
Abstract
Improving quality is an essential goal of rice breeding and production. However, rice quality is not solely determined by genotype, but is also influenced by the environment. Phenotype plasticity refers to the ability of a given genotype to produce different phenotypes under different environmental conditions, which can be a representation of the stability of traits. Seven quality traits of 141 hybrid combinations, deriving from the test-crossing of 7 thermosensitive genic male sterile (TGMS) and 25 restorer lines, were evaluated at 5 trial sites with intermittent sowing of three to five in Southern China. In the Yangtze River Basin, it was observed that delaying the sowing time of hybrid rice combinations leads to an improvement in their overall quality. Twelve parents were identified to have lower plasticity general combing ability (GCA) values with increased ability to produce hybrids with a more stable quality. The parents with superior quality tend to exhibit lower GCA values for plasticity. The genome-wide association study (GWAS) identified 13 and 15 quantitative trait loci (QTLs) associated with phenotype plasticity and BLUP measurement, respectively. Notably, seven QTLs simultaneously affected both phenotype plasticity and BLUP measurement. Two cloned rice quality genes, ALK and GL7, may be involved in controlling the plasticity of quality traits in hybrid rice. The direction of the genetic effect of the QTL6 (ALK) on alkali spreading value (ASV) plasticity varies in different cropping environments. This study provides novel insights into the dynamic genetic basis of quality traits in response to different cropping regions, cultivation practices, and changing climates. These findings establish a foundation for precise breeding and production of stable and high-quality rice.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Rice is one of the important staple crops globally, and hybrid rice technology has greatly increased rice production, ensuring global food security. Along with the progression of society and the enhancement of living standards, there has been a gradual increase in the demand for rice quality (Li et al. 2023). Rice quality is also an important and complex trait, including eating and cooking quality, milling quality, apparent quality, and nutritional quality. The quality of rice directly impacts its commercial value and palatability. Improving the quality of hybrid rice has always been the goal pursued by breeders. Over the years, both traditional and modern molecular breeding techniques have been employed to enhance the quality of hybrid rice steadily (Tian et al. 2009; Zhang et al. 2016). However, the quality traits of rice are susceptible to environmental factors such as light, temperature, and humidity, resulting in variations (Liu et al. 2013; Lu et al. 2022). Therefore, how to mitigate the impact of environmental factors on rice quality and enhance its stability is also one of the concerns in improving hybrid rice.
Phenotype plasticity refers to the ability of the same genotype to produce different phenotypes in different environments (Sultan 2000), reflecting the relationship between organisms and their environment, which is widely present in plants (Bradshaw 1965). Phenotype plasticity is related to the adaptability and stability of plants (Chevin et al. 2013; Finlay and Wilkinson 1963). From an evolutionary perspective, varieties with high phenotype plasticity exhibit stronger adaptability to the environment (Des Marais et al. 2013; Bonamour et al. 2019). However, in the context of crop production, plants with lower phenotype plasticity exhibit greater stability. Consequently, implementing techniques to decrease the phenotype plasticity of crops and enhance their stability becomes crucial in enabling the expression of desired traits across a broader range of locations.
Phenotype plasticity is under genetic control and can be targeted for artificial improvement in crop breeding (Gage et al. 2017). To achieve this goal, based on scientific quantification methods for plasticity, efforts have been made to study the genetic architecture of crop plasticity and analyze the QTLs across various crops (Wang et al. 2015a; Kadam et al. 2017; Jin et al. 2023). Exploratory studies on rice have revealed the genetic structure and potential QTLs underlying the plasticity of yield-related traits (Kikuchi et al. 2017; Mu et al. 2022). Although the precise functions of these QTLs remain unclear, they provide new insights and methods for artificially selecting and improving crop phenotype plasticity. Quality-related traits are influenced by the environment, exhibiting phenotype plasticity. However, previous studies on phenotype plasticity in rice have primarily focused on yield-related traits, but there has been limited research on the patterns and genetic architecture of phenotype plasticity in quality traits, which are crucial for breeding advancements. Enhancing rice quality stability has implications for enhancing the potential and commercial value of rice varieties. Therefore, understanding the patterns of phenotype plasticity and genetic structure of quality-related traits in rice will provide better references for breeding improvements.
Parents selection is crucial in hybrid rice breeding (Chen et al. 2019). However, the challenge of selecting ideal parental materials from a large population to induce strong heterosis is significant. Therefore, breeders use the combining ability to assess the breeding value of parental materials in hybrid production (Sprague and Tatum 1942). By identifying the combining ability of parents in phenotype plasticity, breeders can predict the performance of hybrid combinations, thereby enhancing the efficiency and stability of hybrid rice production (Abd El-Aty et al. 2022).
The large-scale phenotypic analysis is an important foundation for plasticity research. In this study, a total of 141 hybrid rice combinations were obtained from 7 TGMS lines and 25 restorer lines. These combinations were planted in five locations in Southern China in the 2020 summer season and arranged 3 to 5 intermittent sowings in each trial location. Seven quality traits and their phenotype plasticity were investigated, including amylose content (AC), alkali spreading value (ASV), gel consistency (GC), chalkiness degree (CD), percentage of grains with chalkiness (PGWC), transparency (TP), and milled rice ratio (MRR). We analyzed the combining ability of phenotype plasticity for 32 parental materials and elucidated the genetic structure of phenotype plasticity for quality-related traits. Genetic effects and candidate gene analyses were conducted on the identified QTLs. The statistical results suggest that delaying sowing date is beneficial for enhancing rice quality in the Yangtze River basin. We utilized a model to evaluate the phenotype plasticity of each hybrid and identify TGMS and restorer lines that exhibit improved quality stability in hybrid rice breeding. Furthermore, our research uncovered the genetic basis of phenotypic plasticity in rice quality traits and discovered QTLs associated with quality plasticity while predicting candidate genes. These findings offer theoretical guidance for determining the optimal sowing date of high-quality rice and enhancing the quality stability of hybrid rice.
Materials and methods
Plant materials and field experiments
In this study, a total of 141 hybrid rice combinations were obtained by crossing 7 TGMS lines with 25 restorer lines. The TGMS lines include Jing4155S (J4155S), LongZhen36S (LZ36S), HuaYue468S (HY468S), ZhenXiangS (ZXS), HuaXuan302S (HX302S), LongKe638S (LK638S), and HuaWei338S (HW338S). The restorer lines include HuaHui8612 (HH8612), HuaHui7810 (HH7810), HuaHui3135 (HH3135), HuaHui7503 (HH7503), TaiSi (TS), LongKeSiMiao13 (LKSM13), HuaHui8012 (HH8012), HuaHui2646 (HH2646), HuaHui6210 (HH6210), HuaHui3748 (HH3748), DiZhan (DZ), HuaHui8037 (HH8037), HuaHui5106 (HH5106), HuaHui8549 (HH8549), YuZhan (YZ), HuaHui8700 (HH8700), HuaHui8954(HH8954), HuaHui4153 (HH4153), R1206, R1212, R1377, R1988, WuShanSiMiao (WSSM534), HuaZhan (HZ), and Jin4 (J4). All parental lines are elite parents in hybrid rice production and sourced from the Core Germplasm Bank of Commercial Hybrid Rice Breeding in Yuan Longping High-tech Agriculture Co., Ltd. (Supplementary Table S1, S2).
The field experiments were conducted in the summer season of 2020 in five locations including SC-GH (Guanghan, Sichuan Province, 104°25′ E, 30°99′ N), HN-LS (Lingshui, Hainan Province, 109°45′ E, 18°22′ N), HN-CS (Changsha, Hunan Province, 112°59′ E, 28°12′ N), HB-EZ (Ezhou, Hubei Province, 114°52′ E, 30°23′ N), and AH-HF (Hefei, Anhui Province, 117°17′ E, 31°52′ N). Among the five trial locations, HN-LS is situated in the south China rice cropping region, SC-GH is located in the upper Yangtze River rice cropping region, while AH-HF, HN-CS, and HB-EZ are situated in the middle and lower Yangtze River rice cropping region. In SC-GH, three intermittent sowings were arranged from April 1 to May 1, with a 10-day interval between sowings. In HB-EZ and AH-HF, four intermittent sowings were arranged from April 15 to June 1, with a 15-day interval between sowings. In HN-CS, five intermittent sowings were arranged from April 10 to June 10, with a 15-day interval between sowings. In HN-LS, four intermittent sowings were arranged from June 15 to July 30, with a 15-day interval between sowings (Supplementary Table S3). Seedlings of 5-leaf age were transplanted. Each material was planted in a five-row plot with eight individuals in each row at a spacing of 20 cm × 26.5 cm density in the field and standard management practices throughout the growing period. At maturity, nine uniform plants in the middle of each plot were harvested.
Parental DNA extraction and whole genome sequencing
After germination of parental seeds, young leaves were collected and immediately flash-frozen in liquid nitrogen, and then the samples were stored at − 80℃ for future use. DNA extraction was performed using the FastPure Plant DNA Isolation Mini Kit (Vazyme, Jiangsu, China). The concentration of the extracted DNA was evaluated using a NanoDrop spectrophotometer (NanoDrop Technologies, Wilmington, DE, USA) and a Qubit 3.0 fluorometer (Life Technologies, Carlsbad, CA, USA). To assess the purity and integrity of the DNA, 1% agarose gel electrophoresis was conducted. For library preparation, a short-read library with a DNA-fragment insert size of 200–400 bp was generated using 1 μg of genomic DNA. The library preparation was carried out following the manufacturer’s instructions using a library preparation kit compatible with DNBSEQ-T7 (BGI, Shenzhen, China). Subsequently, paired-end (PE) sequencing was performed on a DNBSEQ-T7 platform using the PE 150 model.
Measurement of quality trait
A minimum of 200 g seeds from each accession were used for measuring the seven quality traits, including AC, ASV, GC, CD, PGWC, TP, and MRR. The CD, PGWC, and TP were measured according to NY/T 2334–2013 (Chinese Ministry of Agriculture Standards). CD, PGWC, and TP were measured with a Microtek Scan Wizard EZ scanner and rice quality analyzer SC-E software (Hangzhou Wanshen Detection Technology Co., Ltd., Hangzhou, China). The AC, GC, and ASV were measured according to Chinese National Standards GB/T 15683–2008(GB/T 22294–2008 and NY/T 83–2017), respectively.
Measurement of overall quality and meteorological condition
To evaluate the overall quality of the hybrids, we followed the quality standards for cooking rice varieties set by the Chinese Ministry of Agriculture and Rural Affairs (NY/T 593–2021). Here, a record is specified as the quality performance of a specific hybrid, which has been planted at a trial location on a particular sowing date. We assessed the quality of all 2774 records in our experiment by applying the standard to each of them.
In order to analyze the correlation between quality characteristics and meteorological conditions, we have also summarized the meteorological data collected at different times within each day into the following meteorological factors: minimum temperature, maximum temperature, average temperature, day/night temperature difference, surface solar radiation, accumulated rainfall, and number of rainy days (Cheng and Zhu 1998). In our analysis, we consider a day with more than 1 mm of rainfall as a rainy day. Spearman correlation was conducted between each quality trait and meteorological factors using the SciPy library in Python.
Analysis of phenotype plasticity
According to the reaction norm model, the phenotypic record \({y}_{ij}\) of the \(i\) th hybrid line observed under the \(j\) th environment can be modeled as follows:
where \(\mu\) is the mean value of the trait, \({g}_{i}\) is the main effect of the \(i\) th line, and \({h}_{j}\) is the main effect of the \(j\) th environment. \({b}_{i}{h}_{j}\) represents the interaction term (G-by-E) between \(i\) th hybrid line (genotype) and \(j\) th environment. \({\epsilon }_{ij}\) is the error term.
Here, we introduced the Finlay–Wilkinson regression (FW) as the representation of phenotype plasticity. FW reorganized Eq. 1 into follows:
If we consider \({h}_{j}\) as the independent variable of the function\({y}_{ij}=f({h}_{j})\), then the slope of this function \((1+{b}_{i })\) indicates the expected trait variation (\(\Delta {y}_{i}\)) in traits with changes in unit environmental effects (\(\Delta {h}_{j}=1\)), capable of indicating the phenotype plasticity.
To calculate the plasticity of quality traits at different locations and sowing stages, we selected the hybrid lines with multi-location field trials and at least three intermittent sowings per location from the 141 hybrids. We used the FW package implemented in R by Lian et al. (Lian and De Los Campos 2016) and applied the Bayesian method for regression.
Analysis of combining ability
Combining ability analysis is a statistical technique employed in plant breeding to assess the genetic potential of parents and predict the performance of their hybrids. The analysis considers both general combining ability (GCA), which measures the average performance of a parent across different crosses, and specific combining ability (SCA), which measures the performance of a specific parent in specific crosses. Since there are no extra replications for each sowing management in our experiment, we could not properly evaluate the SCA for each hybrid. For convenience, we simply treated the SCA as 0. Consequently, the phenotype plasticity of hybrid \({p}_{ij}\), which has parents i and j, can be modeled as follows:
where \(\mu\) is the mean value of the phenotype plasticity, \(\epsilon\) is the error term, and \({GCA}_{i}\) represents the breeding value of phenotype plasticity for \(i\) th parent. Since the distribution of \({p}_{ij}\) has a mean value of \(\mu\), the sum of the GCAs of all parents is 0 (i.e., \({\sum }_{i}{GCA}_{i}=0\)). We used the sommer package (Covarrubias-Pazaran 2016) in R to calculate the GCA in a mixed model, in which the parents and crosses are considered as random effects. If there are no significant additive effects detected from the parents for the plasticity of hybrid traits, the model will indicate a GCA of 0, which will be later displayed in the paper.
Analysis of BLUP
The analysis for various traits of each location is conducted by fitting a linear mixed model and computing best linear unbiased predictors (BLUPs) with the lme4 R package (Bates et al. 2015):
where Y represents the phenotypic records, the parentheses indicate random effects, “1|” denotes groups, and “:” refers to interactions. LINE indicates to the hybrid lines, and ENV indicates to different sowing stages or different locations. We utilized the fitted random effects of hybrid lines as the representation of the overall genetic effects of each genotype for these traits while eliminating the environmental effects.
Variant detection and annotation
The sequencing data was aligned to the Oryza sativa Nipponbare reference genome (IRGSP1.0) using BWA mem (Li 2013). PCR and optical duplicates were removed using Picard software (Broad Institute). SNP and InDel detection were performed using GATK HaplotypeCaller (McKenna et al. 2010). To minimize false positives, SNPs that did not meet the following criteria were filtered out: QUAL < 30.0, QD < 2.0, SOR > 3.0, FS > 60.0, MQ < 40.0, MQRankSum < 12.5, or ReadPosRankSum < 8.0. Similarly, InDels that did not meet the following criteria were filtered out: QUAL < 30.0, QD < 2.0, FS > 200.0, MQ < 40.0, or ReadPosRankSum < 20.0. SNPs and InDels were annotated using the software SnpEff (Cingolani et al. 2012) based on their impact on genes. The annotation categorized them into different types, such as synonymous mutations, non-synonymous mutations, frameshift mutations, and others. SnpEff utilized Ensembl release 41 as the gene annotation version for this analysis.
Genome-wide association analysis
The GEMMA (Zhou and Stephens 2014) software was utilized to conduct Genome-Wide Association Studies (GWAS) for BLUP and FW values of different traits and locations, with a mixed linear model (MLM) fitted. The kinship matrix was used as a random effect, while the first three principal components from principal component analysis (PCA) were included as fixed effects in the MLM. PCA analysis was performed using the smartPCA program implemented in the Eigensoft package (Patterson et al. 2006). The significance threshold for GWAS analysis was determined using the Bonferroni correction method, which means dividing 0.05 by the number of SNPs (n) in the analysis. Here, the threshold was set at p < 5.32 × 10−8 (i.e., − log10(p) < 7.27). The entire genome was partitioned into LD blocks based on an LD (linkage disequilibrium) threshold of r2 = 0.6 using the gpart R package (Kim et al. 2019). LD blocks with a distance of less than 1 Mb are merged. LD blocks that contain significant SNPs were considered as candidate QTLs.
Results
Effect of sowing date and meteorological factors on quality of hybrid rice
The rice quality traits of the hybrids collected from 5 trial locations, with multiple intermittent sowing arrangements, were measured, and the distributions of the trait values are illustrated in Fig. 1a and Supplementary Table S4. Nearly all quality traits in these rice cropping regions exhibited significant correlations with the sowing date, suggesting a general contribution of sowing date to rice quality trait variation (Supplementary Table S5). As sowing dates were delayed, we observed the mean of AC increasing by 2.37–3.97% in AH-HF, 3.82–5.12% in HN-CS, 4.37–5.32% in HB-EZ, and 0.77–0.89% in SC-GH, while decreasing 0.32–0.75% in HN-LS. In contrast, the mean of CD decreased by 1.31–6.2 in AH-HF, 2.71–5.03 in HN-CS, 2.85–7.82 in HB-EZ, and 2.34–2.07 in SC-GH, while increasing by 0.84–1.89 in HN-LS in tandem with the delay in sowing dates (Fig. 1b–g).

Variation in the trait of hybrid rice combinations across different locations and sowing stages in the southern rice region of China. a The five surveyed sites span China’s southern rice region, where 7 quality traits were phenotyped for hybrid rice varieties. b–g Boxplot of seven quality traits including b chalkiness degree (CD), c alkali spreading value (ASV), d amylose content (AC), e gel consistency (GC), f percentage of grains with chalkiness (PGWC), and g milled rice ratio (MRR) measured at the five sites in different sowing stages. h–j The number of rice varieties reached quality grade 3 or superior quality according to NY/T 593–2021 at each trial location on different sowing dates
We assessed the overall quality of the hybrids by referring to the Chinese Ministry of Agriculture and Rural Affairs’s quality standard for cooking rice variety (NY/T 593–2021). Out of the 2774 records examined, 872 of them exhibited high-quality with grade 3 or superior performance. These high-quality records are sourced from 124 hybrid varieties and shown in all five trial locations and all sowing dates. For all trial locations except HN-LS, the improvement of rice quality correlates with the postponement of sowing dates, with a greater number of records achieving quality grade 3 or higher grades (Fig. 1h–l).
To assess the meteorological variation more precisely, we decomposed it into the following meteorological factors: minimum temperature, maximum temperature, average temperature, day/night temperature difference, surface solar radiation, accumulated rainfall, and number of rainy days (Cheng and Zhu 1998) (Supplementary Fig. S1). Existing studies have shown that the meteorological factors during the grain filling stage, particularly the initial 15 days following full heading, have a considerable influence on rice quality (Wu et al. 2016; Yan et al. 2021). Thus, we employed the mean values of the aforementioned meteorological factors during this time period for further evaluation. We performed an analysis to investigate the correlation between meteorological factors and quality traits. Due to the non-normal distribution of the quality traits data, we computed Spearman correlation coefficients between meteorological factors and these quality traits (Fig. 2). Nearly all quality traits displayed a significant correlation with multiple meteorological factors in each trial location. The correlation trend of each meteorological factor with each quality trait differed across different trial locations. More specifically, at HN-LS, the minimum, maximum, and average temperatures exhibit significant positive and negative correlations with AC and GC, respectively. At the other four locations, there were significant negative and positive correlations between these meteorological factors and AC and GC, respectively. Likewise, at HN-LS, surface solar radiation was positively correlated with AC and negatively correlated with GC. However, at the other four trial locations, contrary correlation trends were observed. In terms of rainy days and accumulated rainfall, our research suggests that increased rainy days are potentially associated with lower levels of AC and higher levels of CD. The significant impact of accumulated rainfall on quality traits was observed, but a more general pattern was not observed in our experiment. Moreover, we considered the rice quality grade as a trait and investigated its correlation with meteorological factors as a reference. For trial locations in the rice cropping region of the upper, middle, and lower reaches of the Yangtze River (i.e., SC-GH, AH-HF, HN-CS, and HB-EZ), there exists a significant correlation between better rice quality and the minimum, maximum, and average temperatures.

The heatmap illustrates Spearman’s correlation coefficients between meteorological factors and quality traits across five trial locations. T denotes temperature. The green color of each box indicates positive relationships between meteorological factors and the corresponding quality trait, while the red color indicates negative relationships. Significant correlations are denoted by asterisks, as determined by a two-tailed t-test (**p < 0.001; ***p < 0.0001). r stands for correlation coefficients
General combining ability analysis for phenotype plasticity of quality traits
In this study, we evaluated the stability of quality traits for 141 hybrid combinations across five trial locations using the Finlay-Wilkinson regression (FW), a measurement of phenotypic plasticity (Supplementary Table S6) (Fig. 3). The combining ability for phenotypic plasticity was conducted to identify potential parents capable of producing hybrid combinations with stable quality traits. Figure 4 shows the GCA value of the hybrid parents in relation to the plasticity of the seven quality traits. Notably, parents with a low GCA value for plasticity are more likely to derive hybrid combinations with stable quality performance.

The boxplot illustrates the phenotype plasticity of hybrid rice across five trial locations for the following traits: a chalkiness degree (CD), b percentage of grains with chalkiness (PGWC), c transparency (TP), d milled rice ratio (MRR), e amylose content (AC), f alkali spreading value (ASV), g gel consistency (GC)

The heatmap illustrates the GCA value for phenotype plasticity in quality traits, as observed between TGMS and restorer lines across various trial locations. Each grid number represents the GCA value for the trait. The red grid denotes a positive GCA value, the blue grid denotes a negative GCA value, and the white grid denotes a zero GCA value
Three TGMS lines (ZXS, HX302S, and HY468S) and nine restorer lines (HH8012, HH7503, DZ, LKSM13, HH2646, HH5106, HH8549, YZ, and WSSM534) consistently exhibited comprehensive low GCA values (≤ 0) for plasticity across more than three trial locations. It indicates their potential to consistently produce hybrid combinations with stable quality traits across different cropping regions and varied sowing management. Interestingly, parents with superior quality tend to exhibit low GCA values for plasticity. The 12 parental lines with low plasticity GCA value are all of high quality and have been widely used in high-quality hybrid rice development. We analyzed the quality statistics from yield testing trials of nationally approved hybrid rice varieties derived from all tested parental lines from 2016 to 2022. Notably, the hybrid rice varieties derived from parents with low plasticity GCA values exhibited a higher rate of high quality compared to those derived from parents with high plasticity GCA values (Supplementary Table S7).
Genome-wide association analysis for phenotype plasticity and BLUP measurement of quality traits
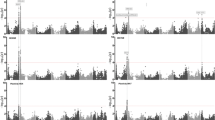
The genome-wide association study (GWAS) was conducted independently for the plasticity of each quality trait across different trial locations. This analysis revealed a total of 13 QTLs associated with grain quality plasticity (Fig. 5a–b; Table 1). Among these, four QTLs were detected in two trial locations, two QTLs were detected in three trial locations, and the remaining seven QTLs were only detected in a single trial location. These findings underscore the complex genetic basis of plasticity for quality traits in different regions.

QTLs associated with plasticity and BLUP for seven traits across five trial locations. a QTLs associated with the plasticity of different traits across different locations. Each dot represents an SNP, and the size of the dot is proportional to its − log10(p)-value, as indicated on the right. Loci with a − log10(p)-value exceeding the genome-wide significance threshold are highlighted in red. b Manhattan plot overlays GWAS results of plasticity for the seven traits across five locations. The black horizontal dashed line indicates Bonferroni-corrected genome-wide significance, and the vertical gray lines indicate the positions of detected QTLs. c Manhattan plot overlays GWAS results of BLUP for the seven traits across five locations. The vertical red lines indicate the positions of detected QTLs that overlap in both plasticity and BLUP. d QTLs associated with the BLUP of different traits across different locations
In the case of AC plasticity, we identified six QTLs were identified on chromosomes 3, 9, 10, 11, and 12. Among these QTLs, four were identified in both HN-CS and AN-HF, while two were exclusively identified in HN-CS. As for CD plasticity, we detected five QTLs were detected on chromosomes 2, 3, 4, 7, and 8. Notably, all QTLs were identified in a single trial location, either HB-EZ or AN-HF.
We identified QTL6 on chromosome 6 as being associated with the plasticity of two key taste quality traits (ASV and GC). The effect of QTL6 on ASV plasticity was detected in both AH-HF and SC-GH, while its effect on GC plasticity was observed exclusively in HB-EZ. These findings suggest that the plasticity of ASV and GC may share a similar genetic basis.
A single QTL, QTL7, located on chromosome 7, was found to be associated with the plasticity of MRR. This association was observed across three trial locations including HB-EZ, HN-CS, and SC-GH.
Furthermore, we employed the best linear unbiased prediction (BLUP) approach to mitigate the impact of non-genetic factors and calculate the genetic effect values for these quality traits (Supplementary Table S8). We also conducted a GWAS for the BLUP measurement. This analysis identified a total of 15 QTLs for the BLUP measurement of seven quality traits across five trial locations (Fig. 5c–d, Supplementary Table S9). Among these QTLs, seven QTLs (QTL2, QTL3, QTL6, QTL7, QTL11, QTL12, and QTL13) were also identified in the previous GWAS of plasticity (Supplementary Table S10). These overlapping QTLs, identified in both GWAS results, could play a crucial role in regulating quality traits and responding to the quality plasticity observed in diverse cropping environments.
Analysis of QTLs genetic effects and prediction of candidate genes
We evaluated the genetic effects of all 13 plasticity QTLs (Fig. 6a–f, Fig. S2). Among these, two QTLs (QTL6 and QTL7) were detected in three trial locations, thereby establishing them as stable major QTLs for rice quality plasticity. As these two QTLs also demonstrated an effect on the BLUP measurement, further investigation into the genetic effects of these QTLs could lay a foundation for molecular breeding strategies aimed at enhancing both the quality and stability of hybrid rice.

Genetic effects of QTLs and prediction of candidate genes. a–f Boxplots of plasticity of quality traits in hybrid combinations containing the different leading SNPs of QTLs. a Plasticity of GC in hybrid combinations containing the different leading SNPs of QTL6 in HB-EZ. b–c Plasticity of ASV in hybrid combinations containing the different leading SNPs of QTL6 in AH-HF and SC-GH. d–f The plasticity of MRR in hybrid combinations containing the different leading SNPs of QTL7 in HB-EZ, HN-LS, and SC-GH. Uppercase letters indicate statistically significant differences at p < 0.01; lowercase letters indicate statistically significant differences at p < 0.05. g The relationships between the QTL6 leading SNP and its candidate gene. h The haplotypes identified by combinations of QTL6 leading SNP and candidate gene FNPs. i The relationships between the QTL7 leading SNP and its candidate gene
The QTL6 was found to be associated with ASV and GC. The leading SNP (Chr6:6,722,905) of QTL6 had a − log10 (p value) of 7.83 in AH-HF, 13.61 in HB-EZ and 31.03 in SC-GH. Three genotypes of the leading SNP of QTL6 were identified. A significance analysis revealed that CC genotypes showed significantly lower GC plasticity compared to CT and TT genotypes in HB-EZ. It is worth noting that the direction of the genetic effects of the identical ASV plasticity allele varied across different cropping environments. In the AH-HF, CC genotypes exhibited significantly lower ASV plasticity compared to varieties with CT and TT genotypes. Conversely, in the SC-GH, CC genotypes showed significantly higher ASV plasticity compared to varieties with CT and TT genotypes (Fig. 6a–c). The QTL7 was associated with MRR. The leading SNP (Chr7:24,665,290) of QTL7 had a − log10 (p value) of 15.69 in HN-LS, 9.93 in HB-EZ, and 7.67 in SC-GH. The leading SNP alleles exhibited significantly different MRR plasticity, as shown in Fig. 6d–f. Notably, the AA genotypes displayed the lowest MRR plasticity value.
Within the associated genomic region of the QTL6 and QTL7, a total of 6 and 18 annotated genes were found, respectively. Among these genes, ALK and GL7, known for their functional annotations in controlling rice quality, were pinpointed as candidate genes for further analysis in rice quality plasticity. The ALK gene, located within QTL6, is a key gene controlling rice gelatinization temperature which encodes the soluble starch synthase II-3. The GL7/GW7 gene, located within QTL7, encodes a protein that is homologous to the LONGIFOLIA proteins found in Arabidopsis thaliana. This protein is known to regulate cell elongation, thereby affecting grain length and grain shape.
Discussion
Rice quality is one of the important goals in hybrid rice breeding improvement. The formation of rice grain quality is under genetic control and also influenced by the environment. Numerous studies have been undertaken to explore the impact of sowing dates on rice quality. However, due to differences in location, tested varieties, climatic conditions, and other factors, the outcomes of these studies lack consistency. Yao et al. observed that a delay in the sowing date leads to an improvement in the appearance quality of rice, but results in a decrease in the eating and cooking quality (Yao et al. 2011). Wang et al. found that a delay in the sowing date causes the CD, PGWC, AC, and GC to exhibit a declining trend (Wenting et al. 2021).
In this study, we analyzed the rice quality across multiple sowing dates in five trial regions located in Southern China and gathered the meteorological data for each region. It was observed that in the Yangtze River basin trials, the amount of high-quality rice (≥ grade 3 set by the Chinese Ministry of Agriculture) increases as the sowing date is delayed. The different sowing date essentially influences the weather conditions in rice growing. Previous studies indicated that an inappropriate sowing date will expose rice to unfavorable climatic conditions during the grain filling stage, resulting in a decline in rice quality (Cheng and Zhong 2001).
It has been proved that light, temperature, and rainfall during the grain filling stage are the pivotal climatic factors influencing rice quality (Resurreccion et al. 1977; Cheng and Zhu 1998). This study quantified and analyzed several crucial meteorological factors during the grain filling stage to assess their relationship with rice quality and endeavored to improve the precision of selecting the appropriate sowing date in high-quality hybrid rice production. Deng et al. found that the average daily temperature ranges of 22–27 °C in the grain filling stage are recommended to achieve high grain yield and quality for irrigated rice in the Yangtze River basin (Deng et al. 2015). Our results suggest that lower average daily temperature can contribute to improving the quality of rice, supporting the conclusion from Deng et al. In addition, we found that the impact of meteorological factors on rice quality exhibited varied trends among trial locations, and even a single factor may affect multiple quality traits in contrasting directions, resulting in a complex effect on overall rice quality. For instance, surface solar radiation was positively correlated with AC and negatively correlated with GC at HN-LS, while contrary correlation trends were observed at the other four trial locations. This difference may be due to the fact that HN-LS is located in the South China rice cropping region, with persistent high temperatures and abundant rainfall in the dry season, and has distinct climate conditions compared to the other trial locations within the Yangtze River basin. Simultaneously, the decomposed meteorological factors are unavoidably related in different ways. For instance, rainfall consistently results in a more significant temperature decrease within AH-HF, HN-CS, and HB-EZ in the middle and lower reaches of the Yangtze River basin in comparison to SC-GH and HN-LS. This discrepancy can be attributed to the lower altitude and the prevalence of hot and sunny weather in the summer. Concurrently, rain can reduce solar radiation, so SC-GH in the upper reaches of the Yangtze River has low solar radiation due to cloudy and rainy weather. As the quality of rice is the result of the combined effects of multiple meteorological factors, further analysis and modeling based on more experiments are necessary.
It is crucial to develop varieties with stable phenotypes that are less sensitive to environmental changes. This will help mitigate the adverse effects of frequent extreme climate events on rice yield and quality, guaranteeing the production of high-quality rice. This study employs phenotypic plasticity measurement to evaluate the stability of different quality traits in hybrid rice combinations. Combining ability analysis is a useful approach to selecting ideal parent lines for hybrid rice breeding (Chen et al. 2019). Through the analysis of combining ability in phenotype plasticity, we discovered that the TGMS lines ZXS, HX302S, and HY468S and the restorer lines HH8012, HH7503, DZ, LKSM13, HH2646, HH5106, HH8549, YZ, and WSSM534 showed low GCA effects on five or more quality traits plasticity across three rice cropping regions; they could be recommended for utilization in rice hybrid breeding programs to improve the stability of rice quality. A noteworthy result was found that parents with superior quality tend to exhibit low GCA values for plasticity and have more potential to develop a hybrid with optimal quality stability. ZXS is an indica TGMS line with high-quality approved in Hunan province in 2020. Its milled rice rate is 63.2%, chalky grains rate is 7%, chalkiness degree is 1.3%, amylose content is 16.1%, gel consistency is 60 mm, alkali digestion value is 7.0, and transparency is grade 1 (China Rice Data Center). A total of ten new hybrid rice varieties of ZXS have been nationally or provincially approved and certificated, and their quality has all reached the grade 2 or grade 1. The quality of ZhenLiangYouYuZhan (ZXS/YZ), a nationally approved new hybrid rice variety of ZXS, met grade-1 in the new variety regional trial and was awarded the Gold Award for Taste Evaluation of High-Quality Rice at the 4th National High-Quality Rice Competition in 2023. In our experimental field settings, the ZXS/YZ combination showed high-quality with grade 3 or superior performance in 12 out of 19 records, and these records occurred in all locations except for HB-EZ.
To uncover the genetic basis of phenotype plasticity in rice quality traits, GWAS for the quality plasticity was performed and identified 13 plastic QTLs. In line with previous studies (Zan and Carlborg 2020), we determined that plasticity is polygenic and exhibits a variable genetic basis for rice quality traits across regions. The genetic effects of plastic QTL can change in different locations. For instance, the QTL6 is a multi-effect QTL associated with the plasticity of ASV and GC, where genetic effects on ASV differ among trial locations. We also identified seven plastic QTLs overlapped in the GWAS of BLUP measurement of quality traits, indicating these QTLs could regulate the quality traits and respond to the plasticity of quality traits. This further validates previous research that there may be a connection between the genetic regulation of traits and plasticity (Zan and Carlborg 2020; Jin et al. 2023).
In the present study, we endeavored to identify the candidate genes of two major plasticity QTLs for rice quality. The region of these two major QTLs contains 24 annotated genes and 2 of them (ALK and GL7) are known to regulate the rice quality. ALK is the key gene controlling rice gelatinization temperature, which is closely associated with the eating and cooking quality in rice (Gao et al. 2011). According to previous studies, there are three main alleles of ALK, including ALKa, ALKb, and ALKc, with ALKc controlling high gelatinization temperature, and ALKa and ALKb controlling low gelatinization temperature ( Chen et al. 2020; Huang et al. 2021). In this study, we identified two alleles of ALK including ALKb and ALKc in our parental rice accessions. Interestingly, we found that the leading SNP genotype of QTL6 was completely linked with ALK. Specifically, the CC genotype of the leading SNP of QTL6 was linked with ALKc while the TT genotype was linked with ALKb (Fig. 6g–h). Sequence analysis showed that among the above low plasticity GCA parents, all carried ALKb except HH7503 and HH5106, which carried ALKc. To some extent, this result explains although rice varieties with low to medium GT are preferred by consumers, those carrying ALKc alleles that regulate high GT are still commonly employed in rice breeding, potentially due to their plasticity levels.
The other plasticity candidate gene, the GL7 locus, plays a significant role in grain size diversity and has been utilized in rice breeding. Wang et al. demonstrated that the copy number variation (CNV) at the GL7 locus led to the difference in the rice grain size (Wang et al. 2015b). Whole genome sequencing analysis of parental rice accessions revealed that the CNV at GL7 was linked with the leading SNP of QTL7. The varieties containing the CNV at GL7 carry the AA genotype of the leading SNP of QTL7 and the remainder carry the GG genotype (Fig. 6i). Rice varieties with multi-copy at GL7 locus exhibit lower plasticity in terms of milled rice ratio, which can be attributed to their long-grain phenotype. Further studies will be necessary to determine the effect of these candidate genes on phenotypic plasticity and their pleiotropic effects on quality traits and plasticity. These QTLs and candidate genes have the potential to contribute to improving rice quality and maintaining the stability of quality in breeding.
References
Abd El-Aty MS, Katta YS, El-Abd AEMB et al (2022) The combining ability for grain yield and some related characteristics in rice (Oryza sativa L.) under normal and water stress conditions. Front Plant Sci 13:866742. https://doi.org/10.3389/fpls.2022.866742
Bates D, Mächler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48. https://doi.org/10.18637/jss.v067.i01
Bonamour S, Chevin L-M, Charmantier A, Teplitsky C (2019) Phenotypic plasticity in response to climate change: the importance of cue variation. Philos Trans R Soc B: Biol Sci 374:20180178. https://doi.org/10.1098/rstb.2018.0178
Bradshaw AD (1965) Evolutionary significance of phenotypic plasticity in plants. Adv Genet 13:115–155. https://doi.org/10.1016/S0065-2660(08)60048-6
Broad Institute (2023) Picard Tools - By Broad Institute. http://broadinstitute.github.io/picard/
Chen J, Zhou H, Xie W et al (2019) Genome-wide association analyses reveal the genetic basis of combining ability in rice. Plant Biotechnol J 17:2211–2222. https://doi.org/10.1111/pbi.13134
Chen Z, Lu Y, Feng L et al (2020) Genetic dissection and functional differentiation of ALKa and ALKb, two natural alleles of the ALK/SSIIa gene, responding to low gelatinization temperature in rice. Rice 13:39. https://doi.org/10.1186/s12284-020-00393-5
Cheng F, Zhong L (2001) Variation of rice quality traits under different climate conditions and its main affected factors. Chin J Rice Sci 15:187–191. https://doi.org/10.16819/j.1001-7216.2001.03.006. (in Chinese)
Cheng F, Zhu B (1998) Present research on the effect of meteoroecological factors on rice quality. Chin J Agrometeorol 19:39–45 (in Chinese)
Chevin L-M, Gallet R, Gomulkiewicz R et al (2013) Phenotypic plasticity in evolutionary rescue experiments. Philos Trans R Soc B: Biol Sci. https://doi.org/10.1098/rstb.2012.0089
China Rice Data Center (2020) ZhenXiangS == database of Chinese rice varieties and their genealogy. https://www.ricedata.cn/variety/varis/623567.htm. Accessed 27 Jul 2023
Cingolani P, Platts A, Wang LL et al (2012) A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly (austin) 6:80–92. https://doi.org/10.4161/fly.19695
Covarrubias-Pazaran G (2016) Genome-assisted prediction of quantitative traits using the R package sommer. PLoS One 11:e0156744. https://doi.org/10.1371/journal.pone.0156744
Deng N, Ling X, Sun Y et al (2015) Influence of temperature and solar radiation on grain yield and quality in irrigated rice system. Eur J Agron 64:37–46. https://doi.org/10.1016/j.eja.2014.12.008
Des Marais DL, Hernandez KM, Juenger TE (2013) Genotype-by-environment interaction and plasticity: exploring genomic responses of plants to the abiotic environment. Annu Rev Ecol Evol Syst 44:5–29. https://doi.org/10.1146/annurev-ecolsys-110512-135806
Finlay K, Wilkinson G (1963) The analysis of adaptation in a plant-breeding programme. Aust J Agric Res 14:742. https://doi.org/10.1071/AR9630742
Gage JL, Jarquin D, Romay C et al (2017) The effect of artificial selection on phenotypic plasticity in maize. Nat Commun 8:1348. https://doi.org/10.1038/s41467-017-01450-2
Gao Z, Zeng D, Cheng F et al (2011) ALK, The key gene for gelatinization temperature, is a modifier gene for gel consistency in rice (Oryza sativa L.). J Integr Plant Biol 53:756–765. https://doi.org/10.1111/j.1744-7909.2011.01065.x
Huang L, Tan H, Zhang C et al (2021) Starch biosynthesis in cereal endosperms: an updated review over the last decade. Plant Commun 2:100237. https://doi.org/10.1016/j.xplc.2021.100237
Jin M, Liu H, Liu X et al (2023) Complex genetic architecture underlying the plasticity of maize agronomic traits. Plant Commun 100473. https://doi.org/10.1016/j.xplc.2022.100473
Kadam NN, Tamilselvan A, Lawas LMF et al (2017) Genetic control of plasticity in root morphology and anatomy of rice in response to water deficit. Plant Physiol 174:2302–2315. https://doi.org/10.1104/pp.17.00500
Kikuchi S, Bheemanahalli R, Jagadish KSV et al (2017) Genome-wide association mapping for phenotypic plasticity in rice: genome-wide association mapping and rice plasticity. Plant Cell Environ 40:1565–1575. https://doi.org/10.1111/pce.12955
Kim SA, Brossard M, Roshandel D et al (2019) Gpart: human genome partitioning and visualization of high-density SNP data by identifying haplotype blocks. Bioinformatics 35:4419–4421. https://doi.org/10.1093/bioinformatics/btz308
Li H (2013) Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 1303:3997v1
Li L, Zheng X, Wang J et al (2023) Joint analysis of phenotype-effect-generation identifies loci associated with grain quality traits in rice hybrids. Nat Commun 14:3930. https://doi.org/10.1038/s41467-023-39534-x
Lian L, De Los Campos G (2016) FW: An R Package for Finlay-Wilkinson regression that incorporates genomic/pedigree information and covariance structures between environments. G3 (Bethesda) 6:589–597. https://doi.org/10.1534/g3.115.026328
Liu Q, Wu X, Ma J et al (2013) Effects of high air temperature on rice grain quality and yield under field condition. Agron J 105:446–454. https://doi.org/10.2134/agronj2012.0164
Lu X, Wang L, Xiao Y et al (2022) Grain quality characterization of hybrid rice restorer lines with resilience to suboptimal temperatures during filling stage. Foods 11:3513. https://doi.org/10.3390/foods11213513
McKenna A, Hanna M, Banks E et al (2010) The genome analysis toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res 20:1297–1303. https://doi.org/10.1101/gr.107524.110
Mu Q, Guo T, Li X, Yu J (2022) Phenotypic plasticity in plant height shaped by interaction between genetic loci and diurnal temperature range. New Phytol 233:1768–1779. https://doi.org/10.1111/nph.17904
Patterson N, Price AL, Reich D (2006) Population structure and eigenanalysis. PLoS Genet 2:e190. https://doi.org/10.1371/journal.pgen.0020190
Resurreccion AP, Hara T, Juliano BO, Yoshida S (1977) Effect of temperature during ripening on grain quality of rice. Soil Sci Plant Nutr 23:109–112. https://doi.org/10.1080/00380768.1977.10433027
Sprague GF, Tatum LA (1942) General vs. specific combining ability in single crosses of Corn1. Agron J 34:923–932. https://doi.org/10.2134/agronj1942.00021962003400100008x
Sultan SE (2000) Phenotypic plasticity for plant development, function and life history. Trends Plant Sci 5:537–542. https://doi.org/10.1016/S1360-1385(00)01797-0
Tian Z, Qian Q, Liu Q et al (2009) Allelic diversities in rice starch biosynthesis lead to a diverse array of rice eating and cooking qualities. Proc Natl Acad Sci USA 106:21760–21765. https://doi.org/10.1073/pnas.0912396106
Wang Y, Mette MF, Miedaner T et al (2015) First insights into the genotype–phenotype map of phenotypic stability in rye. J Exp Bot 66:3275–3284. https://doi.org/10.1093/jxb/erv145
Wang Y, Xiong G, Hu J et al (2015) Copy number variation at the GL7 locus contributes to grain size diversity in rice. Nat Genet 47:944–948. https://doi.org/10.1038/ng.3346
Wenting W, Wenpei C, Ke X et al (2021) Effects of early- and late-sowing on starch accumulation and associated enzyme activities during grain filling stage in rice. Rice Sci 28:191–199. https://doi.org/10.1016/j.rsci.2021.01.008
Wu Y-C, Chang S-J, Lur H-S (2016) Effects of field high temperature on grain yield and quality of a subtropical type japonica rice-Pon-Lai rice. Plant Prod Sci 19:145–153. https://doi.org/10.1080/1343943X.2015.1128091
Yan H, Wang S, Wang X et al (2021) Performance of different rice varieties under high temperature and its relationship with field meteorological factors. Chin J Rice Sci 35:617–628. https://doi.org/10.16819/j.1001-7216.2021.210509. (in Chinese)
Yao Y, Huo Z, Zhang H et al (2011) Effects of sowing date on yield and quality of direct seeding rice of different types and varieties. Sci Agric Sin 44:3098–3107 (in Chinese)
Zan Y, Carlborg Ö (2020) Dissecting the genetic regulation of yeast growth plasticity in response to environmental changes. Genes 11:1279. https://doi.org/10.3390/genes11111279
Zhang C, Zhao D, Li Q et al (2016) Progresses in research on cloning and functional analysis of key genes involving in rice grain quality. Sci Agric Sin 49:4267–4283. https://doi.org/10.3864/j.issn.0578-1752.2016.22.002. (in Chinese)
Zhou X, Stephens M (2014) Efficient multivariate linear mixed model algorithms for genome-wide association studies. Nat Methods 11:407–409. https://doi.org/10.1038/nmeth.2848
Funding
This work was supported by grants from National Natural Science Foundation of China (U19A2032), Science and Technology Innovation Program of Hunan (2021NK1001, 2022NK1010, 2023NK1010), and Shenzhen Science and Technology Project (202320D314).
Author information
Authors and Affiliations
Contributions
YZY, XML, and KW conceived and designed experiments. RRC, DXL, and JF contributed to data analysis and the first draft of the manuscript. RRC and YJW contributed to data collection. CJF, PQ, XWZ, ZBS, KH, LL, and WZ were in charge of filed management. YZY, XML, and KW supervised and complemented the manuscript writing. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chen, R., Li, D., Fu, J. et al. Exploration of quality variation and stability of hybrid rice under multi-environments. Mol Breeding 44, 4 (2024). https://doi.org/10.1007/s11032-024-01442-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11032-024-01442-3