
Abstract
Aims/hypothesis
The blood perfusion of individual pancreatic islets is highly variable, with a subgroup of islets having high perfusion and blood vessels responsive to further blood flow increase induced by glucose. This study tested the hypothesis that there is heterogeneity between islets with regard to beta cell proliferation, function and gene expression based on differences in their blood perfusion.
Methods
Fluorescent microspheres were injected into the ascending aorta, and then microsphere-containing and non-microsphere-containing pancreatic islets were isolated for investigation. By this procedure, the 5% of islets with the greatest blood perfusion were identified for study. Islet endothelial cells were isolated separately to investigate the role of improved vascular support in the observed differences.
Results
The vascular network was found to be more dense and tortuous in microsphere-containing than other islets. The most highly blood-perfused islets also had a higher rate of beta cell proliferation, superior beta cell function and a markedly different gene expression from other islets. Cultured islets exposed to islet endothelial cell products had a similarly increased beta cell proliferation rate, yet significantly fewer changes in gene expression than observed in the most highly blood-perfused islets.
Conclusions/interpretation
A novel heterogeneity between islets was observed, with superior beta cell proliferation, function and gene expression in a subpopulation of islets identified by high blood perfusion. In contrast with a previously described population of low-oxygenated, sleeping islets, which are recruited into functionality when needed, the presently described heterogeneity is shown to remain in vitro after islet isolation.
Similar content being viewed by others

Introduction
Pancreatic islets are richly vascularised [1, 2]. A pivotal role of endothelial cell signalling in early embryonic development of beta cells has been described [3], and endothelial cells have also been shown to be important for postnatal beta cell function and growth. Vascular basement proteins such as laminins produced by the endothelial cells promote insulin gene expression and beta cell proliferation and enhance glucose-stimulated insulin release [4, 5]. We also recently showed that endothelial-derived thrombospondin-1 is important for beta cell differentiation and function [6]. Moreover, hepatocyte growth factor production can be induced by vascular endothelial growth factor-A (VEGF-A) and insulin in islet endothelial cells, further contributing to beta cell proliferation during increased functional demand [7]. These findings raise the possibility of potential differences in endocrine function between subpopulations of pancreatic islets based on their vascular support.
Interestingly, the degree of blood perfusion in different islets seems to be highly variable. Although islets constitute only 1–2% of the pancreatic volume, they receive 5–10% of the whole pancreatic blood flow [8, 9]. However, when islet blood flow is measured by microsphere techniques, a small fraction of islets may contain several microspheres, whereas ~90% of the islets are totally devoid of microspheres, i.e. their blood perfusion is below the detection limit [10]. Repeated microsphere injections show that, primarily, islets containing microspheres after the first injection also contain microspheres after repeated microsphere injections, indicating that the heterogeneity in blood perfusion remains constant over time [11].
Islet blood perfusion is normally tightly regulated at the arteriolar level by nervous, endocrine and metabolic mechanisms [12, 13]. A dose- and time-dependent preferential increase in blood flow is induced by glucose [13, 14]. This suggests that appropriate blood perfusion is essential for islet metabolism and insulin secretion. However, administration of glucose only seems to increase the blood perfusion of those islets that have the highest blood perfusion [10, 11]. It is not known whether any functional differences exist between these highly blood-perfused islets and other islets.
The present study tested the hypothesis that highly blood-perfused islets constitute a subgroup with better vascular support of beta cell function and growth. We used fluorescent microspheres to identify highly blood-perfused islets, followed by separate isolation of microsphere-containing and non-microsphere-containing islets for investigation.
Methods
Experimental animals
Male Wistar–Furth rats 10–12 weeks of age were purchased from Scanbur, Sollentuna, Sweden. All experimental procedures were approved by the Animal Ethics Committee of Uppsala University, and followed Principles of Laboratory Animal Care (NIH publication no. 85–23, revised 1985).
Chemicals
All chemicals were purchased from Sigma-Aldrich (St Louis, MO, USA) unless otherwise mentioned.
Islet blood flow
Blood flow measurements in Wistar–Furth rats were performed 10 min after injection of 1 ml 30% (wt/vol.) d-glucose as previously described [15], except for the use of green fluorescent microspheres (10 μm; E-Z Trac; IMT, Irvine, CA, USA).
Islet isolation
Islets were isolated by collagenase digestion [16] and hand picked. They were then dichotomously sorted under a fluorescence microscope by the presence or absence of microspheres. Each experiment was in most cases based on material from one animal. However, the number of islets containing microspheres was too low in some animals to perform a particular assay, and in these cases both microsphere-containing islets and control islets were pooled from two or more animals. The number of control islets from each animal in the experiment was therefore proportional to the fraction of microsphere-containing islets from each animal.
Islet size
Microsphere-containing and non-microsphere-containing islets were harvested separately and homogenised in groups of 10. Aliquots of the aqueous homogenate were used for DNA measurements by fluorophotometry (PicoGreen dsDNA Quantitation kit; Molecular Probes, Eugene, OR, USA).
Endocrine cell composition
Islets containing or not containing microspheres were separately fixed in paraformaldehyde, embedded in paraffin, cut into 5 μm-thick sections and stained with primary antibodies against insulin (1:400 dilution; polyclonal guinea pig serum; Fitzgerald, Concord, MA, USA) or glucagon (1:1,000 dilution; mouse monoclonal; Abcam, Cambridge, UK). Sections were counterstained with haematoxylin. They were then examined by a point-counting method [17], where the number of intersections overlapping insulin- and glucagon-positive cells were counted by light microscopy. The relative insulin- and glucagon-positive cell areas in islets containing or not containing microspheres from each animal were calculated.
Islet vascular density
To visualise the islet blood vessels with a multiphoton microscope, we used isolectin IB4 from Bandeiraea simplicifolia conjugated to Alexa Fluor 568 dye (Molecular Probes). The isolectin was administered 5 min before glucose and subsequent fluorescent microsphere injections. Islets containing or not containing microspheres were separately fixed in paraformaldehyde, embedded in 1% (wt/vol.) agarose and then scanned with a two-photon microscope (Zeiss LSM710 NLO). The objective used for imaging was the Zeiss W Plan-Apochromat 20×/1.0 DICM27 75 mm. Isolectin was excited with a MaiTai laser at 780 nm and 7.5% using MBS 760. Emission light was collected by NDD-R using long-pass filter LP555 in combination with band-pass filter BP565-610. The scanned images were analysed with IMARIS7.1.1 (Bitplane Scientific Software, South Windsor, CT, USA).
Isolation and culture of islet endothelial cells
Outgrowth of mesenchymal cells from islets was stimulated as described previously [7]. Cells were detached with 0.25% (wt/vol.) trypsin/EDTA solution at 37°C. Endothelial cells were extracted from the resultant cell suspension by a Dynabead method [18] and separated from contaminating cells by the use of Bandeiraea simplicifolia-coated Dynabeads, achieving a purity of >90% [7, 19]. Detailed characterisation of the endothelial cells has been described previously [5, 19].
Generation of islet endothelial cell culture medium
Dynabead-purified endothelial cells were cultured for 2 days in endothelial cell culture medium (CM) (RPMI 1640 medium with 11.1 mmol/l glucose, and supplemented with 20% [vol./vol.] fetal calf serum, 100 μg endothelial cell growth supplement and 2 mmol/l l-glutamine). The wells were washed with standard islet CM (RPMI 1640 medium with 11.1 mmol/l glucose, and supplemented with 10% [vol./vol.] fetal calf serum and 2 mmol/l l-glutamine) to remove all endothelial cell CM, and fresh CM was then added to the wells. The medium was collected 24 h later and centrifuged for 2 min at 600 g to remove cells. The endothelium-conditioned CM (ECCM) was then stored at −70°C.
Beta cell proliferation
Freshly isolated islets containing or not containing microspheres, fixed in paraformaldehyde, embedded in paraffin, sectioned and stained for insulin as described above, were also stained for the proliferation-associated protein, Ki67 (1:200 dilution; mouse monoclonal; Dako, Glostrup, Denmark), and counterstained with haematoxylin. Other islets were isolated from non-microsphere-injected rats and cultured for 7 days, to deplete endogenous blood vessels [20], before exposure for 24 h to ECCM or CM. Ki67-labelled beta cells were counted and expressed as a percentage of the total number of investigated beta cells. In the same sections, all other islet cells (cells negative for insulin) were also investigated for the percentage positive for Ki-67. From each group of islets (microsphere- or non-microsphere-containing, ECCM- or CM-exposed), a mean number of ~2,600 beta cells and ~1,800 other islet cells was investigated per animal.
Glucose-stimulated insulin release
Groups of 10 freshly isolated islets divided into those containing or not containing microspheres were investigated for capacity to release insulin at low (1.67 mmol/l) and high (16.7 mmol/l) glucose concentrations [5]. Insulin content of incubation media and homogenates was determined with a rat insulin ELISA kit (Mercodia, Uppsala, Sweden).
(Pro)insulin biosynthesis
Duplicate groups of 10 freshly isolated islets containing microspheres were compared with those devoid of microspheres with regard to (pro)insulin and total protein biosynthesis. Assays were performed as described previously [16].
Glucose oxidation
Duplicate groups of 10 freshly isolated islets, divided into those containing or not containing microspheres, were analysed for glucose oxidation rates [21].
Islet gene expression
Total RNA was isolated from freshly isolated microsphere-containing islets and non-microsphere-containing islets with RNeasy Micro kit (Qiagen, Hilden, Germany). Quadruple samples comprising 10–12 islets from each of these two groups in Wistar–Furth rats were analysed for gene expression, with mean values for microsphere-containing and non-microsphere-containing islets in each experiment calculated and used in the statistical analysis. One or two animals were needed to obtain enough microsphere-containing islets for an experiment. If islets were obtained from two animals, the islets from each animal were randomly assorted to the four groups of 10–12 islets in an experiment. We then performed the number of experiments stated in the Results section and Table 1 with different quadruple samples. Similarly, total RNA was isolated from islets cultured for 7 days, followed by exposure for 24 h to ECCM or CM. All of the samples were treated with RNase-free DNase (Qiagen). Conversion of RNA into complementary DNA was performed with the QuantiTect Reverse Transcription Kit (Qiagen). Custom-made TaqMan Array Micro Fluidic Cards with Applied Biosystems 7900HT Fast Real Time PCR system (Foster City, CA, USA) were used to profile gene expression through the comparative Ct method of relative quantification, with investigated genes (see electronic supplementary material [ESM] Table 1) analysed according to the manufacturer’s instructions and 18S rRNA as housekeeping gene.
Statistical analysis
All values are given as means ± SEM. Student’s unpaired or paired t tests were used for comparisons of two groups of parametric data, whereas the Mann–Whitney rank sum test or Wilcoxon signed rank test was used for non-parametric data.
Results
Investigated animals (n = 21) had a mean arterial blood pressure of 111 ± 3 mmHg and blood glucose concentration of 15.3 ± 1.1 mmol/l at the time of microsphere injection 10 min after glucose administration.
Identification and microsphere content of isolated islets
Analysis of reference blood samples showed a mean value of 847 ± 46 microspheres/ml arterial blood, i.e. each microsphere was found to indicate 1.2 ± 0.2 μl blood. Approximately 500 islets could be isolated from each of these animals, and 19 ± 2 retrieved islets in each animal contained microspheres, i.e. had a detectable blood perfusion ≥1.2 ± 0.2 μl/min (Fig. 1a).

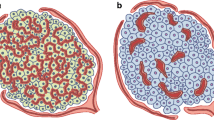
Morphological characterisation of isolated microsphere- and non-microsphere-containing islets. a Fluorescence combined with bright field microscopic image of an islet containing microspheres. Relative area of insulin-positive (b) and glucagon-positive (c) cells in islets with or without microspheres. Multiphoton confocal images of vascularity in an islet without microspheres (d) and an islet with microspheres (microsphere not visualised using this filter; e) after intravascular visualisation of blood vessels by IB4 isolectin. Quantification of vascular density (f) and average vessel diameter (g) in islets with or without microspheres. Scale bar is 50 μm in (a) and 60 μm in (d) and (e). All values are given as means±SEM from four to seven animals in each group, where for each analysis a mean number of 16.1 ± 2.6 microsphere-containing or non-microsphere-containing islets was investigated per animal. *p < 0.05 compared with non-microsphere-containing islets
Islet size and endocrine cell composition
Microsphere-containing islets were of mixed size, and the DNA content of microsphere- and non-microsphere-containing islets was similar (163 ± 20 [n = 6] vs 162 ± 18 ng DNA/10 islets [n = 6], respectively) indicating islets of equal size distribution in the two groups. The relative area of beta and alpha cells was similar in microsphere- and non-microsphere-containing islets (Fig. 1b,c).
Islet vascular density
Microsphere-containing islets had more blood vessels than non-microsphere-containing islets (Fig. 1d–f). Moreover, capillaries in microsphere-containing islets were more tortuous and wider than in non-microsphere-containing islets (Fig. 1g).
Beta cell proliferation
There was a higher rate of beta cell proliferation in highly blood-perfused islets, as identified by microspheres, compared with other islets (Fig. 2a). Similarly, other islet cells also had a higher proliferation rate in microsphere-containing islets (0.5 ± 0.1 vs 1.4 ± 0.2% Ki67-positive cells in non-microsphere- and microsphere-containing islets, respectively; eight experiments in both groups). In separate experiments, exposure of cultured islets to islet endothelial cell products (ECCM) was found to increase beta cell proliferation in exposed islets compared with control islets (Fig. 2b), whereas the proliferation of other islet cells did not differ (0.7 ± 0.3 vs 1.0 ± 0.3% Ki67-positive cells in CM- and ECCM-exposed islets, respectively; seven experiments in both groups).

Expression of the proliferation marker Ki67 in beta cells of highly blood-perfused islets as identified by microspheres when compared with other islets (a), and in islets exposed to islet endothelial cell products (ECCM) or CM (b). c Micrograph showing beta cell proliferation (insulin, red; Ki67, brown) in one of the former islets. Scale bar is 100 μm. All values are given as means±SEM from seven to eight experiments in each group. *p < 0.05 compared with non-microsphere-containing islets and islets exposed to CM, respectively
Islet function
Highly blood-perfused islets showed increased glucose-stimulated insulin release, whereas there was only a tendency to increase insulin secretion at low glucose concentration (p = 0.06; Fig. 3a). Superior capacity to respond to an acute glucose load did not seem to reflect differences in insulin production, since (pro)insulin biosynthesis and insulin content were similar in the two groups of islets (Fig. 3b,c). Moreover, leucine-dependent total protein biosynthesis was similar in the two groups of islets (837 ± 221 [n = 6] vs 1,021 ± 289 [n = 6] for control and microsphere-containing islets, respectively). Likewise, there did not seem to be any differences in mitochondrial function as assessed by a glucose oxidation rate assay (Fig. 3d). Gene arrays, however, revealed the differences delineated below in the expression of genes crucial for proper endocrine cell function between microsphere- and non-microsphere-containing islets.

Beta cell function in highly blood-perfused microsphere-containing islets compared with other islets. a Glucose-stimulated insulin release (black bars, islets without microspheres; grey bars, islets with microspheres); b (pro)insulin biosynthesis; c insulin content; d capacity for glucose oxidation. All values are given as means±SEM from five to six experiments in each group. *p < 0.05 compared with non-microsphere-containing islets
Islet gene expression
As there were no differences in glucose oxidation rates or (pro)insulin biosynthesis between the two groups of islets, the insulin gene and genes associated with mitochondrial function were not studied. Instead, we focused on genes involved in insulin exocytosis and intercommunication between cells and alpha cell and beta cell differentiation, as well as possible causes and consequences of improved vascularisation. Microsphere-containing islets had higher mRNA levels of the angiogenic factor Vegfa (Table 1). They showed higher mRNA levels for several extracellular matrix components, the alpha1, alpha3 and alpha4 chains of collagen type IV, the beta2 chain of laminin and vitronectin (Table 1). Moreover, several transcription factors important for beta cell function were expressed to a greater extent in microsphere-containing islets, namely the clock components Arntl (also known as Bmal1) and Clock (Table 1). Other factors involved in insulin exocytosis, such as Kcnj11, Grx-1 (also known as Glrx) and Gpr40 (also known as Ffar1), were also increased in microsphere-containing islets (Table 1). Moreover, the transcriptional regulator Isl-1 and transcriptional factors c-Maf (also known as Maf), Mafa, Mafb, Mafk, Nkx6-1 and Pdx1—of particular importance for pancreatic embryonic development but also adult beta or alpha cell function—were higher in microsphere-containing islets (Table 1).
Expression of Glut-2 (also known as Slc2a2) or of the gene encoding the main glycolytic enzyme, glucokinase, was not increased in microsphere-containing islets (four experiments), whereas hexokinase 1, normally present in low amounts in islet cells, showed higher expression in microsphere-containing islets (Table 1). Likewise, expression of the gene ephrin-A5—which is central to cell–cell communication in islets—was upregulated in microsphere-containing islets, as was Iapp (Table 1). Expression of the other genes evaluated did not differ between microsphere-containing and other islets (ESM Table 1).
In separate experiments, islets were exposed to islet endothelial cell products (ECCM) to assess whether the observed differences in gene expression between microsphere- and non-microsphere-containing islets reflected differences in vascular support. Few gene expression changes were, however, observed in ECCM-exposed islets compared with control islets, with only the genes encoding the beta2 chain of laminin, c-MAF and ephrin-A5 similarly increased, paralleling microsphere-containing islets (Table 2). Gene expression of Gjd2 (also known as Cx36) and the beta1 chain of laminin was also altered in ECCM-exposed islets (Table 2). Expression of the other genes evaluated did not differ (ESM Table 1).
Discussion
In this study, we characterised a subgroup of islets with higher blood perfusion than other islets. Interestingly, these highly blood-perfused islets had a higher rate of beta cell proliferation, superior capacity to respond with insulin release to a glucose challenge and markedly different gene expression compared with other islets.
In our analysis, ~5% of the isolated islets were found to contain microspheres. As the microspheres disperse in the circulation in proportion to the blood perfusion of different tissues, the presence of microspheres in islets indicates an average blood perfusion ≥1.2 ± 0.2 μl/min after glucose administration. However, by chance, islets with lower blood perfusion may in rare cases also contain microspheres. Isolation of microsphere-containing islets followed by three-dimensional vascular reconstruction of consecutive sections stained for endothelium showed that the vascular network was more dense, with wider and more tortuous capillaries, than in other islets. This indicates that their mean higher blood perfusion reflects not only more blood vessels, but also greater blood perfusion of the individual blood vessels. In islets, VEGF-A constitutively secreted from beta cells has been shown to be crucial for the early formation, branching and maintenance of the vascular network [9, 22, 23]. VEGF-A may, however, also directly increase the capillary diameter through the VEGF2 receptor [24]. Interestingly, our gene expression studies identified increased expression of Vegfa in microsphere-containing islets, providing a likely mechanism for their higher vascular density and increased capillary width. Their greater blood perfusion and preferential responsiveness with further increases in blood flow after a glucose load may also reflect their higher functional activity. Indeed, the glucose-dependent increase in blood flow of islets has been shown to depend on vagal nervous signals combined with generation of vasoactive metabolites, predominantly adenosine, in islet tissue exposed to a prolonged metabolic demand [13]. It is likely that these islets generate more adenosine as a result of increased beta cell functional response to glucose.
We herein identify for the first time a subpopulation of islets with a higher rate of beta cell proliferation than other islets, namely the most highly blood-perfused islets. To investigate the role of this greater vascular support, we isolated islet endothelial cells and exposed cultured islets to ECCM. Indeed, an increased rate of beta cell proliferation was observed in such islets compared with control islets, showing that islet endothelial paracrine factors can increase beta cell proliferation. This suggests a contributing effect of endothelial cell signals to a higher rate of beta cell proliferation in microsphere-containing islets. Previous studies have shown that exposure of isolated murine islets to vascular basement proteins, predominantly laminins, stimulates beta cell proliferation [4]. Our gene expression analyses also verified upregulation of several basement membrane proteins in the islets with greater blood perfusion, which probably reflects their increased number of endothelial cells. The beta2 chain of laminin has been shown to be prominent in vascular basement membranes in human islets also [25]. Upregulation of the clock components Arntl and Clock in higher blood-perfused islets may also contribute to their higher rate of beta cell proliferation (compare with the study of Marcheva et al. [26]), although these genes were not upregulated in ECCM-exposed islets. Likewise, the transcriptional regulator Isl-1 (upregulated in microsphere-containing islets, but not in ECCM-exposed islets) has been identified as crucial for sustained beta cell proliferation and the prevention of apoptosis in postnatal beta cells [27]. It is noteworthy that a difference from the studies of retrieved microsphere-containing islets was that ECCM-exposed islets needed to be precultured in order to deplete their endogenous endothelial cells, which may have affected the responsiveness of their gene expression.
Previous studies have described heterogeneities in function between individual beta cells [28, 29]. Our results suggest this also to be the case for whole islets. Recently, we showed that the 20–25% of islets with the lowest blood perfusion normally have low oxygenation and constitute a metabolically dormant subpopulation of islets, i.e. providing a functional reserve of endocrine cells potentially activated when needed [30]. That pancreatic islets in vivo can accumulate the low oxygenation marker pimonidazole was also recently shown in retrieved islets using western blot, although this could not be corroborated in that study by immunohistochemistry [31]. In this study, we identified another subpopulation of islets, the 5% with the greatest blood perfusion, which produce greater glucose-stimulated insulin release than other islets although containing a similar relative area of beta cells and amounts of insulin. However, in contrast with the islets with lowest blood perfusion, they have rates of (pro)insulin and total protein biosynthesis similar to those of other islets. Moreover, their mitochondrial function, as assessed by measurement of rates of glucose oxidation, is similar to that in other islets. Instead, a number of differences in gene expression were observed, indicating other mechanisms for the improved beta cell secretion capacity. The clock components Arntl and Clock showed, as mentioned above, greater expression in the highly blood-perfused islets. These components do not solely increase beta cell growth; Arntl also augments insulin release in response to glucose, KCl, exendin-4, forskolin and 8-bromo-cAMP, indicating its importance for insulin exocytosis [26]. In fact, pancreas-specific loss of Arntl causes chronic hyperglycaemia [26]. Furthermore, Clock is necessary for beta cell function, which is related to involvement in insulin exocytosis [26]. Other genes involved in insulin exocytosis, Kcnj11 and Grx-1, were also found to be upregulated in the islets with higher blood perfusion. Variants of KcnJ11 encoding the Kir 6.2 subunit of the ATP-sensitive potassium (KATP) channel have previously been shown to be among the strongest predictors of type 2 diabetes development [32, 33], and heterozygous mutations in this gene cause permanent neonatal diabetes [34]. Grx-1 mediates NADPH-dependent stimulation of calcium-dependent insulin secretion [35]. Gpr40 mediates fatty acid potentiation of glucose-stimulated insulin secretion [36] and can, when overexpressed, augment glucose-stimulated insulin secretion [37], possibly by increasing intracellular inositol phosphate levels [38]. Gpr40 was upregulated in the more highly blood-perfused islets, potentially contributing to the superior function found in these islets.
Another group of genes found to be predominantly expressed in the more highly blood-perfused islets were those of disparate transcription factors, principally known for their importance during pancreatic embryonic development. Among these were: Isl1, a direct regulator of Mafa transcription in beta cells [27]; Pdx1, also important for beta cell-specific Mafa expression [39]; and Mafa, as well as the other MAF transcription factors c-Maf, Mafb, Mafk and Nkx6-1. Pdx1 is important for adult beta cells to maintain phenotype and Pdx1 +/− mice are glucose intolerant, suggesting that gene dosage is critical for these cells [40]. Mafa has previously been reported to be a key regulator of glucose-stimulated insulin release from adult islet beta cells [41], although a decrease in beta cell mass and glucose toxicity to the cells may contribute to the reported dysfunction in diabetic animals. Moreover, Mafa is important for neonatal rat beta cells to acquire glucose-responsive insulin secretion [42], whereas Mafk acts to antagonise the function of Mafa [43]. Mafb and c-Maf are not expressed in adult beta cells, being instead salient to adult islet alpha cell function [44]. The Nkx6.1 homeodomain transcription factor has been shown to regulate glucose-stimulated insulin secretion in adult islet beta cells, primarily by suppressing genes impairing nutrient-stimulated insulin release [45].
Expression of glucokinase did not differ in the more highly blood-perfused islets, although, quite surprisingly, hexokinase was upregulated. Hexokinase is known to be present in low amounts in pancreatic endocrine cells [46], but, if upregulated, produces a leftward shift of the normal concentration-dependent activation of glucose-induced insulin release [47]. This could decrease the threshold for glucose-induced insulin release or increase insulin secretion at basal plasma glucose levels [48]. Such a tendency (p = 0.06) was also observed for the more highly blood-perfused islets in the present study. However, the increased expression of hexokinase in the latter may also to some extent reflect the 50% increase in their vascular density, i.e. the presence of more endothelial cells.
Another interesting finding was the upregulation of the gene for ephrin-A5 in the more highly blood-perfused islets. Ephrin-A5 has previously been shown to underpin cell–cell communication between beta cells in islets, and lack of this ephrin results in decreased capacity for glucose-stimulated insulin release while not affecting insulin production [49]. Interestingly, the islet hormone Iapp was expressed to a higher extent in richly blood-perfused islets. Although the importance of this remains obscure, it may indicate a predisposition to amyloid formation in these highly functional islets if not compensated via improved drainage resulting from the high blood perfusion. Clinically, amyloid formation occurs unevenly between islets, with islets in peripheral regions of the pancreas being more prone to developing such deposits [50].
Using the model of ECCM exposure of cultured islets, we have previously shown that endothelial-derived factors, predominantly laminins, augment glucose-stimulated insulin release [5]. The present study reveals that the intercommunication between endothelial cells and beta cells is more extensive. This involves VEGF-A, which is more highly produced in the beta cells of some islets, thereby inducing a more abundant vascular network. These islets are highly blood perfused, providing optimal oxygenation, glucose sensing and disposal of secreted islet hormones. Islet endothelial paracrine signalling, as assessed by exposure of islets to ECCM, also directly supports the endocrine cells and induces changes in expression of the genes encoding the beta1 and beta2 chains of laminin, in conjunction with c-Maf, ephrin-A5 and Gjd2. However, most functional changes in gene expression observed in microsphere-containing islets were not induced by ECCM, which may either reflect changed responsiveness of gene expression due to the necessary culture or indicate that these changes were inherent in this subgroup of islets. The induction of a more abundant vascular network supported by VEGF-A would then be part of this inherent capacity.
In conclusion, we herein report a novel heterogeneity between islets with regard to beta cell proliferation, function and gene expression. A subpopulation of islets with the highest blood perfusion was studied and found to have a higher rate of beta cell proliferation, as well as superior function reflecting several differences in gene expression compared with other islets. In contrast with the previously described population of low-oxygenated, sleeping islets, which are recruited into functionality when needed, the presently described heterogeneity is shown to remain in vitro after islet isolation.
Abbreviations
- CM:
-
Culture medium
- ECCM:
-
Endothelium-conditioned culture medium
- VEGF-A:
-
Vascular endothelial growth factor-A
References
Bonner-Weir S, Orci L (1982) New perspectives on the microvasculature of the islets of Langerhans in the rat. Diabetes 31:883–889
Nyman LR, Wells KS, Head WS et al (2008) Real-time, multidimensional in vivo imaging used to investigate blood flow in mouse pancreatic islets. J Clin Invest 118:3790–3797
Lammert E, Cleaver O, Melton D (2001) Induction of pancreatic differentiation by signals from blood vessels. Science 294:564–567
Nikolova G, Jabs N, Konstantinova I et al (2006) The vascular basement membrane: a niche for insulin gene expression and beta cell proliferation. Dev Cell 10:397–405
Johansson A, Lau J, Sandberg M, Borg LA, Magnusson PU, Carlsson PO (2009) Endothelial cell signalling supports pancreatic beta cell function in the rat. Diabetologia 52:2385–2394
Olerud J, Mokhtari D, Johansson M et al (2011) Thrombospondin-1: an islet endothelial cell signal of importance for {beta}-cell function. Diabetes 60:1946–1954
Johansson M, Mattsson G, Andersson A, Jansson L, Carlsson PO (2006) Islet endothelial cells and pancreatic beta-cell proliferation: studies in vitro and during pregnancy in adult rats. Endocrinology 147:2315–2324
Brunicardi FC, Stagner J, Bonner-Weir S et al (1996) Microcirculation of the islets of Langerhans. Long beach veterans administration regional medical education center symposium. Diabetes 45:385–392
Brissova M, Shostak A, Shiota M et al (2006) Pancreatic islet production of vascular endothelial growth factor-a is essential for islet vascularization, revascularization, and function. Diabetes 55:2974–2985
Jansson L (1996) Microsphere distribution in the pancreas of anesthetized rats. Alloxan stimulates the blood flow to all islets whereas glucose only affects the blood perfusion of a subgroup of islets. Int J Pancreatol 20:69–74
Carlsson PO, Kallskog O, Bodin B, Andersson A, Jansson L (2002) Multiple injections of coloured microspheres for islet blood flow measurements in anaesthetised rats: influence of microsphere size. Ups J Med Sci 107:111–120
Carlsson PO, Jansson L, Ostenson CG, Kallskog O (1997) Islet capillary blood pressure increase mediated by hyperglycemia in NIDDM GK rats. Diabetes 46:947–952
Carlsson PO, Olsson R, Kallskog O, Bodin B, Andersson A, Jansson L (2002) Glucose-induced islet blood flow increase in rats: interaction between nervous and metabolic mediators. Am J Physiol Endocrinol Metab 283:E457–E464
Nyman LR, Ford E, Powers AC, Piston DW (2010) Glucose-dependent blood flow dynamics in murine pancreatic islets in vivo. Am J Physiol Endocrinol Metab 298:E807–E814
Jansson L, Hellerstrom C (1983) Stimulation by glucose of the blood flow to the pancreatic islets of the rat. Diabetologia 25:45–50
Andersson A, Westman J, Hellerstrom C (1974) Effects of glucose on the ultrastructure and insulin biosynthesis of isolated mouse pancreatic islets maintained in tissue culture. Diabetologia 10:743–753
Weibel E (ed) (1979) Practical methods for biological morphometry. Academic Press, London
Jackson CJ, Garbett PK, Nissen B, Schrieber L (1990) Binding of human endothelium to Ulex europaeus I-coated Dynabeads: application to the isolation of microvascular endothelium. J Cell Sci 96(Pt 2):257–262
Mattsson G, Danielsson A, Kriz V, Carlsson PO, Jansson L (2006) Endothelial cells in endogenous and transplanted pancreatic islets: differences in the expression of angiogenic peptides and receptors. Pancreatology 6:86–95
Nyqvist D, Kohler M, Wahlstedt H, Berggren PO (2005) Donor islet endothelial cells participate in formation of functional vessels within pancreatic islet grafts. Diabetes 54:2287–2293
Andersson A (1974) Long-term effects of glucose on insulin release and glucose oxidation by mouse pancreatic islets maintained in tissue culture. Biochem J 140:377–382
Lammert E, Gu G, McLaughlin M et al (2003) Role of VEGF-A in vascularization of pancreatic islets. Curr Biol 13:1070–1074
Kamba T, Tam BY, Hashizume H et al (2006) VEGF-dependent plasticity of fenestrated capillaries in the normal adult microvasculature. Am J Physiol Heart Circ Physiol 290:H560–H576
Nakatsu MN, Sainson RC, Perez-del-Pulgar S et al (2003) VEGF(121) and VEGF(165) regulate blood vessel diameter through vascular endothelial growth factor receptor 2 in an in vitro angiogenesis model. Lab Invest 83:1873–1885
Virtanen I, Banerjee M, Palgi J et al (2008) Blood vessels of human islets of Langerhans are surrounded by a double basement membrane. Diabetologia 51:1181–1191
Marcheva B, Ramsey KM, Buhr ED et al (2010) Disruption of the clock components CLOCK and BMAL1 leads to hypoinsulinaemia and diabetes. Nature 466:627–631
Du A, Hunter CS, Murray J et al (2009) Islet-1 is required for the maturation, proliferation, and survival of the endocrine pancreas. Diabetes 58:2059–2069
Pipeleers DG (1992) Heterogeneity in pancreatic beta-cell population. Diabetes 41:777–781
Pipeleers D, Kiekens R, Ling Z, Wilikens A, Schuit F (1994) Physiologic relevance of heterogeneity in the pancreatic beta-cell population. Diabetologia 37(Suppl 2):S57–S64
Olsson R, Carlsson PO (2011) A low-oxygenated subpopulation of pancreatic islets constitutes a functional reserve of endocrine cells. Diabetes 60:2068–2075
Sato Y, Endo H, Okuyama H et al (2011) Cellular hypoxia of pancreatic beta-cells due to high levels of oxygen consumption for insulin secretion in vitro. J Biol Chem 286:12524–12532
Lyssenko V, Jonsson A, Almgren P et al (2008) Clinical risk factors, DNA variants, and the development of type 2 diabetes. N Engl J Med 359:2220–2232
Scott LJ, Mohlke KL, Bonnycastle LL et al (2007) A genome-wide association study of type 2 diabetes in Finns detects multiple susceptibility variants. Science 316:1341–1345
Gloyn AL, Pearson ER, Antcliff JF et al (2004) Activating mutations in the gene encoding the ATP-sensitive potassium-channel subunit Kir6.2 and permanent neonatal diabetes. N Engl J Med 350:1838–1849
Reinbothe TM, Ivarsson R, Li DQ et al (2009) Glutaredoxin-1 mediates NADPH-dependent stimulation of calcium-dependent insulin secretion. Mol Endocrinol 23:893–900
Itoh Y, Kawamata Y, Harada M et al (2003) Free fatty acids regulate insulin secretion from pancreatic beta cells through GPR40. Nature 422:173–176
Nagasumi K, Esaki R, Iwachidow K et al (2009) Overexpression of GPR40 in pancreatic beta-cells augments glucose-stimulated insulin secretion and improves glucose tolerance in normal and diabetic mice. Diabetes 58:1067–1076
Alquier T, Peyot ML, Latour MG et al (2009) Deletion of GPR40 impairs glucose-induced insulin secretion in vivo in mice without affecting intracellular fuel metabolism in islets. Diabetes 58:2607–2615
Raum JC, Gerrish K, Artner I et al (2006) FoxA2, Nkx2.2, and PDX-1 regulate islet beta-cell-specific mafA expression through conserved sequences located between base pairs −8118 and −7750 upstream from the transcription start site. Mol Cell Biol 26:5735–5743
Dutta S, Bonner-Weir S, Montminy M, Wright C (1998) Regulatory factor linked to late-onset diabetes? Nature 392:560
Zhang C, Moriguchi T, Kajihara M et al (2005) MafA is a key regulator of glucose-stimulated insulin secretion. Mol Cell Biol 25:4969–4976
Aguayo-Mazzucato C, Koh A, El Khattabi I et al (2011) Mafa expression enhances glucose-responsive insulin secretion in neonatal rat beta cells. Diabetologia 54:583–593
Shimohata H, Yoh K, Morito N, Shimano H, Kudo T, Takahashi S (2006) MafK overexpression in pancreatic beta-cells caused impairment of glucose-stimulated insulin secretion. Biochem Biophys Res Commun 346:671–680
Artner I, Hang Y, Mazur M et al (2010) MafA and MafB regulate genes critical to beta-cells in a unique temporal manner. Diabetes 59:2530–2539
Schisler JC, Jensen PB, Taylor DG et al (2005) The Nkx6.1 homeodomain transcription factor suppresses glucagon expression and regulates glucose-stimulated insulin secretion in islet beta cells. Proc Natl Acad Sci U S A 102:7297–7302
Schuit F, Moens K, Heimberg H, Pipeleers D (1999) Cellular origin of hexokinase in pancreatic islets. J Biol Chem 274:32803–32809
German MS (1993) Glucose sensing in pancreatic islet beta cells: the key role of glucokinase and the glycolytic intermediates. Proc Natl Acad Sci U S A 90:1781–1785
Heimberg H, de Vos A, Vandercammen A, van Schaftingen E, Pipeleers D, Schuit F (1993) Heterogeneity in glucose sensitivity among pancreatic beta-cells is correlated to differences in glucose phosphorylation rather than glucose transport. EMBO J 12:2873–2879
Konstantinova I, Nikolova G, Ohara-Imaizumi M et al (2007) EphA-Ephrin-A-mediated beta cell communication regulates insulin secretion from pancreatic islets. Cell 129:359–370
Borromeo CM, Pottier X, In’t Veld PA et al (2005) Heterogeneity in distribution of amyloid-positive islets in type-2 diabetic patients. Virchows Arch 446:232–238
Acknowledgements
We gratefully acknowledge the skilled technical assistance of Birgitta Bodin, Astrid Nordin, Eva Törnelius and My Quach. We are also grateful to Beau Corazon for linguistic revision (Department of Medical Cell Biology, Uppsala University).
Funding
The study was generously supported by the JDRF, the Swedish Research Council, EFSD/Novo Nordisk Programme 2010, AFA Insurance, the Swedish Diabetes Association, the Swedish Juvenile Diabetes Foundation, the Novo Nordisk Foundation, Diabetes Wellness Sverige, the Thuring Foundation, the Anér Foundation, the Swedish Society for Medical Research, Goljes Memorial Fund and the Family Ernfors Fund.
Duality of interest
The authors declare that there is no duality of interest associated with this manuscript.
Contribution statement
JL, JS, LG, ÅJ and P-OC conceived and designed the study, and participated in the analysis and interpretation of the data. P-OC drafted the manuscript, and all other authors revised it critically for intellectual content. All authors approved the final version of the paper.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM Table 1
(PDF 99 kb)
Rights and permissions
About this article
Cite this article
Lau, J., Svensson, J., Grapensparr, L. et al. Superior beta cell proliferation, function and gene expression in a subpopulation of rat islets identified by high blood perfusion. Diabetologia 55, 1390–1399 (2012). https://doi.org/10.1007/s00125-012-2476-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00125-012-2476-6