
Abstract
Evidence has revealed that DDOST plays an important role in cancer development and progression. However, there are no reports on functions of DDOST in cervical tumorigenesis. Hence, we investigated the relationship of DDOST with prognosis, mutation, promoter methylation, immune cell infiltration, and drug sensitivity using bioinformatics techniques. Our results demonstrated that DDOST was significantly upregulated in a variety of tumor types and correlated with poor prognosis, including cervical cancer. Cox regression analysis dissected that high DDOST expression was associated with poor survival in cervical cancer patients. Immune infiltration analysis defined that DDOST was negatively correlated with CD8 T cells and NK cells. Strikingly, the sensitivity to multiple drugs was negatively correlated with the expression of DDOST. Therefore, our findings uncovered that DDOST could play an essential role in the tumor microenvironment and tumor immune regulation in cervical cancer, which indicated that DDOST could be a useful biomarker for prognosis and a potential therapeutic target for cancer treatment.
Similar content being viewed by others


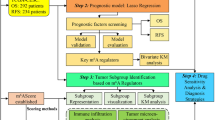
1 Introduction
Cervical cancer is one of the common gynecological cancers [1]. The number of patients who die each year due to cervical cancer exceeds 260,000, making cervical cancer the second most lethal malignancy in women in the world [2]. Human papillomavirus (HPV) infection has been found to be associated with cervical cancer developed [3]. The treatments include surgery, chemotherapy, and immunotherapy. Chemotherapy agents for cervical cancer often include cisplatin, paclitaxel, and topotecan [4]. Cervical oncogenesis is correlated with dysregulation of genetics and epigenetics, including noncoding RNAs [5].
Despite precancerous screening and emerging treatment options, cervical cancer remains one of the leading causes of death among women in developing countries [6,7,8]. When cervical cancer patients present with metastasis and recurrence, their prognosis is worse [9]. Therefore, based on its molecular mechanisms that remain to be elucidated, the search for new prognostic markers as well as the development of new therapeutic approaches is of great significance for cervical cancer [10,11,12].
DDOST (dolichyl-diphosphooligosaccharide–protein glycosyltransferase non-catalytic subunit), encodes a component of the oligosaccharide transferase (OST) complex and is related to the N-glycosylation of proteins [13]. DDOST was reported to be not associated with diabetic nephropathy in type 1 diabetes [14]. DDOST expression was regulated by dietary advanced glycated end-products and statins and angiotensin receptor inhibitors in peripheral mononuclear cells from patients with type 1 diabetes [15]. In congenital disorders of glycosylation, whole-exome sequencing discovered the mutations of DDOST [16]. Furthermore, DDOST was identified to interact with PPP1CC2 in transgenic mouse embryonic stem cells [17]. Several studies revealed that DDOST might be an oncogene in several tumor types. In esophageal squamous cell carcinoma (ESCC) patients, DDOST displayed missense variants, indicating that these variants might be diagnostic biomarkers for ESCC diagnosis [18]. Using integrated analysis of gene expression and DNA methylation profiles, one study found that DDOST gene was one of core genes in bladder cancer development [19]. However, the role of DDOST in cervical cancer was still unclear. Thus, we aim to explore the function of DDOST in cervical tumorigenesis.
Our study utilized multiple publicly available data to perform a systematic analysis of the pan-cancer role of DDOST in tumorigenesis. The underlying molecular mechanisms of DDOST in cervical cancer as well as pan-cancer differential expression, prognosis, genetic alterations, immune infiltration and sensitivity to targeted agents were explored. Quantitative real-time PCR assay and immunohistochemistry (IHC) were performed to validate the role of DDOST in cervical cancer. This study provides a theoretical basis for gaining insight into the role of DDOST in tumors.
2 Materials and methods
2.1 Differential expression analysis and data processing
The expression information of DDOST in 31 types of normal tissues was obtained from the GTEx portal, and the expression of DDOST was compared between 33 types of cancer tissues and 31 types of normal tissues (no normal tissues available for MESO and UVM, MESO: mesothelioma, UVM: uveal melanoma) by combining the data from TCGA with the data from GTEx. Both TCGA and GTEX data were downloaded from UCSC Xena database (https://xenabrowser.net/datapages/), tumor tissues were obtained from TCGA database, and normal tissues were derived from TCGA database and GTEx database.
2.2 Survival analysis
We obtained data from TCGA database to reveal the correlation of DDOST expression with patient prognosis, mainly in relation to overall survival (OS), disease-free interval (DFI), progression-free interval (PFI), and disease-specific survival (DSS). We analyzed the survival of all 33 cancer types, and the results were shown by forest plots and Kaplan–Meier curves from the univariate Cox regression analysis. The survminer R packages and survival were used for analysis and visualization.
2.3 Mutation analysis of the DDOST gene
The cbioportal database was selected for DDOST gene mutation analysis. Through this website, we obtained information on DDOST gene mutation frequency, mutation type, copy number alterations, and promoter methylation in pan-cancer tissues, including cervical cancer, from TCGA database, using human methylation 450 K array methylation data.
2.4 Enrichment analysis of DDOST
We leveraged GSEA to analyze the underlying molecular mechanism of DDOST in 33 cancer types. Analysis and visualization were performed using the R package clusterProfiler. We downloaded 50 HALLMARK pathways from the MsigDB database (http://www.gsea-msigdb.org/gsea/msigdb/index.jsp) and used GSVA package to score HALLMARK pathways and calculate the correlation of DDOST expression with pathways in pan-cancer, including cervical cancer.
2.5 Tumor microenvironment analysis
Tumor microenvironment analysis for each tumor demonstrates the fraction of pathways differentially represented in the high and low DDOST gene expression groups, and a heatmap was used to demonstrate the correlation of genes with pathway scores [20]. The correlation between DDOST expression and the TME-related signatures in pan-cancer was explored. From the ImmuCellAI database (bioinfo.life.hust.edu.cn/web/ImmuCellAI/), we downloaded pan-cancer immune infiltration data. The correlation of DDOST expression with immune cell infiltration was calculated and displayed.
2.6 DDOST related drug sensitivity analysis
Using GDSC (https://www.cancerrxgene.org/) database, we downloaded the half maximal inhibitory concentration (IC50) and gene expression data of tumor cells, analyzed the relationship between DDOST and drug (IC50), including ABT737, WEHI-539, Sabutoclax, PF-4708671, KU-55933, and Trametinib, and plotted the correlation between the expression of DDOST in each drug and IC50, respectively.
2.7 Cell culture
CaSki (HPV16), SiHa (HPV16), HeLa (HPV18), C33A (HPV negative) and Ect1/E6E7 cells were purchased from the American Type Culture Collection. HeLa, SiHa and Ect1/E6E7 cells were cultured in DMEM medium. C33A and CaSki cells were cultures in RPMI 1640 medium. Cell medium were supplemented with 10% FBS and 1% penicillin/streptomycin. Cells were maintained in 37 °C incubator with 5% CO2.
2.8 Quantitative real-time PCR assay
Total RNA (1 g) was extracted from cells using RNA-easy Isolation Reagent (R701-01, Vazyme) following the manufacturer’s instructions. Reverse transcription was performed using TransScript All-in-One First-Strand cDNA Synthesis SuperMix for qPCR (P20607, TransGen Biotech, China), and the reaction was blended with PerfectStart Green qPCR SuperMix (P20604,TransGen Biotech, China) and run on a BIO-RAD CFX96 Real time PCR system as follows: 94 °C 30 s, 94 °C 5 s and 60 °C 30 s for 40 cycles. The sequences of primers were: DDOST: forward 5′-GCT CAC ATT CAA GAC CGC TG-3′; reverse 5′-CGT GAT CCA GCA GCT CTC AA-3′. Beta actin: forward 5′-ACA GAG CCT CGC CTT TGC CGA T-3′; reverse 5′-GGC CTC GTC GCC CAC ATA GGA-3′. Quantitative real-time PCR was conducted as described previously [21].
2.9 Immunohistochemistry (IHC)
We collected 40 cervical cancer tissues and their adjacent non-tumor tissues. The tissues were formalin-fixed, paraffin-embedded, and sliced into 4 M section. We used xylene to deparaffinize and rehydrated in ethanol. Then, microwave was used to retrieve antigen, and anti-DDOST antibody (1:500, Rabbit polyclonal antibody, Abcam Company, ab204314) was applied for incubation with the slices overnight at 4 °C. The slices were washed by PBS three times, and incubated with the relevant secondary antibody for 45 min. Then, IHC were conducted as described previously [22]. The human tissue study was approved by the ethical committee of the Second Affiliated Hospital of Wenzhou Medical University.
2.10 Statistical analysis
The GraphPad Prism 7.0 software was used for statistical analysis. The difference between two groups was analyzed with the Student’s t test. The difference among multiple groups was analyzed by ANOVA (Analysis of variance). P < 0.05 was considered significant. All data are presented as mean ± standard deviation (SD).
3 Results
3.1 Expression levels of DDOST in multiple cancer types and normal tissues
We first performed a joint analysis of TCGA and GTEx data and found that DDOST expression was differed between 30 types of cancer samples and normal tissues. Among them, DDOST was highly expressed in 27 cancer types, including CESC, and lowly expressed in 3 cancer types compared with corresponding normal tissues (Fig. 1A). Based on the TCGA database, we analyzed the expression levels of DDOST in tumor tissues of 33 cancer types and ranked them from high to low by mean expression of DDOST (Fig. 1B). All cancer types expressed DDOST, with the highest level of expression in TGCT and the lowest in KICH. Next, we analyzed the DDOST expression levels among normal tissues using the GTEx data. Expression level of DDOST was highest in bone marrow and lowest in brain (Fig. 1C).

DDOST expression analysis in Pan-cancer. A, Pan-cancer differential expression of DDOST between tumor tissues from TCGA and normal tissues from TCGA and GTEx database is illustrated. B, DDOST expression in tumor tissues from TCGA database. C, DDOST expression in normal tissues from GTEx database. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001
We also analyzed the DDOST expression based on TCGA data and found that DDOST was overexpressed in most tumor types, including CESC (Fig. 2A). Using single cell expression profiling of cervical cancer, we revealed that DDOST was universally expressed in tumor microenvironment (TME), including CD8 T cells, fibroblasts and SMC. The most highly expression of DDOST was in malignant cells (Fig. 2B).

DDOST expression in single cell sequence data. A. The differential expression of DDOST between tumor tissues and normal tissues was presented. B. DDOST expression in single cell sequence data of cervical cancer using TISCH database. *P < 0.05, **P < 0.01, ***P < 0.001
3.2 Correlation analysis between DDOST expression and gene alterations
We further evaluated the gene variation landscape of DDOST in different tumor samples, including mutation, amplification, and deletion types, through the cBioportal database. As shown in Fig. 3A, DDOST had the highest alteration frequency (> 5%) in patients with CHOL, which was predominantly “Deep Deletion”. The correlation of DDOST expression with copy number was shown in Fig. 3B. In cervical cancer, DDOST expression was positively correlated with copy number. Correlation between DDOST gene expression and promoter methylation was revealed, and there was no significant correlation between DDOST expression and its methylation in cervical cancer (Fig. 3C), suggesting that hypomethylation of DDOST promoter could lead to high expression of DDOST in cervical cancer.

The gene alteration of DDOST in pan-cancer. A, CNA and mutation frequency of DDOST in TCGA pan-cancer were accessed from cBioPortal database. B, The correlation of DDOST mRNA expression and linear copy-number value in indicated tumor types from TCGA. C, The correlation of DDOST mRNA expression and promoter methylation level in indicated tumor types from TCGA
3.3 Pan-cancer analysis of the prognostic value of DDOST
We evaluated the relationship between DDOST expression and patient prognosis in a pan-cancer, with survival metrics as OS, DSS, DFI, and PFI. Cox regression analysis of 33 tumor types showed that the expression of DDOST was significantly associated with prognosis in multiple cancer types, especially cervical cancer. Cervical cancer patients with high expression of DDOST had worse OS (Fig. 4A), DSS (Fig. 4B), DFI (Fig. 4C), and PFI (Fig. 4D). Furthermore, Kaplan–Meier survival curves showed that in 16 cancer types, DDOST was a high-risk factor for shorter OS, including cervical cancer (Fig. 5).

Univariate regression analysis of DDOST in pan-cancer. A–D, Forest map shows the univariate cox regression analysis results of DDOST in TCGA pan-cancer, including OS (A), DSS (B), DFI (C), and PFI (D)

Overall survival analysis of DDOST in pan-cancer. Kaplan–Meier survival analysis results of DDOST in indicated tumor types from TCGA pan-cancer
3.4 Enrichment analysis of DDOST
To explore the biological significance of DDOST in different tumor tissues, we analyzed the correlation between DDOST expression and 50 HALLMARK pathways. We found that DDOST was most strongly associated with Glycolysis, Unfolded protein response, and PI3K/AKT/mTOR signaling (Fig. 6). We next performed correlation analysis of DDOST with all mRNAs in cervical cancer and demonstrated gene expression of Top50 positively (Fig. 7A) and negatively (Fig. 7B) correlated with DDOST, respectively. DDOST was closely associated with RNA splicing via transesterification reactions in GO terms (Fig. 7C), protein processing in endoplasmic reticulum in KEGG terms (Fig. 7D), and late phase of HIV life cycle and most cell cycle-related pathways in Reactome terms (Fig. 7E) in cervical cancer.

GSVA of DDOST. The GSVA results of DDOST based on 50 HALLMARK pathways in pan-cancer. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001

GSEA of DDOST. A–B, The expression of top 50 genes positively or negatively associated with DDOST expression in cervical cancer. C–E The top 20 GSEA results of DDOST were showed based on GO (C), KEGG (D), and Reactome (E) pathways in cervical cancer
3.5 Association of DDOST with TME
Next, we explored the correlation between DDOST expression and the TME-related signatures in pan-cancer. As shown in Fig. 8A, DDOST expression was significantly correlated with various pathway scores in pan-cancer, among which were mainly DNA damage repair and EMT related pathways. In cervical cancer, DNA damage repair and EMT related pathways were enriched in DDOST high-expression group (Fig. 8B).

TME analysis of DDOST. A. The correlation of DDOST with indicated TME-related pathways in pan-cancer. B. The score of indicated TME-related pathways in high- and low-DDOST expression group in cervical cancer. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001
We also analyzed the correlation of DDOST with immune cell infiltration in the TME of cervical cancer. A total of 10 immune cells were related with DDOST (Fig. 9A). DDOST expression in cervical cancer was positively correlated with cancer-promoting immune cell Tregs and negatively correlated with cancer-suppressing immune cells, such as CD8 T cells and NK cells (Fig. 9B). We divided cervical cancer patients into two groups according to the median DDOST expression. The Treg cell infiltration level was higher, while the infiltration levels of CD8 T cells and NK cells were lower in the high DDOST expression group (Fig. 9C).

Immune infiltration analysis. A–B, The relationship between DDOST expression and infiltration levels of indicated immune cells in cervical cancer. C. The infiltration levels of indicated immune cells in high- and low-DDOST expression group in cervical cancer. *P < 0.05, **P < 0.01, ****P < 0.0001. P = 0 means p < 0.0001
3.6 Association of DDOST with drug sensitivity
We analyzed the relationship of DDOST and drug resistance to provide a suitable medication selection for patients with high expression DDOST. The results indicated that, among 192 anti-tumor drugs in GDSC database, DDOST expression was positively correlated with the IC50 of 32 drugs, such as ABT737, WEHI-539, Sabutoclax, cisplatin, oxaliplatin, and gemcitabine, and negatively correlated with the IC50 of 11 drugs, such as PF-4708671, KU-55933, dasatinib, and Trametinib (Fig. 10, Supplementary Table 1). The above results suggested that DDOST was highly likely to be applied in the clinic as a drug target.

Drug resistance analysis of DDOST. The correlation between DDOST expression and IC50 values of indicated anti-cancer drugs. P = 0 means p < 0.0001
3.7 Experimental verification of DDOST expression in cervical cancer
Finally, we conducted experimental validation on the expression of DDOST in cervical cancer. Through qPCR detection, we found that the expression of DDOST was higher in tumor cell lines than in normal cell lines (P < 0.01, Fig. 11A). Through immunohistochemistry, we found that the protein level of DDOST was higher in tumor tissues than in adjacent non-tumor tissues in 40 cervical cancer patients (Fig. 11B, C). These results validate the findings of bioinformatics analysis.

Experimental verification of DDOST expression in cervical cancer A, The expression of DDOST in indicated cell lines via qRT-PCR assay. B–C, The protein expression of DDOST in cervical and tumor tissues via immunohistochemical assay
4 Discussion
Glycosylation, the attachment of carbohydrates to polypeptides, is an important form of protein post-translational modification in eukaryotes and is present in more than half of proteins [23,24,25]. Evidence suggests that glycosylation of proteins is non negligible in the process of tumor progression [26,27,28]. In particular, OST-mediated protein glycosylation is a key player in signal transduction, protein folding, and degradation [29, 30]. It has also been confirmed that glycosylation of protein is closely related to immune escape of tumor cells [31,32,33]. DDOST, a subunit of the OST complex, plays a critical role in N-glycosylation. Due to that there are no researches on DDOST in cervical cancer, in the current study, we explored the role of DDOST in cervical carcinogenesis.
One study used integrated bioinformatics analysis and validated that DDOST was correlated with progression and growth in colon adenocarcinoma [34]. DDOST expression was increased and correlated with tumor metastasis in cutaneous squamous cell carcinomas [35]. Endocytosis-associated protein SCAMP3 was highly expressed and associated with poorer survival and progression in hepatocellular carcinoma (HCC). DDOST expression was positively associated with SCAMP3 in HCC patients [36]. One group reported that DDOST was involved in breast oncogenesis and could be a valuable biomarkers for breast cancer development [37]. Suppression of ribophorin 1 triggered endoplasmic-reticulum-stress-induced apoptosis in part via upregulation of DDOST in breast cancer [38]. Downregulation of DDOST reduced cell proliferation in hepatocellular carcinoma (HCC) [39]. Moreover, DDOST could be a prognostic biomarker and correlated with immune infiltrates in HCC [40]. One study showed that DDOST associated with malignancies and immune TME in gliomas [41]. DDOST may serve as an unfavourable biomarker for gliomas [42]. In view of this, our study comprehensively investigated role of DDOST in pan-cancer and cervical cancer. Joint analysis of TCGA and GTEx databases revealed that DDOST was significantly upregulated in 27 of 33 tumor types, including cervical cancer. Experimental verification of DDOST expression in cervical cancer cells and tumor tissues confirmed this conclusion.
Zhu et al. found that high expression of DDOST was linked to poorer survival in HCC patients. DDOST was linked to cell cycle and immune response by the PPAR pathway. Specifically, DDOST expression was associated with immune infiltration of Th2 cell and cytotoxic cells [40]. Similarly, DDOST was revealed to be associated with worse prognosis in glioma patients. DDOST was correlated with immune microenvironment in gliomas, which was related to infiltration of B cells, CD4 + T cells, tumor-associated macrophages and CAFs [41]. MELK (maternal embryonic leucine zipper kinase) was elevated in glioma tissues and associated with poorer survival. DDOST induced the tumor immunosuppressive microenvironment in gliomas [43]. The TME is the cellular environment in which tumor cells reside, and its composition includes extracellular matrix, soluble molecules, and tumor stromal cells [44]. Using single cell expression profiling of cervical cancer, we revealed that DDOST was universally expressed in TME. Among TME components, the most highly expression of DDOST was in malignant. We also analyzed the correlation of DDOST with immune cell infiltration in the TME of cervical cancer. DDOST expression in cervical cancer is positively correlated with cancer-promoting immune cell Tregs and negatively correlated with cancer-suppressing immune cells, such as CD8 T cells and NK cells. These results indicated that patients with high expression of DDOST may be in immunosuppressive TME, which may be one of the main reasons that DDOST causes the poor prognosis of patients.
We evaluated the relationship between DDOST expression and patient prognosis in a pan-cancer. Cox regression analysis of 33 tumor types showed that the expression of DDOST was significantly associated with prognosis in multiple cancer types, especially cervical cancer. Cervical cancer patients with high expression of DDOST had worse OS, DSS, DFI, and PFI. Furthermore, Kaplan–Meier survival curves showed that in 16 tumor species, DDOST was a high-risk factor for OS, including cervical cancer. These results indicated that DDOST was a potential oncogene and risk factor in cervical cancer and most tumors. GSVA and GSEA revealed that DDOST was most strongly associated with glycolysis, unfolded protein response, PI3K/AKT/mTOR signaling, and most cell cycle-related pathways. It is known that PI3K/AKT/mTOR pathway is involved in TME and regulation of immune response [45, 46]. These results suggest that DDOST may have a complex role in influencing the cell cycle and TME. Next, we explored the correlation between DDOST expression and the TME-related signatures in pan-cancer. DDOST expression was significantly correlated with various pathway scores in pan-cancer, among which were mainly DNA damage repair and EMT related pathways. Finally, drug sensitivity analysis also suggests that DDOST could be a potential target for anti-cancer therapy.
Taken together, our findings suggest that DDOST could serve as a prognostic biomarker in cervical cancer as well as in a variety of tumors. High DDOST expression confers poor prognosis of tumor patients. In addition, the expression of DDOST is closely related to the immunosuppressive TME. It is required to explore the function of DDOST in cervical cancer cells, including cell proliferation, apoptosis, cell cycle, migration and invasion. Moreover, in vivo study is necessary to determine the role of DDOST in cervical tumorigenesis. Furthermore, the mechanism of DDOST-involved cervical oncogenesis needs to be clarified. These findings may help elucidate the role of DDOST in tumorigenesis and progression and provide a reference for achieving precision cancer therapy and personalized immunotherapy in the future.
Data availability
All the data will be provided on reasonable request from the corresponding author.
Abbreviations
- NK cells:
-
Natural killer cells
- PPP1CC2:
-
Phosphoprotein phosphatase 1 catalytic subunit gamma 2
- PCR:
-
Polymerase chain reaction
- FBS:
-
Fetal bovine serum
- DMEM:
-
Dulbecco's Modified Eagle Medium
- RPMI:
-
Roswell Park Memorial Institute
- PI3K:
-
Phosphoinositide 3-kinase
- mTOR:
-
Mammalian target of rapamycin
- TME:
-
Tumor microenvironment
- EMT:
-
Epithelial–mesenchymal transition
- SCAMP3:
-
Secretory carrier membrane protein 3
- PPAR:
-
Peroxisome proliferator-activated receptor
- CAFs:
-
Cancer-associated fibroblasts
References:
Siegel RL, Miller KD, Wagle NS, Jemal A. Cancer statistics, 2023. CA Cancer J Clin. 2023;73:17–48.
Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71:209–49.
Zhang Z, Ma Q, Zhang L, et al. Human papillomavirus and cervical cancer in the microbial world: exploring the vaginal microecology. Front Cell Infect Microbiol. 2024;14:1325500.
Rosen VM, Guerra I, McCormack M, et al. Systematic review and network meta-analysis of bevacizumab plus first-line topotecan-paclitaxel or cisplatin-paclitaxel versus non-bevacizumab-containing therapies in persistent, recurrent, or metastatic cervical cancer. Int J Gynecol Cancer. 2017;27:1237–46.
Folliero V, Dell’Annunziata F, Chianese A, et al. Epigenetic and genetic keys to fight hpv-related cancers. Cancers. 2023. https://doi.org/10.3390/cancers15235583.
Rotman J, den Otter LAS, Bleeker MCG, et al. PD-L1 and PD-L2 expression in cervical cancer: regulation and biomarker potential. Front Immunol. 2020;11:596825.
Perkins RB, Wentzensen N, Guido RS, Schiffman M. Cervical cancer screening: a review. JAMA. 2023;330:547–58.
Alias NA, Mustafa WA, Jamlos MA, Alkhayyat A, Rahman KSA, R QM. Improvement method for cervical cancer detection: a comparative analysis. Oncol Res. 2021;29:365–76.
Yu S, Li X, Zhang J, Wu S. Development of a novel Immune Infiltration-based gene signature to predict prognosis and immunotherapy response of patients with cervical cancer. Front Immunol. 2021;12:709493.
Kakotkin VV, Semina EV, Zadorkina TG, Agapov MA. Prevention strategies and early diagnosis of cervical cancer: current state and prospects. Diagnostics. 2023. https://doi.org/10.3390/diagnostics13040610.
Das S, Babu A, Medha T, et al. Molecular mechanisms augmenting resistance to current therapies in clinics among cervical cancer patients. Med Oncol. 2023;40:149.
Liu H, Ma H, Li Y, Zhao H. Advances in epigenetic modifications and cervical cancer research. Biochim Biophys Acta Rev Cancer. 2023;1878:188894.
Yamagata T, Tsuru T, Momoi MY, et al. Genome organization of human 48-kDa oligosaccharyltransferase (DDOST). Genomics. 1997;45:535–40.
Hoverfelt A, Sallinen R, Soderlund JM, et al. DDOST, PRKCSH and LGALS3, which encode AGE-receptors 1, 2 and 3, respectively, are not associated with diabetic nephropathy in type 1 diabetes. Diabetologia. 2010;53:1903–7.
Santos-Bezerra DP, Machado-Lima A, Monteiro MB, et al. Dietary advanced glycated end-products and medicines influence the expression of SIRT1 and DDOST in peripheral mononuclear cells from long-term type 1 diabetes patients. Diab Vasc Dis Res. 2018;15:81–9.
Jones MA, Ng BG, Bhide S, et al. DDOST mutations identified by whole-exome sequencing are implicated in congenital disorders of glycosylation. Am J Hum Genet. 2012;90:363–8.
MacLeod G, Varmuza S. Tandem affinity purification in transgenic mouse embryonic stem cells identifies DDOST as a novel PPP1CC2 interacting protein. Biochemistry. 2012;51:9678–88.
Donner I, Katainen R, Tanskanen T, et al. Candidate susceptibility variants for esophageal squamous cell carcinoma. Genes Chromosom Cancer. 2017;56:453–9.
Zhang Y, Fang L, Zang Y, Xu Z. Identification of core genes and key pathways via integrated analysis of gene expression and dna methylation profiles in bladder cancer. Med Sci Monit. 2018;24:3024–33.
Zeng D, Li M, Zhou R, et al. Tumor microenvironment characterization in gastric cancer identifies prognostic and immunotherapeutically relevant gene signatures. Cancer Immunol Res. 2019;7:737–50.
Liu Y, Xu Y, Jiang W, Ji H, Wang ZW, Zhu X. Discovery of key genes as novel biomarkers specifically associated with HPV-negative cervical cancer. Mol Ther Methods Clin Dev. 2021;21:492–506.
Lin M, Zhang J, Bouamar H, Wang Z, Sun LZ, Zhu X. Fbxo22 promotes cervical cancer progression via targeting p57(Kip2) for ubiquitination and degradation. Cell Death Dis. 2022;13:805.
Eichler J. Protein glycosylation. Curr Biol. 2019;29:R229–31.
Schjoldager KT, Narimatsu Y, Joshi HJ, Clausen H. Global view of human protein glycosylation pathways and functions. Nat Rev Mol Cell Biol. 2020;21:729–49.
Reily C, Stewart TJ, Renfrow MB, Novak J. Glycosylation in health and disease. Nat Rev Nephrol. 2019;15:346–66.
Chen M, Assis DM, Benet M, et al. Comparative site-specific N-glycoproteome analysis reveals aberrant N-glycosylation and gives insights into mannose-6-phosphate pathway in cancer. Commun Biol. 2023;6:48.
Huang N, Wang Q, Chen CY, et al. N-glycosylation on Asn-57 is required for the correct HAI-2 protein folding and protease inhibitory activity. Glycobiology. 2023;33:203–14.
Zhang Y, Li J, Huang Y, et al. Improved antitumor activity against prostate cancer via synergistic targeting of Myc and GFAT-1. Theranostics. 2023;13:578–95.
Harada Y, Ohkawa Y, Kizuka Y, Taniguchi N. Oligosaccharyltransferase: A gatekeeper of health and tumor progression. Int J Mol Sci. 2019. https://doi.org/10.3390/ijms20236074.
Matsuda-Lennikov M, Biancalana M, Zou J, et al. Magnesium transporter 1 (MAGT1) deficiency causes selective defects in N-linked glycosylation and expression of immune-response genes. J Biol Chem. 2019;294:13638–56.
Mikolajczyk K, Kaczmarek R, Czerwinski M. How glycosylation affects glycosylation: the role of N-glycans in glycosyltransferase activity. Glycobiology. 2020;30:941–69.
Matsumoto Y, Ju T. Aberrant glycosylation as Immune therapeutic targets for solid tumors. Cancers. 2023. https://doi.org/10.3390/cancers15143536.
Bangarh R, Khatana C, Kaur S, et al. Aberrant protein glycosylation: implications on diagnosis and Immunotherapy. Biotechnol Adv. 2023;66:108149.
Hu M, Fu X, Si Z, et al. Identification of differently expressed genes associated with prognosis and growth in colon adenocarcinoma based on integrated bioinformatics analysis. Front Genet. 2019;10:1245.
Shapanis A, Lai C, Smith S, et al. Identification of proteins associated with development of metastasis from cutaneous squamous cell carcinomas (cSCCs) via proteomic analysis of primary cSCCs. Br J Dermatol. 2021;184:709–21.
Zhou A, Liu H, Tang B. Comprehensive evaluation of endocytosis-associated protein scamp3 in hepatocellular carcinoma. Pharmgenomics Pers Med. 2020;13:415–26.
Madrid FF, Grossman LI, Aras S. Mitochondria autoimmunity and MNRR1 in breast carcinogenesis: a review. J Cancer Immunol. 2020;2:138–58.
Ding J, Xu J, Deng Q, et al. Knockdown of Oligosaccharyltransferase subunit ribophorin 1 Induces endoplasmic-reticulum-stress-dependent cell apoptosis in breast cancer. Front Oncol. 2021;11:722624.
Lin S, Cao Y, Zhu K, et al. Identification of a novel prognostic signature based on n-linked glycosylation and its correlation with immunotherapy response in hepatocellular carcinoma. J Hepatocell Carcinoma. 2023;10:1749–65.
Zhu C, Xiao H, Jiang X, Tong R, Guan J. Prognostic biomarker DDOST and its correlation with immune infiltrates in hepatocellular carcinoma. Front Genet. 2021;12:819520.
Chang X, Pan J, Zhao R, et al. DDOST correlated with malignancies and immune microenvironment in gliomas. Front Immunol. 2022;13:917014.
Liu J, Feng C, Liu M, et al. An immune-related multi-omics analysis of dolichyl-diphosphooligosaccharide protein glycosyltransferase in glioma: Prognostic value exploration and competitive endogenous RNA network identification. IET Syst Biol. 2023;17:271–87.
Yang H, Zhou H, Wang G, et al. MELK is a prognostic biomarker and correlated with immune infiltration in glioma. Front Neurol. 2022;13:977180.
Chen Y, McAndrews KM, Kalluri R. Clinical and therapeutic relevance of cancer-associated fibroblasts. Nat Rev Clin Oncol. 2021;18:792–804.
Tian LY, Smit DJ, Jucker M. The role of PI3K/AKT/mTOR signaling in hepatocellular carcinoma metabolism. Int J Mol Sci. 2023. https://doi.org/10.3390/ijms24032652.
Mafi S, Mansoori B, Taeb S, et al. mTOR-Mediated regulation of immune responses in cancer and tumor microenvironment. Front Immunol. 2021;12:774103.
Author information
Authors and Affiliations
Contributions
Jie Mei performed the study and wrote the manuscript. Liuliu Pan, Min Huang, Dandan Bao, Hui Gao performed the study and edited the manuscript. Danhan Wang designed the experiments, supervised the study, and revised the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
This work was approved by the Ethics Committee of The Second Affiliated Hospital of Wenzhou Medical University.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mei, J., Pan, L., Huang, M. et al. DDOST is associated with tumor immunosuppressive microenvironment in cervical cancer. Discov Onc 15, 69 (2024). https://doi.org/10.1007/s12672-024-00927-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12672-024-00927-z