Y., Phillips, M. J. & Rambaut, A. Relaxed phylogenetics and dating with confidence. PLoS biology 4, e88, doi:10.1371/journal.pbio.0040088 (2006).
Ferreira, M. A. R. & Suchard, M. A. Bayesian analysis of elapsed times in continuous-time Markov chains. The Canadian Journal of Statistics/La Revue Canadienne de Statistique 36, 355–368 (2008).
Lartillot, N. & Philippe, H. Computing Bayes factors using thermodynamic integration. Systematic biology 55, 195–207, doi:10.1080/10635150500433722 (2006).
Baele, G. et al. Improving the accuracy of demographic and molecular clock model comparison while accommodating phylogenetic uncertainty. Molecular biology and evolution 29, 2157–2167, doi:10.1093/molbev/mss084 (2012).
Xie, W., Lewis, P. O., Fan, Y., Kuo, L. & Chen, M. H. Improving marginal likelihood estimation for Bayesian phylogenetic model selection. Systematic biology 60, 150–160, doi:10.1093/sysbio/syq085 (2011).
Fan, Y., Wu, R., Chen, M. H., Kuo, L. & Lewis, P. O. Choosing among partition models in Bayesian phylogenetics. Mol Biol Evol 28, 523–532, doi:10.1093/molbev/msq224 (2011).
Rambaut, A., Suchard, M. A., Xie, D. & Drummond, A. J. Tracer version 1.6 Accessed January 2016. Available: http://beast.bio.ed.ac.uk/Tracer (2014).
Brooks, S. P. & Gelman, A. General methods for monitoring convergence of iterative simulations. Journal of Computational and Graphical Statistics 7, 434–455 (1998).
Lemey, P., Rambaut, A., Welch, J. J. & Suchard, M. A. Phylogeography takes a relaxed random walk in continuous space and time. Molecular biology and evolution 27, 1877–1885, doi:10.1093/molbev/msq067 (2010).
Rambaut, A. FigTree Version 1.4.3 Accessed January 2016. Available: http://tree.bio.ed.ac.uk/software/figtree/(2016).
Bielejec, F. et al. SpreaD3: Interactive Visualization of Spatiotemporal History and Trait Evolutionary Processes. Mol Biol Evol 33, 2167–2169, doi:10.1093/molbev/msw082 (2016).
Acknowledgements
This study was funded by the Swine Health Information Center, the National Pork Board, and MnDrive.
Author information
Authors and Affiliations
Contributions
All authors contributed in the conception and design of this study. Dr. M.A.A. designed and ran the statistical analyses, wrote the draft of the manuscript and prepared all the figures presented. Dr. A.G.A. contributed in data organization, running the statistical analyses and interpretation of the results. Dr. R.B.M. acquired the data from the producers and contributed to the interpretation of the results. Dr. A.M.P. supervised the analyses, contributed to the interpretation of the results and coordinated the communication and efforts among the co-authors. All authors contributed to the editing and comprehensive revision of this manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Alkhamis, M.A., Arruda, A.G., Morrison, R.B. et al. Novel approaches for Spatial and Molecular Surveillance of Porcine Reproductive and Respiratory Syndrome Virus (PRRSv) in the United States. Sci Rep 7, 4343 (2017). https://doi.org/10.1038/s41598-017-04628-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-04628-2
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
