
Abstract
Wheat-legume temporary intercropping with legume devitalization in late winter can increase the N self-sufficiency of cropping systems and improve the N nutrition of wheat as a cash crop. However, this practice has been scarcely investigated. In this study, carried out in a Mediterranean environment with cold winters, we compared 14 treatments over 3 years with different weather patterns: (A) pure stands of common wheat (Triticum aestivum L.) grown at 0, 40, 80, 120, 160 kg N ha−1; (B) pure stands of faba bean (Vicia faba L. minor), pea (Pisum sativum L.), and squarrose clover (Trifolium squarrosum L.); (C) temporary intercrops of wheat and faba bean, pea, or clover; and (D) permanent intercrops of wheat and the same legumes. In the temporary intercrops, all legumes improved the wheat N availability compared with the unfertilized control. The “N effect” of legumes for wheat was consistent across years and proportional to the competitive ability of the legumes, i.e., faba bean > pea > clover. The higher the legume competitive ability, the higher was its N accumulation before devitalization, and the higher/earlier was the N supplied to wheat. Our findings demonstrate that the date of legume devitalization represents a key factor to be managed each year in order to maximize legume N supply while preventing excessive legume competition, which could undermine wheat growth and yield. Such a modulation was not possible in permanent intercrops, where legume competition depressed wheat grain yield. The N supplied with legume devitalization increased wheat grain N accumulation during grain filling. This is the first work comparing temporary and permanent intercrops of wheat with different legumes and over different seasons. Overall, temporary intercropping appears to be a feasible and efficient tool for the sustainable management of N nutrition in winter wheat.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
N nutrition in winter wheat in the Mediterranean environment is frequently hampered in fall-winter by either low temperature, which slows the soil organic matter mineralization, or high rainfall, which causes soil N depletion via both NO3 leaching and N volatilization (Tosti et al. 2014; Volpi et al. 2018). Such a N shortage limits wheat growth, mainly from tillering to stem elongation (Ferrise et al. 2015), compromising yield (Benincasa et al. 2016). Mineral N fertilization using split top-dress applications represents the traditional form of N management in conventional wheat (Blandino et al. 2015), as it meets crop N requirements while reducing the risk of high N loss. In organic wheat, splitting the rate is generally not feasible because most organic fertilizers need to be broadcast all at once before sowing, and this results in an insufficient N supply to wheat, which is detrimental for plant growth and yield formation. However, even in the case of top-dress application, the N availability from organic fertilizers in winter may be low and late (Tosti et al. 2016), resulting in a limited yield, mainly in terms of a low number of grains per unit area (Benincasa et al. 2017a). In addition, the low winter N accumulation in vegetative organs predisposes the grain protein content to be below the target values required for baking and pasta (Giuliani et al. 2015). The management of soil N fertility based only on extrafarm fertilizers (both mineral and organic) seems not to be sustainable over the long term (Thorup-Kristensen et al. 2012). For this reason, strategies are needed to increase (i) N self-sufficiency and (ii) N use efficiency at the plant, crop, and whole-rotation scales (Benincasa et al. 2017b; Dresbøll and Thorup-Kristensen 2014). Among these strategies, cereal-legume intercropping appears to be a useful agronomic solution for wheat production (Bedoussac and Justes 2010).
It is widely accepted that cereal-legume permanent intercropping (PIC), i.e., growing species together from establishment to final harvest, can provide good N nutrition for the companion cereal (Bedoussac et al. 2015). Unfortunately, this is partly counterbalanced by competition from the legume toward the cereal for resources other than N (Verret et al. 2017). In addition, PIC implies technical management issues (e.g., weed and disease control, mechanization of cultivation practices), which need to be addressed in order to promote the adoption of this practice (Brooker et al. 2015). Thus, PIC appears mainly suitable for feed, where it provides a double harvest with increased protein yield.
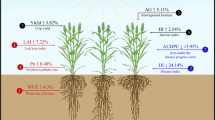
An alternative technique to PIC is represented by temporary intercropping (TIC, Fig. 1), where the cereal and the legume are sown in alternate rows and spaced sufficiently to allow legume devitalization and incorporation into the soil at the beginning of cereal stem elongation (Tosti and Guiducci 2010). In this case, the cereal is intended to be the sole high-value cash crop, devoted to the bakery or pasta industry, in which high protein content is required. The legume devitalization at the beginning of stem elongation is aimed toward preventing late interspecific competition and providing N release from the incorporated biomass, which works similarly to side-dress organic fertilization for the cereal (e.g., the green manure effect). Previous research on durum wheat-faba bean TIC indicated that the N released from the incorporation of faba bean was low (48 ± 3 kg N ha−1 on average) and not timely enough to increase wheat yield, whereas the grain protein content was much increased (Tosti and Guiducci 2010). However, the results are likely to depend on the environmental conditions, legume species, and TIC management. Compared to companion legumes, wheat is advantaged by high soil mineral N (Nmin) at sowing and freezing temperatures in winter, which further depress legume growth. As far as TIC is concerned, the green manuring effect of legumes depends on their growth rate and N fixation during the growing season. In particular, slow growth in late winter does not match the need for early devitalization. Several legumes have been studied in terms of their use in intercropping with wheat (Alonso-Ayuso et al. 2014; Costanzo and Bàrberi 2014). Among these, faba bean (Vicia faba L. minor), pea (Pisum sativum L.), and squarrose clover (Trifolium squarrosum L.) are three of the main green manure crops used in the Mediterranean environment (Benincasa et al. 2010) but not for intercropping with wheat.

Wheat-legume temporary intercropping with legume devitalization in late winter-early spring. The devitalization date varied year by year with the legume species according to its growth stage in order to modulate the competition toward wheat. The legume biomass incorporation into the soil increased wheat nitrogen status and grain nitrogen content
Faba bean has been demonstrated to perform very well in the Mediterranean environment, with a good growth rate in winter (provided frost-tolerant cultivars are used), which guarantees high N accumulation and weed suppression thanks to its tall size (Benincasa et al. 2010). Tosti and Guiducci (2010) intercropped wheat with faba bean and observed an overall positive outcome in terms of both wheat yield and protein content. In a greenhouse experiment, Benincasa et al. (2012) studied early interference between wheat and faba bean and found that the competitive ability of both species is strictly related to their seeding density and that this interaction can be quite easily managed. However, when the interaction is not properly managed, faba bean can take advantage on wheat, especially in rainy years (Mariotti et al. 2015).
Pea has good frost resistance and is known to have a growing cycle that perfectly lines up to that of wheat (Annicchiarico and Iannucci 2007). Moreover, the complementarity between these species in terms of canopy architecture has been shown to produce an excellent smothering effect on weeds (Verret et al. 2017).
Squarrose clover has been widely investigated in several Mediterranean areas; it has good N accumulation potential, and its growing cycle is similar to that of wheat, but very few studies on wheat-clover intercropping are available (Saia et al. 2016). Compared to the former two species, clover appears to be less competitive due to its slow initial growth rate and its slow growth recovery after winter (Hekneby et al. 2006).
The variability of outcomes reported for these species in the literature clearly suggests that a meaningful picture can be drawn only by considering different locations and conducting trials repeated in multiple years (i.e., with different seasonal weather).
Finally, although a substitutional approach (i.e., intercropping vs. mineral fertilization) is simplistic, including pure wheat plots fertilized with mineral N in intercropping trials may help with the determination of the timing of N provisioning to wheat from legume biomass. Moreover, this approach may offer a tangible reference for farmers who consider moving toward such an innovative technique.
Hence, the aim of this 3-year study was to evaluate the performance of wheat under temporary and permanent intercropping with three different legume species and including pure stands of the legumes and pure stands of wheat fertilized with increasing mineral fertilizer N rates as controls.
2 Materials and methods
2.1 Experimental site
The field experiment was carried out in three consecutive years (2009/10, 2010/11, and 2011/12) at the experimental station (FIELDLAB) of the Dept. of Agricultural, Food, and Environmental Sciences of the University of Perugia, Italy. The FIELDLAB is located in the Tiber River alluvial plain at 42.956°N, 12.376°E, 163 m asl. The soil is a typical Fluventic Haplustept clay-loam soil (20% sand, 46% silt, and 34% clay, 1.4 Mg m−3 bulk density), subalkaline (pH = 7.8), poor in organic matter (12 g om kg−1, C/N ratio = 11) and extractable phosphorus (29.9 mg P2O5 kg−1, Olsen method), and rich in exchangeable potassium (258 mg K2O kg−1, ammonium-acetate method).
In each year, the previous crop had been sunflower. The soil was fertilized with 90 kg P2O5 ha−1 as superphosphate and 90 kg K2O ha−1 as potassium sulfate just after the sunflower harvest.
Seed bed preparation consisted of shallow plowing (0.25 m depth) followed by harrowing. The mineral N content in the 0–0.90 m soil layer at sowing was 72, 56, and 117 kg N ha−1 in the three experimental years, respectively. Weather data were collected by an automatic meteorological station within the FIELDLAB.
2.2 Treatments and crop management
A total of 14 treatments per year were compared using a randomized block design with three replicates (plot size = 54 m2). The treatments included (A) pure stands of soft wheat (Triticum aestivum L., cv Aubusson) at five increasing N fertilization rates, i.e., 0 (N0), 40 (N40), 80 (N80), 120 (N120), and 160 kg N ha−1 (N160), as ammonium nitrate, split ½ at tillering and ½ at the beginning of stem elongation; (B) pure stands of faba bean (PURF, Vicia faba L. minor, cv Torre lama scuro), pea (PURP, Pisum sativum L, cv Corallo, dwarf, wild type), and squarrose clover (PURC, Trifolium squarrosum L.); (C) temporary intercrops (TICs) of wheat and (i) faba bean (TICF), (ii) pea (TICP), and (iii) squarrose clover (TICC); (D) permanent intercrops (PICs) of wheat and the same leguminous species (PICF, PICP, PICC). Both PIC and TIC were based on the additive principle (i.e., the same sowing densities were adopted in mixture and in pure crops; thus, each species in the intercropping treatments had half the available space as that of pure stands). Wheat and legumes were simultaneously sown on 30 October 2009, 15 November 2010, and 03 November 2011 with wide row spacing (0.375 m apart) using a plot driller, with sowing densities of 400 kernels m−2 for wheat, 80 seeds m−2 for faba bean, 100 seeds m−2 for pea, and 900 seeds m−2 for clover. In the intercropping treatments, the legumes were sown in single rows in the middle of the wheat inter-row space.
In each year, plant emergence regularly occurred approximately 2 weeks after sowing, and the initial plant density was > 90% of the target density in all species.
In the PIC treatments, wheat and legumes grew together until the maturity of both species, while in the TIC treatments, legume plants were killed in spring and incorporated into the topsoil (≈ 0.15 m depth) using split rotary hoeing. No visible damage was caused to wheat plants from this operation.
Legume devitalization in the TIC treatments was planned in order to maximize the N accumulation of the legume component and keep the competitive interaction with wheat to a minimum. Therefore, the legume devitalization date varied on the basis of the legume biomass in relation to the wheat growth stage as follows: (i) at the beginning of wheat stem elongation (Zadoks 32; 23 March 2010, 05 April 2011, 28 March 2012) in TICF; (ii) during mid wheat stem elongation (Zadoks 33; 31 March 2010, 12 April 2011, and 04 March 2012) in TICP; and (iii) at the end of stem elongation (Zadoks 39; 15 April 2010) or heading (Zadoks 59; 09 May 2011, and 08 May 2012) in TICC.
The fields hosting the experiment presented very low volunteer plant pressure, and the crop was therefore easily maintained by hand to be free of weeds. At maturity, the legumes in pure stands and in the PIC treatments were harvested by hand just before the wheat harvest and threshed in the laboratory. Wheat was harvested on 02 July 2010, 23 June 2011, and 23 June 2012 using a plot combine harvester.
2.3 Plant sampling and N analysis
The aboveground biomass and N accumulation of wheat and legumes were determined on the legume devitalization dates at wheat heading (Zadoks 59; 15 May 2010, 10 May 2011, 10 May 2012) and at wheat maturity (02 July 2010, 24 June 2011, 24 June 2012). On each sampling date, the aboveground biomass of wheat and legumes was harvested by hand at the ground level from an area of 1.35 m2. At maturity, the vegetative parts and grains of wheat were kept separate to determine the dry matter accumulation in straw and grains. All samples were oven dried at 85 °C until they reached a constant weight and then ground to a fine powder for the analysis of N content. The N content was measured with Kjeldahl digests using an autoanalyzer (FlowSys Systea, Italy).
The performance in the intercrop and interspecific competition treatments was estimated using the land equivalent ratio (LER; Willey and Osiru 1972) calculated for the legume devitalization dates on a total aboveground N accumulation basis, as follows:
where NWintercrop and NLEGintercrop represent the N accumulation in the intercropped wheat and legumes, respectively, while NWpure and NLEGpure represent the N accumulation in the pure, unfertilized stands, respectively.
2.4 SPAD and wheat N nutritional status
SPAD readings were carried out using wheat leaves from early stem elongation to the beginning of leaf senescence using a SPAD 502 (Minolta Ltd., Osaka, Japan). On each sampling date, measurements were performed in the middle part of the leaf blade (excluding the midvein) of 10 leaves per plot that were randomly chosen among the main culms. Measurements were taken on the youngest fully expanded leaves during stem elongation and on the flag leaves from the booting stage onwards.
At wheat heading, SPAD measurements were also carried out on 10 additional flag leaves per plot. The leaf blades were then harvested, and their area was immediately measured using an automatic area meter (Delta-T Devices Ltd., Cambridge, UK). The leaf blades from each plot were then pooled together, oven dried (at 85 °C until a constant weight was reached), weighed, and ground for Kjeldahl N analysis in order to calculate the average specific leaf N weight (SLNW, g N m−2 leaf area).
2.5 Data analysis
Analysis of variance (ANOVA) was performed according to a randomized block design with three replicates using the software R (R Core Team 2014). Due to the lack of variance homogeneity across years, each year was analyzed separately. The pooled standard error of the mean (SEM) was used as a variability index within each year.
3 Results and discussion
3.1 Weather and implications for soil mineral N content
In the first two growing seasons, the mean air temperatures were in line with the local 30-year average, while the fall-winter was colder in the third year (further details are available in Tosti et al. 2016). Rainfall varied greatly from year to year in terms of both total amount and distribution: years 2009/10r and 2010/11 were both rainy, with an overall amount (from November to June) of 713 and 627 mm, respectively, while only 341 mm of rainfall was recorded in 2011/12.
In 2009/10, rainfall was abundant during wheat tillering (240 mm, January and February) and the beginning of grain filling (108 mm, May), while in 2010/11, 54% of the total rainfall occurred just after sowing (335 mm, November and December), and then another 18% fell in March (113 mm). The third year was unusually dry in fall-winter: from November to March, 138 mm of rain was recorded, accounting for less than one half of the cumulated ET0 (i.e., rain < 0.5 ET0); it was rainy in April (108 mm, rain > 1 ET0) and again very dry in May and June (94 mm), except for 70 mm that fell in the third week of May, just at the beginning of wheat grain filling.
As a consequence, deep drainage was abundant in the first 2 years (251 and 225 mm in 2009/10 and 2010/11, respectively) and none in 2011/12. This affected the soil mineral N availability, which was low in the first 2 years and quite high in the third. Further details on deep drainage and soil mineral N content can be found in Tosti et al. (2016).
3.2 N accumulation associated with legume devitalization
The N accumulation of unfertilized pure wheat varied across years according to the mineral soil N availability at sowing and the weather conditions during the growth cycle (Table 1).
As far as the intercrops are concerned, data on N accumulation recorded on the legume devitalization dates were the same in the TIC and PIC treatments, and thus the mean value of intercrops (ICs) is reported in Fig. 2.

N accumulation (kg ha−1) in the aboveground biomass of wheat and legumes in intercrops (ICs) with faba bean (ICF), pea (ICP), and clover (ICC) on the date of legume devitalization. Numbers within columns indicate partial LER values for wheat (calculated with respect to the N accumulation in unfertilized pure wheat) and legumes (calculated with respect to the N accumulation in pure legumes)
The legume N accumulation in intercrops was markedly reduced compared to that in pure stands (i.e., the legume partial LER was much lower than 1), except in the case of faba bean in 2009/10, which showed an LER value (0.92) that was not significantly different from 1 (Fig. 2). However, the legume LER was still higher than 0.5 (i.e., the expected value when intraspecific competition equals interspecific competition in two-species mixtures) in all years for ICF and in 2 out of 3 years for ICP; therefore, in these cases, the interaction between the legumes and wheat was neutral. These results are in line with findings by Agegnehu et al. (2008) for faba bean and Monti et al. (2016) for pea. On the other hand, although clover was always the last leguminous species to be devitalized, clover N accumulation was very low (particularly in 2010/11), with a partial LER that was always lower than 0.3 (Fig. 2). Thus, clover plants were the most affected by wheat competition. This is because clover has a low plant height (Cornelissen et al. 2003) and a slow resumption of growth if late winter temperatures remain low, as frequently happens in our inner region of central Italy, which differs from the coastal and southern zones, where clover performance is generally very good (Annicchiarico and Pagnotta 2012).
The low legume N accumulation on the devitalization date mainly occurred due to low aboveground biomass accumulation (data not shown) and a lower N concentration compared to that observed in the pure legumes, namely, 2.6% ICC vs 3.2% PURC (SEM = 0.09) in 2009/10, 2.0% ICP vs 3.4% PURP (SEM = 0.17) and 2.2% ICC vs 2.7% PURC (SEM = 0.14) in 2010/11 and 3.9% ICF vs 4.6% PURF (SEM = 0.12) in 2011/12.
Intercropping slightly affected the wheat aboveground N accumulation, which never significantly differed from that in the unfertilized pure wheat. The highest differences in terms of N accumulation between intercropped and control (N0) wheat were 28.1 vs. 20.2 kg (± 3.55) N ha−1 in ICF (2009/10), 61.5 vs. 68.9 kg (± 5.44) N ha−1 in ICP (2011/12, and 33.6 vs. 42.2 kg (± 7.97) N ha−1 in ICC (2009/10) across the three legumes during the three experimental years (pooled SEM in brackets). The partial LER of wheat was always higher than that of legumes and never significantly different from 1. These results are in line with findings by Thilakarathna et al. (2016) and would confirm the facilitation via rhizosphere interactions observed in faba bean-wheat intercropping by Wang et al. (2017). During the fall-winter period, wheat was always the strongest competitor, as expected in our environment. Due to the high clay content in the soil, Nmin at sowing was sufficient, even in the rainiest year (2010/11), to allow good initial growth in the cereal, and then low winter temperatures (Tosti et al. 2016) limited legume growth more than wheat growth.
The total N accumulation of each intercrop (i.e., the sum of the N accumulation in the two companion species) on the legume devitalization date was always higher than that of each species in pure stands (i.e., the total LER was always higher than 1). ICF was always the most efficient, with a maximum LER greater than 2 in 2009/10.
3.3 SPAD values during the crop cycle
In each year, the time-course of SPAD readings in pure wheat was the same for all mineral N controls (Fig. 3), i.e., substantially stable or slightly increasing during stem elongation and declining after heading; however, the SPAD values differed according to the mineral N rate applied (i.e., N160 > N120 > N80 > N40 > N0). Moreover, the absolute SPAD values were affected by the base soil Nmin availability (i.e., confounding mineral N dose, 2009/10 ≅ 2010/11 < 2011/12 on average).

Time-course (day of the year, DOY) of the variation in SPAD readings of leaves of wheat grown in temporary (TICs) and permanent (PICs) intercrops with faba bean (TICF and PICF, respectively), pea (TICP and PICP, respectively), and clover (TICC and PICC, respectively) and in control treatments, where pure-stand wheat was fertilized with mineral N at the rates of 0 kg ha−1 (N0), 80 kg ha−1 (N80), and 160 kg ha−1 (N160) in 2009/2010, 2010/2011, and 2011/2012. In each graph, the arrows pointing toward the x axis indicate the legume devitalization (left) and wheat heading (right) dates. Vertical bars indicate ± 1 SEM
In intercrops, the wheat SPAD values varied with (i) the year, (ii) the legume companion, and (iii) the duration of companion species coexistence (i.e., TIC or PIC, Fig. 3). In 2009/10, the SPAD values in TICF and TICP were similar to those in N0 during stem elongation (DOY = 83) and increased up to those of N80 just after wheat heading (DOY = 130, Fig. 3), while they were always similar to those of N0 in TICC. In all PICs, the SPAD values increased from late stem elongation (DOY = 118) until initial grain filling (DOY = 140) and reached values close to those of N80 in PICF and PICP and between those of N0 and N80 in PICC. In 2010/11, the trends were similar to those recorded in the previous year. In all TICs, the values were lower than those in N0 in early stages and slightly higher from stem elongation onwards (Fig. 3), while in the PICs, the SPAD values increased with time up to those recorded in N160 in PICF, between N80 and N160 in PICP, and slightly above N0 in PICC. In 2011/12, the SPAD values were between those of N0 and N80 in all intercrops except for PICF, in which the SPAD values approximated those recorded in N160 (Fig. 3).
Overall, the SPAD readings strongly correlated to the N content per unit leaf area (SLNW) at heading (R2 = 0.821, df = 31), and SPAD was thus a good indicator of wheat N status. According to the SPAD readings, the wheat N status was improved by the presence of legumes, confirming previous evidence for both temporary (Tosti and Guiducci 2010) and permanent intercropping (Bedoussac et al. 2015).
Based on the SPAD readings, the wheat N status from stem elongation to approximately heading appeared to benefit from PICs more than from TICs. This apparently contradictory result can be explained by the effect of the legume cogrowth duration on wheat biomass accumulation. In fact, the wheat stem density in PICs strongly declined (in terms of both reduced tillering and later self-thinning) due to legume competition, especially in case of faba bean and pea. Thus, N was shared among the few stems that remained, i.e., diluted in a small amount of wheat biomass. Conversely, the legume devitalization in TICs allowed wheat plants to grow undisturbed and spread in the above- and belowground space of the missing legume, and this caused a great increase in wheat biomass and consequent N dilution.
3.4 N uptake in wheat at maturity and from heading to maturity
At maturity, the total N uptake of pure wheat in the mineral controls increased linearly with the fertilization rate (Table 1). The rates of wheat N uptake per unit of mineral N added were 0.59 ± 0.066 kg kg−1 (R2 = 0.964) in 2009/10, 0.48 ± 0.041 (R2 = 0.978) in 2010/11 and 0.68 ± 0.087 (R2 = 0.953) in 2011/12. The wheat N uptake in TICs varied according to the amount of N incorporated into the soil with the legume biomass (Table 1).
In TICF, the wheat N uptake was higher than in N0 in all years (+ 56% in 2009/10, + 25% in 2010/11 and + 15% in 2011/12), while in TICP and TICC, it was higher than in N0 only in 2009/10 (+ 57 and + 13%, respectively) and in 2010/11 (+ 32 and + 25%, respectively). Thus, in terms of the wheat total N uptake, the legume made the difference only when it delivered an amount of N that was relevant compared to the soil Nmin.
In the first two experimental years, TIC enhanced active N absorption during wheat grain filling, and this N uptake was higher in TIC compared to pure wheat (at any N fertilization dose). These findings are confirmed by the SPAD trends observed in TIC (Fig. 3), so even if a small amount of N was supplied to wheat with the incorporation of the legume biomass in the soil, the efficiency of such N uptake was remarkable and far from being marginal in terms of wheat plant N nutrition. Specifically, the extent of the N effect of the TIC treatments on wheat N nutrition was related to the considered leguminous species: the N absorbed by wheat after heading accounted for 27% (2009/10) and 43% (2010/11) of the total N uptake at maturity in TICF, for 21 and 52%, respectively, in TICP and for 31 and 44%, respectively, in TICC, compared with + 10 and + 35% recorded in the unfertilized control (N0). It is worth noting that the wheat LER for N was approximately 1 (Fig. 2); thus, after heading, the higher wheat N uptake in TICs compared to N0 was a result of the N supplied from the devitalized legume biomass. This N was easily available to wheat thanks to the low C/N ratio of the fresh legume biomass at that stage (Alonso-Ayuso et al. 2014; Benincasa et al. 2010).
On the other hand, wheat N uptake in PICs was reduced compared to both the N0 and TICs treatments in all years, and especially in 2009/10 (Table 1), when legumes were well established and grown in winter and could then become very competitive toward wheat in spring. In this year, the competition was so severe that wheat showed a net loss of N during grain filling (Table 1), i.e., the wheat plants were actually suppressed, causing a loss of wheat biomass and N. Negative performance in the cereal component at final harvest has been observed in several works on permanent intercrops (Bedoussac and Justes 2010; Verret et al. 2017), often related to high legume competition (Mariotti et al. 2009). One might assume that the increased N availability in TICs compared to PICs arose from enhanced soil organic matter mineralization caused by hoeing. This side effect cannot be completely excluded, but it likely had very little influence, given the low soil OM content (1.2%) and the low depth of the hoeing operation. In 2011/12, the wheat N accumulation from heading to maturity was always very low, and the differences among treatments were negligible. This is because most of the N was taken up in the early stages due to the high soil Nmin, and thus late N uptake was less important (i.e., the amount of N was lower both as an absolute value and compared to the total). Thus, again, the effect of intercrops depends on the interaction between companion species as affected by the soil and climate conditions. However, in TICs, the farmer has an additional choice of whether to modulate the interaction between species in favor of the cereal. He may establish the date of legume devitalization in order to increase the N supply and then devitalize the legume before its competition undermines wheat performance (Fayaud et al. 2014). The more competitive the legume (depending on the environmental circumstances), the earlier it will have to be removed. Species (and cultivars) actually differ in terms of their growth habit and adaptation to the environment (Cornelissen et al. 2003; Ramírez-García et al. 2015). In our study, the sequence of leguminous species devitalization was the same in all years (i.e., first faba bean, then pea, and finally clover), but the date changed according to legume and wheat growth performance.
3.5 Grain N accumulation and yield
The grain N accumulation of wheat in pure stands was always proportional to the mineral N rate, while it varied in TICs depending on the year and legume species (Fig. 4). In particular, it was always higher than in N0, except in TICC in 2011/12, and reached values closer to N80 in TICF and TICP in 2009/10 as a maximum.

Wheat grain N accumulation (kg ha−1), N concentration (N% in dry matter), and grain yield (t ha−1) in relation to the amount of N provided (kg ha−1) in the mineral-fertilized control treatments (0, 40, 80, 120, 160 kg N ha−1) by legume aboveground biomass incorporation in temporary intercrops (TICs) with faba bean (TICF), pea (TICP), and clover (TICC) and by coexistence with each of the three legumes in permanent intercrops (PICs). The N transferred to wheat from legumes in permanent intercrops was unknown and set to zero in the graphs. Bars represent ± 1 SEM
In PICs, N accumulation in wheat grains was always lower than that in N0 and the corresponding TIC, with the most severe reduction observed in 2009/10, when it dropped to almost zero in PICF.
The grain N concentration of pure wheat increased with the mineral N rate from N0 to N120 in 2009/10 and from N0 to N160 in 2010/11 and 2011/12 (Fig. 4). In TICs, it was always higher than in N0, except for TICC in 2011/12, and up to values recorded in N120 for all TICs in 2009/10 and for TICF and TICP in 2010/11. In the other cases, it was close to (TICC 2010/11, TICF 2011/12) or lower (TICP 2011/12) than the values recorded in N40.
In PICs, the N concentration of wheat grains was very high and higher than that in TICs in all years for PICF and PICP and in 2 years out of 3 in PICC (2009/10 and 2011/12).
Combining the data on wheat grain N accumulation and concentration shows the different effects of TICs and PICs on wheat grain yield and quality. In TICs, the increased N allocation to grains resulted from a higher grain N concentration associated with slight variations in grain yield. Only TICF (+ 26%) and TICP (+ 44%) in 2009/10 gave a wheat grain yield higher than that in N0 (Fig. 4). Although the N supplied as a result of legume incorporation was generally available too late to increase yield, it was effective in increasing the grain protein content beyond a mere N dilution effect. This is of relevance considering that low protein content is the main limitation to the use of organic wheat for baking and pasta (Giuliani et al. 2015). TICs did not increase grain yield, and PICs severely depressed it: the high grain N concentration in PICS cannot be considered as positive in light of such a dramatic (unacceptable) drop in grain yield. In fact, the lower the N accumulation in wheat grains, the higher was their N% concentration, which is clearly related to the N dilution effect in cereal-legume intercropping (Bedoussac et al. 2015). As discussed above, the results vary year by year depending on legume competition, with no way to modulate it in PICs, while the date of legume devitalization represents a key factor in TICs.
4 Conclusions
Results allow some relevant information to be drawn in terms of the management of wheat-legume temporary intercropping, highlighting the criteria and the effects to consider in the choice of (i) legume companions and (ii) the timing of devitalization.
In TICs, the ranking of the three legumes in terms of their “N effect” toward wheat was consistent in the 3 years, i.e., faba bean > pea > clover. The N effect was correlated with the competitive ability of the legume. The higher the legume competitive ability, the higher was the legume biomass and N accumulation during the cogrowth period, and thus the higher (and earlier) was the N supply provided to wheat from legume devitalization after winter. The legume devitalization date becomes a key factor to be managed year after year according to legume growth, as affected by environmental factors. The rainfall amount and/or low soil mineral N availability and mild winters enhance legume competition against wheat and vice versa. A compromise will take into account the amount of N that has been accumulated and is thus supplied by the legume (direct advantage) and the need to eliminate legume competition before it undermines wheat growth (indirect advantage). Such a modulation was not feasible in permanent intercrops, where higher legume competitive ability resulted in a lower grain yield of wheat. Thus, if the final aim is a high cereal grain yield, such permanent intercrops are not recommended. The N supplied as a result of legume devitalization slightly increased the total N uptake of wheat but was relevant to grain N accumulation, i.e., it caused no grain yield variation but caused an increased grain N concentration. In contrast, in permanent intercropping, the wheat grain yield was severely compromised, and the very high grain N% concentration, due only to the dilution effect, cannot be considered a positive outcome. In conclusion, temporary intercropping is an efficient tool for the sustainable management of N nutrition in winter wheat.
References
Agegnehu G, Ghizaw A, Sinebo W (2008) Yield potential and land-use efficiency of wheat and faba bean mixed intercropping. Agron Sustain Dev 28:257–263. https://doi.org/10.1051/agro:2008012
Alonso-Ayuso M, Gabriel JL, Quemada M (2014) The kill date as a management tool for cover cropping success. PLoS One 9(10):e109587. https://doi.org/10.1371/journal.pone.0109587
Annicchiarico P, Iannucci A (2007) Winter survival of pea, faba bean and white lupin cultivars in contrasting Italian locations and sowing times, and implications for selection. J Agric Sci 145:611–622. https://doi.org/10.1017/S0021859607007289
Annicchiarico P, Pagnotta MA (2012) Agronomic value and adaptation across climatically contrasting environments of Italian red clover landraces and natural populations. Grass Forage Sci 67:597–605. https://doi.org/10.1111/j.1365-2494.2012.00887.x
Bedoussac L, Justes E (2010) Dynamic analysis of competition and complementarity for light and N use to understand the yield and the protein content of a durum wheat–winter pea intercrop. Plant Soil 330:37–54. https://doi.org/10.1007/s11104-010-0303-8
Bedoussac L, Journet EP, Hauggaard-Nielsen H, Naudin C, Corre-Hellou G, Jensen ES, Prieur L, Justes E (2015) Ecological principles underlying the increase of productivity achieved by cereal-grain legume intercrops in organic farming. A review. Agron Sustain Dev 35:911–935. https://doi.org/10.1007/s13593-014-0277-7
Benincasa P, Tosti G, Tei F, Guiducci M (2010) Actual N availability from winter catch crops used for green manuring in maize cultivation. J Sustain Agric 34:705–723. https://doi.org/10.1080/10440046.2010.507452
Benincasa P, Pace R, Tosti G, Tei F (2012) Early interspecific interference in the wheat/faba bean (Triticum aestivum/ Vicia faba ssp. minor) and rapeseed/squarrosum clover (Brassica napus var. oleifera/Trifolium squarrosum) intercrops. Ital J Agron 7:39–45. https://doi.org/10.4081/ija.2012.e24
Benincasa P, Farneselli M, Tosti G, Bonciarelli U, Lorenzetti MC, Guiducci M (2016) Eleven-year results on soft and durum wheat crops grown in an organic and in a conventional low input cropping system. Ital J Agron 11:77–84. https://doi.org/10.4081/ija.2016.726
Benincasa P, Reale L, Tedeschini E, Ferri V, Cerri M, Ghitarrini S, Falcinelli B, Frenguelli G, Ferranti F, Ayano BE, Porfiri O, Rosati A (2017a) The relationship between grain and ovary size in wheat: an analysis of contrasting grain weight cultivars under different growing conditions. Field Crop Res 210:175–182. https://doi.org/10.1016/j.fcr.2017.05.019
Benincasa P, Tosti G, Guiducci M, Farneselli M, Tei F (2017b) Crop rotation as a system approach for soil fertility management in vegetables. In Tei F. et al. (ed) Advances in research on fertilization management of vegetable crops, advances in horticulture. Springer, Cham, pp 115–148. doi: https://doi.org/10.1007/978-3-319-53626-2_5
Blandino M, Vaccino P, Reyneri A (2015) Late-season nitrogen increases improver common and durum wheat quality. Agron J 107:680–690. https://doi.org/10.2134/agronj14.0405
Brooker RW, Bennett AE, Cong WF, Daniell TJ, George TS, Hallett PD, Hawes C, Iannetta PPM, Jones HG, Karley AJ, Li L, McKenzie BM, Pakeman RJ, Paterson E, Schöb C, Shen J, Squire G, Watson CA, Zhang C, Zhang F, Zhang J, White PJ (2015) Improving intercropping: a synthesis of research in agronomy, plant physiology and ecology. New Phytol 206:107–117. https://doi.org/10.1111/nph.13132
Cornelissen JHC, Lavorel S, Garnier E, Diaz S, Buchmann N, Gurvich DE, Reich PB, ter Steege H, Morgan HD, van der Heijden MGA, Pausas JG, Poorter H (2003) A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust J Bot 51:335–380. https://doi.org/10.1071/BT02124
Costanzo A, Bàrberi P (2014) Functional agrobiodiversity and agroecosystem services in sustainable wheat production. A review. Agron Sustain Dev 34:327–348. https://doi.org/10.1007/s13593-013-0178-1
Dresbøll DB, Thorup-Kristensen K (2014) Will breeding for nitrogen use efficient crops lead to nitrogen use efficient cropping systems? A simulation study of G×E×M interactions. Euphytica 199:97–117. https://doi.org/10.1007/s10681-014-1199-9
Fayaud B, Coste F, Corre-Hellou G, Gardarin A, Duerr C (2014) Modeling early growth under different sowing conditions: a tool to predict variations in intercrop early stages. Eur J Agron 52:180–190. https://doi.org/10.1016/j.eja.2013.09.009
Ferrise R, Bindi M, Martre P (2015) Grain filling duration and glutenin polymerization under variable nitrogen supply and environmental conditions for durum wheat. Field Crop Res 171:23–31. https://doi.org/10.1016/j.fcr.2014.10.016
Giuliani MM, De Palermo C, Mentana A, Pompa M, Giuzio L, Masci S, Centonze D, Flagella Z (2015) Differential expression of durum wheat gluten proteome under water stress during grain filling. J Agric Food Chem 63:6501–6512. https://doi.org/10.1021/acs.jafc.5b01635
Hekneby M, Antolin MC, Sanchez-Diaz M (2006) Frost resistance and biochemical changes during cold acclimation in different annual legumes. Environ Exp Bot 55:305–314. https://doi.org/10.1016/j.envexpbot.2004.11.010
Mariotti M, Masoni A, Ercoli L, Arduini I (2009) Above- and below-ground competition between barley, wheat, lupin and vetch in a cereal and legume intercropping system. Grass Forage Sci 64:401–412. https://doi.org/10.1111/j.1365-2494.2009.00705.x
Mariotti M, Masoni A, Ercoli L, Arduini I (2015) Nitrogen leaching and residual effect of barley/field bean intercropping. Plant Soil Environ 61:60–65. https://doi.org/10.17221/832/2014-PSE
Monti M, Pellicanò A, Santonoceto C, Preiti G, Pristeri A (2016) Yield components and nitrogen use in cereal-pea intercrops in Mediterranean environment. Field Crop Res 196:379–388. https://doi.org/10.1016/j.fcr.2016.07.017
R Core Team (2014) R: a language and environment for statistical computing. 3rd ed. Vienna, Austria. Available: http://www.R-project.org/
Ramírez-García J, Carrillo JM, Ruiz M, Alonso-Ayuso M, Quemada M (2015) Multicriteria decision analysis applied to cover crop species and cultivars selection. Field Crop Res 175:106–115. https://doi.org/10.1016/j.fcr.2015.02.008
Saia S, Urso V, Amato G, Frenda AS, Giambalvo D, Ruisi P, Miceli GD (2016) Mediterranean forage legumes grown alone or in mixture with annual ryegrass: biomass production, N2 fixation, and indices of intercrop efficiency. Plant Soil 402:395–407. https://doi.org/10.1007/s11104-016-2837-x
Thilakarathna MS, McElroy MS, Chapagain T, Papadopoulos YA, Raizada MN (2016) Belowground nitrogen transfer from legumes to nonlegumes under managed herbaceous cropping systems. A review. Agron Sustain Dev 36:58. https://doi.org/10.1007/s13593-016-0396-4
Thorup-Kristensen K, Dresbøll DB, Kristensen HL (2012) Crop yield, root growth, and nutrient dynamics in a conventional and three organic cropping systems with different levels of external inputs and N recycling through fertility building crops. Eur J Agron 37:66–82. https://doi.org/10.1016/j.eja.2011.11.004
Tosti G, Guiducci M (2010) Durum wheat-faba bean temporary intercropping: effects on nitrogen supply and wheat quality. Eur J Agron 33:157–165. https://doi.org/10.1016/j.eja.2010.05.001
Tosti G, Benincasa P, Farneselli M, Tei F, Guiducci M (2014) Barley-hairy vetch mixture as cover crop for green manuring and the mitigation of N leaching risk. Eur J Agron 54:34–39. https://doi.org/10.1016/j.eja.2013.11.012
Tosti G, Farneselli M, Benincasa P, Guiducci M (2016) Nitrogen fertilization strategies for organic wheat production: crop yield and nitrate leaching. Agron J 108:1–12. https://doi.org/10.2134/agronj2015.0464
Verret V, Gardarin A, Pelzer E, Mediene S, Makowski D, Valantin-Morison M (2017) Can legume companion plants control weeds without decreasing crop yield? A meta-analysis. Field Crop Res 204:158–168. https://doi.org/10.1016/j.fcr.2017.01.010
Volpi I, Antichi D, Ambus PL, Bonari E, Nassi O, Di Nasso N, Bosco S (2018) Minimum tillage mitigated soil N2O emissions and maximized crop yield in faba bean in a Mediterranean environment. Soil Tillage Res 178:11–21. https://doi.org/10.1016/jstill2017.12.016
Wang GZ, Li HG, Christie P, Zhang FS, Zhang JL, Bever JD (2017) Plant-soil feedback contributes to intercropping overyielding by reducing the negative effect of take-all on wheat and compensating the growth of faba bean. Plant Soil 415:1–12. https://doi.org/10.1007/s11104-016-3139-z
Willey RW, Osiru DSO (1972) Studies on mixtures of maize and beans (Phaseolus vulgaris) with particular reference to plant population. J Agric Sci 79:517–529. https://doi.org/10.1017/S0021859600025909
Acknowledgements
This experiment was part of the NITBIO project funded by MIPAAF (Ministero Italiano delle Politiche Agricole, Alimentari e Forestali, Italy), scientific coordinator Prof. Marcello Guiducci.
The authors thank the FIELDLAB staff for their important contributions to crop management and data collection.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
About this article

Cite this article
Guiducci, M., Tosti, G., Falcinelli, B. et al. Sustainable management of nitrogen nutrition in winter wheat through temporary intercropping with legumes. Agron. Sustain. Dev. 38, 31 (2018). https://doi.org/10.1007/s13593-018-0509-3
Accepted:
Published:
DOI: https://doi.org/10.1007/s13593-018-0509-3