
Abstract
Obesity is a growing health risk with few successful treatment options and fewer still that target both obesity and obesity-associated comorbidities. Despite ongoing scientific efforts, the most effective treatment option to date was not developed from basic research but by surgeons observing outcomes in the clinic. Bariatric surgery is the most successful treatment for significant weight loss, resolution of type 2 diabetes and the prevention of future weight gain. Recent work with animal models has shed considerable light on the molecular underpinnings of the potent effects of these ‘metabolic’ surgical procedures. Here we review data from animal models and how these studies have evolved our understanding of the critical signalling systems that mediate the effects of bariatric surgery. These insights could lead to alternative therapies able to accomplish effects similar to bariatric surgery in a less invasive manner.
Similar content being viewed by others

Introduction
Over 78 million American adults are currently classified as obese [1]. Together with the increase in obesity in the adult population, there has been a disproportionate rise in severe obesity (e.g. patients with a BMI >40 kg/m2) [2]. Unfortunately, individuals with severe obesity are more likely to have diabetes, heart disease and an increased risk of stroke [1], underscoring the need for effective treatment options for both obesity and its associated comorbidities. Indeed, an ideal treatment would lead to weight loss as well as alleviation of the associated comorbidities (e.g. type 2 diabetes) simultaneously.
There are currently a limited number of treatment options available for obesity and only a subset have proved successful in the long term. Behavioural interventions, such as energy-restricted diets and exercise, result in a temporary loss of 5–10% body weight when successful [3], and the few approved obesity medications have a limited range of efficacy [4]. In contrast, some bariatric surgical procedures, as discussed below, have proved to be particularly effective at weight reduction, with patients maintaining ∼30% weight loss for periods of 10 years or more [5, 6].
Despite the repeatedly observed success of bariatric surgical procedures, the underlying mechanisms leading to the metabolic improvements are unclear. Animal models of bariatric surgery allow the unique ability to systemically test mechanistic hypotheses to reveal how and why bariatric surgery leads to significant and sustained weight loss.
‘Metabolic’ surgeries produce weight loss-independent resolution of diabetes
There are multiple types of bariatric surgery; each is similar in that, on average, they produce significant and sustained weight loss. However, not all bariatric surgical procedures result in the resolution of type 2 diabetes independent of weight loss. The ability of a surgical procedure to lead to metabolic benefits independent of weight loss suggests that other effects of the surgery contribute to the observed metabolic improvements [7].
Roux-en-Y gastric bypass and vertical sleeve gastrectomy
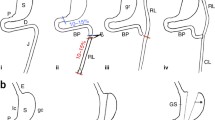
The bariatric surgical procedures that lead to the resolution of type 2 diabetes independent of body weight loss are sometimes referred to as ‘metabolic’ surgeries. (For a review on the various types of bariatric surgeries see [8, 9].) For the purposes of this review, we will focus on the two metabolic procedures that are the most commonly used in the USA and produce sustained, substantial weight loss and resolution of type 2 diabetes: Roux-en-Y gastric bypass (RYGB), and vertical sleeve gastrectomy (VSG) (Fig. 1).

Anatomical arrangement of metabolic surgeries
In RYGB, a small stomach pouch is created and connected to the mid-jejunum. The remainder of the stomach and the proximal intestine remains in the body and is bypassed by nutrient flow. Thus, this procedure results in a decrease in stomach volume as well as a change in nutrient flow. RYGB has been successfully replicated in rodents [10, 11] and, as in humans, leads to significant, sustained weight loss and an improvement in glucose tolerance independent of weight loss [12]. Because micronutrients are predominantly absorbed in the distal gut, the re-routing of nutrients in RYGB leads to micronutrient malabsorption and so individuals who undergo this procedure need to take vitamin supplements [13]. This is replicated in rodent models, where RYGB has been associated with malabsorption of 25-hydroxyvitamin D and calcium [14].
The VSG procedure is technically simpler than RYGB [15, 16]. During VSG, ∼80% of the stomach is excised along the major curvature. Since the only modification is to the size of the stomach, the route of nutrient flow remains the same and the risk of macro- and micro-nutrient malabsorption is minimal. VSG has been successfully modelled in both rats and mice, producing effects similar to those observed in humans, leading to a significant and sustained decrease in body weight as well as an improvement in glucose tolerance independent of weight loss [10, 11, 17, 18].
Weight loss and diabetes resolution in humans
While each procedure is anatomically distinct, on average, both RYGB and VSG patients will experience a reduction in body weight of ∼30% and long-term resolution of type 2 diabetes [19–22]. However, there is discrepancy among human studies indicating less weight loss and a lower frequency of diabetes resolution from VSG in comparison to RYGB [22–25]. Although the majority of evidence points to metabolic surgery producing long-term weight loss [6], this is not always the case. For example, one study reported that ∼20% of RYGB patients regain weight after metabolic surgery [26]. While VSG is still a relatively new procedure, some long-term follow-up studies suggest a smaller weight regain percentage of about 4% [27].
For some patients, diabetes remission lasts as long as 15 years. A recent study by Sjöström et al indicates that this is the case for ∼30% of bariatric surgery patients [28]. It is worth noting that Sjöström et al used pooled results from three different surgeries: RYGB, vertical banded gastroplasty (stomach stapling) and gastric band. Compared with RYGB, the other two surgeries are associated with less weight loss and less diabetes resolution. While this could have contributed to the more conservative rate of resolution compared with short-term outcome studies, a 30% resolution rate still reflects important health and medical cost improvements of surgery relative to other treatment methods.
Weight loss and glucose tolerance in rodents
RYGB and VSG lead to similar reductions in body weight, predominantly from the loss of fat mass, in both rats and mice [10, 29–31]. While few rodent studies have focused on long-term glucose control, numerous studies mirror the glucose improvement observed in humans (as discussed in the following sections). Notably, metabolic surgery is successful in rodent models under a variety of conditions. Significant and sustained body weight reduction is observed in both rats and mice with diet-induced [11, 32, 33] and genetic obesity [34, 35], and in both male and female animals [36]. The tendency for weight regain after metabolic surgery has also been observed in rodent models [37], indicating a conserved physiological adaptation to the surgeries. Furthermore, the ability of the metabolic surgery to consistently lead to a loss in body weight despite alterations in these variables underscores the fact that these models share features with the clinical effects observed in humans.
Research with animal models
The use of animal models of metabolic surgery allows for an additional comparison with pair-fed control groups to assess weight-loss-independent effects of surgery. Pair-fed controls are fed the same amount of energy as consumed by the animals receiving the metabolic surgical procedure, and allows for control of both food intake and weight gain, which is difficult to control for in clinical studies. As discussed in more detail in the following sections, animals pair-fed to so that their energy intake matches that of VSG animals show less of an improvement in glucose homeostasis, despite losing a similar amount of weight [11]. In the case of RYGB, pair-fed groups lose less weight than the RYGB animals [38], and thus weight-matched controls are often used. In agreement with the animal data, resolution of type 2 diabetes in humans has been reported before significant weight loss occurs and sometimes before the patient is discharged from the hospital [39]. Importantly, these data demonstrate that a portion of the glucose improvement from metabolic surgeries is not exclusively due to weight loss but from another weight-independent mechanism.
The mechanism(s) that leads to the success of metabolic surgery is unknown. One possibility is that distinct anatomical differences between the surgeries lead to distinctly different mechanisms of weight loss and diabetes resolution. For example, altered nutrient flow, either by hindgut delivery of nutrients or by duodenal exclusion, has been proposed as a key mechanism driving the success of RYGB [40]. However, recent data have revealed that the gastric emptying rate is extraordinarily rapid after both RYGB and VSG, resulting in similar distal nutrient exposure [41]. Given the similar changes in body weight, glucose tolerance and gastric emptying between VSG and RYGB, an alternative hypothesis focuses on the similarities between the two procedures (e.g. similar changes in gut hormones, bile acids) as a way of elucidating critical pathways involved in weight reduction and glucose regulation. It is important to note that it is as yet unclear what mechanisms are involved, and indeed a mix of both overlapping and non-overlapping mechanisms could be at play.
Metabolic surgery success is not due to restriction or malabsorption
Historically, bariatric surgery was hypothesised to lower body weight primarily via physical manipulation of the surgery to restrict the stomach size and/or re-routing of the intestine to cause a malabsorption of energy. However, this hypothesis has been challenged by multiple datasets from both humans and rodent models.
First, rodents who undergo restrictive procedures are nonetheless capable of increasing their food intake. For example, animals receiving an energy-restricted diet after VSG show a reduction in body weight beyond the weight loss from the surgery [30]. When then allowed free access to food, VSG animals will increase their energy intake to regain weight. Interestingly, the now freely feeding VSG animals only consume enough energy to return to the pre-restriction body weight trajectory and continue to remain lighter than non-operated controls. Importantly, animals that have undergone metabolic surgery can become hyperphagic when physiologically challenged. Another example is seen post surgery in lactating rats. During lactation, the energy required to feed pups via breast milk requires a dramatic increase in food intake. Lactating female rats that have undergone VSG show a pronounced increase in food intake identical to that observed in lactating rats with entirely normal stomachs [36]. These observations highlight two important findings: (1) animals that have undergone metabolic surgery remain on a body weight trajectory below that of a non-operated animal, and (2) post-surgical animals retain the ability to consume more energy, indicating that it is not physical restriction of the stomach that is limiting food intake [30].
There is much debate about whether RYGB leads to malabsorption and, if so, to what extent this accounts for weight loss. Evidence in both humans and rodents indicates that RYGB causes some malabsorption of fat; however, in both species, malabsorption is typically minimal and appears to be unrelated to the degree of weight loss [5, 42]. Moreover, malabsorption is not the primary mechanism of action determining the success of VSG, yet both rodent models and human VSG patients successfully maintain a reduction in body weight and an improvement in glucose tolerance. Thus, malabsorption does not appear to be a critical underlying mechanism involved in the reduction of body weight.
RYGB and VSG: multiple endpoints demonstrate improved metabolic regulation
Metabolic surgeries such as RYGB and VSG affect a number of metabolic endpoints, from food intake to blood glucose regulation. As discussed above, many of these changes, such as diabetes resolution, are independent of body weight loss and collectively demonstrate various aspects of improved metabolic health.
Energy homeostasis
In general, a reduction in body weight can occur through a reduction in food intake, an increase in energy expenditure, or a combination of the two. As an adaptation to dieting and weight loss, there tends to be a reduction in the amount of energy used, thereby preserving body weight and body fat [43, 44]. In both VSG and RYGB a reduction in food intake appears to be a key contributor to the reduction of body weight and body fat [10, 11, 30]. Indeed, after surgery, both humans [45] and rodent models [30] exhibit an early reduction in energy intake. Controlling for energy intake by using pair-fed controls, VSG studies report that pair-fed animals lose a similar amount of weight to those receiving the metabolic surgery [10, 11, 30]. These data indicate that in the case of VSG, food intake appears to be the primary contributor to the reduction in body weight. Supporting this, VSG has been reported to result in no change in energy expenditure [30].
While RYGB also results in a reduction in food intake, some studies have reported that pair-fed animals lose less weight than those that received RYGB [38], indicating that food intake alone may not account for all the weight loss achieved during RYGB. However, data on the impact of RYGB on energy expenditure are variable. This has been more controversial in humans, with some reports finding a decrease in overall energy expenditure [46]. In contrast, energy expenditure has been shown to increase in rodent RYGB studies [38]. This discrepancy may be due to differences between humans and animal models, and also the methods and formulas in which energy expenditure is expressed (for a review, see [8]). Moreover, these results are confounded by how long after the surgery energy expenditure was assessed. Immediately after surgery, both humans and rodents experience a severe reduction in energy intake and prominent negative energy balance, which can independently impact energy expenditure. We conclude that while energy expenditure may play some initial role in weight loss after RYGB, it is not likely to be the sole contributor to weight loss and has even less impact on long-term sustained weight reduction.
Glucose homeostasis
Type 2 diabetic patients are characterised by elevated fasting glucose, impaired glucose tolerance, insulin resistance and impaired beta cell function. After VSG and RYGB, fasting glucose levels are markedly improved [12, 17]. While fasting glucose is known to improve with weight reduction, it has become clear that these changes in fasting glucose after surgery are weight independent, as they often occur before significant weight loss is observed [47] and do not occur to the same degree in pair-fed controls [10].
Both RYGB and VSG are characterised as metabolic surgeries because of their ability to improve glucose tolerance [11, 20]. This effect occurs quickly, before significant weight loss is observed [18, 39, 48]. Data from animal models is particularly important for determining weight-independent effects, since the energy intake of human metabolic surgery patients is severely restricted during the peri-operative period. Energy restriction alone greatly affects fasting glucose levels, as well as insulin sensitivity and beta cell function, in a manner similar to that observed following RYGB [49]. In fact, some studies hypothesise that the energy restriction during the peri-operative period may account for the metabolic effects of VSG and RYGB [49, 50]. However, in rodent models, energy restriction alone cannot explain multiple aspects of glucoregulation observed during VSG or RYGB, including changes in gut hormones, hepatic insulin sensitivity and early phase insulin secretion (see later sections and [7]).
In animal models, RYGB and VSG lead to substantial improvements in glucose tolerance that are greater than the improvement in body weight-matched, pair-fed animals, demonstrating the ability of the metabolic surgery to improve glucose tolerance beyond weight loss alone. This improvement in glucose tolerance may be partly explained by the impact of the surgery on insulin secretion/action. Both RYGB and VSG produce substantial improvements in glucose homeostasis via an early increase in hepatic insulin sensitivity [11, 51]. This increase in hepatic insulin sensitivity is greater than that observed in pair-fed controls, demonstrating the ability of these metabolic surgeries to improve insulin sensitivity through a weight-independent mechanism [11, 18]. After the initial increase in hepatic sensitivity, metabolic surgeries lead to a later increase in insulin sensitivity in skeletal muscle, which is likely to be due to the weight loss [52]. In both humans and animal models, insulin secretion is increased by VSG and RYGB compared with non-operated, weight-matched individuals, suggesting improved beta cell function [53, 54]. In response to a mixed meal, rodent models of metabolic surgery respond similarly to humans, exhibiting an increase in insulin release greater than that observed in non-operated, sham and pair-fed controls [11]. Although controversial [55], intestinal gluconeogenesis has been proposed to increase following a surgery that produces no restriction in stomach size but involves exclusion of upper gastrointestinal (GI) tract nutritional exposure [56]. It has been postulated that the increase in glucose production by the intestine enhances portal vein glucose sensing and contributes to improved glucose homeostasis and can help account for the weight-independent metabolic improvements not observed for other bariatric surgical procedures. While each of the above can contribute to glucose homeostasis after metabolic surgery, it remains to be determined which aspects have the largest impact in the clinical setting.
Gut hormones
An evolving hypothesis for the mechanism of metabolic surgery is rooted in the impact these surgeries have on gut hormone secretion. Indeed, the levels of a number of gut hormones are beneficially changed after VSG and RYGB. Glucagon-like peptide-1 (GLP-1) increases insulin secretion, suppresses glucagon release [57], inhibits hepatic glucose production [58–60] and decreases food intake [61, 62]. Interestingly, postprandial GLP-1 increases tenfold after RYGB and VSG [10], and these increases in GLP-1 are not seen in pair-fed controls. GLP-1 has therefore been implicated as a possible mechanism leading to body weight loss and an improvement in glucose homeostasis after bariatric surgery. Administration of a GLP-1 receptor antagonist blunts the insulin response to a mixed nutrient liquid meal [11]. However, animals lacking the GLP-1 receptor successfully lose weight and exhibit improved glucose tolerance after metabolic surgery [32, 63]. While long-acting GLP-1 agonists are the only type 2 diabetes medications that cause weight loss, the degree of weight loss is considerably less, with estimates of around a third of that achieved by metabolic surgery [64]. Similarly, chronic treatment with pharmacological doses of long-acting GLP-1 agonists alone fail to produce the same magnitude of effects as the degree of weight loss caused by RYGB and VSG and, indeed, these drugs are often used in combination with other pharmacological interventions to treat type 2 diabetes [64–66]. Moreover, GLP-1 antagonists fail to impair the otherwise improved glucose tolerance after VSG in humans [67]. Together, these data demonstrate that GLP-1 alone cannot account for the overall metabolic effect of metabolic surgeries, but we cannot rule out the possibility that high GLP-1 levels in concert with other hormonal changes that occur after surgery might contribute to the potent effects of these procedures on glucose regulation.
Another gut hormone, peptide YY (PYY) decreases food intake, increases insulin sensitivity [68], and is increased after both RYGB and VSG [11, 69, 70]. PYY may play a pivotal role in RYGB-induced weight loss because mice lacking PYY lose less weight initially after RYGB compared with wild-type mice receiving the same procedure [71]. Similarly, an inverse relationship has been established in humans between weight regain after RYGB and plasma PYY levels [37].
Unlike GLP-1 and PYY, the hormone ghrelin elicits food intake [72–74]. Ghrelin is produced in the stomach, duodenum [75] and pancreas [76] and levels are reduced after VSG [18, 77, 78]. Ghrelin response after RYGB is controversial, with some reports indicating a reduction in levels [79], while others show no change [80]. Yet, it appears that the reduction in ghrelin is not essential for the effects of metabolic surgery, at least in an animal model of VSG. Ghrelin-deficient mice respond normally to VSG, including a reduction in body weight and an improvement in glucose tolerance [29]. It is clear that multiple GI hormones change following RYGB and VSG, and it is plausible that each has an important role to play; however, no single gut hormone has emerged as the critical conductor for the metabolic success of VSG or RYGB.
Leptin
Leptin is a satiety hormone released in proportion to adipose tissue. A reduction in body weight results in a reduction in circulating leptin which, in turn, triggers increased hunger, decreased satiety and a decrease in energy expenditure [81]. Thus, it is not surprising that leptin levels decrease after metabolic surgery [30, 48, 70, 82, 83], but, interestingly, the decrease is greater than that observed in weight-matched pair-fed controls. Supporting the view that the enhanced reduction of leptin is weight-loss independent, humans undergoing metabolic surgery also display a reduction in plasma leptin before significant weight loss occurs [83]. Importantly, despite the reduction in leptin levels following metabolic surgery, patients do not display many of the characteristic responses associated with low levels of leptin. For example, patients receiving metabolic surgery actually have decreased hunger and increased satiety [84]. Thus, it has been hypothesised that metabolic surgeries increase leptin sensitivity. However, animal models lacking leptin receptors, which are thus are unable to modulate leptin sensitivity, exhibit normal reductions in body weight in response to VSG [34] and RYGB [35]. Further research is needed to determine why the reduction of leptin in metabolic surgery patients fails to elicit typical responses to promote a positive energy balance.
Lipid handling
Metabolic surgery leads to improved lipid profiles, including cholesterol and plasma triacylglycerols. Some studies indicate that surgery can improve plasma lipid profiles in 70% of patients [85] in a timeframe similar to that for the resolution of type 2 diabetes. On average, both RYGB and VSG improve HDL-cholesterol, and reduce total cholesterol and LDL-cholesterol [85–87]. Moreover, both RYGB and VSG reduce plasma triacylglycerols [85]. Rats also show a reduction, specifically in postprandial plasma triacylglycerols, after VSG owing to intestinal rather than hepatic lipid handling, and this occurs in a weight-loss independent manner [88]. While postprandial hepatic lipid handling may not play a critical role in postprandial plasma triacylglycerols, hepatic triacylglycerol levels themselves are reduced to a greater extent than in pair-fed animals after VSG, again highlighting the weight loss-independent effects of the surgery [89].
Moreover, lipids themselves can exert a powerful stimulus to the gut–brain–liver axis, and it is possible that as a result of the anatomical changes from metabolic surgery, lipids exert beneficial effects on glucose regulation via actions in the intestine. Direct administration of lipids into the upper intestine suppresses hepatic glucose production [90] and this effect can be blocked by inhibiting gut sensory fibres in the brain, or by subdiaphragmatic or hepatic vagotomy. This indicates that lipids activate an intestine–brain–liver neural axis that impacts hepatic glucose production [90] and may be a key contributor to the improved metabolism of patients who undergo metabolic surgery.
Feeding behaviour
Metabolic surgery alters behavioural feeding patterns and food choice. Metabolic surgery patients consistently report a decrease in food intake and meal size [45]. Similarly, animal models of RYGB and VSG result in the consumption of smaller, more frequent, meals [30, 91–93]. There is even evidence that bariatric surgery can improve disrupted feeding patterns in patients with night eating syndrome—a syndrome characterised by >25% of energy intake in the late evening and night [94]. Other aspects of feeding are also altered. In rodent models, metabolic surgery can alter macronutrient preference, decreasing the consumption of fat and increasing the consumption of carbohydrates [92, 93, 95–97].
Novel molecular targets of bariatric surgery
The research highlighted in the previous sections dispels many of the common mechanisms implicated in the success of bariatric surgery, i.e. neither endocrine nor mechanical explanations are sufficient to explain the success of surgery. Thus, current research focuses on potential molecular mechanisms that drive the success of surgery. There are two recent areas of research that have been driving many hypotheses surrounding bariatric surgery. The first is the observation that bile acids are increased after RYGB in humans [98] and similar results have been observed after VSG in rodent models [88]. Bile acids have traditionally been thought to act as lipid detergents but have recently been found to be involved in important signalling mechanisms critical for body weight and glucose regulation [99–101]. In addition, the trillions of species of bacteria that reside in our intestines have been implicated in a more integrated role regulating homeostasis. In this section we describe data demonstrating the link, not only within, but also across, these two systems to the success of bariatric surgery.
Bile acid signalling
Primary bile acids are produced in the liver through the oxidation of cholesterol. Primary bile acids can then be conjugated with a glycine or taurine amino acid to form bile salts that act as a detergent to hydrolyse lipids for absorption. Alternatively, primary bile acids can be secreted into the gut lumen, where dehydroxylation leads to secondary bile acids. Like primary bile acids, secondary bile acids can also be conjugated in the liver to form bile salts. An increase in bile acid circulation directly increases the digestion and absorption of lipids, thus promoting essential effects for lipid trafficking. Moreover, bile acids have been found to act as hormones by acting on both membrane and nuclear receptors within the intestine and liver, with the end result of improved triacylglycerol and glucose metabolism [99–101]. The role of bile acids as a hormone in metabolic signalling makes them particularly interesting to explore for the effects of metabolic surgery.
Interestingly, serum bile acids are more than doubled in patients receiving RYGB compared with gastric banding, a bariatric surgery that results in less weight loss and fewer weight-dependent improvements in comorbidities [102]. This elevation of bile acids during the RYGB metabolic surgery led researchers to test how manipulation of bile acids alone could affect metabolism. In fact, using bile diversion surgery to increase circulating bile acids and bile salts results in improvements similar to those produced by metabolic surgery, including weight loss, improved glucose tolerance and increased postprandial GLP-1 secretion [103]. Similarly, ileum interposition, primarily a rodent surgery model in which a piece of the ileum is excised and sutured within the jejunum, increases bile acids, improves glucose tolerance, and leads to increases in secretion of GLP-1 [104].
Bile acids bind to a nuclear transcription factor called farnesoid X receptor (FXR). Recent studies demonstrate that FXR signalling is a key player in the link between bile acids and metabolism. Indeed, the FXR appears to be crucial for both weight loss and glucose homeostasis from bariatric surgery, as animal models lacking the FXR fail to maintain lower body weights after VSG and instead increase their energy intake to compensate for early post-surgical weight loss [105]. Furthermore, FXR knockout mice have a physically restricted stomach size after VSG similar to wild-type VSG animals; however, FXR knockout mice still fail to maintain a lowered body weight, again pointing to the mechanism of weight loss of bariatric surgery going beyond mere physical restriction of the stomach [105].
Bile acids also bind to TGR5 (also known as G protein-coupled bile acid receptor 1, GPBAR1), a G protein-coupled receptor found in the gut. Activation of TGR5 by bile acid signalling modulates GLP-1 secretion [106], thus making it an interesting target for future studies examining how metabolic surgery, bile acids and glucose homeostasis are linked. However, further studies are necessary to determine the role of TGR5.
Gut microbiota
The gut microbiota refers to the intricate community of bacteria in the gut composed of several different phyla and numerous strains. The composition of these communities has been shown to be sensitive to dietary changes as well as to changes in body weight, and a wide range of data link micobiota composition to obesity and metabolic diseases [107]. Initial studies on obese individuals found significant changes in phyla in terms of the relative prevalence of Firmicutes and Bacteroidetes, with an increase in Firmicutes and a reduction of Bacteroidetes compared with lean counterparts [108]. Similarly, the ob/ob mouse displays elevated Firmicutes and reduced Bacteroidetes compared with lean controls [109]. However, the relative direction of these changes was not supported in later studies [110], which may be partly due to an inconsistency in methodology (see review [107]). For this reason, many animal studies include transplant procedures, which can help elucidate the cause and effect between microbiota and disease states. For example, colonisation of germ-free mice with the microbiota of an obese individual leads to increased body fat compared with colonisation with the microbiota of a lean individual [111].
Metabolic surgery results in a significant change to the gut microbiome [112]. For example, RYGB changes the gut microbiota of an obese individual to a population more similar to that observed in a lean individual [113, 114]. Moreover, germ-free mice receiving the microbiota of an RYGB mouse exhibit significant weight loss and reduced body fat, demonstrating the importance of the microbiota in generating an obese or lean state [115]. Beyond body weight, the microbiota population may also contribute to the resolution of type 2 diabetes. For example, type 2 diabetic patients who receive microbiota from a healthy individual exhibit an improvement in insulin sensitivity [116]. Therefore, a ‘lean’ microbiota population may contribute to the sustained success of metabolic surgery.
Interestingly, there is a correlation between the gut microbiota and bile acids, independent of direct gut manipulation. Non-operated mice with faecal microbiota transplants from mice with RYGB exhibit elevated bile acids and a microbial population similar to ‘lean’ mice [115]. Unfortunately, from this data it is difficult to ascertain whether the microbiota or the bile acids are the first to change. However, data from VSG in the FXR knockout mouse helps to elucidate this relationship [105]. While both FXR knockout and wild-type mice undergo a similar alteration of the gut with VSG, only wild-type mice show a ‘lean’ microbiota phenotype [105]. These data suggest that bile acid-driven FXR signalling may be key to both changes in gut microbiota, as well as the associated weight loss and improvement in glucose metabolism.
Summary and future directions
Obesity affects a large proportion of the population and is associated with substantial health risks, putting a premium on new treatment options. While diet and exercise rarely produce sustained reductions in body weight, metabolic surgery produces larger and more durable improvements in body weight. Importantly, metabolic surgery also leads to an improvement in glucose regulation. Research in pair-fed animal models, in particular, indicates that the improvement in glucose regulation from metabolic surgery is not simply a product of reduced energy intake or weight loss. Despite the original hypothesis behind the design of some human bariatric surgery models, the mechanism of weight loss goes beyond restriction and malabsorption. Metabolic surgery results in marked changes in a variety of gut hormones that can contribute to improvements in glucose regulation and satiety and a reduction in food intake. However, recent data have moved past traditional gut hormones and implicated a role for bile acid signalling and alterations in the gut microbial community as contributors to the potent effects of these surgeries.
Metabolic surgeries were developed and deployed with very little study of preclinical animal models. While it is clear that these animal models will be crucial in helping us understand how these surgeries work, the use of rodent models is not without limitations. For example, the surgeries performed in these animal models can vary between research groups and can be different from what is typically performed in humans because of differences in gut anatomy and the small size of the rodent GI tract. It remains unclear whether these differences in technique impact the metabolic outcome. Nevertheless, rodent models of metabolic surgery have two important implications that will improve our treatment ability in the clinical setting. First, animal models allow for testing of specific hypotheses about the molecular underpinnings of surgical benefits. Identification of the underlying mechanisms is crucial for the development of new therapies that can mimic the benefits of metabolic surgeries while reducing the invasiveness and allowing these solutions to be scaled up for a large target population. Second, animal models allow us to gain an insight into which populations of individuals might most benefit from the currently available procedures. Obesity and diabetes are complex disorders with multiple underlying aetiologies. Metabolic surgeries are likely to be more effective at improving outcomes for some of these underlying aetiologies and not for others. Testing the effects of metabolic surgeries in various animal models allows for a more focused approach, to identify those patients who are most likely to receive a benefit from the procedures, thereby reducing the number of patients who get little or no benefit.
Abbreviations
- FXR:
-
Farnesoid X receptor
- GI:
-
Gastrointestinal
- GLP-1:
-
Glucagon-like peptide-1
- PYY:
-
Peptide YY
- RYGB:
-
Roux-en-Y gastric bypass
- VSG:
-
Vertical sleeve gastrectomy
References
Shields M, Carroll MD, Ogden CL (2011) Adult obesity prevalence in Canada and the United States. NCHS Data Brief: 1–8
Sturm R (2007) Increases in morbid obesity in the USA: 2000–2005. Public Health 121:492–496
Kraschnewski JL, Boan J, Esposito J et al (2010) Long-term weight loss maintenance in the United States. Int J Obes (Lond) 34:1644–1654
Bray GA (2008) Lifestyle and pharmacological approaches to weight loss: efficacy and safety. J Clin Endocrinol Metab 93:S81–S88
Sandoval D (2011) Bariatric surgeries: beyond restriction and malabsorption. Int J Obes (Lond) 35(Suppl 3):S45–S49
Pories WJ, Swanson MS, MacDonald KG et al (1995) Who would have thought it? An operation proves to be the most effective therapy for adult-onset diabetes mellitus. Ann Surg 222:339–350, discussion 350–332
Cohen RV, Rubino F, Schiavon C, Cummings DE (2012) Diabetes remission without weight loss after duodenal bypass surgery. Surg Obes Relat Dis 8:e66–e68
Stefater MA, Wilson-Perez HE, Chambers AP, Sandoval DA, Seeley RJ (2012) All bariatric surgeries are not created equal: insights from mechanistic comparisons. Endocr Rev 33:595–622
Moshiri M, Osman S, Robinson TJ, Khandelwal S, Bhargava P, Rohrmann CA (2013) Evolution of bariatric surgery: a historical perspective. AJR Am J Roentgenol 201:W40–W48
Chambers AP, Stefater MA, Wilson-Perez HE et al (2011) Similar effects of roux-en-Y gastric bypass and vertical sleeve gastrectomy on glucose regulation in rats. Physiol Behav 105:120–123
Chambers AP, Jessen L, Ryan KK et al (2011) Weight-independent changes in blood glucose homeostasis after gastric bypass or vertical sleeve gastrectomy in rats. Gastroenterology 141:950–958
Wickremesekera K, Miller G, Naotunne TD, Knowles G, Stubbs RS (2005) Loss of insulin resistance after Roux-en-Y gastric bypass surgery: a time course study. Obes Surg 15:474–481
Bal BS, Finelli FC, Koch TR (2011) Origins of and recognition of micronutrient deficiencies after gastric bypass surgery. Curr Diabetes Rep 11:136–141
Stemmer K, Bielohuby M, Grayson BE et al (2013) Roux-en-Y gastric bypass surgery but not vertical sleeve gastrectomy decreases bone mass in male rats. Endocrinology 154:2015–2024
Santry HP, Gillen DL, Lauderdale DS (2005) Trends in bariatric surgical procedures. JAMA 294:1909–1917
Abu-Jaish W, Rosenthal RJ (2010) Sleeve gastrectomy: a new surgical approach for morbid obesity. Expert Rev Gastroenterol Hepatol 4:101–119
Rizzello M, Abbatini F, Casella G et al (2010) Early postoperative insulin-resistance changes after sleeve gastrectomy. Obes Surg 20:50–55
Basso N, Capoccia D, Rizzello M et al (2011) First-phase insulin secretion, insulin sensitivity, ghrelin, GLP-1, and PYY changes 72 h after sleeve gastrectomy in obese diabetic patients: the gastric hypothesis. Surg Endosc 25:3540–3550
Nannipieri M, Baldi S, Mari A et al (2013) Roux-en-Y gastric bypass and sleeve gastrectomy: mechanisms of diabetes remission and role of gut hormones. J Clin Endocrinol Metab 98:4391–4399
Buchwald H, Estok R, Fahrbach K et al (2009) Weight and type 2 diabetes after bariatric surgery: systematic review and meta-analysis. Am J Med 122:248–256
Chang SH, Stoll CR, Song J, Varela JE, Eagon CJ, Colditz GA (2014) The effectiveness and risks of bariatric surgery: an updated systematic review and meta-analysis, 2003–2012. JAMA Surg 149:275–287
Trastulli S, Desiderio J, Guarino S et al (2013) Laparoscopic sleeve gastrectomy compared with other bariatric surgical procedures: a systematic review of randomized trials. Surg Obes Relat Dis 9:816–829
Lee WJ, Chong K, Ser KH et al (2011) Gastric bypass vs sleeve gastrectomy for type 2 diabetes mellitus: a randomized controlled trial. Arch Surg 146:143–148
Schauer PR, Bhatt DL, Kirwan JP et al (2014) Bariatric surgery versus intensive medical therapy for diabetes – 3-year outcomes. N Engl J Med 370:2002–2013
Pham S, Gancel A, Scotte M et al (2014) Comparison of the effectiveness of four bariatric surgery procedures in obese patients with type 2 diabetes: a retrospective study. J Obes 2014:638203
Guijarro A, Osei-Hyiaman D, Harvey-White J et al (2008) Sustained weight loss after Roux-en-Y gastric bypass is characterized by down regulation of endocannabinoids and mitochondrial function. Ann Surg 247:779–790
Abd Ellatif ME, Abdallah E, Askar W et al (2014) Long term predictors of success after laparoscopic sleeve gastrectomy. Int J Surg 12:504–508
Sjöström L, Peltonen M, Jacobson P et al (2014) Association of bariatric surgery with long-term remission of type 2 diabetes and with microvascular and macrovascular complications. JAMA 311:2297–2304
Chambers AP, Kirchner H, Wilson-Perez HE et al (2013) The effects of vertical sleeve gastrectomy in rodents are ghrelin independent. Gastroenterology 144:50–52
Stefater MA, Perez-Tilve D, Chambers AP et al (2010) Sleeve gastrectomy induces loss of weight and fat mass in obese rats, but does not affect leptin sensitivity. Gastroenterology 138:2426–2436
Carey DG, Pliego GJ, Raymond RL (2006) Body composition and metabolic changes following bariatric surgery: effects on fat mass, lean mass and basal metabolic rate: six months to one-year follow-up. Obes Surg 16:1602–1608
Wilson-Perez HE, Chambers AP, Ryan KK et al (2013) Vertical sleeve gastrectomy is effective in two genetic mouse models of glucagon-like peptide 1 receptor deficiency. Diabetes 62:2380–2385
Nestoridi E, Kvas S, Kucharczyk J, Stylopoulos N (2012) Resting energy expenditure and energetic cost of feeding are augmented after Roux-en-Y gastric bypass in obese mice. Endocrinology 153:2234–2244
Lopez PP, Nicholson SE, Burkhardt GE, Johnson RA, Johnson FK (2009) Development of a sleeve gastrectomy weight loss model in obese Zucker rats. J Surg Res 157:243–250
Liu B, Kuang L, Liu J (2014) Bariatric surgery relieves type 2 diabetes and modulates inflammatory factors and coronary endothelium eNOS/iNOS expression in db/db mice. Can J Physiol Pharmacol 92:70–77
Grayson BE, Schneider KM, Woods SC, Seeley RJ (2013) Improved rodent maternal metabolism but reduced intrauterine growth after vertical sleeve gastrectomy. Sci Transl Med 5:199ra112
Meguid MM, Glade MJ, Middleton FA (2008) Weight regain after Roux-en-Y: a significant 20% complication related to PYY. Nutrition 24:832–842
Stylopoulos N, Hoppin AG, Kaplan LM (2009) Roux-en-Y gastric bypass enhances energy expenditure and extends lifespan in diet-induced obese rats. Obesity 17:1839–1847
Shah SS, Todkar JS, Shah PS, Cummings DE (2010) Diabetes remission and reduced cardiovascular risk after gastric bypass in Asian Indians with body mass index <35 kg/m2. Surg Obes Relat Dis 6:332–338
Cummings DE, Overduin J, Foster-Schubert KE (2004) Gastric bypass for obesity: mechanisms of weight loss and diabetes resolution. J Clin Endocrinol Metab 89:2608–2615
Chambers AP, Smith EP, Begg DP et al (2014) Regulation of gastric emptying rate and its role in nutrient-induced GLP-1 secretion in rats after vertical sleeve gastrectomy. Am J Physiol Endocrinol Metab 306:E424–E432
Odstrcil EA, Martinez JG, Santa Ana CA et al (2010) The contribution of malabsorption to the reduction in net energy absorption after long-limb Roux-en-Y gastric bypass. Am J Clin Nutr 92:704–713
Schwartz A, Doucet E (2010) Relative changes in resting energy expenditure during weight loss: a systematic review. Obes Rev Off J Int Assoc Study Obes 11:531–547
Corbett SW, Stern JS, Keesey RE (1986) Energy expenditure in rats with diet-induced obesity. Am J Clin Nutr 44:173–180
Warde-Kamar J, Rogers M, Flancbaum L, Laferrere B (2004) Calorie intake and meal patterns up to 4 years after Roux-en-Y gastric bypass surgery. Obes Surg 14:1070–1079
Carrasco F, Papapietro K, Csendes A et al (2007) Changes in resting energy expenditure and body composition after weight loss following Roux-en-Y gastric bypass. Obes Surg 17:608–616
Rubino F, R'Bibo SL, del Genio F, Mazumdar M, McGraw TE (2010) Metabolic surgery: the role of the gastrointestinal tract in diabetes mellitus. Nat Rev Endocrinol 6:102–109
Rubino F, Gagner M, Gentileschi P et al (2004) The early effect of the Roux-en-Y gastric bypass on hormones involved in body weight regulation and glucose metabolism. Ann Surg 240:236–242
Jackness C, Karmally W, Febres G et al (2013) Very low-calorie diet mimics the early beneficial effect of Roux-en-Y gastric bypass on insulin sensitivity and beta-cell function in type 2 diabetic patients. Diabetes 62:3027–3032
Bradley D, Conte C, Mittendorfer B et al (2012) Gastric bypass and banding equally improve insulin sensitivity and beta cell function. J Clin Invest 122:4667–4674
Abbatini F, Rizzello M, Casella G et al (2010) Long-term effects of laparoscopic sleeve gastrectomy, gastric bypass, and adjustable gastric banding on type 2 diabetes. Surg Endosc 24:1005–1010
Magkos F, Bradley D, Schweitzer GG et al (2013) Effect of Roux-en-Y gastric bypass and laparoscopic adjustable gastric banding on branched-chain amino acid metabolism. Diabetes 62:2757–2761
Nannipieri M, Mari A, Anselmino M et al (2011) The role of beta-cell function and insulin sensitivity in the remission of type 2 diabetes after gastric bypass surgery. J Clin Endocrinol Metab 96:E1372–E1379
Romero F, Nicolau J, Flores L et al (2012) Comparable early changes in gastrointestinal hormones after sleeve gastrectomy and Roux-En-Y gastric bypass surgery for morbidly obese type 2 diabetic subjects. Surg Endosc 26:2231–2239
Previs SF, Brunengraber DZ, Brunengraber H (2009) Is there glucose production outside of the liver and kidney? Annu Rev Nutr 29:43–57
Troy S, Soty M, Ribeiro L et al (2008) Intestinal gluconeogenesis is a key factor for early metabolic changes after gastric bypass but not after gastric lap-band in mice. Cell Metab 8:201–211
Meier JJ (2012) GLP-1 receptor agonists for individualized treatment of type 2 diabetes mellitus. Nat Rev Endocrinol 8:728–742
Hvidberg A, Nielsen MT, Hilsted J, Orskov C, Holst JJ (1994) Effect of glucagon-like peptide-1 (proglucagon 78-107amide) on hepatic glucose production in healthy man. Metab Clin Exp 43:104–108
Sandoval DA, Bagnol D, Woods SC, D’Alessio DA, Seeley RJ (2008) Arcuate glucagon-like peptide 1 receptors regulate glucose homeostasis but not food intake. Diabetes 57:2046–2054
Ayala JE, Bracy DP, James FD, Julien BM, Wasserman DH, Drucker DJ (2009) The glucagon-like peptide-1 receptor regulates endogenous glucose production and muscle glucose uptake independent of its incretin action. Endocrinology 150:1155–1164
Turton MD, O’Shea D, Gunn I et al (1996) A role for glucagon-like peptide-1 in the central regulation of feeding. Nature 379:69–72
Gutzwiller JP, Degen L, Heuss L, Beglinger C (2004) Glucagon-like peptide 1 (GLP-1) and eating. Physiol Behav 82:17–19
Mokadem M, Zechner JF, Margolskee RF, Drucker DJ, Aguirre V (2014) Effects of Roux-en-Y gastric bypass on energy and glucose homeostasis are preserved in two mouse models of functional glucagon-like peptide-1 deficiency. Mol Metab 3:191–201
Bode B (2011) Liraglutide: a review of the first once-daily GLP-1 receptor agonist. Am J Manage Care 17:S59–S70
Pratley R, Nauck M, Bailey T et al (2011) One year of liraglutide treatment offers sustained and more effective glycaemic control and weight reduction compared with sitagliptin, both in combination with metformin, in patients with type 2 diabetes: a randomised, parallel-group, open-label trial. Int J Clin Pract 65:397–407
Fujishima Y, Maeda N, Inoue K et al (2012) Efficacy of liraglutide, a glucagon-like peptide-1 (GLP-1) analogue, on body weight, eating behavior, and glycemic control, in Japanese obese type 2 diabetes. Cardiovasc Diabetol 11:107
Jiménez A, Mari A, Casamitjana R, Lacy A, Ferrannini E, Vidal J (2014) GLP-1 and glucose tolerance after sleeve gastrectomy in morbidly obese subjects with type 2 diabetes. Diabetes 63:3372–3377
Vrang N, Madsen AN, Tang-Christensen M, Hansen G, Larsen PJ (2006) PYY(3-36) reduces food intake and body weight and improves insulin sensitivity in rodent models of diet-induced obesity. Am J Physiol Regul Integr Comp Physiol 291:R367–R375
Peterli R, Wolnerhanssen B, Peters T et al (2009) Improvement in glucose metabolism after bariatric surgery: comparison of laparoscopic Roux-en-Y gastric bypass and laparoscopic sleeve gastrectomy: a prospective randomized trial. Ann Surg 250:234–241
Shin AC, Zheng H, Townsend RL, Sigalet DL, Berthoud HR (2010) Meal-induced hormone responses in a rat model of Roux-en-Y gastric bypass surgery. Endocrinology 151:1588–1597
Chandarana K, Gelegen C, Karra E et al (2011) Diet and gastrointestinal bypass-induced weight loss: the roles of ghrelin and peptide YY. Diabetes 60:810–818
Wren AM, Small CJ, Ward HL et al (2000) The novel hypothalamic peptide ghrelin stimulates food intake and growth hormone secretion. Endocrinology 141:4325–4328
Wren AM, Seal LJ, Cohen MA et al (2001) Ghrelin enhances appetite and increases food intake in humans. J Clin Endocrinol Metab 86:5992
Tschop M, Smiley DL, Heiman ML (2000) Ghrelin induces adiposity in rodents. Nature 407:908–913
Kojima M, Hosoda H, Date Y, Nakazato M, Matsuo H, Kangawa K (1999) Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature 402:656–660
Lai KC, Cheng CH, Leung PS (2007) The ghrelin system in acinar cells: localization, expression, and regulation in the exocrine pancreas. Pancreas 35:e1–e8
Bohdjalian A, Langer FB, Shakeri-Leidenmuhler S et al (2010) Sleeve gastrectomy as sole and definitive bariatric procedure: 5-year results for weight loss and ghrelin. Obes Surg 20:535–540
Wang Y, Liu J (2009) Plasma ghrelin modulation in gastric band operation and sleeve gastrectomy. Obes Surg 19:357–362
Cummings DE, Weigle DS, Frayo RS et al (2002) Plasma ghrelin levels after diet-induced weight loss or gastric bypass surgery. N Engl J Med 346:1623–1630
Tymitz K, Engel A, McDonough S, Hendy MP, Kerlakian G (2011) Changes in ghrelin levels following bariatric surgery: review of the literature. Obes Surg 21:125–130
Leibel RL, Rosenbaum M, Hirsch J (1995) Changes in energy expenditure resulting from altered body weight. N Engl J Med 332:621–628
Korner J, Bessler M, Cirilo LJ et al (2005) Effects of Roux-en-Y gastric bypass surgery on fasting and postprandial concentrations of plasma ghrelin, peptide YY, and insulin. J Clin Endocrinol Metab 90:359–365
Woelnerhanssen B, Peterli R, Steinert RE, Peters T, Borbely Y, Beglinger C (2011) Effects of postbariatric surgery weight loss on adipokines and metabolic parameters: comparison of laparoscopic Roux-en-Y gastric bypass and laparoscopic sleeve gastrectomy—a prospective randomized trial. Surg Obes Relat Dis 7:561–568
Schultes B, Ernst B, Wilms B, Thurnheer M, Hallschmid M (2010) Hedonic hunger is increased in severely obese patients and is reduced after gastric bypass surgery. Am J Clin Nutr 92:277–283
Buchwald H, Avidor Y, Braunwald E et al (2004) Bariatric surgery: a systematic review and meta-analysis. JAMA 292:1724–1737
Hady HR, Dadan J, Luba M (2012) The influence of laparoscopic sleeve gastrectomy on metabolic syndrome parameters in obese patients in own material. Obes Surg 22:13–22
Nguyen NT, Varela E, Sabio A, Tran CL, Stamos M, Wilson SE (2006) Resolution of hyperlipidemia after laparoscopic Roux-en-Y gastric bypass. J Am Coll Surg 203:24–29
Stefater MA, Sandoval DA, Chambers AP et al (2011) Sleeve gastrectomy in rats improves postprandial lipid clearance by reducing intestinal triglyceride secretion. Gastroenterology 141(939–949):e931–e934
Myronovych A, Kirby M, Ryan KK et al (2014) Vertical sleeve gastrectomy reduces hepatic steatosis while increasing serum bile acids in a weight-loss-independent manner. Obesity 22:390–400
Wang PY, Caspi L, Lam CK et al (2008) Upper intestinal lipids trigger a gut-brain-liver axis to regulate glucose production. Nature 452:1012–1016
Bueter M, Lowenstein C, Olbers T et al (2010) Gastric bypass increases energy expenditure in rats. Gastroenterology 138:1845–1853
Chambers AP, Wilson-Perez HE, McGrath S et al (2012) Effect of vertical sleeve gastrectomy on food selection and satiation in rats. Am J Physiol Endocrinol Metab 303:E1076–E1084
Zheng H, Shin AC, Lenard NR et al (2009) Meal patterns, satiety, and food choice in a rat model of Roux-en-Y gastric bypass surgery. Am J Physiol Regul Integr Comp Physiol 297:R1273–R1282
Colles SL, Dixon JB (2006) Night eating syndrome: impact on bariatric surgery. Obes Surg 16:811–820
Wilson-Perez HE, Chambers AP, Sandoval DA et al (2013) The effect of vertical sleeve gastrectomy on food choice in rats. Int J Obes (Lond) 37:288–295
le Roux CW, Bueter M, Theis N et al (2011) Gastric bypass reduces fat intake and preference. Am J Physiol Regul Integr Comp Physiol 301:R1057–R1066
Mul JD, Begg DP, Alsters SI et al (2012) Effect of vertical sleeve gastrectomy in melanocortin receptor 4-deficient rats. Am J Physiol Endocrinol Metab 303:E103–E110
Patti ME, Houten SM, Bianco AC et al (2009) Serum bile acids are higher in humans with prior gastric bypass: potential contribution to improved glucose and lipid metabolism. Obesity 17:1671–1677
Parks DJ, Blanchard SG, Bledsoe RK et al (1999) Bile acids: natural ligands for an orphan nuclear receptor. Science 284:1365–1368
Kohli R, Kirby M, Setchell KD et al (2010) Intestinal adaptation after ileal interposition surgery increases bile acid recycling and protects against obesity-related comorbidities. Am J Physiol Gastrointest Liver Physiol 299:G652–G660
Cummings BP, Strader AD, Stanhope KL et al (2010) Ileal interposition surgery improves glucose and lipid metabolism and delays diabetes onset in the UCD-T2DM rat. Gastroenterology 138:2437–2446
Kohli R, Bradley D, Setchell KD, Eagon JC, Abumrad N, Klein S (2013) Weight loss induced by Roux-en-Y gastric bypass but not laparoscopic adjustable gastric banding increases circulating bile acids. J Clin Endocrinol Metab 98:E708–E712
Kohli R, Setchell KD, Kirby M et al (2013) A surgical model in male obese rats uncovers protective effects of bile acids post-bariatric surgery. Endocrinology 154:2341–2351
Strader AD, Vahl TP, Jandacek RJ, Woods SC, D’Alessio DA, Seeley RJ (2005) Weight loss through ileal transposition is accompanied by increased ileal hormone secretion and synthesis in rats. Am J Physiol Endocrinol Metab 288:E447–E453
Ryan KK, Tremaroli V, Clemmensen C et al (2014) FXR is a molecular target for the effects of vertical sleeve gastrectomy. Nature 509:183–188
Thomas C, Gioiello A, Noriega L et al (2009) TGR5-mediated bile acid sensing controls glucose homeostasis. Cell Metab 10:167–177
Karlsson F, Tremaroli V, Nielsen J, Backhed F (2013) Assessing the human gut microbiota in metabolic diseases. Diabetes 62:3341–3349
Ley RE, Turnbaugh PJ, Klein S, Gordon JI (2006) Microbial ecology: human gut microbes associated with obesity. Nature 444:1022–1023
Ley RE, Backhed F, Turnbaugh P, Lozupone CA, Knight RD, Gordon JI (2005) Obesity alters gut microbial ecology. Proc Natl Acad Sci U S A 102:11070–11075
Schwiertz A, Taras D, Schafer K et al (2010) Microbiota and SCFA in lean and overweight healthy subjects. Obesity 18:190–195
Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI (2006) An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 444:1027–1031
Sweeney TE, Morton JM (2013) The human gut microbiome: a review of the effect of obesity and surgically induced weight loss. JAMA Surg 148:563–569
Zhang H, DiBaise JK, Zuccolo A et al (2009) Human gut microbiota in obesity and after gastric bypass. Proc Natl Acad Sci U S A 106:2365–2370
Li JV, Ashrafian H, Bueter M et al (2011) Metabolic surgery profoundly influences gut microbial-host metabolic cross-talk. Gut 60:1214–1223
Liou AP, Paziuk M, Luevano JM Jr, Machineni S, Turnbaugh PJ, Kaplan LM (2013) Conserved shifts in the gut microbiota due to gastric bypass reduce host weight and adiposity. Sci Transl Med 5:178ra141
Vrieze A, Van Nood E, Holleman F et al (2012) Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology 143:913–916
Duality of interest
DMA has no duality of interest associated with this manuscript. DAS receives funding from Ethicon Endo-Surgery, Novo Nordisk and Boehringer Ingelheim International. RJS receives funding from Ethicon Endo-Surgery, Ablaris Therapeutics, Inc., Novo Nordisk, Novartis, Angiochem, Eisai, Forest Pharmaceuticals, Givaudan, Zealand Pharmaceuticals and Boehringer Ingelheim International.
Contribution statement
All authors were responsible for the conception and design of the manuscript, drafting the article and revising it critically for important intellectual content. All authors approved the version to be published.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Arble, D.M., Sandoval, D.A. & Seeley, R.J. Mechanisms underlying weight loss and metabolic improvements in rodent models of bariatric surgery. Diabetologia 58, 211–220 (2015). https://doi.org/10.1007/s00125-014-3433-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00125-014-3433-3