
Abstract
Radionuclides, including 137Cs, were released from the disabled Fukushima Daiichi Nuclear Power Plant and had been deposited broadly over forested areas of north-eastern Honshu Island, Japan. In the forest, 137Cs was highly concentrated on leaf litters deposited in autumn 2010, before the accident. Monitoring of the distribution of 137Cs among functional groups clearly showed the role of the detrital food chain as the primary channel of 137Cs transfer to consumer organisms. Although many studies have reported the bioaccumulation (or dilution) of radioactive materials through trophic interactions, the present results highlight the importance of examining multiple possible pathways (e.g., grazing vs. detrital chains) in the proliferation of 137Cs through food webs. These results provide important insight into the future distribution and transfer of 137Cs within forest ecosystems.
Similar content being viewed by others

Introduction
A large amount of radionuclide was released from the Fukushima Daiichi Nuclear Power Plant (FDNPP) accident after the disastrous earthquake and subsequent tsunami of March 2011. Among the variety of radionuclides released from FDNPP1, including iodine, cesium, strontium and plutonium, cesium 137 (137Cs) is the most worrying radionuclide in the environment, with a half-life of 30 years2. Unlike iodine, which is mainly found in a gaseous form, cesium rapidly binds to aerosols and is thus washed out by rain from contaminated air masses3 and deposited and accumulated on the land surface4. Since most of the Japanese land area is covered by forest, the distribution and transportation of radionuclides within forest ecosystems should be conscientiously monitored (cf. Hashimoto et al.5 and Ohte et al.6).
Many previous studies on the distribution and transfer of radionuclides have focused on “bioaccumulation” and the vertical transition between trophic levels7,8,9. In contrast with this simple view of linear food chains, Polis and Strong10 incorporated a diverse array of connections among species within complex food webs, which are comprised of grazing and detrital food chains. To explore the biological proliferation of 137Cs within forest ecosystems, it is essential to track the 137Cs concentrations of each organism component within complex food webs.
In the case of the FDNPP disaster, because the accident occurred in spring (before leaf emergence), the released 137Cs would have been deposited on the surface of leaf litters on the forest floor. This accumulated 137Cs on leaf litter would likely spread within forest ecosystems through two pathways; grazing and detrital food chains (cf. Polis & Strong10; Fig. 1). Accumulated 137Cs on leaf litter is directly consumed by detritivores and cascades up through detrital food chains. On the other hand, 137Cs in an ionic form could be transferred from leaf litter to plants and thus cascade up through grazing food chains. Therefore, to examine the flow and distribution of radiocesium in a forest ecosystem, the organisms in both terrestrial and aquatic food webs should be carefully monitored. Based on these assumptions, 10 functional groups were chosen as sampling units in the present study (Fig. 1).

Schematic food web of the present study.
Ten functional groups were used as sampling units. Solid lines indicate trophic interactions and dashed lines indicate nutrient flow. Broken lines show spatial movements; e.g., transformation of tadpoles to frogs and dragonflies or supply of leaf litters from forest to stream.
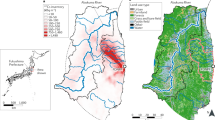
In this study, the results of investigations of the Kami-Oguni River catchment in the northern part of the Fukushima Prefecture (Fig. 2) 15 to 18 months after the accident are reported. The main focus of the field observations was the flow of 137Cs among organisms in the continuum of forest and stream ecosystems. To understand the distribution and transportation of 137Cs within the forest ecosystem, we comprehensively collected the food web components shown in Figure 1 and measured their 137Cs radioactivities and nitrogen stable isotope ratio to evaluate trophic transfer among organisms11.

Maps showing the location of study sites at the Kami-Oguni watershed.
The study area was delineated with the geographic coordinates. These maps were attributed to Zenrin, Kingway Ltd., US Dept. of State Geographer, Mapabc.com, DATA SIO, NOAA, U.S. Navy, NGA, GEBCO, Cnes/Spot Image and DigitalGlobe.
Results
137Cs was concentrated in the litter layer, especially in the litters of F (fragmented litter) and H (humus) layers, although the highest concentration was observed in the A1 layer at site 3 (Table 1). For the sites in secondary deciduous forests (sites 1 and 3), the highest concentrations were observed in the F and A1 (top horizon of the mineral soil) layers, respectively, but for site 2 in a conifer plantation, the highest concentrations were observed in the F layer, with higher concentrations also observed in the surface L (litter) layer. The total accumulations were 55,100, 25,800 and 28,900 Bq/m2 at each site, respectively.
The 137Cs concentrations of each functional group are shown in Figure 3. The glm based on Akaike's Information Criterion (AIC) selected the model with the grouping of functional groups shown in Figure 3 (AICbest = 232.5 vs. AICnull = 265.3). The difference in AIC (ΔAIC) between the second best grouping (terrestrial detritivores as a single independent group with the same grouping as in Figure 3) and the best grouping shown in Figure 3 was 1.9. The 137Cs concentration in leaf litter was highest among all components, reaching more than 300,000 Bq/kg. In the terrestrial food web, 137Cs concentrations in fungi and detritivores were also high compared with other components. Conversely, the concentration in terrestrial herbivores was lowest among all functional groups. The leaves of living plants also showed relatively low concentrations of 137Cs with high variability among samples. In the stream food web, the 137Cs concentration in algae was high at an average of 10,800 Bq/kg. The concentration in detritus in stream was relatively low and was much lower than terrestrial leaf litters. The concentrations in stream consumers and predators were also low, at similar levels to detritus in stream.

137Cs concentrations in each functional group.
Different letters beside each box indicate differences in 137Cs concentrations based on the grouping of functional groups with the model selection using glm. The data below the detection limit were excluded from this analysis. Numbers beside the BDL (below detection level) symbols show the number of specimens in BDL.
Nitrogen stable isotope (δ15N) ratio of the samples was used to confirm trophic levels of sample organisms. Based on this analysis, the glm based on AIC detected the effect of trophic levels on δ15N values (AICtrophic level = 378.3 vs. AICnull = 398.1). The δ15N did not differ between terrestrial and aquatic food webs since the model segregating terrestrial and aquatic food webs was not selected, with ΔAIC between the habitat + trophic level model (AIC = 378.9) and the trophic level model being 0.6. The relationship between 137Cs concentration and nitrogen stable isotope ratio (δ15N) is shown in Figure 5. The glm based on AIC detected the correlation between δ15N values and 137Cs concentration (AICδ15N = 150.3 vs. AICnull = 153.5). In this case, the model segregating the terrestrial and aquatic food webs was not selected, with the AIC of δ15N + habitat model being 152.3. The analyses showed that 137Cs decrease with δ15N in both terrestrial and aquatic food webs.

Changes in δ15N value following the trophic levels.
The gap in the mean values of tropic levels 1 to 2 was 2.05 and level 2 to 3 was 2.63, respectively. The model selection in glm revealed differences in δ15N values among trophic levels.

Relationship between δ15N values and 137Cs concentrations in each sample.
Equation: log(137Cs) = −15.7(δ15N) + 1392.3. The data below the detection limit were excluded from the analysis.
Discussion
The present results show that the total accumulation of 137Cs in the studied forest ranged from 26,000 to 55,000 Bq/m2. A large heterogeneity in 137Cs concentration was observed within the study area. In the present study, 137Cs was highly accumulated in leaf litters and soils in the F to A1-layers (Table 1). In deciduous forest (sites 1 and 3), these litters corresponded with the leaves deposited in autumn 2010, before the accident. The accident took place on 11 March 2011, immediately before the budbreak of deciduous trees and the massive release of radionuclides from FDNPP lasted 2 weeks12. Thus, the 137Cs fell and accumulated on the leaf litters on the forest floor, rather than on the foliage of the forest canopy. This may explain why the 137Cs concentrations in surface leaf litter (A0 to L layers) were less than in the lower layers (Table 1). However, in conifer plantation, a higher concentration was observed in the surface L (litter) layer with a lower concentration in the lower layers (F to A2 layers). This may be caused by the interception fraction of the deposited radiocesium by evergreen coniferous canopies13.
These contaminated leaf litters on the forest floor were inevitability decomposed by biological activities. According to a review by Aerts14, the annual decomposition rate of leaf litter in temperate forest is 0.698±0.329 (sd), on average. Applying this value to the present study, about half of (0.698×0.698 = 0.487) the leaf litter deposited in autumn 2010 had been decomposed by the autumn of 2012, when the present study finished. Although it is unclear whether biological or geochemical processes contributed to this process, the present results clearly show the input of considerable amounts of 137Cs to the detrital chain through trophic levels (Figs. 1, 3). In terrestrial food webs, highly contaminated leaf litters are consumed by detritivores, e.g. earthworms and Bibionidae fly larvae, which were heavily contaminated with 137Cs. In contrast, the uptake of 137Cs through the grazing chain was limited. Although a large variation in 137Cs concentrations among plant species was observed, the average value of these concentrations was relatively low in plants compared with organisms in the other functional groups (Fig. 3). This was suggestive of a lower uptake of 137Cs from contaminated soils by plants. The herbivores, which consumed the less-contaminated living plant tissues, showed low concentrations of 137Cs. However, predatory animals, such as lizards and snakes, were considerably contaminated with 137Cs. This clearly shows the uptake of 137Cs through the detrital chain, but not through the grazing chain and the uptake of 137Cs by aboveground generalist predators corresponds with the concept of detrital infusion into grazing food chains15. Based on studies after the Chernobyl accident, higher contaminations of detritivorous insects than herbivorous and predatory insects were reported16,17. High 137Cs concentrations were also observed in fungi, as has been previously reported18,19. These studies suggested that the higher absorption of potassium by fungi leads to the accumulation of 137Cs in their tissues20,21.
The relationship between 137Cs concentrations and δ15N values increases our understanding of the dynamics of radiocesium within the forest ecosystem. The 137Cs concentrations in each sample element decreased with the δ15N value (Fig. 5), which showed the increasing trend through trophic levels (Fig. 4). These studies showed the decrease in 137Cs concentrations through trophic interactions, which was suggestive of biological dilution and not accumulation of 137Cs. This supports our conclusion that the detrital food chain is the primary channel of 137Cs transfer to consumer organisms. The relatively high level of 137Cs contamination of predatory organisms in this forest is explained by detrital infusion, but not by the biological accumulation of 137Cs through less contaminated grazing chains.
In the stream food web, algae were highly contaminated with 137Cs, but detritus showed relatively low 137Cs concentrations (Fig. 3), which may have been caused by the relatively low retention and recent accumulation of litter falls into the stream. The retention time of leaf litters within streams is known to be shorter than that on forest floors because litter in streams is easily washed out by waterflows22. In contrast, the higher 137Cs concentrations of algae was suggestive of an accumulation of 137Cs. Adam et al.23 reported relatively high accumulation of 137Cs by aquatic algae through the dissolved forms of 137Cs from the contaminated water. Because there were no effective sources of 137Cs within the stream at that moment (Fig. 3), the higher 137Cs concentrations of algae may be caused by leaf litters with higher contamination levels supplied from forest floor. Despite the higher contamination of stream algae, the level of 137Cs concentrations of aquatic consumers and predators were relatively low, which was suggestive of the limited inputs of algal production in stream food webs. The biomass of detritus would be much larger than that of algae, which caused the relatively low 137Cs contamination in aquatic consumers and predators (Fig. 3).
It should be mentioned that these results are from the very initial processes after 137Cs fallout and in the forest area with an intermediate level of 137Cs contamination. Future studies should examine the intake of 137Cs to the grazing chain and the changes in the degree of transfer of 137Cs through food webs in forest ecosystems. It is important to determine when and how the 137Cs is transferred through grazing food chains. In this study, the 137Cs concentrations of living plants were relatively low (Fig. 3), but those of a fern (Dryopteris crassirhizoma) and wild cherry (Potentilla hebiichigo) were fairly high compared with the other elements (Table S1). The reason for this variation is unclear, but the depth of the root system24 or mutual interactions with mycorrhizae25 may affect the variation in 137Cs concentrations. The belowground distribution of 137Cs may affect its uptake by plants. Thus, monitoring of the decomposition of the contaminated leaf litters, which are the source of 137Cs and the subsequent diffusion of 137Cs, are important in predicting the dynamics of 137Cs within forest ecosystems. It is also important to determine whether the transfer rate of 137Cs through the food chain varies over time or among sites with different contamination levels. In the present study, most of the measured food web components were annuals, but several amphibians, reptiles and other animals and tree species were perennial and may potentially accumulate (or dilute) the 137Cs, as in bioaccumulation7. Although many previous studies demonstrated the bioaccumulation of 137Cs by a variety of organisms8,9,26 these studies did not examine the effect of food web structure on the distribution of 137Cs among organisms. Furthermore, because the dynamics of 137Cs are known to be affected by the relative amount of the other elements, such as K and stable Cs (133Cs)27, the examination of 137Cs in highly contaminated areas is required to fully understand the distribution and transportation of radionuclides at broader spatio-temporal scales.
Methods
Samples
The study was conducted in a secondary forest at the headwater of the Kami-Oguni River catchment, located 53 km from the FDNPP (Fig. 2). According to a radioactivity survey (December 2012) conducted using aircraft survey devices, the air dose rate in this region was 1.9–3.8 mSv/h and the total deposition rate of 137Cs was estimated as 300,000–600,000 Bq/m228. The upstream area of the study catchment was composed mostly of forested areas, while farmland consisting mainly of paddy fields was the dominant land used in the middle to downstream area of the catchment. The study area for the samplings was a forest-stream ecotone. In the forest, the dominant tree species are oak (Quercus serrata), Japanese zelkova (Zelkova serrata) and other broad-leaved deciduous tree species. Some of the forest is used for plantations of Japanese cedar (Cryptomeria japonica) and cypress (Chamaecyparis obtusa) for timber production. All samples were taken from the study area and identified to the species and genus level, after which they were classified according to the criteria shown in Fig. 1 (see also Table S1). The sampling points for each sample were selected haphazardly within the study area. The samples were collected from 16 May to 27 July 2012, with the exception of two samples for Bibionidae larvae which were collected on 5 September 2012 and three samples for stream algae which were collected on 27 February 2013.
In addition to these collections, litters and soils of the A0 (L, F and H), A1 and A2 layers, respectively, were sampled from 0.09 m2 of forest floor at three sites (sites 1, 2 and 3), which were chosen within the study area; two from secondary deciduous forests (sites 1 and 3) and one from a cedar plantation (site 2). The total amount of 137Cs over a fixed area (1 m2) was calculated from the sum of all layers. The depth of the lower end of the A2 layer was 10 to 12 cm.
All samples were dried for 48 h at 60°C and powdered using a mortar and pestle. For small organisms, several individuals were mixed and powdered together for the measurements. At least 200 mg of samples were gathered and the data on the 137Cs concentrations and nitrogen stable isotope ratio (δ15N) were obtained from the same sample.
Sample analysis
Germanium semiconductor detectors were used for the measurements of the 137Cs concentrations of all samples. Gamma-ray spectrometry was conducted using germanium detectors (Seiko EG&G). An efficiency calibration of the detectors was made with volume radioactivity standard gamma sources (MX0333U8, Japan Radioisotope Association). The standard reference material JSAC-0471 (the Japan Society for Analytical Chemistry) was used for an accuracy check. The measured values were corrected for the sampling day.
Nitrogen stable isotope (δ15N) ratios were measured using SerCon ANCA GSL elemental analyser interfaced to a SerCon Hydra 20–20 continuous flow isotope ratio mass spectrometer. Nitrogen isotopic compositions were normalized by using the N-1 standard ( = 1.36‰) and are reported relative to atmospheric nitrogen29.
Statistical analysis
To determine the best grouping of the functional group in terms of the 137Cs concentration, a generalized linear model (glm) with a normal distribution and log-link function was utilized with the model selection based on Akaike's Information Criterion (AIC). All possible groupings of the combinations of all trophic groups were examined. The model with the lowest AIC value was selected as the preferred grouping30. The effects of trophic levels and the habitat (terrestrial vs. aquatic) on the δ15N values were analysed using glm. The best model explaining the variation of δ15N values was selected based on AIC. The relationship of δ15N value and 137Cs concentrations with examining the effect of the habitat (terrestrial vs. aquatic) were also analysed using glm. The best model explaining the variation of 137Cs concentrations was selected based on AIC. All analyses were performed in R version 2.13.131.
References
Zheng, J. et al. Isotopic evidence of plutonium release into the environment from the Fukushima DNPP accident. Sci. Rep. 2, 304 (2012).
Yasunari, T. J. et al. Cesium-137 deposition and contamination of Japanese soils due to the Fukushima nuclear accident. Proc. Natl. Acad. Sci. USA 108, 19530–19534 (2011).
Masson, O. et al. Tracking of airborne radionuclides from the damaged Fukushima Dai-Ichi nuclear reactors by European networks. Environ. Sci. Technol. 45, 7670–7677 (2011).
Ishii, N. et al. Deposition in Chiba Prefecture, Japan, of Fukushima Daiichi Nuclear Power Plant fallout. Health Physics 104, 189–194 (2013).
Hashimoto, S., Ugawa, S., Nanko, K. & Shichi, K. The total amounts of radioactively contaminated materials in forests in Fukushima, Japan. Sci. Rep. 2, 416–420 (2012).
Ohte, N. et al. Diffusion and export dynamics of 137Cs deposited on the forested area in Fukushima after the nuclear power plant accident in March 2011: Preliminary results. International Symposium on Environmental Monitoring and Dose Estimation of Residents after Accident of TEPCO's Fukushima Daiichi Nuclear Power Station, Kyoto University Research Reactor Institute., Kyoto, 25–32 (2012). <http://www.rri.kyoto-u.ac.jp/anzen_kiban/outcome/Proceedings_for_Web/Topics_1-07.pdf> (accessed 2012.11.26).
Kitchings, T., Digregorio, D. & Van Voris, P. A review of ecological parameters in vertebrate food chains. Radioecology and Energy Resources. Proceedings of the Fourth National Symposium on Radioecology, 304–313 (1976). <http://pdw.hanford.gov/arpir/pdf.cfm?accession=D196018276> (accessed 2012.11.26).
Rowan, D. & Rasmussen, J. B. Bioaccumulation of radiocesium by fish: the influence of physicochemical factors and trophic structure. Can. J. Fish. Aquat. Sci. 51, 2388–2410 (1994).
Wang, W. X., Ke, C., Yu, K. N. & Lam, P. K. S. Modeling radiocesium bioaccumulation in a marine food chain. Marine Ecol. Progress Series 208, 41–50 (2000).
Polis, G. A. & Strong, D. R. Food web complexity and community dynamics. Am. Nat. 141, 813–846 (1996).
Minagawa, M. & Wada, E. Stepwise enrichment of 15N along food chains: further evidence and the relation between 15N and animal age. Geochimica et Cosmochimica Acta 48, 1135–1140 (1984).
Tokyo Electric Power Company, Estimation of the released amount of radioactive materials into the atmosphere as a result of the accident in the Fukushima Daiichi Nuclear Power Station (2012). <http://www.tepco.co.jp/en/press/corp-com/release/betu12_e/images/120524e0201.pdf> (accessed 2012.11.26).
Kato, H., Onda, Y. & Gomi, T. Interception of the Fukushima reactor accident-derived 137Cs, 134Cs and 131I by coniferous forest canopies. Geophys. Res. Lett. 39, L20403 (2012).
Aerts, R. Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: a triangular relationship. Oikos 79, 439–449 (1997).
Miyashita, T., Takada, M. & Shimazaki, A. Experimental evidence that aboveground predators are sustained by underground detritivores. Oikos 103, 31–36 (2003).
Strand, P., Beresford, N. & Avila, R. (eds) Deliverable 1; Identification of Candidate Reference Organisms from a Radiation Exposure Pathways Perspective. A project within the EC 5th Framework Programme (2001). <https://wiki.ceh.ac.uk/download/attachments/115802176/fasset_d1.pdf>(accessed 2012.11.26).
Rudge, S. A., Johnson, M. S., Leah, R. T. & Jones, S. R. Biological transport of radiocaesium in a semi-natural grassland ecosystem. 1. Soils, vegetation and invertebrates. J. Environ. Radioactivity 19, 173–198 (1993).
Battiston, G. A. et al. Radioactivity in mushrooms in northeast Italy following the Chernobyl accident. J. Environ. Radioactivity 9, 53–60 (1989).
Bazala, M. A., Golda, K. & Bystrzejewska-Piotrowska, G. Transport of radiocesium in mycelium and its translocation to fruitbodies of a saprophytic macromycete. J. Environ Radioactivity 99, 1200–1202 (2008).
Eckl, P., Hofmann, W. & Tüurk, R. Uptake of natural and man-made radionuclides by lichens and mushrooms. Radiation Environ. Biophys. 25, 43–54 (1986).
Kalač, P. A review of edible mushroom radioactivity. Food Chemistry 75, 29–35 (2001).
Shibata, H., Mitsuhashi, H., Miyake, Y. & Nakano S. Dissolved and particulate carbon dynamics in a cool-temperate forested basin in northern Japan. Hydrological Process 15, 1817–1828 (2001).
Adam, C., Garnier-Laplace, J. & Baudin, J. P. Bioaccumulation of 110mAg, 60Co, 137Cs and 54Mn by the freshwater crustacean Daphnia magna from dietary sources (Scenedesmus obliquus and Cyclotella meneghiana). Water, Air and Soil Pollution 136, 125–146 (2002).
Nimis, P. L., Bolognini, G. & Giovani, C. Radiocontamination patterns of vascular plants in a forest ecosystem. Forests and Radioactivity 157, 181–188 (1994).
Vinichuk, M., Mårtensson, A., Ericsson, T. & Rosén, K. Effect of arbuscular mycorrhizal (AM) fungi on 137Cs uptake by plants grown on different soils. J. Environ. Radioactivity 115, 151–156 (2013).
Avery, S. V. Fate of cesium in the environment: distribution between the abiotic and biotic components of aquatic and terrestrial ecosystems. J. Environ. Radioactivity 30, 139–171 (1996).
Davis, J. J. Cesium and its relationships to potassium in ecology. in Radioecology (ed. Schultz, V. & Klement, A. W.) 539–556 (Reinhold, New York, 1963).
MEXT (Japanese Ministry of Education, Culture, Sports, Science and Technology) and DOE (the US Department of Energy), Results of Airborne Monitoring by the Ministry of Education, Culture, Sports, Science and Technology and the US Department of Energy. (2011). <http://www.mext.go.jp/component/english/__icsFiles/afieldfile/2011/05/10/1304797_0506.pdf> (accessed 2012.11.26).
Lajtha, K. & Michener, R. H. Stable Isotopes in Ecology and Environmental Science (Blackwell Scientific Publications, Oxford., 1994).
Crawley, M. J. The R Book (John Wiley and Sons, Chichester., 2007).
R Development Core Team, R: A language and Environment for Statistical Computing (R Foundation for Statistical Computing, Vienna, 2011).
Hoover, M. D. & Lunt, H. A. A key for the classification of forest humus types. Soil Sci. Soc. Am. J. 16, 368–370 (1952).
Acknowledgements
This study was supported by a grant (24248027) for scientific research from the Ministry of Education, Culture, Sports, Science and Technology Japan Society for the Promotion of Science.
Author information
Authors and Affiliations
Contributions
M.M. and N.O. designed the study and wrote the manuscript with input from all other authors. T.S. and Y.I. collected samples and performed the calculations. N.I. and K.T. conducted the radioisotope analyses.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Dataset 1
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article
Cite this article
Murakami, M., Ohte, N., Suzuki, T. et al. Biological proliferation of cesium-137 through the detrital food chain in a forest ecosystem in Japan. Sci Rep 4, 3599 (2014). https://doi.org/10.1038/srep03599
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep03599
This article is cited by
-
Radiocesium contaminations and transfer in cyclorrhaphous flies (Diptera: Muscidae, Calliphoridae) at three distances from the Fukushima Dai-ichi Nuclear Power Plant after the 2011 accident
Applied Entomology and Zoology (2022)
-
Fungal spore involvement in the resuspension of radiocaesium in summer
Scientific Reports (2019)
-
Comparison of radioactive and stable cesium uptake in aquatic macrophytes affected by the Fukushima Dai-ichi Nuclear Power Plant accident
Journal of Radioanalytical and Nuclear Chemistry (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
