
Abstract
Environmental conditions for trees that established in central Fennoscandia shortly after the final retreat of the Weichselian ice sheet remain poorly understood. In this study we examine tree rings of five well-preserved Pinus sylvestris (Scots pines) that grew in the area in front of the retreating ice sheet in northern Sweden. They became buried in flood sediments deposited by a glacial outburst flood (jökulhlaup) about 9.5–9.9 kyr cal bp and the aim of our study was to search for information regarding damage from fires and bioclimatic conditions in their ancient tree ring records. Our analysis, providing a glimpse into the local early Holocene environment in north-central Sweden, suggests that: 1, there were repeated fires (four fire events detected) during the early Holocene; and 2, bioclimatic conditions when the ancient pines were growing resembled those of modern sub-alpine pine woods. The latter is indicated by their patterns of tree ring growth (growth rate and variation), which were statistically similar to those of pines growing in sub-alpine woods with an open canopy, but different from pines in protected and managed boreal forests. Lower δ13C for the ancient latewood in comparison to pine wood from trees growing near the Scandinavian mountains before the 1850s were probably caused both by stomata fractionation due to lower atmospheric CO2 during the early Holocene and by the moist local environment created by the nearby ancient Ancylus lake, which preceded the Baltic Sea. Periods with cloudy and cold summers were also indicated by the occurrence of ‘false rings’. Finds of charred fragments of Calluna vulgaris (heather, ling), an understory shrub that can burn even with a relatively high moisture content, suggest that heath vegetation was crucial to make fire a reoccurring ecological factor in the area during the early Holocene.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Our understanding about environmental conditions prevailing in terrestrial ecosystems directly after the retreat of the Weichselian ice sheet in northern Fennoscandia is limited. The primary reason for this knowledge gap is that natural archives that both span the early Holocene period and offer information about terrestrial ecosystems with an annual resolution, such as tree rings, have been lacking from north-central Sweden where the last remnants of the ice sheet lingered as late as ca. 9.2–9.5 kyr cal bp (Hughes et al. 2016; Stroeven et al. 2016). The late deglaciation of north-central Sweden makes it easy to assume that the region was covered by a continuous blanket of ice until at least 10 kyr cal bp and that terrestrial sources of proxy data such as pollen could be used for environmental reconstruction from ca. 9 kyr cal bp (Seppä et al. 2009; Mauri et al. 2015). For tree rings, the oldest dendrochronological time series (about 7.4 kyr cal bp) from northern Fennoscandia comes from the northernmost part of the area that was deglaciated several millennia earlier than the central part (Grudd et al. 2002). Indeed, the absence of early Holocene tree rings from the north-central part of the area covered by the former ice sheet seems unsurprising, given its late deglaciation. However, fragments of Picea abies (Norway spruce) and Pinus sylvestris (Scots pine) dating back to the late Weichselian glaciation have been found on presumed nunataks which protruded through the ice sheet in the Scandinavian mountains (Kullman 2008). Unfortunately, the wood from these samples was too degraded for detailed dendrochronological analysis of tree growth. Other indications, suggesting that the arrival of trees directly followed the retreating ice margin in the central part of Fennoscandia during the early Holocene, come from studies detecting spruce DNA fragments in 10 kyr old sediments (Nota et al. 2022). It has even been suggested from pollen records that tree taxa, such as Larix (larch) and Betula (birch), had colonized debris on top of the stagnant Weichselian ice sheet around the time of deglaciation (Zale et al. 2018).
The lack of well-preserved tree ring records from the early Holocene in Fennoscandia has led researchers to attempt the extraction of growth rates from charcoal located within archaeological deposits, but problems with shrinking during heating as well as unknown sample origins, from trunks or branches, made it difficult to compare these tree ring analyses with those of modern specimens (Carcaillet 2017). Findings from well-preserved tree rings from the first pioneer trees that established in front of the decaying ice sheet in northern Scandinavia could provide detailed information about bioclimatic conditions and possible fire regimes in this region. Here, the existence of fires seems indeed uncertain at this presumed early stage of vegetational succession, as insufficient amounts of burnable materials may have limited them. It takes decades for a burnable forest floor layer to form in mature boreal forests (Schimmel 1993; Schimmel and Granström 1997). While most studies seem to agree that a pine-birch woodland dominated the area in the centuries after the ice sheet retreated from central Fennoscandia (Bergman et al. 2005), it is less certain when fire became an ecological factor in this early environment. Our limited knowledge of fire frequency and intensity in the early post-glacial landscape hampers our understanding of both early ecosystem dynamics and climate-driven fire regimes. Forest fires are a significant ecological factor that regulates the ecological dynamics as well as light and nutrient availability in boreal forests (Schimmel 1993; Schimmel and Granström 1997; Niklasson and Granström 2000; Thonicke et al. 2001; Wardle et al. 2003; Zackrisson et al. 2004). Therefore, mimicking natural fire frequency and intensity is a common aim of forestry management programs (Angelstam 1998).
Our knowledge of fire regimes in the post-glacial landscape is mainly based on the analysis of charcoal fragments from lake sediments or peat. Charcoal found in dated lake sediments from southern Sweden suggests that local fire events started several millennia after deglaciation (Hannon et al. 2018). In contrast, similar analyses in northern Sweden suggest that fires began when the degrading ice sheet was still present in the region (Carcaillet et al. 2007, 2012). However, while the size of charcoal pieces in sedimentary or peat archives can be used to differentiate between local fires (large fragments) and regional fires (small fragments), tree ring analysis is the best technique to date local fire frequency with a high resolution (Lageard et al. 2000).
The overall purpose of this study was to assess the information on fire frequencies and bioclimatic conditions preserved in the tree ring record of some uniquely well-preserved Pinus sylvestris trees which were buried in a massive flood deposit in northern Fennoscandia. Their age (ca. 9.6 kyr cal bp) suggests that they were growing near the last remnants of the degrading Weichselian ice sheet. We aimed to retrieve; 1, possible evidence of early fire regimes; and 2, information on bioclimatic conditions for tree growth during the early formation of the woodland ecosystem by analysing the tree ring record. The latter was assessed using measurement of tree ring width and δ13C analysis of the latewood as a proxy for drought stress during the summer season.
Materials and methods
Site description
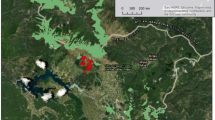
The study site (64°36”N, 18°42”E) is situated in north-central Sweden within the municipality of Lycksele (Fig. 1a). The best estimates from deglaciation models (Hughes et al. 2016; Stroeven et al. 2016) suggest that this region was deglaciated ca. 9.9–10 kyr cal bp (Fig. 1b, c). However, Hughes et al. (2016) considered uncertainties in their deglaciation data and suggested that the study area may have been under an ice sheet many kilometres from its edge as late as 10 kyr cal bp. Some models also propose deglaciation of the study site as late as 9.5 kyr cal bp (Parducci et al. 2012). The excavated pit was situated within a glaciofluvial or fluvial floodplain (Fig. 2a). The site is at ~ 218 m a.s.l. and several hundred metres from the present river course of the Umeälven (mean water level at 211 m).

Location of the study site (red star on all maps); a, within Fennoscandia; b, in relation to the retreating Weichselian ice sheet in the study region around 10 kyr cal bp according to Hughes et al. (2016), with the relative uncertainty indicated by three possible situations: maximum extent (dashed black line), minimum extent (dotted black line) and most probable extent (white line); c, in relation to the modelled deglaciation margins (yellow lines) between 10.4 and 9.7 kyr cal bp according to Stroeven et al. (2016). The maps in panels b and c are modified from Hughes et al. (2016) and Stroeven et al. (2016), respectively. YD, Younger Dryas

a, The excavated site with the fallen pine tree (T1), wrapped in plastic before its removal to Trädmuseet (Forest Museum) in Lycksele, visible in the centre (red arrow); b, Close up picture of the remaining branches of T1. Note that the tree bark was intact (but fell off upon drying) and there were no signs of deadwood beetles, which indicates that the trees were rapidly buried and sealed off from the soil surface; c, the disrupted roots of T1 still with embedded stones and inclusions of a silt-rich till, suggesting short distance transport of the fallen tree by water; d, the excavation, showing the positioning of a partly standing pine tree (T3, red arrow) situated near the fallen pine (T1); e, overview of the excavation sites where the positions and the tilting of the three pine trees found in the pit are shown with red arrows; f, two fire scars (injuries) at the base of T1 (two red arrows) from two separate points in time
In 1986, two Pinus sylvestris tree trunks ca. ~1.8 m long and with roots were excavated in a construction project ca. 100 m from the excavation site (64°36’07,87”N, 18°42’20,57”E) and were taken to Skogsmuseet (the Forestry Museum) in Lycksele and preserved there. In 2005, a ground penetrating radar (GPR) survey by Malå Geoscience/RAYCON identified additional objects (potentially trees) buried at 2–4 m depth at two nearby sites. We selected one of the most promising sites where our excavation revealed the presence of a fallen 12 m long P. sylvestris tree (Fig. 2a), with branches and bark still attached to the trunk (Fig. 2b). The tree (T1) was lying between depths of about 2.7 m (top) down to about 3.4 m (roots). We found large lenses of stones and silt within the roots of pine T1 and we assume that this glacial till was the original growth substrate. During the excavation, two additional pine trunks (T2 and T3), not previously detected by the ground penetrating radar, were found protruding from the sediment deposit (Fig. 2d, e). The deep burial (> 4 m) of these trunks made it impossible to retrieve the roots. The original fallen tree (T1) also had two evident fire scars (Fig. 2f). The setting and the vertical stratigraphy of the excavated 10 m deep deposit in which the trees were found is shown in ESM Fig. S1.
To study the formation of the deposit as well as the material in which the trees had grown, we used a Hypro GH8 percussion corer. Cores from a 10 m deep sediment sequence were scrutinized for the main sedimentary layers (ESM) and plant macrofossils. Samples of plant material from the sediment core which were used for dating are referred to as “macrofossil samples” in the following text, and dated wood samples from the buried trunks or branches as “megafossil samples”.
Radiocarbon dating
Radiocarbon dating was done by Beta Analytic Inc., Miami, Florida, using standard Accelerator Mass Spectrometry (AMS). All samples were dated using individual macrofossils or wood from individual megafossils to avoid dating samples with remains of different ages, which is known to be a problem in this area (Zale et al. 2018). The AMS 14C dates were calibrated using BetaCal v. 3.21 (Table 1).
Dendrochronological analyses and comparison with modern tree rings
Tree ring analyses were done on the five P. sylvestris trees (N = 5), the three recently excavated tree trunks (T1-3), as well as the two found in 1986 that are preserved at Skogsmuseet (the Forestry Museum) in Lycksele (T4 and T5). Seven cores for tree ring analysis were extracted using a 12 mm diameter increment borer mounted on a power drill, and in three cases a disc sample or ‘tree cookie’ was also taken. The samples were then prepared according to standard dendrochronological techniques (Baudet et al. 2020).
We compared our early Holocene tree ring dataset to three modern pine chronologies from sites in northern Sweden: one from a boreal forest in Lycksele and two from a sub-alpine wood in Tjeggelvas nature reserve. The Lycksele chronology covers 470 years (1531–2000) and the Tjeggelvas ones 332 years (1690–2021) for the site located in the forest zone at 475 m and 218 years (1803–2021) for the site located at the P. sylvestris treeline at 625 m. We also compared our dataset to tree ring data available from the Riksskogstaxeringen (Swedish National Forest Inventory) for managed forests in the Lycksele area covering 67 years (1946–2013).
Latewood δ13C values are a common proxy for drought stress due to the reduced photosynthetic discrimination against 13C in favour of 12C during drought periods (Eilmann et al. 2010). In P. sylvestris from north-central Sweden, increased δ13C in pine wood correlates with warm and dry summers with high atmospheric pressure (Seftigen et al. 2011). We sampled latewood for δ13C analysis from the longest individual time series (T1 and T2) with a Proxxon TBM 220 with a 0.1–0.3 mm drill bit and connected to a digital microscope with a 50× magnification. Sampled latewood was analysed for δ13C using an Isotope Ratio Mass Spectrometer, EA-IRMS.
Statistics
We compared growth rates and growth variability for the ancient wood samples with similar data from the more recent Tjeggelvas and Riksskogstaxeringen inventory datasets to search for modern analogues that matched our buried pine trees. We compared average growth rates (mm yr− 1) per wood sample and also the coefficient of variation (CV):
Here, Stdev ring width refers to the standard deviation (mm) of the whole tree ring sequence and Mean ring width is the average ring increment (mm) for the same sequence; hence, the coefficient of variation is a growth rate normalized for the variability in ring width. In addition, we tested for differences in the δ13C values from the latewood of the buried pines and from trees growing in central Sweden, previously analysed by Seftigen et al. (2011). Differences between our buried tree trunks and modern pines were assessed using a one-way analysis of variance (ANOVA). Growth rates were log-transformed and residual ANOVA plots were visually inspected and evaluated to fulfil the assumption of normality. A Tukey’s range test was used to assess the significance of differences between groups. The following groups were tested: 1, pines growing in the valley at 475 m below the treeline in Tjeggelvas; 2, pines growing in sub-alpine birch woods near the treeline at 675 m in Tjeggelvas; 3, pines from a protected boreal forest in the study region (County of Västerbotten); 4, pines from managed boreal forests in the study region. All tests were done using IBM SPSS statistics v.28.
Results
In our sediment core from the flood deposit, the first occurrence of macrofossils (Calluna vulgaris, Salix spp., Betula spp. and Pinus sylvestris) was found at a depth of ~ 4.2 m and with an age ranging between 9.8 and 9.5 kyr cal bp (Table 1). Here, Calluna vulgaris (heather) was partly burned (charred). Many macrofossils were found in the deposit without any apparent chronological order. Grouped together with the wood samples from the museum (T4 and T5), the best age estimates (mean 14C age ± std. error) for the megafossils (N = 6) and macrofossils (N = 8) are 9.5–9.7 and 9.6–9.8 kyr cal bp, respectively (Table 1).
Environmental conditions inferred from the tree rings
The tree rings from the buried pines were very well preserved and indicated a highly variable growth history (Fig. 3a-c). Many rings displayed a latewood with irregularities in colour and length and a ‘false ring’ suggesting short periods with a rapid reduction in growth, followed by a return to normal growth. Compression wood was common, but we detected no apparent disturbance from summer frost in the rings. However, besides occurrence of false rings from short periods of reduced growth, we noted the frequent presence of scattered narrow rings from years with slow growth, interspersed among wide rings from fast growth. Two of the trunks (T1 and T5) had two fire scars each, with 19 (T1) and 20–23 (T5) years between the fires (return intervals), respectively. Judging by the encroachment wood grown over the damage and the surface of injury, the two fires separated by the 19 year gap had a rather moderate intensity, whereas the latest burning of T5 might have been of high intensity. The trees were 12 (T1) and 23 (T5) years old when the first fire occurred.

a, series of tree rings from the ancient Pinus sylvestris tree (T5) excavated from the Lycksele site; b, a series of tree rings from a P. sylvestris growing in Tjeggelvas nature reserve at 625 m.; c, a series of tree rings from a P. sylvestris growing in Tjeggelvas nature reserve at 475 m. Note that the ancient tree in a shows a later period of increased growth compared to those currently growing at Tjeggelvas
We compared growth rates and variation in growth (CV) of pines from the early Holocene with present day examples from woods and forests (protected sub-alpine birch woods, protected boreal forest and managed boreal forests). We found differences between these trees in terms of log growth rates (F4, 437=24.23; P < 0.001) and CV (F4, 437=25.84; P < 0.001). The post-hoc statistical test suggested that the buried pine trees were comparable to those growing in recent sub-alpine birch woods, but different from ones from modern boreal forests (Fig. 4). The growth rates of the buried trees were lower, but showed higher variation in growth between years than trees in protected and managed boreal forests. The δ13C of the latewood of the buried trees was lower (F2, 199=131.80; P < 0.001) than that of pine trees grown during ad 1730–1850 in central Sweden (Fig. 5). The differences between the two age classes remained when correcting the δ13C of the buried trees for the fractionation effect caused by the lower CO2 concentration in the atmosphere during the early Holocene (F2, 199=51.18; P < 0.001).

Boxplots of tree ring growth rates (above) and variance in growth (below) for pines growing in four different modern environments, protected sub-alpine birch woodland, protected sub-alpine treeline, protected boreal forest and managed boreal forest, and early Holocene pine trees. The boxes show the 25 and 75% quartiles and whiskers represent the 1.5QR. Outliers are shown as black filled diamond shapes. Similar letters above the boxplots indicate that data are not statistically different (p > 0.05) according to a Tukey post hoc test, and different letters represent significant differences between boxes. Note that the statistics provided for growth rates are based on log-transformed data

Boxplot of the δ13C values for pine wood grown in central Sweden between 1730 and 1850 (pre-Suess effect) as determined by Seftigen et al. (2011), compared to values for the early Holocene pine trees (T1 and T2, N = 131). Values for the ancient pine tree rings which have been corrected for the increased stomata fractionation due to lower CO2 concentration in the atmosphere during the early Holocene are also shown, on right panel. The latter correction is based on the isotope fractionation observed for Pinus contorta as a function of naturally elevated CO2 (Sharma and Williams 2009). Note that the latter study represents the most extensive, whole-ecosystem isotope fractionation experiment on natural elevated CO2 and Pinus sp. Box limits show the 25 and 75% quartiles and whiskers represent the 1.5QR. Different letters above the boxplots indicate statistically significant differences (p > 0.05) according to a Tukey post hoc test. Background colours show the general correlation between δ13C values and either cold/wet (blue) or warm/dry (pink) growth conditions
Discussion
Fire regimes in the pioneer post-glacial woodland
From the four fire scars found on the analysed pine trunks, it is evident that fire was an active factor in the landscape at a time where remnants of the degrading ice sheet was still present in north-central Sweden. The detection of past fires in these Scots pine tree rings from 9.5 to 9.7 kyr cal bp is, to our knowledge, the oldest direct evidence of such early fires in this region. Fire regimes during the early Holocene in northern Sweden have so far been inferred from indirect lines of proxy evidence from lake sediments, where charcoal size has been used to distinguish between local and regional fires (Carcaillet et al. 2007), so our findings are important for settling any uncertainties relating to early local fires.
Fire return intervals of 19–23 years for T1 and T5 are intriguingly short. Detailed studies using tree ring chronologies from boreal landscapes have suggested a typical fire return interval of 100–300 years before the start of modern forestry (Zackrisson 1977; Morgan et al. 2001). Studies using sedimentary archives to discover fire frequency have suggested similar fire return intervals during the early Holocene in southern Sweden, ranging between ca. 100 years (Olsson et al. 2010) and ca. 300 years (Carcaillet et al. 2007). Nevertheless, a shorter mean fire return interval of 10–90 years, where variations are driven by site-specific conditions, has been observed in some unmanaged boreal pine forest stands (Zackrisson 1977; Engelmark 1984; Lageard et al. 2000; Ivanova et al. 2010; Ryzhkova et al. 2022). With these lower estimates in mind, our inferred fire frequency during the early Holocene may also have direct analogues in modern boreal forests. We acknowledge that our limited number of samples makes it difficult to estimate how common such short fire return intervals were at a larger regional scale in the early Holocene environment. However, a return interval of few decades is mostly impossible to detect using the time resolution usually provided by lake sediment chronologies, so that charcoal from several fires with short return rates might seem to correspond to a single event.
Fire frequency and Holocene vegetation in Fennoscandia are inextricably interlinked (Cui et al. 2013; Molinari et al. 2020), and it seems possible that the dominance of fire resistant trees such as P. sylvestris during the early Holocene was at least partly caused by a high frequency of low intensity fires, for it is well-established knowledge that fires give pine a competitive advantage over other trees (Rogers et al. 2015). Therefore, it seems plausible that frequent fires gave pine a competitive advantage over Picea abies (spruce) during the early Holocene, which may have played a role in the unexpectedly long time lag between the local establishment of spruce in Fennoscandia more than 10,000 years ago and its main migration wave which occurred many millennia later (Nota et al. 2022). Moreover, forest fires vigorously stimulate nitrogen cycling in boreal forests (Neary et al. 1999; Zackrisson et al. 2004), and if fires occurred every few decades, it seems likely that they were crucial for maintaining productivity in the early Holocene post-glacial landscape. This possible role of fire is important to highlight in this context, given that the trees in the study area are believed to have become established before the arrival of nitrogen fixing taxa such as Alnus (alder), which became established several thousand years later (Bergman et al. 2005; Kullman and Öberg 2015).
The occurrence of macrofossils of mosses, Betula and Pinus, which were detected in the flood sediments burying the pine trunks, is consistent with previous suggestions that a Betula-Pinus woodland with an open canopy established itself early at the south-western edge of the ice sheet in central Sweden (Bergman et al. 2005; Berglund 2008), and that this type of woodland became established a millennium later in northern Fennoscandia (Sjögren and Damm 2019). Pines growing in sub-alpine birch woods with an open canopy also seem like the best analogue for the early Holocene pine tree rings. Statistically, the growth pattern in the tree ring record of these pines is comparable to that of pines growing in protected sub-alpine birch woods in the Scandinavian mountains (Scandes). That is, the buried trunks showed a higher variation in ring growth than pine from modern boreal forest, suggesting a more variable growth environment in the early Holocene. Furthermore, the δ13C of the latewood of the ancient tree rings was clearly lower than the values of more modern analogues from central Sweden, where lower δ13C values are typically interpreted to represent lower summer temperatures or moister conditions (Hilasvuori et al. 2009; Seftigen et al. 2011; Esper et al. 2018). However, the early Holocene atmosphere contained only about 265 ppm CO2, while in the 1850s it was about 288 ppm (Flückiger et al. 2002), and when the ancient trees were growing, the lower carbon dioxide concentration is expected to have caused increased stomata fractionation (O’Leary 1988). Established regressions between atmospheric CO2 concentrations and fractionation for Pinus suggest that the ~ 22 ppm lower CO2 concentration during the early Holocene probably caused a 0.4‰ lower δ13C in the pine wood (Sharma and Williams 2009). Despite correcting for this fractionation, the ancient pine trees still have δ13C values ~ 0.6‰ lower than pine wood from the early 19th century. We note that pine wood growing near shorelines in northern Sweden has a -0.7‰ lower δ13C than from inland at comparable latitude (Esper et al. 2018). Therefore, we interpret the lower δ13C for the early Holocene pines to be a result of their growing environment near the lake shore, as the excavated site was part of the Ancylus lake shoreline following deglaciation (Björck 1995). This interpretation is in line with the occurrence of false rings in the tree ring record. Their presence suggests that cold and wet summers periodically suppressed growth, as such rings are usually interpreted as indications of periods with these conditions which are typical of more maritime climates (Nöjd and Hari 2001). Lower CO2 in the atmosphere in combination with growing near the shore of the Ancylus lake are the most likely causes of the low δ13C values in the early Holocene tree rings. However, we cannot exclude the possibility that lower δ13C in the ancient trees was to some extent caused by colder summers compared to the early 19th century. The observation of scarce narrow rings interspersed within a series of wider rings in our tree ring records indicate substantial drops in air temperatures during some summers (Cook and Kairiukstis 2013). Our interpretation of periodically colder summer temperatures is also supported by pollen records indicating 0–2 °C cooler summers in our study area around 9–10 kyr cal bp in comparison to the 21st century (Mauri et al. 2015). However, we emphasize that the inferred cooler climate may have been of very local character, as multiproxy reconstructions, based on pollen, chironomids and diatoms have suggested warmer summers in both the northern and southern parts of the Scandinavian mountains around this time (Bigler et al. 2002; Paus and Haugland 2017). Nevertheless, it seems possible that remains of the stagnant ice sheet had a cooling impact on the local climate, as deglaciation maps of Fennoscandia predict that remnants of the ice sheet may have persisted in central Sweden until ca. 9.6 kyr cal bp (Hughes et al. 2016; Stroeven et al. 2016). Indeed, outburst floods from collapsing ice-walled lakes together with meltwater from higher up in the Umeälven catchment are the most likely causes of the burial of the excavated trees. Floods of this type in the early Holocene have previously been described for river valleys in northern Sweden (Regnéll et al. 2019).
During the early Holocene, fire regimes are assumed to have been climatically controlled, as the inferred warm and dry climate would have favoured them (Olsson et al. 2010). However, in our study area the frequent fires in the early Holocene occurred in an environment best described as a sub-alpine woodland with a moist climate influenced by the nearby Ancylus lake. A previous study has shown that forest fires in Fennoscandia are associated with conditions prevailing in the northern North Atlantic and that they can become more frequent during cold periods with unstable conditions (Drobyshev et al. 2016). Our finding is also in agreement with other fire regime reconstructions suggesting that high frequencies of small fires characterize cool and wet periods in Fennoscandia (Remy et al. 2023). Calluna shrubs may have provided living material that burns even with a relatively high moisture content in maritime climates (Davies et al. 2009). Charred Calluna macrofossils in our studied deposit indicate that heather played an important role as combustible material at the site.
Conclusions: of ice and fire
Our findings represent a snapshot of a local early Holocene environment. It should only be generalised to the whole region with caution, given the limited number of samples and the local character of the data. However, our results indicate that although remnants of the Weichselian ice sheet were still present in north-central Sweden ca. 9.5–9.9 kyr cal bp, fire was already acting as a major ecological factor despite the presumed early succession stage of the post-glacial landscape. At this early time, patterns of tree ring growth were comparable with those found in pines growing within protected sub-alpine birch woods today. Despite a local climate influenced by the adjacent Ancylus lake, this early Holocene forest had an intriguing short fire return interval of less than 25 years. The short intervals indicate that burnable materials were abundant just a few centuries after deglaciation and that fires played an important role when the first Pinus and Betula woods became established in northern Fennoscandia.
Data Availability
Tree ring data in support of the findings of this study are available upon request from Lars Östlund: Lars.Ostlund@slu.se.
References
Angelstam PK (1998) Maintaining and restoring biodiversity in European boreal forests by developing natural disturbance regimes. J Veg Sci 9:593–602
Baudet M, Josefsson T, Östlund L (2020) New insights into estimating the age of old Scots pine from increment cores with stem rot. Dendrochronologia 64:125782
Berglund M (2008) Time-transgressive early Holocene vegetational succession following shore displacement: a case study from central Sweden. Boreas 37:87–101
Bergman J, Hammarlund D, Hannon G, Barnekow L, Wohlfarth B (2005) Deglacial vegetation succession and Holocene tree-limit dynamics in the Scandes Mountains, west-central Sweden: stratigraphic data compared to megafossil evidence. Rev Palaeobot Palynol 134:129–151
Bigler C, Larocque I, Peglar SM, Birks HJB, Hall RI (2002) Quantitative multiproxy assessment of long-term patterns of Holocene environmental change from a small lake near Abisko, northern Sweden. Holocene 12:481–496
Björck S (1995) A review of the history of the Baltic Sea, 13.0–8.0 ka bp. Quat Int 27:19–40
Carcaillet C (2017) Unlimited fuel wood during the middle Mesolithic (9650–8300 cal. Yr bp) in northern Sweden: fuel typology and pine-dominated vegetation inferred from charcoal identification and tree-ring morphology. Holocene 27(1):370–1378
Carcaillet C, Bergman I, Delorme S, Hornberg G, Zackrisson O (2007) Long-term Fire frequency not linked to prehistoric occupations in northern Swedish boreal forest. Ecology 88:465–477
Carcaillet C, Hörnberg G, Zackrisson O (2012) Woody vegetation, fuel and Fire track the melting of the scandinavian ice-sheet before 9500calyrBP. Quat Res 78:540–548
Cook ER, Kairiukstis LA (eds) (2013) Methods of dendrochronology: applications in the environmental sciences (ebook edition). Springer, Dordrecht
Cui Q-Y, Gaillard M-J, Lemdahl G, Sugita S, Greisman A, Jacobson GL, Olsson F (2013) The role of tree composition in Holocene Fire history of the hemiboreal and southern boreal zones of southern Sweden, as revealed by the application of the Landscape Reconstruction Algorithm: implications for biodiversity and climate-change issues. Holocene 23:1747–1763
Davies GM, Legg CJ, Smith AA, MacDonald AJ (2009) Rate of spread of Fires in Calluna vulgaris-dominated moorlands. J Appl Ecol 46:1054–1063
Drobyshev I, Bergeron Y, de Vernal A, Moberg A, Ali AA, Niklasson M (2016) Atlantic SSTs control regime shifts in forest Fire activity of Northern Scandinavia. Sci Rep 6:22532
Eilmann B, Buchmann N, Siegwolf R, Saurer M, Cherubini P, Rigling A (2010) Fast response of Scots pine to improved water availability reflected in tree-ring width and δ13C. Plant Cell Environ 33:1351–1360
Engelmark O (1984) Forest Fires in the Muddus National Park (northern Sweden) during the past 600 years. Can J Bot 62:893–898
Esper J, Holzkämper S, Büntgen U et al (2018) Site-specific climatic signals in stable isotope records from Swedish pine forests. Trees 32:855–869
Flückiger J, Monnin E, Stauffer B et al (2002) High-resolution Holocene N2O ice core record and its relationship with CH4 and CO2. Glob Biogeochem Cycles 16:10–11
Grudd H, Briffa KR, Karlén W, Bartholin TS, Jones PD, Kromer B (2002) A 7400-year tree-ring chronology in northern Swedish Lapland: natural climatic variability expressed on annual to millennial timescales. Holocene 12:657–665
Hannon GE, Halsall K, Molinari C, Boyle J, Bradshaw RHW (2018) The reconstruction of past forest dynamics over the last 13,500 years in SW Sweden. Holocene 28:1791–1800
Hilasvuori E, Berninger F, Sonninen E, Tuomenvirta H, Jungner H (2009) Stability of climate signal in carbon and oxygen isotope records and ring width from Scots pine (Pinus sylvestris L.) in Finland. J Quat Sci 24:469–480
Hughes ALC, Gyllencreutz R, Lohne ØS, Mangerud J, Svendsen JI (2016) The last eurasian ice sheets – a chronological database and time-slice reconstruction, DATED-1. Boreas 45:1–45
Ivanova GA, Ivanov VA, Kukavskaya EA, Soja AJ (2010) The frequency of forest Fires in Scots pine stands of Tuva, Russia. Environ Res Lett 5:015002
Kullman L (2008) Early postglacial appearance of tree species in northern Scandinavia: review and perspective. Quat Sci Rev 27:2467–2472
Kullman L, Öberg L (2015) New aspects of High-Mountain Palaeobiogeography: a synthesis of data from Forefields of receding glaciers and ice patches in the Tärna and Kebnekaise Mountains, Swedish Lapland. Arctic 68:141–152
Lageard JGA, Thomas PA, Chambers FM (2000) Using Fire scars and growth release in subfossil Scots pine to reconstruct prehistoric Fires. Palaeogeogr Palaeoclimatol Palaeoecol 164:87–99
Mauri A, Davis BAS, Collins PM, Kaplan JO (2015) The climate of Europe during the Holocene: a gridded pollen-based reconstruction and its multi-proxy evaluation. Quat Sci Rev 112:109–127
Molinari C, Carcaillet C, Bradshaw RHW, Hannon GE, Lehsten V (2020) Fire-vegetation interactions during the last 11,000 years in boreal and cold temperate forests of Fennoscandia. Quat Sci Rev 241:106408
Morgan P, Hardy CC, Swetnam TW, Rollins MG, Long DG (2001) Mapping Fire regimes across time and space: understanding coarse and fine-scale Fire patterns. Int J Wildland Fire 10:329–342
Neary DG, Klopatek CC, DeBano LF, Ffolliott PF (1999) Fire effects on belowground sustainability: a review and synthesis. For Ecol Manage 122:51–71
Niklasson M, Granström A (2000) Numbers and sizes of Fires: long-term spatially explicit Fire history in a Swedish boreal landscape. Ecology 81:1484–1499
Nöjd P, Hari P (2001) The effect of temperature on the radial growth of Scots pine in northernmost Fennoscandia. For Ecol Manage 142:65–77
Nota K, Klaminder J, Milesi P et al (2022) Norway spruce postglacial recolonization of Fennoscandia. Nat Commun 13:1333
O’Leary MH (1988) Carbon isotopes in photosynthesis. Bioscience 38:328–336
Olsson F, Gaillard M-J, Lemdahl G et al (2010) A continuous record of Fire covering the last 10,500 calendar years from southern Sweden – the role of climate and human activities. Palaeogeogr Palaeoclimatol Palaeoecol 291:128–141
Parducci L, Jørgensen T, Tollefsrud MM et al (2012) Glacial Survival of Boreal Trees in Northern Scandinavia. Science 335:1083–1086
Paus A, Haugland V (2017) Early- to Mid-holocene forest-line and climate dynamics in southern Scandes mountains inferred from contrasting megafossil and pollen data. Holocene 27:361–383
Regnéll C, Mangerud J, Svendsen JI (2019) Tracing the last remnants of the scandinavian ice sheet: ice-dammed lakes and a catastrophic outburst Flood in northern Sweden. Quat Sci Rev 221:105862
Remy CC, Magne G, Stivrins N et al (2023) Climatic and vegetational controls of Holocene wildfire regimes in the boreal forest of northern Fennoscandia. J Ecol 111:845–860
Rogers BM, Soja AJ, Goulden ML, Randerson JT (2015) Influence of tree species on continental differences in boreal Fires and climate feedbacks. Nat Geosci 8:228–234
Ryzhkova N, Kryshen A, Niklasson M et al (2022) Climate drove the Fire cycle and humans influenced Fire occurrence in the East European boreal forest. Ecol Monogr 92:e1530
Schimmel J (1993) On Fire. Fire behavior, fuel succession and vegetation response to Fire in the Swedish boreal forest. Swedish University of Agricultural Sciences, Umeå. Dissertation
Schimmel J, Granström A (1997) Fuel succession and Fire behavior in the Swedish boreal forest. Can J for Res 27:1207–1216
Seftigen K, Linderholm HW, Loader NJ, Liu Y, Young GHF (2011) The influence of climate on 13C/12C and 18O/16O ratios in tree ring cellulose of Pinus sylvestris L. growing in the central Scandinavian Mountains. Chem Geol 286:84–93
Seppä H, Bjune AE, Telford RJ, Birks HJB, Veski S (2009) Last nine-thousand years of temperature variability in Northern Europe. Clim Past 5:523–535
Sharma S, Williams DG (2009) Carbon and oxygen isotope analysis of leaf biomass reveals contrasting photosynthetic responses to elevated CO2 near geologic vents in Yellowstone National Park. Biogeosciences 6:25–31
Sjögren P, Damm C (2019) Holocene vegetation change in northernmost Fennoscandia and the impact on prehistoric foragers 12 000-2000 cal. A bp – a review. Boreas 48:20–35
Stroeven AP, Hättestrand C, Kleman J et al (2016) Deglaciation of Fennoscandia. Quat Sci Rev 147:91–121
Thonicke K, Venevsky S, Sitch S, Cramer W (2001) The role of Fire disturbance for global vegetation dynamics: coupling Fire into a dynamic global vegetation model. Glob Ecol Biogeogr 10:661–677
Wardle DA, Hörnberg G, Zackrisson O, Kalela-Brundin M, Coomes DA (2003) Long-term effects of wildfire on ecosystem properties across an island area gradient. Science 300:972–975
Zackrisson O (1977) Influence of forest Fires on the North Swedish Boreal Forest. Oikos 29:22–32
Zackrisson O, DeLuca TH, Nilsson M-C, Sellstedt A, Berglund LM (2004) Nitrogen fixation increases with successional age in boreal forests. Ecology 85:3327–3334
Zale R, Huang Y-T, Bigler C et al (2018) Growth of plants on the late Weichselian ice-sheet during Greenland interstadial-1? Quat Sci Rev 185:222–229
Acknowledgements
We want to thank Iréne Gustafson, Anna-Maria Rautio and Ture Hennerson, all affiliated with Skogsmuseet (the Forestry Museum) in Lycksele, for providing samples from the pine trees that were originally excavated in the 1980s. We also want to thank Oscar Lundgren for assistance during latewood sampling and acknowledge funding from Vetenskapsrådet (Swedish research council) (2017–04548) and the Wallenberg Academy Fellow program to J. Klaminder.
Funding
Open access funding provided by Umea University.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
Authors have no financial or non-financial interests that are directly or indirectly related to the work of this study.
Additional information
Communicated by M.-J. Gaillard.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Klaminder, J., Fassl, M., Baudet, M. et al. Landscape of ice and fire – uniquely well-preserved Scots pine trunks reveal forest fires near the retreating Weichselian ice margin. Veget Hist Archaeobot 33, 519–527 (2024). https://doi.org/10.1007/s00334-023-00974-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-023-00974-6