
Abstract
Aims/hypothesis
Hypoxic pulmonary vasoconstriction is an essential mechanism to prevent hypoxaemia in lung diseases. Insulin is known to be a systemic vasodilator but its effect on the pulmonary circulation is not known. Inhaled particulate insulin can generate locally high concentrations in the lung which could be physiologically important.
Methods
We therefore studied the effects of insulin in vitro on isolated rat pulmonary artery in a small vessel myograph.
Results
We have shown that pulmonary artery vasodilatation with insulin occurs in a dose-dependent manner. Pre-constriction with PGF2α can be abolished (105.7±2.9%, mean±SEM) and pre-constriction with hypoxia reduced (68.9±6.5%) by pharmacologically relevant concentrations of insulin. The characteristic phasic vasoconstriction by pulmonary vessel to hypoxia is substantially modified, resulting in sustained vasodilatation.
Conclusions/interpretation
These effects could be clinically important for patients using inhaled insulins who have acute or occult chronic lung disease.
Similar content being viewed by others

Hypoxic pulmonary vasoconstriction (HPV) is an adaptive mechanism which matches perfusion to ventilation in order to minimise systemic hypoxaemia. In acute lung disease with ventilation-perfusion mismatch—e.g. acute asthma, pneumonia—HPV shunts blood away from poorly ventilated alveoli. Any exogenous or endogenous stimulus which reverses HPV can potentially worsen ventilation-perfusion mismatch. In chronic lung disease with more generalised hypoxia, HPV becomes a maladaptation, leading to chronic pulmonary hypertension. Reversal of HPV in this situation merely allows greater pulmonary arterial inflow to hypoxic alveoli, leading to further desaturation with potentially serious consequences where critical hypoxia already exists. Although insulin is known to be a systemic vasodilator, whose mode of action involves nitric oxide (NO), cycooxygenase pathway, adenosine and ATP-sensitive K+ channels, its effect on pulmonary vessels was previously not known. If it were an active pulmonary vasodilator, it may have important clinical implications. At least one selective pulmonary vasodilator (nitric oxide) has been shown to worsen arterial hypoxaemia by reversal of HPV and resulted in greater dispersion of blood flow in chronic obstructive airway disease [1].
With inhaled insulin, only a small percentage of the inhaled particles are absorbed into systemic circulation. It is possible that exposure in supra-physiological doses occurs at the alveolar-arterial interface. To date, clinical studies of inhaled insulin have excluded people with lung disease. We have undertaken an in vitro study to describe the reactivity characteristics of rat pulmonary arteries in response to insulin.
Materials and methods
Vessel preparation
Small side-branch pulmonary arteries were dissected from male Wistar rats (body mass 220–250 g, reared following the principles of laboratory animal care) and mounted on two 40 µm stainless steel wires in the organ bath of a small vessel myograph (Cambustion, Cambridge, UK) as described by [2]. The vessels were bathed in PSS (120 mmol/l NaCl, 4.7 mmol/l KCl, 1.17 mmol/l MgSO4, 25 mmol/l NaHCO3, 1.18 mmol/l KH2PO4, 5.5 mmol/l Glucose, 2.5 mmol/l CaCl2 and 26.9 µmol/l EDTA) and 95%O2, 5%CO2 bubbled into the bathing solution which was at a constant 37°C. Length tension curves were then plotted for each vessel before loading to a tension equivalent to a transmural pressure of 17.5 mmHg, the normal in vivo resting tension.
All vessels were left to equilibrate at their resting tension for 1 h. All experiments were carried out isometrically. At the end of the equilibration period when baseline tension was steady a maximal KCl contraction (80 mmol/l) was measured. The bath was washed three times with fresh PSS pre heated to 37°C and the vessel allowed to relax, until a stable baseline tension returned. This was repeated twice more to ensure reproducibility of the contractile response within each vessel. Next PGF2α (100 µmol/l) was added to the bathing solution with subsequent administration of ACh (1 µmol/l) once a maximal contraction had been achieved, to indicate endothelial integrity.
The effect of insulin on the PGF2α constriction
Vessels dissected (n=17) with a diameter 383±26 µm were mounted and preliminary data obtained as described above. After a period of equilibrium PGF2α (100 µmol/l) was added to the bathing solution to achieve maximum contraction and was allowed to stabilise for 45 min (results will be expressed as a percentage of this contraction). Cumulative doses of insulin (0.1 µmol/l–300 µmol/l) were then added to the organ bath at 5-min intervals or until the result had reached a plateau.
The effect of insulin on hypoxic pulmonary vasoconstriction (HPV)
Vessels dissected (n=11) with a diameter 449±36 µm were mounted and preliminary data obtained as described above. Once this preliminary data had been obtained the following experiments were done. PGF2α (5 µmol/l) was added to the bathing solution to achieve a submaximal contraction and was allowed to stabilise for 5 min; 95% N2/5% CO2 was then bubbled through the organ baths and lids were placed on the top of the organ bath. A four-phase response was observed as described previously [3, 4] (results will be expressed as a percentage of the phase four contraction). Cumulative doses of insulin (0.001 µmol/l–30 µmol/l) were then added to the organ bath at 5-min intervals or until the result had reached a plateau.
Data analysis
Values were expressed as mean±standard error (SE) of the mean. A significant difference between the mean values was determined using the appropriate ANOVA test while using the SPSS software package. In myograph experiments individual contractions are expressed as a percentage of the maximal induced tone for that vessel, and the mean values calculated for the number of vessels in each group. This was done to standardise responses between vessels, since magnitude of contraction varied between vessels. The potency of a substance was expressed as the negative logarithm of the concentration required to produce half the maximal relaxation (EC50). This was calculated for each experimental vessel to make a statistical comparison. Significance was assumed if the p value was less than 0.05.
Results
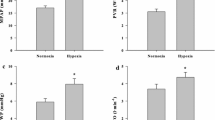
Vessel diameter, maximum contraction to PGF2α or HPV, maximal relaxation obtained at the highest doses of insulin and EC50 are shown in Table 1. There was no significant difference in vessel size between vessels pre-constricted to PGF2α or HPV (383±26, 449±36 µm respectively, p=0.14). The last two KCl (100 mmol/l) contractions of the initial three contractions for each individual vessel were of a similar magnitude in the PGF2α group (3.30±0.28, 2.76±0.24 mN/mm, p=0.15) and also in the HPV group (0.89±0.11, 0.96±0.14 mN/mm, p=0.42) indicating reproducibility of the contractile response for those vessels. All of the vessels were shown to have an intact endothelium by relaxant responses to ACh (1 µmol/l) in the PGF2α group (26±4% reversal of PGF2α 100 µmol/l contraction) and also in the HPV group (19±2% reversal of PGF2α 100 µmol/l contraction). Vessels did, however, significantly differ in levels of induced tone, PGF2α pre-constricted vessels developed more induced tone (2.5±0.2mN/mm) compared to HPV pre-constricted vessels (0.8±0.1 mN/mm) (p>0.001). Insulin was also shown to be more potent in vessels pre-constricted to HPV (6.2±0.2) compared to vessels pre-constricted to PGF2α (4.5±0.0) (p>0.001).
Dose response curves to insulin in PGF2α and HPV groups are shown (Fig. 1, 2). These results show that insulin vasodilates rat pulmonary resistance vessels pre-constricted with PGF2α and HPV in a dose-dependant manner; however, insulin is capable of completely reversing the PGF2α pre-constriction (105.7±2.9%) compared to the HPV pre-constriction, which is only partially abolished (68.9±6.5%).

Dose response curve to insulin against PGF2α contraction

Dose response curve to insulin against HPV contraction
Of interest, vessels pre-constricted to HPV also exhibited a triphasic response to higher doses of insulin, an initial dilatation is followed by constriction and then a further longer lasting dilatation. This whole triphasic response was complete on average 5 min after the introduction of the insulin to the organ bath (Fig. 3).

Schematic presentation of modification of phasic HPV response by insulin. The top panel shows characteristic four-phase response of the pulmonary vessel to hypoxia: i.e. vasodilatation (1), vasoconstriction (2), vasodilation (3) and vasoconstriction (4). The bottom panel characterises hypoxic response in the presence of insulin in which initial vasodilatation (1) is followed by vasoconstriction (2) and finally by sustained vasodilatation (3)
Discussion
Gas exchange depends crucially on the proper matching of ventilation and perfusion. Depending on the relative effectiveness of ventilation and perfusion, the possible ratio in any alveolar-capillary unit ranges from zero, where lack of ventilation leads to arteriovenous shunt with no gas exchange, to infinity, where there is no perfusion. The hypothetical 'ideal' ratio is one where ventilation is equivalent to perfusion but even in health, there is some ventilation-perfusion mismatch as there is normally a gradient of blood flow from bases to the apices and a similar, albeit less marked, gradient of ventilation from the apices to the bases. Hypoxic pulmonary vasoconstriction is the primary physiological response to respiratory insult, reducing perfusion of inadequately ventilated areas of the lung, thereby attempting to restore ventilation-perfusion matching. Reversal of HPV has been shown to cause dispersal of blood flow to hypoventilated areas and worsen hypoxaemia [1]. Whereas it is unlikely for the inhaled route to be the chosen mode of insulin delivery in patients with overt clinical lung disease, many patients, particularly smokers, are unaware that they have lung disease. To date, clinical trials [5] of inhaled insulins have carefully excluded any subject with known pulmonary dysfunction; therefore, it is only possible to conclude that in healthy subjects inhaling insulin does not (at least in the short term) lead to significant respiratory impairment.
In this in vitro model we have shown that insulin has a dose-dependent vasodilatory effect on prostaglandin-induced pulmonary artery vasoconstriction. We have also shown a substantial modification of the phased vasoconstriction response to hypoxia, which is the most important physiological pulmonary vasoconstrictor. We have characterised these responses and produced an accurate measure of their magnitude. These changes observed at concentrations several orders of magnitude less than particulate or aerosolised U500-insulin preparation can be expected to be produced by local pulmonary deposition.
It is invariably observed that only 10 to 20% of inhaled insulin reaches the systemic circulation [6, 7, 8]; thus locally-mediated pharmacological effects in the lung are not only conceivable but very likely to be important. It is therefore imperative that these potential effects be studied in detail in humans, with clinical trials specific to different lung diseases.
Abbreviations
- ACh:
-
Acetylcholine
- HPV:
-
hypoxic pulmonary vasoconstriction
- PGF2α :
-
prostaglandin F2α
- EC50 :
-
negative logarithm of the concentration required to induce half maximal vasodilatation (i.e., a measure of vasodilatory potency)
References
Barbera JA, Roger N, Roca J, Higenbottam TW, Rodriguez-Rorsin R (1996) Worsening of pulmonary gas exchange with nitric oxide inhalation in chronic obstructive pulmonary disease. Lancet 347:436–440
Rogers TK, Stewart AG, Morice AH (1992) Effect of chronic hypoxia on rat pulmonary resistance vessels: vasodilatation by atrial natriuretic peptide. Clin Sci (Lond) 83:723–729
Rogers TK, Thomson JS, Morice AH (1997) Inhibition of hypoxic pulmonary vasoconstriction in isolated rat resistance arteries by ANP. Eur Respir J 10:2061–2065
Wanstall JC, O'Brien E (1996) In vitro hypoxia on rat pulmonary artery: effects on contractions to spasmogens and role of KATP channels. Eur J Pharmacol 303:71–78
Skyler JS, Cefalu WT, Kourides IA et al. (2001) Efficacy of inhaled human insulin in type 1 diabetes mellitus: a randomised proof-of-concept study. Lancet 357:331–335
Wigley FW, et al. (1971) Insulin across respiratory mucosae by aerosol delivery. Diabetes 20:552–556
Laube BL, Benedict GW, Dobs AS (1998) Time to peak insulin level, relative bioavailability, and effect of site of deposition of nebulized insulin in patients with noninsulin-dependent diabetes mellitus. J Aerosol Med 11:153–173
Steiner S, et al. (2002) Technosphere/Insulin—proof of concept study with a new insulin formulation for pulmonary delivery. Exp Clin Endocrinol Diabetes 110:17–21.
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Aye, M., Sheedy, W., Harrison, R. et al. Pulmonary vasodilation in the rat by insulin in vitro could indicate potential hazard for inhaled insulin. Diabetologia 46, 1199–1202 (2003). https://doi.org/10.1007/s00125-003-1172-y
Received:
Revised:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00125-003-1172-y