
Abstract
Transgenic plants serve as attractive systems for the production and delivery of subunit vaccines, thus expression of an enterotoxigenic Escherichia coli (ETEC) antigen in an edible plant may lead to the development of a viable oral vaccine against cholera and ETEC diarrhea. In this study, expression of the heat labile toxin B subunit (LTB) from ETEC was performed in lettuce, and its immunological characterization was investigated. A total of 27 independent transgenic lines were established following Agrobacterium-mediated transformation. Selected lettuce lines were subjected to GM1-ELISA to confirm the proper quaternary structure of the LTB protein. Levels of accumulation of the pentameric LTB reached up to 0.05% of the total soluble protein (TSP) in T1 and T2 progenies of these lines. Oral immunization of Balb/c mice was conducted using three weekly doses of lettuce-derived LTB. This elicited specific and significant antibody responses in both serum and intestinal tissues. Moreover, mice immunized with lettuce-derived LTB showed diminished intestinal fluid accumulation following challenge with the cholera toxin. This study demonstrated that this plant-based vaccine may contribute to immunization practices against diarrheal diseases.
Similar content being viewed by others

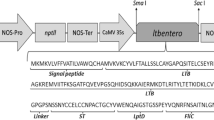
Introduction
Diarrheal diseases remain the second most frequent contributors to mortality of children in developing countries. There is a great need to develop effective vaccines against the major bacterial enteropathogens, including Enterotoxigenic Escherichia coli (ETEC), that result in significant rates of morbidity and mortality in children up to 5 years of age, and also affecting adults in these countries (Black 1993; Sack et al. 1971). ETEC is caused by E. coli strains that produce either one or both toxins, the heat-stable toxin (ST) and/or the heat-labile toxin (LT), and participate in the production of profuse watery diarrhea. Anti-toxin immunity has been reported to provide at least short-term protection against ETEC (Peltola et al. 1991; Clements et al. 1991). Interestingly, LT structure and function are highly similar to those of the cholera toxin (CT) as both contain A and B subunits, whose amino acid sequences are 80% similar, and they lead to diarrhea via persistent synthesis of cAMP following the delivery of the A subunit to the cytosol of target cells (Spangler 1992). The B subunits of LT (LTB) and CT (CTB) are non-toxic, and they are potent mucosal immunogens. Immunization with LTB increases the antitoxin response that also extends to the CTB subunit, thereby providing equally strong protection against oral LT and/or CT challenge (Klipstein et al. 1984; Clements et al. 1988). CTB has been used to develop a vaccine against ETEC, but its production requires costly fermentation and purification processes (Qadri et al. 2000).
Due to its importance as a candidate vaccine against ETEC, LTB has been expressed in bacterial systems and in yeast (Hirst 1991; Rezaee et al. 2005). In recent years, plants have been used to express antigenic proteins against myriad diseases as they serve as low-cost “biofactories” and vehicles for these functional protective antigens (Hoffmeisterová et al. 2008; Kim et al. 2006, 2011; Landell et al. 2008; Lau and Korban 2009). These alternative subunit vaccines have several advantages, such as low-cost of production and friendly administration (Mason et al. 2002).
Several plant models, such as tobacco, potato, maize, ginseng, carrot, and watercress have been utilized to assess the feasibility of developing a plant-based vaccine against ETEC (Haq et al. 1995; Mason et al. 1998; Chikwamba et al. 2002; Kang et al. 2006; Rosales-Mendoza et al. 2007; Loc et al. 2011). In general, these studies have confirmed that plant-derived LTB retains both its antigenicity and immunogenicity compared with the native LTB. Haq et al. (1995) have indicated that inclusion of the microsomal retention signal SEKDEL in the plant construct increases LTB accumulation in plant tissues. Later, Mason et al. (1998) have reported that using a plant-optimized synthetic gene also results in increased LTB accumulation in potato leaves and tubers.
Development of models based on food crops that are consumed raw would help in designing practical strategies that are compatible with oral administration of these subunit vaccines. As lettuce (Lactuca sativa L.) is readily consumed in fresh form, unprocessed, and has been evaluated as a production platform for the hepatitis B virus surface antigen (Kapusta et al. 2001) and the Measles virus hemagglutinin (MV-H) protein (Webster et al. 2006) with promising results, we have elected to evaluate it for expression and oral delivery of the LTB antigen. In this study, we have successfully developed transgenic lettuce lines expressing an LTB-encoding synthetic gene and evaluated levels of expression of the LTB antigen over two generations. Moreover, mice orally-fed freeze-dried transgenic lettuce leaves have produced both serum and mucosal antibodies against LTB protein, and are protected following challenge with a cholera toxin.
Materials and methods
Plant material, construct, and bacterial cell culture
Two commercial loose-leaf lettuce cvs. Potosina (Asgrow, Rio Rico, AZ, USA) and ‘Green Wave’ (Takii Seed Co., Ltd., Kyoto, Japan; gifted by Dr. Masaru Nakano, Niigata University) were used in this study. Cotyledons from seven-day-old seedlings were excised near the cotyledonary node, wounded six times with a scalpel along the abaxial surface, and used for transformation following the protocol described by Curtis et al. (1994).
The pBI-LTB binary vector, containing a synthetic LTB gene, optimized for plant expression and including the SEKDEL endoplasmic reticulum retention signal at the 3′ end, under the control of the CaMV35S promoter was used (Rosales-Mendoza et al. 2007).
A bacterial cell suspension culture of Agrobacterium tumefaciens strain LBA4404 harboring the pBI-LTB vector was grown in a YM liquid medium supplemented with 50 mg/l kanamycin for a period of 3 days, and then diluted 1:10 with the same medium to a final concentration of OD600nm = 0.2.
Plant transformation
Cotyledonary explants were dipped in an overnight-grown bacterial cell culture for a period of 10 s. Explants were then blotted dry onto a sterilized paper towel, and co-cultivated for a period of 2 days onto a medium consisting of Murashige and Skoog (MS) (1962) salts and vitamins, and supplemented with 0.05 mg/l naphthaleneacetic acid (NAA), 0.25 mg/l kinetin, 2.0 mg/l 2,4-diclorophenoxyacetic acid (2,4-D), and 2.0 g/l casein.
Following co-cultivation, explants were rinsed with sterilized deionized water, blotted dry on a sterilized paper towel, and placed onto a selection medium consisting of MS salts and Gamborg’s B5 vitamins, and supplemented with 0.02 mg/l NAA, 0.5 mg/l 6-benzyladenine (BA), 100 mg/l kanamycin, and 500 mg/l cefotaxime. All explants were incubated in 150 × 100 mm petri plates, and each plate contained 15 explants. All plates were maintained in a controlled environment chamber under a 16 h photoperiod, providing 100 μmol m−2 s−1 light intensity, and at 25°C. Explants were subcultured onto a fresh selection medium once every 2 weeks. Developing KanR shoots, 1 cm in length, were transferred to a rooting medium consisting of MS salts and vitamins, and supplemented with 50 mg/l kanamycin and 250 mg/l cefotaxime. Rooted plantlets were acclimatized and transferred to a greenhouse, and grown under a 16 h photoperiod, of 100 μmol m−2 s−1 light intensity, 30% relative humidity, and at 25°C.
Upon flowering, T0 plants were self-pollinated, and seeds collected upon drying. To obtain T1 plants, seeds were germinated in petri plates containing MS medium supplemented with 100 mg/l kanamycin. A similar protocol was used to obtain T2 plants.
Detection of the LTB gene by PCR and Southern blot analyses
Total DNA from leaf tissues of both transformed and non-transformed (wild-type) plants was isolated according to Michiels et al. (2003). The presence of the transgene(s) was assessed by a duplex PCR assay using two sets of primers to amplify the LTB and nptII genes (LTB-forward, 5′CCATGGGGATCCATGAATAA; LTB-reverse 5′AACTCATCTTTCTCAGAATTTTCCA, and nptII-forward, 5′TATTCGGCTATGACTGGGCA; nptII-reverse, 5′GCCAACGCTATGTCCTGAT). Primers were added, at a final concentration of 1 μM each, to a 50 μl PCR mixture containing 50 ng of purified DNA, 1.5 mM magnesium chloride, 2.5 U of Taq DNA polymerase, and 1 mM dNTPs. PCR reactions were carried out using the following amplification conditions: 94°C for 2 min; 35 cycles consisting of 95°C for 30 s, 58°C for 60 s, 72°C for 60 s; and a final extension of 72°C for 5 min. PCR products were detected by ethidium bromide staining following electrophoresis on a 1.0% agarose gel.
For Southern blot analysis, DNA samples (20 μg) were digested with HindIII, electrophoresed on a 1.0% agarose gel, and then transferred to a positively charged nylon membrane. A DIG-labeled LTB probe was obtained by using the PCR DIG Labeling Mix following the manufacturer’s instructions (Roche Co., Mannheim, Germany). Blots were hybridized overnight at 42°C in a DIG easy hybridization buffer. Then, the membrane was washed in an SSC buffer series (2–0.5×), and a specific signal was detected using a Dig Detection Kit following the manufacturer’s instructions (Roche Co.).
Analysis of transgenic lines for GM1 binding and LTB protein content
Approximately 100 mg of leaf tissue was ground in liquid nitrogen, and homogenized in 500 μl of protein extraction buffer [25 mM sodium phosphate (pH 6.6), 100 mM NaCl, 0.5% Triton X-100 (v/v), 10 g ml−1 leupeptin (w/v), and 1 mM PMSF]. Homogenates were centrifuged at 14,000 rpm for 15 min at 4°C, and total soluble protein (TSP) concentrations were determined according to Bradford (1976). The LTB protein content in the soluble extract was estimated using ganglioside-dependent ELISA (GM1-ELISA), according to Chikwamba et al. (2002). Briefly, assay plates were coated with 1.5 μg of Type III GM1 ganglioside (Sigma, St Louis, MO, USA). After 1 h of incubation at 25°C, plates were washed with PBST and blocked with 5% fat free dry milk for 1 h, also at 25°C. Plates were then washed with PBST and the plant extract was added. These plates were incubated for 1 h at 37°C, washed, and rabbit anti-LTB antiserum (1:1,500 dilution; Biogenesis, Kingston, NH, USA) was added. All plates were incubated for an additional 1 h at 37°C. The secondary antibody, alkaline phosphatase-conjugated mouse anti-rabbit IgG (Sigma), was added, and plates incubated for another 1 h at 25°C. After washing, the substrate, alkaline phosphatase buffer supplemented with 1 mg/ml ρ-nitrophenyl phosphate, was added. After 30 min of incubation, the OD at 405 nm was measured. These ELISA readings were compared with those obtained with purified LTB standards (kindly gifted by Dr. John Clements, Tulane University, New Orleans, LA, USA) to estimate the amount of LTB expressed in transgenic plants.
Western blot analysis was performed using protein extracts that were boiled for 10 min. Gel electrophoresis was carried out in 15% acrylamide using 50 μg of total soluble protein. The gel was blotted onto a nitrocellulose membrane (Amersham Biosciences, Buckinghamshire, UK). After blocking in PBST plus 5% Blocking agent (Amersham Biosciences, Buckinghamshire, UK), the blots were incubated with a rabbit anti-LTB antiserum (1:1,500 dilution) (Biogenesis). A horseradish peroxidase conjugated anti-rabbit antibody (Amersham Biosciences, Buckinghamshire, UK) was applied for 2 h at 25°C. Antibody binding was detected by incubation with a SuperSignal West Dura substrate (Pierce Biotechnology, Rockford, IL) following the manufacturer’s instructions.
Oral immunization with LTB
This feeding study was approved by the Institutional Animal Care and Use Committee, and the animals were handled in accordance with the Mexican federal regulations for animal experimentation and care (NOM-062-ZOO-1999, Ministry of Agriculture, Mexico).
Leaf tissues from mature T1 plants (line L-41) were harvested, freeze-dried, ground, and stored at room temperature. Prior to conducting the immunogenicity study, the LTB was quantified using a GM1-ELISA assay.
The immunogenicity test was carried out using three groups (n = 5) of 12- to 14-week-old male Balb/c mice subjected to the following three treatments, administered orally: (1) 0.260 g of freeze-dried transgenic lettuce tissue resuspended in deionized water; (2) 0.260 g of wild-type lettuce freeze-dried tissue; and (3) 10 μg of pure bacteria-derived LTB. The immunization scheme consisted of three weekly doses, at days 0, 4, and 14, administered to test mice that were subjected to overnight fasting. At day 21, serum samples were obtained from blood extracted by cardiac puncture from ether-anesthetized mice. The small and large intestines were harvested, and intestinal washes were obtained using 3 ml of cold RPMI medium, and these were mixed with 250 μl of 10 mM p-hydroxymercuribenzoate. Samples were centrifuged at 12,000×g at 4°C for 10 min, and the supernatants stored at −70°C until antibody content analysis.
Antibody assays
Antibody levels were determined by indirect ELISA for the production of anti-LTB antibodies. Volumes of 50 μl were used throughout all ELISA assays, and the wells were washed three times between each step using 300 μl of phosphate-buffered saline Tween-20 (PBST). Serum and mucosal samples were diluted 1:50 and 1:2, respectively, and were added to wells previously coated with 5 μg/ml of LTB protein in coating buffer, and blocked with 5% non-fat milk. Labelling was done using alkaline horseradish peroxidase-conjugates of either anti-mouse IgG (Pierce, Rockford, IL), IgA (Zymed Laboratories, San Francisco, CA), IgG1 (biotinylated; Zymed), or IgG2a (biotinylated; Zymed). Plates incubated with biotinylated antibodies for IgG subclass analysis, washed with PBST, and conjugated horseradish peroxidase-streptavidin was added.
Labelling was detected with a substrate solution (0.5 mg/ml o-phenylenediamine, 0.01% H2O2, 50 mM citrate buffer, pH 5.2), and the enzymatic reaction was stopped with 25 μl of 2.5 M H2SO4. The OD at 492 nm was measured in a Multiskan Ascent (Thermo Electron Corporation, Waltham, MA) microplate reader.
Neutralizing activity against CT challenge
The sealed intestinal loop model developed to measure protection against enterotoxin activity was used as described by Richardson et al. (1984). Following immunization with either LTB-producing leaf tissue, WT leaf tissue, or pure LTB, mice were fasted at day 21, but providing water ad libitum.
The mice challenge experiment consisted of intragastric administration of 10 μg of purified cholera toxin (CT) (Sigma–Aldrich Chemical Co., St Louis, MO) diluted in 0.5 ml of 10% NaHCO3. The unchallenged group of mice was treated with 0.5 ml of 10% NaHCO3. After a period of 6 h, mice were euthanized, and the small intestines were ligated, carefully dissected, and weighed. Fluid accumulation (FA), (mg of intestine)/(g of body weight), was calculated for each test animal as follows: FA = G/(B−G) × 1,000, where G is the weight of the gut plus fluid in grams and B is the body weight in grams.
Statistical analysis
ELISA data were expressed as the geometric means of values obtained per group, and the error bars represent standard deviations. Comparisons between absorbance readings and fluid accumulation values were performed using analysis of variance.
Results
Developing transgenic lettuce plants and determining in vitro activity of the LTB recombinant protein
Genetic transformation of lettuce plants was mediated by A. tumefaciens infection using a previously described pBI-LTB vector (Rosales-Mendoza et al. 2007). KanR shoots were observed over a period of 4 weeks, and after 3 months of culture, transformed plants with well-developed roots were established. KanR plantlets from the two lettuce cultivars were successfully regenerated. No differences in number of KanR plantets were observed between the two lettuce cultivars.
A total of 27 transformed lines, 13 from cv. Potosina and 14 from cv. Green Wave, were obtained from independent transformation events, acclimatized, and transferred to the greenhouse.
The presence of LTB and nptII transgenes was confirmed using duplex PCR showing amplicons of the expected molecular weight, 400 and 700 bp, respectively (Fig. 1a). No PCR product was present in reactions containing DNA from untransformed plants. Southern blot analysis confirmed integration of the transgenes in all PCR-positive transgenic lines. Transgenic lines 19 and 39 carried two copies of the transgene; while line 41 carried a single copy of the transgene (Fig. 1b).

a Duplex PCR analysis showing the presence of 400 bp and 700 bp amplicons, corresponding to the LTB and nptII genes, respectively. Lanes 1, genomic DNA from untransformed lettuce plant; 2–5, genomic DNA from T1 independent lettuce transgenic lines, where 2–4 were designated as L-19, L-39, and L-41; 6, positive control for nptII; and 7, positive control for LTB. b Southern blot analysis revealing the insertion of the transgene. Lines L-19, L-39, and L-41 correspond to three representative transgenic lines; WT, untransformed lettuce plant
The ability of the lettuce-derived LTB to bind to the GM1 receptor was assessed using an ELISA assay. All three representative transgenic lines were positive for this assay, thus indicating that LTB retains its quaternary structure. Values obtained in this assay were used to determine the percentage of total soluble plant protein (%TSP) in the sample using a standard curve constructed using a purified LTB.
In general, T0 plants from cv. Potosina showed higher LTB contents, ranging 0.01–0.07%TSP, than those from cv. Green Wave, ranging 0.005–0.07% TSP. Initial ELISA screening identified transgenic lines L19, L39, and L41 from cv. Green Wave as having the highest LTB contents. However, most transgenic lines from cv. Potosina failed to set seeds; while, untransformed plants produced high amounts of seeds.
Following seed production, T1 plants for the high LTB producing lines, all from cv. Green Wave, were successfully grown in kanamycin-containing MS medium, transferred to ex-vitro conditions, and grown in the greenhouse in plastic pots containing a soil mix.
LTB levels in mature leaves of T1 plants from cv. Green Wave reached up to 0.05% TSP (Fig. 2a). Transgenic lines L19 and L41 showed the highest accumulation of LTB. After freeze-drying, leaf tissues were ground, and LTB content was measured by ELISA. The LTB contents were ~20 μg/g for transgenic line L-19 and 30 μg/g for transgenic line L-41.

a GM1-ELISA analysis showing LTB content and GM1 binding. GM1-ELISA using anti-LTB antiserum is carried out to determine LTB content in T1 plants from individual transgenic lines. Total soluble protein (TSP) extracted from T1 mature lettuce plants is used. TSP from wild-type lettuce is used as a negative control. Expression levels of LTB are represented as % TSP. b Western blot analysis showing presence of LTB (12 kDa) in T1 lettuce plants. Lanes 1–3, bacterial LTB protein (20, 50, and 100 ng, respectively); 4, protein extract (50 μg) from line L-19; 5, protein extract (50 μg) from line L-41; and 6, protein extract from untransformed lettuce plant
Western blot analysis using protein extracts from freeze-dried tissues revealed the presence of an immunoreactive protein of the expected molecular weight (12 kDa) as shown in Fig. 2b. When these freeze-dried tissues were stored at room temperature for a period of 1 year, the LTB content did not show any significant decrease in these transgenic lines (data not shown). Therefore, transgenic line L-41 was selected for conducting immunological assays.
In vivo activity
Oral immunogenicity of the lettuce-derived LTB was evaluated in test mice fed water-reconstituted plant tissue containing ~8 μg of lettuce-derived LTB over three dosages, and anti-LTB antibody levels were then determined (Fig. 3). Mice fed wild-type lettuce did not show any detectable signal, and were not significantly different from that of untreated animals. Whereas, mice fed LTB-containing lettuce produced significant levels of anti-LTB antibodies, and these were similar to those elicited by mice administered pure LTB (Fig. 3a). These findings indicated that lettuce-derived LTB was immunogenic following oral delivery in mice.

Anti-LTB antibody responses demonstrating oral immunogenicity of lettuce-derived LTB. Anti-LTB serum IgG (a) and intestinal IgA (b) antibody levels were determined by ELISA. Three weekly doses of either 10 μg of pure recombinant LTB (rLTB) or 8 μg of lettuce-derived LTB (L-LTB), as well as untransformed wild-type lettuce (L-WT) were administered to Balb/c mice by oral feeding. Serum samples and intestinal washes were taken at day 21 and subjected to ELISA analysis. A492 values ± SD from each experimental group (n = 5) are shown. Significant differences (P < 0.05) are indicated as follows: (*) vs. the negative control group (L-WT), (+) vs. the positive control group (rLTB)
Significant anti-LTB IgA antibody responses were elicited by lettuce-derived LTB, but at lower levels than those detected following immunization with pure LTB (Fig. 3b). These results demonstrated that lettuce-derived LTB was an effective mucosal immunogen by the oral route.
The sealed intestinal adult mouse loop assay was used to assess the neutralizing activity of lettuce-derived LTB against oral CT challenge (Fig. 4). In mice fed WT lettuce, a mean fluid accumulation (FA) value of 85.0 mg/g was detected, and this was significantly higher than that observed (53.0 mg/g) in the negative control group; i.e., mice treated with the NaHCO3 solution alone. In contrast, the mean FA value for mice immunized with either pure LTB or lettuce-derived LTB were not significantly different from the negative control group. These findings indicated that oral immunization with lettuce-derived LTB induced protection against CT.

Fluid accumulation observed in mice after challenge with the cholera toxin, and showing the protective effect of lettuce-derived LTB oral immunization. Three weekly doses of pure recombinant LTB (rLTB) or lettuce-derived LTB (L-LTB), as well as untransformed wild-type lettuce (L-WT) were administered to Balb/c mice by oral feeding. At day 21, the L-WT group received 500 μl bicarbonate buffer (L-WT NaHCO3); whereas, the other groups, L-WT, rLTB, and L-LTB, were challenged with 10 μg of cholera toxin (CT) dissolved in the same buffer volume. After 6 h, fluid accumulation (FA) was recorded. Mean FA values ± SD from each group (n = 5) are shown. The cutoff value (mean FA ± SD of the negative control group, cWT/NaHCO3) is indicated by the discontinuous horizontal line, and the mean FA increase above it is noted for each challenged group. Statistically significant differences (P < 0.05), compared to the negative control group (L-WT-), are indicated by an asterisk
T1 seeds were also grown on the selection medium to generate T2 plants. When these plants were evaluated for levels of expression of the LTB transgene, similar levels of LTB to those observed for T1 progeny were detected (data not shown).
Discussion
As LTB has immunoprotective and adjuvant properties, it has been produced in several plants in an effort to assess the feasibility of oral immunization using a low-cost source of the recombinant protein. Different food (potato, rice, soybean, maize, carrot, and watercress) and non-food (tobacco, ginseng) crops have been used as models for the production of LTB (Haq et al. 1995; Mason et al. 1998; Moravec et al. 2007; Chikwamba et al. 2002; Kang et al. 2006; Rosales-Mendoza et al. 2007; Kim et al. 2010; Loc et al. 2011). Kim et al. (2007) have reported on the synthesis and assembly of the LTB protein into oligomeric structures of pentameric size in lettuce, reaching expression levels of up to 1.0–2.0% of the total soluble protein.
Recently, Loc et al. (2011) reported on the expression of an optimized LTB gene codon in watercress. Western blot and GM1-ELISA analyses revealed that the watercress-derived LTB protein was antigenic and assembled into a pentameric form. This was similar to findings obtained in lettuce in this study. However, none of the transgenic watercress plants were transferred to ex vitro conditions, and therefore subsequent generations were not analysed for transgene stability and LTB protein accumulation. Moreover, immunogenicity of watercress-derived LTB protein in test animals was yet to be conducted.
In this study, several transgenic lines from two different cultivars of lettuce have been successfully developed. The frequency of transformation was similar for both cultivars of lettuce, and this was also comparable to that reported by Curtis et al. (1994). Molecular analysis confirmed the presence of LTB and nptII genes in transgenic plants. As expected, different transgenic lines expressed varying levels of the LTB recombinant protein. Levels of LTB in T1 lettuce plants reached up to 0.05% TSP. This content was comparable to those reported previously in potato, reaching up to 0.19% TSP (Mason et al. 1998), and carrot, reaching up to 0.3% TSP (Rosales-Mendoza et al. 2007).
Following freeze-drying of lettuce leaf tissue, LTB levels of up to 30 μg/g were calculated, and these levels remained the same for at least 1 year when freeze-dried tissues were stored at room temperature. Western blot analysis using protein extracts from freeze-dried tissues revealed presence of an immunoreactive protein of the expected molecular weight (12 kDa) as shown in Fig. 2b. When these freeze-dried tissues were stored at room temperature for a period of 1 year, and then reanalyzed by Western blot analysis, there was no significant decrease in levels of the antigenic protein (data not shown). This indicated that freeze-dried lettuce tissue could serve as a reliable form for storing the recombinant LTB protein for at least for 1 year.
In a previous study, LTB accumulation of up to 2% TSP is detected based on ELISA analysis wherein plates have been coated with total protein extracts, and thus both monomeric and oligomeric forms of LTB have been detected (Kim et al. 2007). However, it is important to point out that quantification of the recombinant LTB has not been carried out based on GM1-ELISA. In this study, GM1-ELISA is used to quantify LTB in lettuce-derived leaf tissues as pentamers are the ones with biological activity and induce neutralizing activity.
For those transgenic lettuce lines of cv. Potosina that showed highest levels of LTB content yet failed to set seed, it is likely that this is attributed to toxic effects of the recombinant LTB protein. A similar observation has been reported in transgenic potato lines expressing LTB wherein plants expressing highest levels of LTB resulted in stunting of shoot growth as well as poor tuber yield (Mason et al. 1998). It has been hypothesized that pore-forming LTB qualities influence ion partitioning within plant cell compartments, which in turn could alter growth patterns. This may also be responsible for the observed lack of seed set in our transgenic lettuce lines.
It is interesting to note that LTB levels in the highest-LTB T1 lettuce line were similar to those detected in the T2 progeny, thus suggesting stability of these levels in advanced generations of lettuce. This is in contrast to findings reported on expression of a recombinant antigenic protein, F glycoprotein of the Respiratory syncytial virus (RSV), wherein RSV-F protein levels decreased in advanced generations of tomato (Lau and Korban 2010).
To assess the induction of humoral responses, mice were immunized by the oral route. Results indicated that lettuce-derived LTB from transgenic line L-41 elicited specific systemic and mucosal immune responses following a three weekly-dosing scheme used in this study. Furthermore, a neutralizing activity against the cholera toxin was observed in this study as demonstrated by fluid accumulation following CT challenge. Interestingly, mice immunized with ~8 μg of lettuce-derived LTB were capable of reducing fluid accumulation to levels similar to those in mice immunized with the pure LTB, thus contributing to full protection against CT. To our knowledge, this is the first report wherein challenge against CT has been assayed and protection has been demonstrated in mice. The neutralizing activity of antibodies elicited by the plant-derived LTB has been reported in soybean, rice, corn, and carrot (Chikwamba et al. 2002; Moravec et al. 2007; Rosales-Mendoza et al. 2008; Kim et al. 2010).
This study provides further characterization of the immunogenicity of the lettuce-derived LTB. Further research should assess whether or not lettuce-derived LTB would induce a long-term immune response in test animals. To investigate the efficacy of this vaccination strategy in a more physiologically relevant system, a model for infection by ETEC strains such as the RITARD rabbit model may be used in the future, using the transgenic lettuce line L-41, as this is the only animal model that elicits similar responses to human ETEC infection (Spira et al. 1981).
In recent years, several attempts have been made to develop oral candidate vaccines against ETEC by the expression of relevant antigens, such as CFAs (Yu and Langridge 2001; Lee et al. 2004), a heat labile enterotoxin mutant (Kang et al. 2004), and the atoxic LT derivative LTB (Mason et al. 1998; Chikwamba et al. 2002; Kang et al. 2006; Rosales-Mendoza et al. 2007). In this study, although the low levels of expression of LTB in lettuce leaves remain an obstacle, the ability of LTB-expressing lettuce to induce protection in mice against CT challenge following oral immunization is encouraging for future vaccine development as large amounts of freeze-dried lettuce leaves can be prepared. Moreover, the recombinant LTB protein remains stable over prolonged storage at room temperature.
As LTB also acts as an effective adjuvant, this immunogen could be used in various formulations by mixing freeze-dried lettuce tissues containing LTB with other plant tissues containing unrelated antigens that are otherwise poorly immunogenic for oral administration. These findings indicate that lettuce may be used as a functional and efficient system for inexpensive large scale vaccination against ETEC as well as for formulation of polyvalent oral vaccines.
There is a great need for to develop inexpensive and effective mucosal delivery systems for vaccine formulations against ETEC diarrhea and cholera (Black 1993). Conventional vaccines are expensive, require parenteral administration, as well as availability of cold chain infrastructure. Parenterally delivered vaccines also require availability of large volumes of hypodermic needles and are likely to be inefficacious due to breakdowns in the cold-chain, especially in developing countries where the need is the highest. For all these reasons, needle-free vaccination has become a priority in vaccine development (Giudice and Campbell 2006). An oral-plant delivery system will facilitate vaccine administration as plant-based vaccines are inexpensive, easily stored at room temperatures when freeze-dried, and can be easily administered after reconstitution with water.
References
Black RE (1993) Epidemiology of diarrheal disease: implications for control by vaccines. Vaccine 11:100–106
Bradford MM (1976) A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Chikwamba R, Cunnick J, Hathaway D, McMurray J, Mason H, Wang K (2002) A functional antigen in a practical crop: LT-B producing maize protects mice against Escherichia coli heat labile enterotoxin (LT) and cholera toxin (CT). Transgenic Res 11:479–493
Clements JD, Sack DA, Harris JR, Chakraborty J, Neogy PK, Stanton B, Huda N, Khan MU, Kay BA, Khan MR (1988) Crossprotection by B subunit-whole cell cholera vaccine against diarrhea associated with heat-labile toxin-producing enterotoxigenic Escherichia coli: results of a large-scale field trial. J Infect Dis 158:372–377
Clements JD, van Loon F, Sack DA (1991) Field trial of oral cholera vaccines in Bangladesh: serum vibriocidal and antitoxic antibodies as markers of the risk of cholera. J Infect Dis 163:1235–1242
Curtis IS, Power JB, Blackhall NW, de Laat AMM, Davey MR (1994) Genotype-independent transformation of lettuce using Agrobacterium tumefaciens. J Exp Bot 45:1441–1449
Giudice EL, Campbell JD (2006) Needle-free vaccine delivery. Adv Drug Deliv Rev 58:68–89
Haq TA, Mason HS, Clements JD, Arntzen CJ (1995) Oral immunization with a recombinant bacterial antigen produced in transgenic plants. Science 268:714–716
Hirst TR (1991) Assembly and secretion of oligomerictoxins in gram negative bacteria. In: Alouf JE, Freer JH (eds) A source book of bacterial protein toxins. Academic Press, London
Hoffmeisterová H, Čeřovská N, Tomáš Moravec T, Helena Plchová H, Folwarczna J, J Velemínsky (2008) Transient expression of fusion gene coding for the HPV-16 epitopes fused to the sequence of potyvirus coat protein using different means of inoculation of Nicotiana benthamiana and Brassica rapa, cv. Rapa plants. Plant Cell Tiss Organ Cult 94:261–267
Kang TJ, Han SC, Kim MY, Kim YS, Yang MS (2004) Expression of non-toxic mutant of Escherichia coli heat-labile enterotoxin in tobacco chloroplasts. Protein Expr Purif 38:123–128
Kang TJ, Lee WS, Choi EG, Kim JW, Kim BG, Yang MS (2006) Mass production of somatic embryos expressing Escherichia coli heat-labile enterotoxin B subunit in Siberian ginseng. J Biotechnol 121:124–133
Kapusta J, Modelska A, Pniewski T, Figlerowicz M, Jankowski K, Lisowa O, Plucienniczak A, Koprowski H, Legocki AB (2001) Oral immunization of human with transgenic lettuce expressing hepatitis B surface antigen. Adv Exp Med Biol 495:299–303
Kim YS, Kim B-G, Kim T-G, Kang T-J, Yang M-S (2006) Expression of a cholera toxin B subunit in transgenic lettuce (Lactuca sativa L.) using Agrobacterium-mediated transformation system. Plant Cell Tiss Organ Cult 87:203–210
Kim TG, Kim MY, Kim BG, Kang TJ, Kim YS, Jang YS, Arntzen CJ, Yang MS (2007) Synthesis and assembly of Escherichia coli heat-labile enterotoxin B subunit in transgenic lettuce (Lactuca sativa). Protein Expr Purif 51:22–27
Kim TG, Kim BG, Kim MY, Choi JK, Jung ES, Yang MS (2010) Expression and immunogenicity of enterotoxigenic Escherichia coli heat-labile toxin B subunit in transgenic rice callus. Mol Biotechnol 44:14–21
Kim MY, Kim T-G, Yoo H-S, Yang M-S (2011) Expression and assembly of ApxIIA toxin of Actinobacillus pleuropneumoniae fused with the enterotoxigenic E. coli heat-labile toxin B subunit in transgenic tobacco. Plant Cell Tiss Organ Cult. doi:10.1007/s11240-010-9877-3
Klipstein FA, Engert RF, Clements JD, Houghten RA (1984) Differences in cross-protection in rats immunized with the B subunits of cholera toxin and Escherichia coli heat-labile toxin. Infect Immun 43:811–816
Landell FGC, Rivera GG, Flores TJO, Martínez MB, Orozco EO (2008) Expression and functional analysis of an Entamoeba histolytica truncated adhesin in tomato plants. Plant Cell Tiss Organ Cult 94:1–9
Lau JM, Korban SS (2009) Analysis and stability of the Respiratory Syncytial Virus antigen in a T3 generation of transgenic tomato plants. Plant Cell Tiss Organ Cult 96:335–342
Lee JY, Yu J, Henderson D, Langridge WH (2004) Plant-synthesized E. coli CFA/I fimbrial protein protects Caco-2 cells from bacterial attachment. Vaccine 23:222–231
Loc NH, Song NV, Tien NQD, Minh TT, Nga PTQ, Kim T-G, Yang M-S (2011) Expression of the Escherichia coli heat-labile enterotoxin B subunit in transgenic watercress (Nasturtium officinale L.). Plant Cell Tiss Organ Cult. doi:10.1007/s11240-010-9835-0
Mason HS, Haq TA, Clements JD, Arntzen CJ (1998) Edible vaccine protects mice against Escherichia coli heat-labile enterotoxin (LT): potatoes expressing a synthetic LT-B gene. Vaccine 16:1336–1343
Mason H, Warzecha H, Mor T, Arntzen C (2002) Edible plant vaccines: applications for prophylactic and therapeutic molecular medicine. Trends Mol Med 8:324–329
Michiels A, Van den Ende W, Tucker M, Van Riet L, Van Laere A (2003) Extraction of high-quality genomic DNA from latex-containing plants. Anal Biochem 315:85–89
Moravec T, Schmidt MA, Herman EM, Woodford-Thomas T (2007) Production of Escherichia coli heat labile toxin (LT) B subunit in soybean seed and analysis of its immunogenicity as an oral vaccine. Vaccine 25:1647–1657
Peltola H, Siitonen A, Kyronseppa H, Simula I, Mattila L, Oksanen P, Kataja MJ, Cadoz M (1991) Prevention of travellers’ diarrhoea by oral B-subunit/whole-cell cholera vaccine. Lancet 338:1285–1289
Qadri F, Wenneras C, Ahmed F, Azaduzzamar M, Saha D, Albert MJ, Sack RB, Svennerholm AM (2000) Safety and immunogenicity of an oral, inactivated enterotoxigenic Escherichia coli plus cholera toxin B subunit vaccine in Bangladeshi adults and children. Vaccine 18:2704–2712
Rezaee MA, Rezaee A, Moazzeni SM, Salmanian AH, Yasuda Y, Tochikubo K, Pirayeh SN, Arzanlou M (2005) Expression of Escherichia coli heat-labile enterotoxin B subunit (LTB) in Saccharomyces cerevisiae. J Microbiol 43:354–360
Richardson SH, Giles JC, Kruger KS (1984) Sealed adult mice: new model for enterotoxin evaluation. Infect Immun 43:482–486
Rosales-Mendoza S, Soria-Guerra RE, de Jesus Olivera-Flores MT, Lopez-Revilla R, Arguello-Astorga GR, Jimenez-Bremont JF, Garcia-de la Cruz RF, Loyola-Rodriguez JP, Alpuche-Solis AG (2007) Expression of Escherichia coli heat-labile enterotoxin b subunit (LTB) in carrot (Daucus carota L.). Plant Cell Rep 26:969–976
Rosales-Mendoza S, Soria-Guerra RE, López-Revilla R, Moreno-Fierros L, Alpuche-Solís A (2008) Ingestion of transgenic carrots expressing the Escherichia coli heat-labile enterotoxin B subunit protects mice against cholera toxin challenge. Plant Cell Rep 27:79–84
Sack RB, Gorbach SL, Banwell JG, Jacobs B, Chatterjee BD, Mitra RC (1971) Enterotoxigenic Escherichia coli isolated from patients with severe cholera-like disease. J Infect Dis 123:378–385
Spangler BD (1992) Structure and function of cholera toxin and the related Escherichia coli heat-labile enterotoxin. Microbiol Rev 56:622–647
Spira WM, Sack RB, Froehlich JL (1981) Simple adult rabbit model for Vibrio cholerae and enterotoxigenic Escherichia coli diarrhea. Infect Immun 32:739–747
Webster DE, Smith SD, Pickering RJ, Strugnell RA, Dry IB, Wesselingh SL (2006) Measles virus hemagglutinin protein expressed in transgenic lettuce induces neutralising antibodies in mice following mucosal vaccination. Vaccine 24:3538–3544
Yu J, Langridge WH (2001) A plant-based multicomponent vaccine protects mice from enteric diseases. Nat Biotechnol 19:548–552
Acknowledgments
This study was funded by grants no. 37048-B, 56980 and 102109 from CONACYT. The authors wish to thanks Dr. John Clements for providing us with the pure LTB protein.
Author information
Authors and Affiliations
Corresponding author
Additional information
Luzmila Martínez-González and Sergio Rosales-Mendoza are contributed equally to this work.
Rights and permissions
About this article
Cite this article
Martínez-González, L., Rosales-Mendoza, S., Soria-Guerra, R.E. et al. Oral immunization with a lettuce-derived Escherichia coli heat-labile toxin B subunit induces neutralizing antibodies in mice. Plant Cell Tiss Organ Cult 107, 441–449 (2011). https://doi.org/10.1007/s11240-011-9994-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11240-011-9994-7