
Abstract
A valid indicator of central serotonergic neurotransmission would be useful for various diagnostic and psychopharmacological purposes in psychiatry. However, known peripheral serotonergic measures only partially reflect serotonergic function in the brain. Previous findings suggest that the intensity dependence of auditory evoked potentials (AEPs) is closely related to central serotonergic activity. The present study examines the effects of microinjection of a 5-HT1A agonist (8-OH-DPAT) and a 5-HT1A antagonist (spiperone) into the dorsal raphe nucleus (DRN) on AEP recorded epidurally from the primary and secondary auditory cortex in behaving cats. We found a stronger intensity dependence only of AEP from the primary auditory cortex after 8-OH-DPAT, which inhibits the firing rate of serotonergic DRN neurons, and a weaker intensity dependence after spiperone, which increases serotonergic cell firing, as compared to baseline measurements. These results demonstrate that the intensity dependence of AEP is inversely related to serotonergic neuronal activity and that it may be a promising tool for assessing central serotonergic function in humans (e.g., identifying patients with low serotonergic neurotransmission).
Similar content being viewed by others
Main
Disturbed activity of the serotonergic system is assumed to play a major role in such psychiatric disorders as depression, obsessive compulsive and anxiety disorders, as well as alcoholism and schizophrenia (Siever et al. 1991; Murphy et al. 1998). Several new antidepressants and antipsychotics are specifically designed to influence central serotonergic neurotransmission (Cowen 1991). A valid indicator of serotonergic function in the brain could be used to identify psychiatric patients with a serotonergic dysfunction in order to treat them more specifically. However, peripheral measures of serotonin levels or its metabolites only partially reflect central serotonergic activity (Murphy 1990). Several findings from basic and clinical research suggest that a strong stimulus intensity dependence of auditory evoked potentials (AEPs), that is the increase of AEP amplitudes attributable to increasing tone intensity (loudness), is related to low serotonergic activity, and vice versa. The intensity dependence is reduced by serotonin enhancing substances (zimelidine, lithium, ethanol, methamphetamines); this variable predicts the response to serotonin agonists and is related to serotonin-associated personality traits and behavior (sensation seeking, impulsivity, suicidality, antisocial/aggressive behavior), as well as, to 5-HIAA levels in cerebrospinal fluid (Hegerl and Juckel 1993).
Recently, we established an animal model of the intensity dependence in behaving cats (Juckel et al. 1996). We found that the intensity dependence of the cat AEP first positive component with the highest functional similarity to that of human AEP was enhanced by the systemic administration of the serotonin antagonist ketanserin and reduced by the serotonin agonist 8-OH-DPAT, which both may be a postsynaptic effects (Juckel et al. 1997). These changes were found only for the intensity dependence of the primary auditory cortex, which corresponds to the known higher innervation of the primary, rather than the secondary auditory cortex by serotonergic fibers (Lewis et al. 1986). To investigate the relationship between the intensity dependence of AEP and the (presynaptic) activity of serotonergic neurons itself, the somatodendritic 5-HT1A autoreceptors of the serotonergic neurons were influenced by local application of 8-OH-DPAT and spiperone into the dorsal raphe nucleus (DRN). The 5-HT1A agonist 8-OH-DPAT strongly suppresses the firing rate of serotonergic neurons in the cat DRN, while the 5-HT1A antagonist spiperone enhances the firing rate of these neurons (Fornal et al. 1994).
METHODS
The intensity dependence of AEP was recorded epidurally over the primary and secondary auditory cortex (AI, AII) in five adult male freely moving cats, weighting 2.8 to 3.5 kg. The cats were cared for and used in accordance to the Guide for the Care and Use of Laboratory Animals. As recording electrodes, 0.23 mm diameter enamel-coated stainless steel wires, etched for 1 mm at their tips, were used; a stainless steel screw above the bone of the frontal sinus served as reference electrode. The auditory stimuli were 4-kHz tones (sine wave bursts) of 5 ms duration, at four intensity levels (50, 60, 70, 80 dB peak equivalent SPL), and were presented in random order through a tiny bone conductor fixed to the animals’ foreheads (randomized ISI of 1.8–2.2 s; 42 stimuli of each intensity). Electrophysiological data were collected with a sampling rate of 1 kHz in a time range of 50 ms prestimulus to 200 ms poststimulus, and they were corrected for the prestimulus basline, as well as for artifacts. After averaging, slopes of all possible connections between the four amplitude values of the AEP first positive components were calculated for each recording of each cat separately. The median of these slopes (n = 8) was taken as the intensity dependence [μV/10 dB] of the AEP first positive component, recorded from the primary and secondary auditory cortex. A detailed description of the AEP method used is given elsewhere (Juckel et al. 1996, 1997). A 22-gauge stainless steel guide cannula was implanted through the cerebellum at an angle of 40 to 50°. Through this guide cannula, a 5 μl Hamilton syringe (26-gauge) was inserted, so that its tip was in the DRN (stereotaxic coordinates: AP −0.5, L 0.0, V −0.5) (Jacobs et al. 1984). ± 8-OH-DPAT HBr [2 μg/1 μl saline (Sharp et al. 1989)], spiperone (2 μg/1 μl saline), and saline (1 μl, 0.9%) were injected on different occasions separated by at least 48 hours. Injections were done over 1 minute, and the syringe was let in place for another minute to allow the solution to diffuse away from the site. Experiments were performed at the same time in each animal in the morning hours (8:00–10:00 A.M.). AEP recordings were immediately obtained before (baseline) and starting 2, 10, and 30 minutes after injection, while cats were in a quiet waking state. The quiet waking state was defined by usual criteria: cats were sitting in the experimental cage with no apparent sign of noticing their enviroment. Electrophysiologically, this state was characterized by a low-voltage, high-frequency cortical EEG, theta activity of the dorsal hippocampus, and frequent eye movements in the EOG. The localization of the injection sites within the DRN of four cats studied (marked by 1 μl methylblue at the end of the experiments) was verified histologically (Figure 1). In one cat, histology revealed that the tip of the syringe was located shortly behind the cerebellum in the cerebral aqueduct during drug injections. Data were statistically analyzed using analysis of variance (ANOVA) with repeated measures, followed by post hoc comparisons (Student-Newman-Keuls’ test).

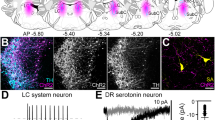
The localization of the injection sites in four animals studied. (P 1.0–P 2.5: the AP stereotaxic positions of the slides, according to Snider and Niemer 1961)
RESULTS
Following injection of either 8-OH-DPAT, spiperone, or saline, all cats remained in the quiet waking state throughout the experiments and showed no apparent behavioral changes. The intensity dependence of the AEP first positive component recorded from the primary auditory cortex was significantly increased 2 minutes after injection of 8-OH-DPAT into the DRN (means ± SEM: 5.48 ± 2.21 vs. 7.15 ± 2.33 μV/10 dB, F(3/9) = 4.06, p < .05; Figure 2); whereas it was significantly decreased 2 minutes after injection of spiperone (8.88 ± 3.03 vs. 7.33 ± 3.26 μV/10 dB, F(3/9) = 3.91, p < .05). The intensity dependence of each cat injected into the DRN showed the same direction of change within the 15-minute pre–post period after injection of 8-OH-DPAT and of spiperone. Original AEP recordings of one cat and their changes after drug application are shown in Figure 3. In the 10 and 30 min recordings after 8-OH-DPAT and spiperone, the intensity dependence tended to return to the baseline values. Control saline injections had no effect on the intensity dependence of the AEP first positive component recorded from the primary auditory cortex (F(3/9) = 1.09, ns). The intensity dependence of the AEP first positive component recorded from the secondary auditory cortex was not significantly altered by the injection of either 8-OH-DPAT or spiperone (F(3/9) = 0.67 and F(3/9) = 0.93, ns; Figure 4). In that cat, in which 8-OH-DAPT, spiperone, and saline were injected into the ventricular system, there was no significant change of the intensity dependence of the AEP first positive component recorded from the primary auditory cortex (before and after 8-OH-DPAT: 12.7, 12.0, 12.5, 12.8, 12.9 μV/10 dB; before and after spiperone: 13.5, 13.7, 13.6, 14.2, 14.0 μV/10 dB; before and after saline: 13.3, 13.0, 13.0, 13.4, 13.1 μV/10 dB).

Intensity dependence of AEP first positive component, recorded epidurally from the primary auditory cortex, before (baseline) as well as 2, 10, and 30 minutes after injection of 8-OH-DPAT, spiperone, and saline into the DRN of the four cats. *p < .05, as compared to the baseline recordings; Student-Newman-Keuls’ multiple comparison test.

Original AEP recordings from the primary auditory cortex of one cat before and 2 min after injection of 8-OH-DPAT or spiperone into the DRN. The AEP first positive component is the first deflection to the bottom after auditory stimulation, which is indicated as y-axis.

Intensity dependence of AEP first positive component, recorded epidurally from the secondary auditory cortex, before (baseline) as well as 2, 10, and 30 minutes after injection of 8-OH-DPAT, spiperone, and saline into the DRN of the four cats
DISCUSSION
The present study is the first to demonstrate an inverse relationship between the intensity dependence of AEP, recorded from the primary, but not of the secondary, auditory cortex, and brain serotonergic neuronal activity in behaving cats. The inactivation of serotonergic DRN neurons by local application of the 5-HT1A agonist 8-OH-DPAT into the DRN increased the intensity dependence of AEP. Reciprocally, the activation of serotonergic DRN neurons by local administration of the 5-HT1A antagonist spiperone decreased the intensity dependence of AEP. Studies have shown that the primary auditory cortex, particularly layer IV, which receives most of the specific thalamic sensory input, is one of the most highly innervated cortical areas by serotonergic fibers (Jacobs and Azmitia 1992; Lewis et al. 1986). Innervation of cortical layer IV is mostly derived from the serotonergic DRN neurons (Wilson and Molliver 1991). This nucleus, well known as main source of serotonergic efferents to the forebrain, is, therefore, in an excellent position to modulate neuronal activity at the initial stage of sensory processing in the primary auditory cortex. This modulation of the sensory function by serotonergic fibers from the raphe nuclei seems be mostly inhibitory. Electrical stimulation of the raphe nuclei or microiontophoretic application of serotonin into the cortex were found to lead to a decrease of amplitudes of sensory evoked potentials as well as of evoked responses of cortical neurons (Samanin et al. 1972; Mantz et al. 1990; Waterhouse et al. 1990). The inhibitory function of the serotonergic system is also supported by the findings of the present study, showing a decrease of the intensity dependence from the primary auditory cortex, as a measure of cortical reagibility to tones of different loudness, after enhancement of serotonergic activity. Thus, the serotonergic systems is possibly aimed to protect the cortex from sensory overstimulation.
However, three major concerns related to the results of the study has to be addressed. First is the variability of the baseline values of the intensity dependence. Such variability was observed only over a time period of several days or weeks. This variability was also seen in previous studies (Juckel et al. 1996, 1997), and could be caused by technical reasons concerning the recording electrode; for example, small changes of the electrode environment over longer time. The changes of the intensity dependence we found after drug application are very unlikely to be time effects within the experimental period of 45 minutes, because the intensity dependence of the AEP first positive component did not intraindividually change after injection of saline. In addition, the intensity dependence recorded simultaneously from the secondary auditory cortex did not change after the injection of 8-OH-DPAT, spiperone, and saline into the DRN. Thus, changes of the intensity dependence recorded from the primary auditory cortex, found in all of the cats, may be caused by the drug effects on the firing rate of DRN serotonergic neurons.
Concerning histology, injection sites outside the DRN as well as tissue damage of the DRN could be factors influencing the results. We found, however, the injection sites of all animals showing significant changes of the intensity dependence after drug application in that area where the DRN is usually located. There was no hint for any significant damage of the DRN tissue, indicating quite normal functioning of the DRN serotonergic cells during the experiments. Given the size of the DRN with 3 to 4 mm in the rostral-caudal plane, it seems very likely that most of the drug solution had exerted its influence only within the DRN. It is also likely that the effects seen with the intensity dependence of the primary auditory cortex after drug application into the DRN were mediated by the somatodentritic 5-HT1A autoreceptors within the DRN, because 5-HT1A receptors are present only on the cell membranes of the DRN serotonergic neurons in this anatomic region (Sotelo et al.1990). Because there was no significant change of the intensity dependence in the cat, in which the drugs were applied into the ventricular system, the changes of the intensity dependence from the primary auditory cortex reported here seem to be site specific for the DRN than the result of any leakage of the injectate into the ventricular system.
Finally, the relationship between the intensity dependence and the serotonergic system seems to be not specific in an exclusive manner. Although the noradrenergic and dopaminergic neurotransmission probably have no direct influence on the intensity dependence of AEP first positive component, atropine administered intraveneously in cats exhibited a clear effect on the intensity dependence of the primary auditory cortex (Juckel et al. 1997). Thus, it is assumed that the serotonergic and cholinergic system interact synergistically in the modulation of the intensity dependence, but an important part of the variance of the intensity dependence may be explained by the serotonergic neurotransmission alone.
In humans, activity from the primary and secondary auditory cortex can be separated by dipole source analysis (Scherg and Von Cramon 1990). Using this method, it was shown that only the intensity dependence of the tangential dipole, reflecting activity of the primary auditory cortex, is related to serotonergic measures (Hegerl and Juckel 1993). Although the extrapolation of the present results in cats to humans must be made cautiously, and the specificity might be limited, some evidence suggests that the intensity dependence of AEP is of clinical value in assessing central serotonergic function. For example, several studies have revealed that patients with low serotonergic activity, indicated by a strong intensity dependence before treatment, respond more favorably to drugs that increase serotonergic neurotransmission, such as fluoxetine (Paige et al. 1994), fluvoxamine (Hegerl and Juckel 1993), fenfluramine (Bruneau et al. 1989), paroxetine (Hegerl et al. 1996), or lithium (Hegerl and Juckel 1993) than patients with normal or high serotonergic activity, as indicated by a weak intensity dependence of AEP. Regarding the fact that brain serotonin cannot be measured directly and that no valid biochemical indicator of the central serotonergic function is available, the finding of a close relationship between the intensity dependence recorded from the primary auditory cortex and central serotonergic activity is of great clinical interest. Thus, the intensity dependence could, at the least, be used as a gross index of the amount of serotonin released into the synapses; this is information that cannot be provided by any other method known today.
References
Bruneau N, Barthelmy C, Roux S, Jouve J, Lelord G . (1989): Auditory evoked potential modifications according to clinical and biochemical responsiveness to fenfluramine treatment in children with autistic behavior. Neuropsychobiology 21: 48–52
Cowen PJ . (1991): Serotonin receptor subtypes: Implications for psychopharmacology. Br J Psychiat 159: 7–14
Fornal CA, Litto WJ, Metzler CW, Marrosu F, Tada K, Jacobs BL . (1994): Single-unit responses of serotonergic dorsal raphe neurons to 5-HT1a agonist and antagonist drug administration in behaving cats. J Pharmacol Exp Ther 270: 1359–1366
Hegerl U, Juckel G . (1993): Intensity dependence of auditory evoked potentials as an indicator of central serotonergic neurotransmission: A new hypothesis. Biol Psychiat 33: 173–187
Hegerl U, Gallinat J, Juckel G, Bauch T, Munke A, Bottlender R, Möller HJ . (1996): ERP as indicator of serotonergic dysfunction and predictor of clinical response to serotonin agonists. Electroencephal Clin Neurophysiol 99: 395 (Abstract)
Jacobs BL, Azmitia EC . (1992): Structure and function of the brain serotonin system. Physiol Rev 72: 165–229
Jacobs BL, Gannon PJ, Azmitia EC . (1984): Atlas of serotonergic cell bodies in the cat brainstem: An immunocytochemical analysis. Brain Res Bull 13: 1–31
Juckel G, Csépe V, Molnár M, Hegerl U, Karmos G . (1996): Intensity dependence of auditory evoked potentials in behaving cats. Electroencephal Clin Neurophysiol 100: 527–537
Juckel G, Molnár M, Hegerl U, Csépe V, Karmos G . (1997): Auditory evoked potentials as indicator of brain serotonergic activity — First evidence in behaving cats. Biol Psychiat 41: 1181–1195
Lewis DA, Campbell MJ, Foote SL, Morrison JH . (1986): The monaminergic innervation of primate neocortex. Human Neuobiol 5: 181–188
Mantz J, Godbout R, Tassin JP, Glowinski J, Thierry AM . (1990): Inhibition of spontaneous and evoked unit activity in the rat medial prefrontal cortex by mesencephalic raphe nuclei. Brain Res 524: 22–30
Murphy DL . (1990): Peripheral indices of central serotonin function in human. Ann NY Acad Sci 600: 282–296
Murphy DL, Andrews AM, Wichems CH, Li Q, Tohda M, Greenberg B . (1998): Brain serotonin neurotransmission: An overview and update with an emphasis on serotonin subsystem heterogeneity, multiple receptors, interaction with other neurotransmitter systems, and consequent implications for understanding the actions of serotonergic drugs. J Clin Psychiat 59: 4–12
Paige SR, Fitzpatrik DF, Kline JP, Balogh SE, Hendricks SE . (1994): Event-related potential (ERP) amplitude/intensity slopes predict response to antidepressants. Neuropsychobiology 30: 197–201
Samanin R, Valzelli L, Gumulka W . (1972): Inhibitory effect of midbrain raphe stimulation on cortical evoked potentials in rats. Psychopharmacology 24: 373–379
Scherg M, Von Cramon D . (1990): Dipole source potentials of the auditory cortex in normal subjects and in patients with temporal lobe lesions. Adv Audiol 6: 165–193
Sharp T, Bramwell SR, Clark D, Grahame-Smith DG . (1989): In vivo measurement of extracellular 5-hydroxytryptamine in hippocampus of the anesthetized rat using microdialysis: Changes in relation to 5-hydroxytryptaminergic neuronal activity. J Neurochem 53: 234–240
Siever LJ, Kahn RS, Lawlor BA, Trestman RL, Lawrence TL, Coccaro EF . (1991): Critical issues in defining the role of serotonin in psychiatric disorders. Pharmacol Rev 43: 509–525
Snider RS, Niemer WT . (1961): A Stereotaxic Atlas of the Cat Brain. Chicago, The University of Chicago Press
Sotelo C, Cholley B, El Mestikawyn S, Gozlan H, Hamon M . (1990): Direct immunohistochemical evidence of the existence of 5-HT1A autoreceptors on serotoninergic neurons in the midbrain raphe nuclei. Eur J Neurosci 2: 1144–1154
Waterhouse BD, Azizi SA, Burne RA, Woodward DJ . (1990): Modulation of rat cortical area 17 neuronal responses to moving visual stimuli during norepinephrine and serotonin microiontophoresis. Brain Res 514: 276–292
Wilson MA, Molliver ME . (1991): The organization of serotonergic projections to cerebral cortex in primates: Regional distribution of axon terminals. Neuroscience 44: 537–553
Acknowledgements
We gratefully acknowledge the encouragement and advice of Barry L. Jacobs and Casimir A. Fornal (Princeton). This study was supported by the Deutsche Forschungsgemeinschaft (He 1680/3, He 916/7), the Hungarian Research Found (OTKA I/3-2595) and the Upjohn Company.
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Juckel, G., Hegerl, U., Molnár, M. et al. Auditory Evoked Potentials Reflect Serotonergic Neuronal Activity—A Study in Behaving Cats Administered Drugs Acting on 5-HT1A Autoreceptors in the Dorsal Raphe Nucleus. Neuropsychopharmacol 21, 710–716 (1999). https://doi.org/10.1016/S0893-133X(99)00074-3
Received:
Revised:
Accepted:
Issue Date:
DOI: https://doi.org/10.1016/S0893-133X(99)00074-3
Keywords
This article is cited by
-
Neurophysiological investigation of auditory intensity dependence in patients with Parkinson’s disease
Journal of Neural Transmission (2021)
-
Effects of intensity, attention and medication on auditory-evoked potentials in patients with fibromyalgia
Scientific Reports (2020)
-
Tobacco use is associated with reduced amplitude and intensity dependence of the cortical auditory evoked N1-P2 component
Psychopharmacology (2016)
-
Altered processing of sensory stimuli in patients with migraine
Nature Reviews Neurology (2014)
-
Loudness dependence of auditory evoked potentials (LDAEP) in clinical monitoring of suicidal patients with major depression: a pilot study
European Archives of Psychiatry and Clinical Neuroscience (2012)
