
Abstract
In the marine environment, the persistence and abundance of Vibrio are related to a number of environmental parameters. The influence of the different environmental variables in determining the Vibrio occurrence could be different in the specific geographic areas around the world. Moreover, oceanographic parameters are generally interdependent and should not be considered separately when their influence on bacterial presence and concentration is tested. In this study, an integrated approach was used to identify key parameters determining the abundance of Vibrio spp in marine samples from the Venetian Lagoon in Italy, which is an important area for fish farming and tourism. Multivariate techniques have been adopted to analyze the dataset: using PCA, it was shown that a relatively high proportion of the total variance in this area was mainly due to two independent variables, namely salinity and temperature. Using cluster analysis, it was possible to categorize different groups with homogeneous features as regards space (“stations”) and time (“seasons”) distribution, as well as to quantify the values of environmental variables and the Vibrio abundances in each category. Furthermore, integrating key environmental factors and bacterial concentration values, it was possible to identify levels of salinity and sea surface temperature which were optimal for Vibrio concentration in water, plankton, and sediment samples. The identification of key environmental variables conditioning Vibrio occurrence should facilitate ocean monitoring, making it possible to predict unexpected variations in marine microflora which determine possible public health risks in coastal areas.
Similar content being viewed by others
Introduction
The impact of climate change on the emergence and re-emergence of many infectious diseases is now recognized [14, 16, 28, 29]. Climatic factors have been shown to influence environment-related diseases such as malaria, dengue, West Nile virus, and Rift Valley fever, and to be significant in cholera epidemics [7, 11, 28]. Oceanic environmental parameters also condition the presence, persistence, and spread of bacteria belonging to the genus Vibrio, which are a predominant part of autochthonous marine microflora. Notably, remote sensing employing satellites has established that Sea Surface Temperature (SST) and Sea Surface Height (SSH) correlate with cholera epidemics in Bangladesh [20, 21]. Other studies in the Bay of Bengal and Peru have identified temperature as a key factor associated with increased counts of Vibrio cholerae and cases of cholera [7, 12, 15]. Environmental parameters may influence Vibrio occurrence differently in the different areas of the world and so there is no general agreement on which parameters are truly determinant in favoring the persistence and abundance of vibrios. For example, it has been shown [9] that in the Baltic Sea, water temperature alone did not significantly affect the abundance of total Vibrio spp or of individual Vibrio populations. In contrast, cyanobacteria-derived organic matter was an important factor regulating the growth and abundance of V. cholerae and Vibrio vulnificus in brackish waters [9]. In a study conducted in Brazil [26], the results obtained in oysters and water samples showed no correlation between temperature, salinity, and bacteria. Moreover, some environmental factors might influence the total Vibrio concentration, while others seem to affect the emergence of cholera cases by influencing the distribution of vibrios or even the contact between pathogens in water and plankton and the host. Sea surface height, for example, is an indicator of incursion of plankton-laden water inland and was found to correlate with cholera outbreaks [21]. It has also been proposed that aeroplankton, under the influence of wind strength and direction, could transport cholera bacteria from one body of water to an adjacent one [24].
There is limited information for the Italian coast about the presence and abundance of Vibrio or about their correlation with environmental variables [2, 5, 6, 8, 32]. Some studies indicate a correlation between seawater temperature and the presence of Vibrio [2, 6, 8, 32], although temperature seems not to be the only factor implicated. In this study, we analyzed the Vibrio spp concentration and distribution, in time and space, in the coastal waters of the Northern Adriatic Sea, where the influence of riverine inputs rich in pollutants, industrial wastes, and nutrients is substantial. This stretch of water, particularly along the coast of the Veneto region, is subject to anthropogenic disturbance, and changes in its environmental characteristics resulting from global warming have recently been detected [3, 17, 30]. The area was chosen for study because of year-round fish farming and because it is a popular recreational bathing zone during the summer. The study began within the framework of the International “VibrioSea” Project (http://www.redgems.org/spip.php?article10), which aimed to develop a satellite-based early warning system for Vibrio-related outbreaks of disease.
Materials and Methods
Study Area and Sampling Sites
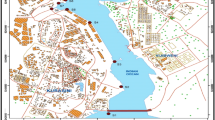
The area included in this study extends north to south from the Venetian Lagoon to the Po river, along a 30-km stretch of coast. Three stations were chosen: San Pietro, Fosson, and Caleri (Fig. 1). The S. Pietro station, which lies in front of the Malamocco inlet, the middle entrance to the Venice Lagoon, was selected as a potentially “low-risk” area, while the other two stations, which lie between the Brenta and Adige rivers (Fosson) and northern to the Po delta (Caleri), are situated in areas considered to be “high risk” on the basis of conditions that favor Vibrio persistence (estuarine-like waters).

Studied area and sampling sites in the Northern Adriatic Sea, facing the Venetian Lagoon
At each station, two sampling sites were chosen, making a total of six sites: one 500 m from the coastline (Caleri1, Fosson1, and S. Pietro1; bottom depths ranging from 4 to 6 m) and the second 3,000 m offshore (Caleri2, Fosson2, and S. Pietro2; bottom depths ranging from 10 to 16 m). To obtain additional samples from waters of low salinity, a seventh site (Foce), located inside the mouth of the Adige river (bottom depth = 4 m) (Fig. 1), was included in the study from May 2007.
Sample Collection and Processing
Water, plankton, and sediment samples were collected twice monthly during the warm season (June to September) and once each month from October to May, covering a period from June 2006 to November 2007 for six stations, while Foce was sampled from May 2007 to November 2007. Two liters of water was collected in the water column using a sterile Niskin plastic bottle at 1 m depth. Mesozooplankton were sampled using a 200-μm-mesh-size plankton net to filter approximately 105 l of water. Surface sediment samples (0–2 cm) were obtained using a Van Veen grab from the three coastal stations only (Caleri1, Fosson1, and S. Pietro1) and were stored in sterile plastic bottles. All samples were transported to the laboratory in an insulated box and were processed within 24 h of collection.
For microbiological studies, water samples (2 l) were concentrated by filtering through a 0.22-μm Millipore nitrocellulose membrane; cells in the filter were suspended in 300 ml of artificial sea water (ASW) [19]. Plankton-containing water samples were filtered using a 100-μm-mesh-size plankton net; the plankton was then weighed and suspended in ASW, homogenized, sonicated three times on ice at a low frequency (15 kHz for 30 s) to detach bacteria, vortexed, and centrifuged. Approximately 5 g of sediment was weighed and suspended in ASW, vortexed for 2 min, sonicated for 2 min as described above, and stored at RT for several minutes.
Environmental Parameter Measurements
Vertical profiles from sea surface to the bottom were determined at each station using a multiparametric probe (Idronaut® mod. 316). Data on temperature (°C), salinity (PSU), dissolved oxygen (% of saturation), pH, and turbidity (NTU) were acquired using a personal computer. To get a more accurate picture of the seston concentration in northern Adriatic coastal waters, total suspended matter (TSM, in milligrams per liter) was calculated on the basis of a regression between turbidity measurements (made using the multiparametric probe) and TSM sample determinations (made concurrently by ISMAR in the Northern Adriatic Sea in the period 2003–2006; linear equation—TSM = 0.606704 turb + 1.864584; correlation—n = 633, r = 0.550, p < 0.01). Additional water samples were collected for chlorophyll laboratory determinations, following a fluorometric protocol [10, 13]. At each site, 1 l of water sampled at 1 m depth was filtered using glass fiber filters (Whatman GF/C, nominal porosity = 0.7 μm). Filters were disrupted using an Ultra-Turrax homogenizer (20,000 rpm) and the pigments extracted in 90% acetone. Fluorescence was measured using a Perkin-Elmer luminometer LS-5B, before and after acidification with HCl 1 N [10]. Standardization with a pure chlorophyll solution and related calculations gave chlorophyll and pheopigment concentrations. Chlorophyll and pheopigments are described individually in the results, while in the statistical analyses (correlation matrix, PCA, and cluster analysis) their concentrations were added to each other and reported as “total pigments”.
Count of Total Vibrio spp in Marine Samples
The most-probable-number method associated with standard PCR (MPN-PCR) was used to enumerate culturable Vibrio spp in water, plankton, and sediment samples, and to confirm the identification of the bacteria as belonging to the genus Vibrio. Five dilution series (1:10) were carried out in ten replicate culture tubes, starting with 0.5 ml of prepared samples and 4.5 ml of modified alkaline peptonate water (m-APW, pH 8.5 and 3.5% NaCl) as the growth medium. Tubes were incubated at 37°C for 7 h and growth was assessed by visual inspection, coupled with standard PCR on cell pellets obtained by centrifugation (10 min, 4,500 rpm) of m-APW tubes showing visible growth. Moreover, a loop from each of the m-APW tubes showing visible growth was streaked onto TCBS-containing plates to confirm the presence of typical Vibrio colonies. DNA for the PCR reactions was prepared using a standard protocol and PCR assays were carried out in a Gene Amp PCR System 9700 Thermal Cycler (Perkin-Elmer). Each reaction was conducted in a final volume of 50 μl, using 100 ng of DNA template, 0.2 μM of forward and reverse primers, 200 μM of each dNTPs, 1.5 mM MgCl2, and 2 U Taq polymerase (5 U/μl; Advanced Biomedicine) in 10× buffer [160 mM (NH4)2SO4, 670 mM Tris–HCl pH 8.8, 0.1% Tween20; Advanced Biomedicine]. A primer pair designed on the 16S rDNA sequence of Vibrio was used (FWD—5′ CTCGCGTCAGGATATGCCTA 3′ and REV—5′ ACAGTACTCTAGT CTGCCAGTTTC 3′). The PCR reaction mix was subjected to an initial activation step at 95°C for 5 min, followed by 30 cycles of denaturation at 95°C for 30 s, annealing for 45 s at 53°C, and extension at 72°C for 45 s, with a final extension at 72°C for 10 min. PCR products were visualized by staining with ethidium bromide (0.5 μg ml−1); the amplicons were separated on a 1.8% agarose gel and photographed using an Image Master VDS (Pharmacia Biotech).
Statistical Analysis
Statistical analysis was carried out on a matrix prepared by assembling data on hydrology and Vibrio concentrations in water, plankton, and sediments. Vibrio in water and plankton were related to surface probe data (1 m depth), while Vibrio in sediments were coupled with hydrology measured at the bottom (±0.5 m above). A non-parametric multivariate multiple regression was utilized [1, 25]. Individual variables were analyzed separately for their relationship with the Vibrio concentration (ignoring other variables), and variables were then subjected to a stepwise forward-selection procedure to develop a model. All non-parametric multivariate multiple regressions were done using the computer programs DISTLM® and STATISTICA® [1, 31], with a previous log10 transformation of biological data (i.e., chlorophyll, pheopigments, total pigments, and Vibrio concentrations) because of their non-normal distribution.
Results
Environmental Parameter Distribution in the Study Area of the Adriatic Sea
A basic statistics analysis relating to the measurements at the surface (average values and relative standard deviations) was performed in the defined area (Supporting information—Fig. S1). Regarding sea surface temperature (SST), minima around 9°C were measured everywhere in January, while maxima, between 27° and 28°C, were observed in July. The slight differences between the average temperatures of the seven stations in 2006 and 2007 (range = 18.8–21.2°C) were not statistically significant.
Sea Surface Salinity
Differences were observed in the spatial distribution of sea surface salinity values: due to the influence of the lagoon water, which is not as diluted as riverine water, salinity values at S. Pietro were higher than elsewhere (S.Pietro1 avg = 35.4 ± 1.4 PSU; S. Pietro2 avg = 35.1 ± 1.6 PSU). The other stations, Fosson and Caleri, showed lower salinities (Fosson1 avg—31.6 ± 3.4 PSU, Fosson2 avg—31.6 ± 4.1 PSU, Caleri1 avg—32.6 ± 3.4 PSU, Caleri2—32.9 ± 2.5 PSU) due to the direct influence of the Brenta, Adige, and Po rivers (respective average discharges over the last 30 years = 93 m3 s−1, 235 m3 s−1, and 1,470 m3 s−1). The lowest salinities (SSSmin = 4.8 PSU; SSSavg = 23.7 ± 9.0 PSU) were measured at Foce, which is located inside the mouth of the Adige river.
Oxygen Saturation and pH
At the surface, dissolved oxygen saturation averaged from 98 (Foce, SD = ±11%) to 108% (Fosson2, SD = ±19%), with no hypoxia phenomena detected during the samplings. pH distribution ranged between averages of 8.15 (±0.12) at Foce and 8.21 U (±0.07) at Caleri2. The lowest pH of 7.90, measured at Foce, corresponded to the salinity minimum of 4.8 PSU, as expected in freshwaters.
Turbidity
Turbidity is a signal of freshwater distribution: in fact, this parameter is closely related to the river runoff, being consequently inverse respect to salinity. In our data set, turbidity maxima were found at Foce (avg = 10.4 NTU ≈ 8.2 mgTSM l−1), inside the Adige river, while turbidity minima (=higher transparency) were measured in front of the Venice Lagoon, at S. Pietro (avg = 2.0 NTU, corresponding to 3.1 mgTSM l−1, both at 500 m and 3,000 m sites), where salinity was higher. Intermediate values were detected at Fosson1, Fosson2, and Caleri2 (≈3.3 NTU, ≈3.9 mgTSM l−1), and at Caleri1, influenced by the Po river (avg = 4.5 NTU ≈ 4.6 mgTSM l−1).
Pigment Concentration
Chlorophyll a concentrations averaged around 1.1–1.9 μg l−1; non-significant differences were computed between 500 and 3,000 m sites. Values >6 μg l−1 were observed at S. Pietro2, Fosson1, Caleri1, and Caleri2, in both 2006 and 2007, between the end of July and the end of August, when the phytoplankton biomass peaked. Pheopigment distribution (the degradation products of living chlorophyll, mainly due to the natural senescence of phytoplankton populations and to the grazing pressure from herbivorous organisms) was similar to that of chlorophyll. Values increased progressively from high to low salinity waters, i.e., from S. Pietro1 and 2 to Fosson2, Caleri2, Caleri1, Fosson1, and Foce. A maximum >5 μg l−1 was measured at Fosson1, at the end of the growth season (October 2006). These studies regarding environmental variables allowed us to accurately characterize the different sampling sites in order to analyze possible correlations between microbiological and environmental data.
Temporal Variation of the Oceanographic Parameters in the Adriatic Coastal Area
Time trends of SST, SSS, and chlorophyll over the entire study period are shown in Fig. 2. As already observed, temperatures were very homogeneous and practically equivalent in all stations. Nevertheless, some differences can be detected between 2006 and 2007: although our samplings started in June 2006 (so we have no information about water temperatures in the preceding months), it was observed that in 2007 water temperatures exceeding 20°C occurred a month earlier than in 2006 (from June–October, 2006 to May–September, 2007). This could be due to the different air temperatures measured in 2006 and 2007: from December 2006 to February 2007, and in April 2007 again, average air temperature was higher than the monthly historical maximum, leading to an increase of air temperature (and consequently water temperature) in the first half of 2007 (data from ARPAV; http://www.arpa.veneto.it/acqua/htm/documenti.asp).

Time trends of SST (sea surface temperature), SSS (sea surface salinity), and chlorophyll a at all stations
As concerns SSS (Fig. 2), this parameter varied over time in a random sequence, oscillating between 35 and 28 PSU in the S. Pietro and Caleri stations (apart from a substantial decrease at Caleri during the summer of 2006). The salinity trend was less homogeneous at Fosson because of its vicinity to the estuary of the river Adige; in some cases, it reached values <20 PSU (Fosson2, 19/09/2006; Foce, 27/08/2007), down to the minimum of 4 PSU (12/06/2007). As for chlorophyll a concentration, it is possible to identify two seasonal peaks corresponding to the periods of maximum SST (i.e., summer) at all the stations, although the second peak, corresponding to the summer of 2007, is not well defined at S. Pietro site (Fig. 2).
The Abundance of Vibrio in Marine Samples
In Water and Plankton
Figure 3 shows the Vibrio concentration in water and plankton during the study period at the Foce, Caleri1, and S. Pietro1 stations, which were regarded as representative sites. Maxima peaks of Vibrio concentration in water were detected during the summers of 2006 and 2007, with major peaks in July and September 2006 and May and August 2007. However, some differences can be observed: there was, for example, a significant delay of about 45 days in reaching the peak of maximum Vibrio concentration in water during summer 2007 at the San Pietro station. Again at this station, the concentration of bacteria in water decreased much more (two logs more on average) at the end of summer 2006 and in April 2007 than bacterial concentrations detected at Caleri and Foce.

Vibrio spp concentrations in water, plankton, and sediments in the three most representative stations S.Pietro1, Foce, and Caleri1
Data on bacterial abundance in plankton were homogenous at the different sampling sites, and in general presented the same temporal distribution as that of vibrios in water. However, an additional peak in bacterial counts in plankton, but not in water, was observed during the winter (December 2006 to February 2007).
Vibrio Abundances in Sediments
Vibrio concentration in sediments was evaluated beginning from August 2006 and showed a maximum peak in that year during the month of October. Afterwards, concentrations remained low until the summer, when bacterial concentration gradually increased, reaching a peak in July–August 2007 and another in October. While data obtained from samples taken in 2006 were homogenous among the different stations, significant differences can be observed during the year 2007: Vibrio concentrations were particularly high at Foce and much lower in S. Pietro during spring (May and June).
Maximal Bacterial Concentrations
Vibrios in water were as abundant as in sediments (about 105 bacteria ml−1), while total Vibrio in plankton reached concentrations as high as 108 bacteria ml−1. Comparing bacterial abundance in 2006 with that in 2007, there was a slight difference between the two years for Vibrio concentration in water, and a difference of about two logs for Vibrio concentration in plankton and in sediments, with the higher concentrations detected in 2006.
The most abundant species of Vibrio isolated in this area was Vibrio alginolyticus, followed by Vibrio harveyi. Of the human pathogenic species of Vibrio, V. cholerae, Vibrio parahaemolyticus, and V. vulnificus, by far the most abundant species was V. parahaemolyticus (195 strains). Few strains of V. cholerae and none of V. vulnificus were ever isolated.
Statistical Tools used to Evaluate Possible Correlations Between Environmental Variables and Vibrio spp Concentration
To relate the presence of Vibrio to environmental parameters, bacteria abundances in water and plankton were coupled with surface oceanographic data (all sampled at 1 m depth). Seasonality was highlighted by the direct dependence of biological cycles on temperature: when temperature peaked, chlorophyll (as phytoplankton biomass) and, consequently, dissolved oxygen increased, along with Vibrio concentrations in water and in plankton. Salinity correlated inversely with turbidity, chlorophyll, and Vibrio in water, so that phytoplankton and bacteria abundances were higher in waters with low salinity. Vibrio in sediments, when correlated with the oceanographic data from close to the bottom, showed a significant inverse relationship only with pH (r = −0.268).
PCA Analysis
PCA analysis is a useful tool for quantifying the contribution of each variable to the environmental variability of a region, and for visualizing the variables in two or more dimensions, called factors [23]. When applied to our environmental dataset (SST, SSS, pH, DO%, turbidity, and total pigments), the analysis showed that a substantial fraction of the total variance (54.3%) was due to factors I and II (factor I—24.4%; factor II—29.9%); of these factors, the environmental variables salinity and temperature were the most significant, accounting for 19% and 15%, respectively, of the variability of the whole system (Supporting information—Fig. S2). Other parameters also contributed, but their influence was mainly indirect, i.e., mediated by salinity or temperature. Indeed, turbidity is inversely linked to salinity (r = −0.733) because of the high seston content of diluted waters; chlorophyll is a direct function of temperature (r = 0.340) because phytoplankton growth is strictly dependent on the seasonal cycles; dissolved oxygen is directly related to chlorophyll, as a final product of photosynthesis (r = 0.455) and, consequently, to temperature (r = 0.504) (Supporting information—Fig. S2).
Cluster Analysis
Cluster analysis is a statistical method for grouping objects of similar kind into categories [18]. We applied this method to the oceanographic matrix (hydrology and Vibrio, in water and plankton) in order to verify the grouping of samples belonging to different stations and seasons in homogeneous clusters (Supporting information—Fig. S3). Results from the first category (“homogeneous stations”) lead to the identification of three groups with similar features (Supporting information—Fig. 3A): group 1, represented by just one station, Foce; group 2, in which Caleri1, Caleri2, Fosson1, and Fosson2 were placed together; and group 3, with S.Pietro1 and S.Pietro2. Within each cluster, the environmental characteristics were very similar.
Subsequently, we organized the whole dataset (oceanographic parameters + Vibrio abundance) within these three categories. The basic statistics for all the variables, calculated for each group, are shown in Table 1.
Group 1: this group is represented only by the station at Foce, which is clearly separated from the others due to the lowest salinities measured here. Oxygen and pH were lowest here, while turbidity and chlorophyll were highest. There was a tenfold difference when bacteria concentration in water and plankton were compared, with much higher Vibrio concentration in plankton. V. parahaemolyticus was isolated at all sampling sites, but was found at highest frequency at the Foce site, where 100% of the samples collected tested positive for this pathogenic species.
Group 2: this grouping of data from the Fosson and Caleri sites, at both 500 and 3,000 m, was characterized by intermediate values for environmental variables, between those of groups 1 and 3. Vibrio concentrations in water were twice as high as in groups 1 and 3, while the concentration on plankton was six times higher than at the other stations.
Group 3: this cluster of data comes from the S. Pietro sites. This station is mainly influenced by waters with marine-like features, having intermediate temperatures, high salinities, oxygen >100%, low turbidity, and low chlorophyll concentration. Here, Vibrio abundances in water and plankton were more or less the same as at Foce. Fewer human pathogenic Vibrio strains were isolated here than in the other groups, with only 45% of samples containing V. parahaemolyticus.
To determine the “homogeneous seasons” categories, a new cluster analysis was performed to detect similarities between different seasonal periods. The analysis produced three further groups, this time called groups A, B, and C (Supporting information—Fig. 3B). As previously, the variables were allocated into these three clusters and means and standard deviations were calculated (Table 2).
Group A (winter data): in this season, biological activity is particularly reduced due to the low temperatures (avgs = 10.1°C); so phytoplankton biomass, along with Vibrio concentrations in water and plankton, reached their minima.
Group B (summer): high water temperature favored the phytoplankton growth (and consequently its biomass, measured as chlorophyll a and dissolved oxygen production) as well as Vibrio presence in water and plankton which reached their maxima. Most (84%) of the Vibrio human pathogenic species analyzed in this study, mainly V. parahaemolyticus, were isolated during this period, when the SST was higher than 22°C.
Group C (spring + autumn): in this group, the studied variables had intermediate values. Concerning pathogenic Vibrio species, 16% of the V. parahaemolyticus strains were isolated during spring or fall, when the SST ranged between 15 and 20°C.
The cluster analysis conducted using bacterial concentration in sediments and environmental variables measured at sea bottom identify three groups corresponding to those described for environmental data recorded in the surface (Table 3). In group A (winter), Vibrio concentration reached its minimum while in group B, corresponding to summer, the concentration is ten times higher. Surprisingly, the highest concentration of bacteria in sediments was detected in group C during intermediate seasons corresponding to spring and autumn (Table 3).
A further analysis was conducted integrating Vibrio concentration and the two key environmental variables, SST and SSS. As shown in Fig. 4, salinities around 30–35 PSU and maximal temperatures (27–29°C) seem to be the optimal range for vibrios concentration in plankton, while the same salinity range but associated with lower temperatures (18–24°C) favored the presence of Vibrio in sediments. Free vibrios in water are favored at lower salinity ranges (20–28 PSU) both at intermediate (17–23°C) and high temperatures (27–29°C).

Combined effect of temperature and salinity on Vibrio abundance in water, plankton, and sediment in all the stations
Discussion
The coastal environment of the North Adriatic Sea is characterized by low sea depth, fine sediments, exchange with water from the Venetian Lagoon, and the contribution of numerous rivers which transport the products of agricultural, urban, and industrial activities. Over the centuries, the Venetian Lagoon has been subjected to the diversion of the main rivers outside the lagoon, altering its original estuarine nature. The area is strongly influenced by the great quantity of river freshwater and by saline water arriving from the Eastern Mediterranean Sea. Anthropogenic pressure must also be taken into account because the Veneto is a region with considerable tourist and recreational development. All these characteristics, together with meteorological and hydrodynamic variability, make this environment very susceptible to sudden changes in chemical–physical properties and consequent biological changes [4, 30]. It has been demonstrated that environmental changes in recent years were associated with the emergence of new pathogens, the re-emergence of diseases which had almost been eradicated, and with changing patterns and distribution of numerous infectious agents [16, 27, 28]. Hence, permanent monitoring of environmental parameters is mandatory to predict and prevent climate-associated diseases and threats.
Most of the studies regarding the influence of environmental parameters on Vibrio occurrence report the effect of individual oceanographic variables on Vibrio presence and/or abundance not always taking into account the inter-relations existing among these parameters [9, 12, 15, 26]. In the present study, we have considered globally a number of parameters to accurately characterize sampling sites in the study area and, in this context, be able to analyze the presence and abundance of the Vibrio population. PCA analysis allowed us to establish that salinity and temperature, with a contribution of 19% and 15%, respectively, of the total variance, are the most important environmental parameters determining the variability of the coastal waters of the Northern Adriatic Sea. However, other environmental parameters, chlorophyll concentration, and turbidity contribute to modulate the specific correlation. In a recent report [22], salinity was indicated as the primary factor governing the temporal and spatial distribution of V. parahaemolyticus, whereas seawater temperature had a secondary effect and only modulated the abundance in periods and areas of reduced salinities. However, the low occurrence and abundance of V. parahaemolyticus during the summer observed by these authors contrasts with previously reported patterns of presence in other areas of the world and with the data in the present study where the highest Vibrio densities were obtained during the warmest months of summer and low or undetectable levels were observed in winter. Data are not completely comparable, however, in that different Vibrio populations seem to be regulated by different environmental variables [9].
The analysis of samples from the station Foce, located within an estuarine area, led to interesting observations. Surprisingly, the peculiar oceanographic features of this site (low salinity, very high turbidity) did not correspond to a significant variation in Vibrio concentrations which, in fact, showed values very similar to those found at S. Pietro, a site located in a completely different area and belonging to another cluster (high salinity, low turbidity). The only difference between the two types of environmental niche concerned Vibrio concentrations in sediments, which was four times lower at S. Pietro sites.
The sites (Caleri1 and 2 and Fosson1 and 2) located close to the main Veneto rivers, the Po, Adige, and Brenta, presented intermediate levels of salinity and turbidity. Here, average Vibrio abundances reached their maximal values, both in water, where they were twice as high as in the other groups, and in plankton, where they were four to six times higher. In sediment, average bacteria concentrations were similar to those at Foce. The seasonal distribution of Vibrio concentrations showed that in winter, when the SST is lower than 15°C, the phytoplankton biomass, as well as Vibrio concentrations in water, plankton, and sediments, reached their minimal values. This was expected as it is known that Vibrio spp tend to be more common in warm waters, notably when temperatures exceed 17°C [9]. In summer, Vibrio counts in water and plankton reached their maxima, while intermediate vibrios concentrations were found in spring and fall. An exception was the concentration of Vibrio spp in sediment during the intermediate seasons, which was higher than in any other period of the year. Comparing these results with other published data, it is quite clear that the correlation environmental factors–bacteria concentration does not function in the same way in the different geographic sites, and that in the different cases one or other oceanic parameter has the highest influence on presence and concentration of Vibrio. Huq et al. [15] found a very strong and consistent relationship between the ecological predictors and cholera incidence for one of the Bangladesh area examined but, although the general trends were relatively strong in the whole area included in the study, they did not find a consistency of prediction in other geographical sites located in the same area. In other studies, the influence of the classical environmental factors, salinity, nutrient concentration, and sea temperature, has not been demonstrated as is the case of Vibrio present in Brazilian oysters [26]. Another source of variability underlying the influence of oceanic factors on Vibrio presence is the specific marine micro-environment as demonstrated by Vezzulli et al. [32] who showed that in the pelagic environment, total variance in culturable Vibrio data was explained by sea surface temperature, salinity, and organic matter concentration while in the benthic environment, sea surface temperature was the only factor that significantly affected culturable Vibrio occurrence. On the basis on all these data, determination of Vibrio occurrence and abundance by environmental conditions appears as a very complex phenomenon which has to be examined using integrated analysis and considering local geographic characteristics.
In the present study, no significant differences were reported between environmental variable values in 2006 and 2007, with the SST values for these two years being the highest since 2000, with the exception of the year 2003. However, a significant difference (about one log on average) was observed between bacteria concentrations recorded in 2006 and 2007. This seems to indicate that other environmental, climatic, and/or biological factors in addition to the “classic” factors like SST, SSS, and nutrients may play a minor role in controlling bacterial concentrations in the marine environment. The influence of wind and climatic events in V. cholerae dissemination demonstrated by Paz and Broza [24] is an example.
In this study, a series of statistical tools were applied to correlate Vibrio concentrations with environmental variables, which were not taken independently but were analyzed using an integrated approach. This was done because frequently the relationship between one or more environmental factors and Vibrio counts, although statistically significant, is not a direct correlation but a relation to which a number of variables contribute. Identification and monitoring of key parameters in this specific geographic area is important to predict unexpected variations in the marine microflora, which determine possible public health risks in coastal areas.
References
Anderson MJ (2011) DISTLM v.5: a FORTRAN computer program to calculate a distance-based multivariate analysis for a linear model. Department of Statistics, University of Auckland, Auckland, http://www.stat.auckland.ac.nz/∼mja/prog/DISTLM_UserNotes.pdf (2011)
Baffone W, Tarsi R, Pane L, Campana R, Repetto B, Mariottini GL, Pruzzo C (2006) Detection of free-living and plankton-bound vibrios in coastal waters of the Adriatic Sea (Italy) and study of their pathogenicity-associated properties. Environ Microbiol 8:1299–1305
Barale V, Schiller C, Tacchi R, Marechal C (2005) Trends and interactions of physical and bio-geo-chemical features in the Adriatic Sea as derived from satellite observations. Sci Total Environ 353:68–81
Bernardi Aubry F, Berton A, Bastianini M, Socal G, Acri F (2004) Phytoplankton succession in a coastal area of the NW Adriatic, over a 10-year sampling period (1990–1999). Continent Shelf Res 24:97–115
Caburlotto G, Gennari M, Ghidini V, Tafi M, Lleo MM (2009) Presence of T3SS2 and other virulence-related genes in tdh-negative Vibrio parahaemolyticus environmental strains isolated from marine samples in the area of the Venetian Lagoon, Italy. FEMS Microbiol Ecol 70:174–182
Caburlotto G, Bradd JH, Lleo MM, Huq A, Colwell RR (2009) Serodiversity and ecological distribution of Vibrio parahaemolyticus in the Venetian Lagoon, Northeast Italy. Environ Microbial Rep 2:151–157
Colwell RR (1996) Global climate and infectious disease: the cholera paradigm. Science 274:2025–2031
Covazzi Harriague A, Brino MD, Zampini M, Albertelli G, Pruzzo C, Misic C (2008) Vibrios in association with sedimentary crustaceans in three beaches of the northern Adriatic Sea (Italy). Mar Pollut Bull 56:574–579
Eiler A, Gonzalez-Rey C, Allen S, Bertilsson S (2007) Growth response of Vibrio cholerae and other Vibrio spp. to cyanobacterial dissolved organic matter and temperature in brackish water. FEMS Microbiol Ecol 60:411–418
EPA (1997) In vitro determination of chlorophyll a and pheophytin a in marine and freshwater algae by fluorescence. Method 445.0; National Exposure Research Laboratory, Office of Research and Development. U.S. Environmental Protection Agency, Cincinnati, p 22
Fisman DN (2007) Seasonality of infectious diseases. Annu Rev Public Health 28:127–143
Gil A, Louis V, Rivera I, Lipp E, Huq A, Lanata C, Taylor D, Russek-Cohen E, Choopun N, Sack R, Colwell RR (2004) Occurrence and distribution of Vibrio cholerae in the coastal environment of Peru. Environ Microbiol 6:699–706
Holm-Hansen O, Lorenzen CJ, Holmes RW, Strickland DH (1965) Fluorometric determination of chlorophyll. ICES J Mar Sci 30:3–15
Hoskisson PA, Trevors JT (2010) Shifting trends in pathogen dynamics on a changing planet. Antonie Van Leeuwenhoek 98:423–427
Huq A, Sack RB, Nizam A, Longini IM, Nair GB, Ali A, Morris JG Jr, Khan MN, Siddique AK, Yunus M, Albert MJ, Sack DA, Colwell RR (2005) Critical factors influencing the occurrence of Vibrio cholerae in the environment of Bangladesh. Appl Environ Microbiol 71:4645–4654
Jones KE, Patel NG, Levy MA, Storeygard A, Balk D, Gittleman JL, Daszak P (2008) Global trends in emerging infectious diseases. Nature 451:990–993
Lejeusne C, Chevaldonné P, Pergent-Martini C, Boudouresque CF, Pérez T (2010) Climate change effects on a miniature ocean: the highly diverse, highly impacted Mediterranean Sea. Trends Ecol Evol 25:250–260
Liu H, Weng Q, Gaines D (2008) Spatio-temporal analysis of the relationship between WNV dissemination and environmental variables in Indianapolis, USA. Int J Health Geograph 7:66
Lleo MM, Bonato B, Benedetti D, Canepari P (2005) Survival of enterococcal species in aquatic environments. FEMS Microbiol Ecol 54:189–196
Lleo MM, Lafaye M, Guell A (2008) Application of space technologies to the surveillance and modelling of waterborne diseases. Curr Opin Biotechnol 19:307–312
Lobitz B, Beck L, Huq A, Wood B, Fuchs G, Faruque A, Colwell RR (2000) Climate and infectious disease: use of remote sensing for detection of Vibrio cholerae by indirect measurement. Proc Natl Acad Sci USA 97:1438–1443
Martinez-Urtaza J, Lozano-Leon A, Varela-Pet J, Trinanes J, Pazos Y, Garcia-Martin O (2008) Environmental determinants of the occurrence and distribution of Vibrio parahaemolyticus in the rias of Galicia, Spain. Appl Environ Microbiol 74:265–274
McArdle BH, Anderson MJ (2001) Fitting multivariate models to community data: a comment on distance-based redundancy analysis. Ecology 82:290–297
Paz S, Broza M (2007) Wind direction and its linkage with Vibrio cholerae dissemination. Environ Health Perspect 115:195–200
Primpas I, Tsirtsi G, Karydis M, Kokkoris GD (2010) Principal component analysis: development of a multivariate index for assessing eutrophication according to the European water framework directive. Ecol Indicators 10:178–183
Ristori CA, Iaria ST, Gelli DS, Rivera IN (2007) Pathogenic bacteria associated with oysters (Crassostrea brasiliana) and estuarine water along the south coast of Brazil. Int J Environ Health Res 17:259–269
Rose JB, Epstein PR, Lipp EK, Sherman BH, Bernard SM, Patz JA (2001) Climate variability and change in the United States: potential impacts on water- and foodborne diseases caused by microbiologic agents. Environ Health Perspect 109(suppl2):211–221
Rosenthal J (2009) Climate change and the geographic distribution of infectious diseases. Ecohealth 6:489–495
Semenza JC, Menne B (2009) Climate change and infectious diseases in Europe. Lancet Infect Dis 9:365–375
Socal G, Acri F, Bastianini M, Bernardi Aubry F, Bianchi F, Cassin D, Coppola J, De Lazzari A, Bandelj V, Cossarini G, Solidoro C (2008) Hydrological and biogeochemical features of the Northern Adriatic Sea in the period 2003–2006. Mar Ecol 29:449–468
StatSoft, Inc (2001) STATISTICA for Windows, Computer program manual.
Vezzulli L, Pezzati E, Moreno M, Fabiano M, Pane L, Pruzzo C, VibrioSea Consortium (2009) Benthic ecology of Vibrio spp and pathogenic Vibrio species in a coastal Mediterranean environment (La Spezia Gulf, Italy). Microb Ecol 58:808–818
Acknowledgments
The Vibrio environmental strains analyzed in this study were isolated within the framework of the international research project VibrioSea, which was co-funded by the Centre National d’Etudes Spatiales (CNES), the Institute Pasteur, France, and the Universities of Verona and Genova, Italy.
The authors wish to express their thanks for helpful discussions with the VibrioSea consortium, which included the following institutions: the University of Verona, University of Genova and ISMAR–CNR Venezia (Italy), CNES, MEDIAS-France, CLS, IFREMER and Institute Pasteur Paris (France), and the Institutes Pasteur in Morocco, Algeria, and Tunisia.
Author information
Authors and Affiliations
Corresponding author
Supplemental Materials
Below is the link to the electronic supplementary material.
Fig. S1
Average values (gray filled bars) and standard deviations (vertical lines) of the main oceanographic parameters at each sampling station (surface data). (DOC 142 kb)
Fig. S2
PCA analysis. Top: projection of the oceanographic parameters, Vibrio in water and in plankton on the plane of the first and second factors. Bottom: scores of each variable on the same factors. (DOC 123 kb)
Fig. S3
Cluster analysis performed on oceanographic surface data, Vibrio in water and in plankton, related to the categories “stations” (a) and “seasons” (b). As results, three main groups for each category (GROUP 1, 2, and 3 for stations; GROUP A, B, and C for seasons) were pointed out. (DOC 76 kb)
Rights and permissions
About this article
Cite this article
Caburlotto, G., Bianchi, F., Gennari, M. et al. Integrated Evaluation of Environmental Parameters Influencing Vibrio Occurrence in the Coastal Northern Adriatic Sea (Italy) Facing the Venetian Lagoon. Microb Ecol 63, 20–31 (2012). https://doi.org/10.1007/s00248-011-9920-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00248-011-9920-x