
Abstract
Recent research has revealed extensive pheromonal parsimony within the large beetle family Cerambycidae, with closely related species producing the same or very similar pheromone components. This article summarizes research that evaluated attraction of cerambycids to individual pheromone components, blends of pheromone components, and combinations of pheromones with host plant volatiles. Field bioassays were carried out, in collaboration with the Pennsylvania Department of Agriculture and the USDA Cooperative Agricultural Pest Survey program, in 10–25 counties of Pennsylvania over 3 years. A total of 15,438 cerambycids of 134 species were captured, including two exotic species. Semiochemical lures attracted significant numbers of beetles in species of the subfamilies Cerambycinae, Lamiinae, and Spondylidinae, but were not attractive to species in the Lepturinae, Parandrinae, and Prioninae. These experiments reconfirmed the behavioral roles of semiochemicals for a number of species that have been studied previously, and yielded new information about semiochemistry of several species. The host plant volatile α-pinene enhanced attraction of species that were conifer specialists, whereas ethanol enhanced attraction of some species of hardwood specialists. The data suggest that species which share dominant pheromone components avoid cross attraction by differing in seasonal activity period, and by antagonistic effects of minor pheromone components on attraction of heterospecifics. This study further supports the concept that with careful choice of components, multiple pheromones can be deployed as single blends, and paired with host plant volatiles, to maximize the number and taxonomic diversity of cerambycid beetles that are attracted to a single lure, so that the number of different lures that must be deployed can be minimized.
Similar content being viewed by others


Introduction
Research during the past decade has revealed substantial pheromonal parsimony within the large beetle family Cerambycidae in terms of both closely related (congeneric) and more distantly related species (in different subfamilies) producing the same or very similar pheromone components. The majority of pheromones identified to date are produced by males and attract both sexes. For example, 3-hydroxyhexan-2-one, the analogous 2,3-hexanediols, and the homologous 8-carbon compounds are common, and often the sole pheromone components for many species in the subfamily Cerambycinae (Millar et al. 2009). (E)-6,10-Dimethyl-5,9-undecadien-2-ol (termed fuscumol) is the shared pheromone component of North American and European species in the genus Tetropium (subfamily Spondylidinae; Silk et al. 2007; Sweeney et al. 2010), and a South American species in the subfamily Lamiinae (Fonseca et al. 2010). In addition, several North American lamiine species are attracted by fuscumol and/or its acetate, which are likely the pheromone components for these species (Mitchell et al. 2011). Mature males of lamiine species in the genus Monochamus, native to Asia, Europe, and North America, produce the pheromone 2-(undecyloxy)-ethanol (henceforth monochamol) to which both sexes are attracted (Pajares et al. 2010; Teale et al. 2011; Allison et al. 2012; Fierke et al. 2012), and the same compound is strongly implicated as a male-produced pheromone component of another two North American Monochamus species (Macias-Samano et al. 2012; unpublished data).
In addition, several female-produced sex pheromones have been identified within the subfamilies Prioninae and Lepturinae, and again, pheromone structures often appear to be shared among congeners. For example, female Prionus californicus Motschulsky (subfamily Prioninae), produce (3R,5S)-3,5-dimethyldodecanoic acid as their primary pheromone component (Rodstein et al. 2011), but the same compound also attracts males of several North American and European congeners (Barbour et al. 2011). Remarkably, females of at least three Tragosoma species (subfamily Prioninae) produce 2,3-hexanediol stereoisomers as sex pheromones, i.e., the same compounds that are male-produced aggregation pheromones of many cerambycine species (Ray et al. 2012b).
An increasing body of literature has shown that the similarity in pheromone composition across cerambycid taxonomic groups results in simultaneous attraction of multiple species to traps baited with single or multiple synthetic pheromone components (e.g., Hanks et al. 2012; Wong et al. 2012). Thus, our initial objective was to assess the variety of species that would be attracted to some of the pheromone components that were known from multiple species. We further anticipated that the results from these trials would provide leads for full identification of the pheromones of additional species, based on the reasonable assumption that a compound that attracted a certain species was likely to be a pheromone component of that species.
When it became clear during the first field season that the pheromone components were broadly attractive to many cerambycid species (see “Results”), we added two further objectives. The first was to determine whether different pheromone components could be combined to make generic lures that could be used to attract multiple species in different tribes and subfamilies to a single trap. Such generic lures would be of enormous practical benefit for detecting and monitoring cerambycids (e.g., Witzgall et al. 2010). Our final objective, developed during the third year, was to determine whether host plant volatiles might enhance attraction of cerambycids to synthetic pheromones.
Thus, in the first year (2009), we tested three pheromone motifs that are known to be common among North American cerambycids, namely six-carbon hydroxyketones (racemic 3-hydroxyhexan-2-one) and diols (generic blend of all four stereoisomers of 2,3-hexanediol), and an inexpensive formulation of racemic (E/Z)-fuscumol and its corresponding acetate. In the second year, based on results from 2009 and other ongoing trials, we extended the library of test treatments to include five compounds: racemic 3-hydroxyhexan-2-one, the diastereomerically pure but racemic (2R*,3R*)- and (2R*,3S*)-2,3-hexanediols, racemic (E/Z)-fuscumol + fuscumol acetate, and monochamol, tested separately. We also deployed a blend of the latter three components to test for agonistic and inhibitory interactions among these three compounds. For comparison with standard monitoring treatments, we also included two different types of generic lures: ethanol + α-pinene, which commonly are used for monitoring and quarantine surveillance of wood-boring insects (e.g., see Brockerhoff et al. 2006b; Witzgall et al. 2010), and fermenting sugar bait, which has been reported to attract certain cerambycid species (Linsley 1961).
The testing strategy was modified in two separate ways in the third year. First, based on results showing that different cerambycid pheromones generally could be combined without substantial inhibitory interactions among the components (Wong et al. 2012), we formulated a generic lure containing six known cerambycid pheromones: racemic 3-hydroxyhexan-2-one, (2R*,3R*)-2,3-hexanediol, racemic (E)-fuscumol, racemic (E)-fuscumol acetate, monochamol, and racemic 2-methylbutan-1-ol [the (R)-enantiomer of 2-methylbutan-1-ol is a pheromone component of species in several genera within the Cerambycinae; Hanks et al. 2007; unpublished data]. Second, based on recent reports that host plant volatiles enhance attraction of cerambycid beetles to pheromones (e.g., Silk et al. 2007; Fonseca et al. 2010; Pajares et al. 2010), and our own preliminary results (Teale et al. 2011; Hanks et al. 2012), we deployed the same generic lure with and without the host plant volatiles ethanol, and α- and β-pinene, to assess the influence and importance of host plant volatiles in combination with pheromones, for a large number of species simultaneously.
Field trials were deployed over the entire field season for each of the 3 years, during which more than 15,000 beetles of 134 species were trapped (see “Results”). This large data set provided substantial amounts of information with regard to the seasonal activity patterns of numerous species, and phenological mechanisms that might limit cross attraction between species that share common pheromone components. Thus, our final, adventitious objective was to mine the data set for trends in pheromone use. In particular, we hypothesized that cerambycid species that shared a common pheromone component would be likely to have different activity periods, and/or that their pheromone blends might contain minor components that inhibited cross attraction of heterospecifics.
Species in other beetle families also were captured in significant numbers during this project, including other xylophagous species (scolytids, bostrichids, buprestids) and predators of xylophagous species (clerids). Data for those groups will be summarized in a separate manuscript.
Materials and methods
Field bioassays were conducted during 2009–2011 by employees of the Pennsylvania Department of Agriculture as part of the USDA Cooperative Agricultural Pest Survey (CAPS) program. Unless stated otherwise, beetles were trapped with 12-funnel Lindgren traps (Contech Enterprises Inc., Victoria, British Columbia, Canada) that were suspended from branches of trees (<10 m above the ground). Trap basins contained propylene glycol as a killing solution. Traps used in 2010 and 2011 had their funnels coated with Fluon® (Thermo-Fisher Scientific, Waltham, MA) to improve trapping efficiency (Graham et al. 2010). Lure blends usually were formulated to contain 25 mg of each isomer per ml of solution, with the carrier being 95 % ethanol (in 2009, 2010) or 100 % isopropanol (2011). Emitters were clear polyethylene sachets (press-seal bags, Bagette model 14770, 5.1 cm × 7.6 cm, 0.05 mm wall thickness, Cousin Corp., Largo, FL). A single trap line was set up at each study site, with traps positioned at least 25 m apart. Traps were serviced at ~2-week intervals, at which time pheromone lures were replaced. Taxonomy follows Monné and Bezark (2012). The gender of beetles was not recorded because the pheromones and attractants that we used generally attract adults of both sexes in similar numbers (e.g., see Silk et al. 2007; Lacey et al. 2009; Teale et al. 2011).
The 2009 experiment tested attraction of beetles to racemic 3-hydroxyhexan-2-one (hereafter 3R*-ketone, synthesized as described in Imrei et al. 2012), 2,3-hexanediol as a mixture of all four stereoisomers (hereafter 2,3-diol; 36:64 [2R*,3R*]:[2R*,3S*], synthesized as described in Hanks et al. 2007), and the ~1:1 blend of (E/Z)-fuscumol and its acetate (hereafter fuscumol + acetate, both 1:1.25 mixtures of racemic (Z)- and (E)-isomers, synthesized as in Mitchell et al. 2011). The bioassay was replicated in 25 counties of Pennsylvania that were dominated by hardwoods (Fig. 1).

Location of study sites in Pennsylvania during field seasons in 2009 (squares), 2010 (circles), and 2011 (stars)
In 2010, we tested the following seven treatments at study sites that were wooded with hardwoods and pines in ten counties (Fig. 1): (1) 3R*-ketone, (2) (2R*,3R*)-2,3-hexanediol (R*R*-diol, synthesized as described in Lacey et al. 2004), (3) (2R*,3S*)-hexanediol (R*S*-diol, Lacey et al. 2004), (4) the blend of 3R*-ketone plus the four hexanediol stereoisomers, (5) a ~1:1 blend of (E/Z)-fuscumol + fuscumol acetate, (6) monochamol (synthesized as in Teale et al. 2011), (7) the plant volatiles ethanol and α-pinene (ultra-high release lures; α-pinene ~92 % ee, Contech Inc., Victoria, BC, Canada). By comparing trap catch data for traps baited with the blend of hydroxyketone and diols with the corresponding data for the hydroxyketone and diols tested separately, we were able to evaluate positive and negative interactions. Plant volatile lures were replaced at monthly intervals. Test treatments also included a fermenting sugar bait (~4 l plastic jug with a ~10-cm diameter hole at the top of one side, containing 0.23 kg brown sugar in ~2 l of water that was allowed to ferment). Because the latter treatment involved a different trap design, and attracted few cerambycid beetles (see Online Resource 1), it was not included in the overall analysis of treatment effects.
The 2011 experiment tested attraction of beetles to a blend of pheromones of diverse cerambycid species that had been developed as a multi-species lure (see Hanks et al. 2012), with and without host plant volatiles (ethanol, α- and β-pinene). The experiment was replicated at study sites in 12 counties (Fig. 1). The multi-species pheromone blend contained 3R*-ketone (50 mg/lure), monochamol (25 mg), (E)-fuscumol + fuscumol acetate (100 mg each), all from Bedoukian Research (Danbury, CN), R*R*-diol (50 mg; Lacey et al. 2004), and racemic 2-methylbutan-1-ol (120 mg; Sigma-Aldrich, St. Louis, MO), dissolved in 1 ml of isopropanol. Lures for host plant volatiles were loaded with α- and β-pinene (ultra-high release, 2.3:1, 62 and 98 % ee, respectively; Product name: P339 Sirex lure, Chemtica Internacional, Santo Domingo, Costa Rica), and high-release ethanol (100 %; Contech Enterprises, Inc.), and were hung alongside pheromone lures for treatments that included both.
For each experiment, we compared the utility of pheromone lures as probes for assessing species diversity of cerambycid beetles in terms of the total number of cerambycid species that were captured during entire seasons per study site, and the number of beetles captured per species (averaged across species within study sites). Differences between treatment means (with study sites as replicates) were tested with the nonparametric Friedman’s Test (PROC FREQ, option CMH; SAS Institute 2001). Similar analyses were conducted separately for the dominant subfamilies (Cerambycinae, Lamiinae, Spondylidinae, Lepturinae) to assess how treatment effects varied with taxonomy. Differences between pairs of means were tested with the REGWQ means-separation test, which controls for maximum experiment-wise error rates (PROC GLM; SAS Institute 2001). The same statistical approach was used to test treatment effects on mean numbers of beetles captured for individual species, separately for each year, blocked by study site and collection period. For the latter analyses, we excluded data for site and date replicates that had fewer than a threshold number of specimens (N = 2–15, depending on the total number captured) that had been calculated so as to optimize sample size per replicate while maintaining sufficient replication for a robust analysis (N > 8 replicates included in each analysis).
We also present data on the flight periods of all species, based on ordinal dates of capture, to facilitate discussion of how species with similar pheromones might avoid cross attraction, and as a contribution to the literature on their biology. For each species, and for each year, we calculated the mean and standard deviation of ordinal dates of capture. The yearly means then were averaged, and another standard deviation was calculated to provide the best estimate of the mean activity period for each species across years. We also averaged the three yearly standard deviations as an estimate of how data were distributed around that grand mean.
In 2011, we conducted a follow-up experiment in Illinois to compare attraction of the cerambycine Megacyllene caryae (Gahan) to R*R*- and R*S*-diols (see “Results”). We used black flight intercept panel traps (corrugated plastic, 1.2 m high × 0.3 m wide, Alpha Scents Inc., West Linn, OR) that were treated with Fluon® and suspended from frames of PVC irrigation pipe (for details, see Graham et al. 2010). Trap basins contained propylene glycol as a killing agent. Lures were plastic sachets, as described above, loaded with 3R*-ketone, R*R*-diol, or R*S*-diol, with one replicate per site. The experiment was conducted during 29 April–3 June 2011 (average daily temperature 17.8 °C, 3.8 mm precipitation; Weather Underground, Inc., Ann Arbor, MI) at three sites in east-central Illinois that were wooded primarily with hardwoods: Allerton Park (Piatt Co.; 39.985342, −88.650147; property of the University of Illinois); Trelease Woods (Champaign Co.; 40.134873, −88.142796; property of the University of Illinois); and Forest Glen Reserve (Vermilion Co.; 40.01516, −87.56771; a county nature preserve). Traps were positioned 10 m apart in linear transects and checked for beetles every 1–3 days, at which time treatments were rotated within transects (by moving entire traps) to control for location effects. Differences between treatment means were tested using the Friedman’s test followed by the REGWQ means-separation test.
Results
During the 3 years of bioassays, we captured a total of 15,438 cerambycid beetles of 134 species (Table 1), including 44 species in 13 tribes of the subfamily Cerambycinae, 50 species in eight tribes of the Lamiinae, 30 species in two tribes of the Lepturinae, one species in each of the Necydaliniae and Parandrinae, two species in the Prioninae, and four species in the Spondylidinae. Also included is a single specimen of a species in the Disteniidae. Two of the captured cerambycid species were exotic: the invasive Callidium rufipenne Motschulsky, native to Asia (Maier 2007), and Phymatodes testaceus (L.), native to Europe (Swift and Ray 2010). Pheromones and other semiochemicals had previously been identified for some of these species, or their identifications were confirmed while this study was in progress (Table 2).
Of all the cerambycid beetles that were captured (Table 1), 55 species were represented by 1–5 specimens, 27 species were represented by 6–20 specimens, and 52 species were caught in numbers greater than 20 individuals. The most common species was the cerambycine Neoclytus a. acuminatus (F.) which was represented by 2,656 specimens. We present the entire data set in a supplement (Online Resource 1) because even low numbers of a species may provide leads or indications as to pheromone chemistry.
Activity periods of cerambycid species began in mid April with the cerambycines Neoclytus caprea (Say) and Stenosphenus notatus (Olivier), the lamiine Leptostylus asperatus (Say), and the lepturine Rhagium inquisitor inquisitor F. Cerambycine species that were best represented in the data set (i.e., with >100 captured in at least 2 years; see Table 1) varied considerably in the duration of their flight periods (Table 3), ranging from a few weeks for Phymatodes amoenus (Say), to 2 months for N. a. acuminatus and Xylotrechus colonus (F.). Many of the lamiine species were active during late June to late July, and the lengths of their flight periods were more uniform than those of the cerambycines (generally ~3 weeks; Table 3). Among the lepturines, only R. i. inquisitor was captured in sufficient numbers for analysis (Table 3). Species in the Spondylidinae emerged in sequence during June, but with considerable overlap among species (Table 3).
During 2009, traps in the 25 counties captured a total of 781 cerambycid beetles (range per county 5–67 beetles, mean 31.3 ± 18.2) of 79 species (range 5–36 species, mean 15.9 ± 9.2). These are relatively small numbers of beetles compared to the subsequent 2 years (see Table 1). Based on an earlier study that showed that trap catches could be increased more than tenfold by treatment of trap surfaces with Fluon® (Graham et al. 2010), it seems likely that many beetles that were attracted in 2009 were not captured. Experimental treatments did not differ significantly in the mean number of cerambycid species that they captured per site during the entire 2009 season (Fig. 2a), but significantly more cerambycine species were captured in traps baited with 3R*-ketone and 2,3-diols than with the fuscumol + acetate treatment. Conversely, the latter treatment captured the greatest number of lamiine and spondylidine species (Fig. 2a). For lepturines, there were no clear preferences for any of the three treatments (means 0.6–1.0 species per site; P > 0.05).

Top mean (±1 SE) number of cerambycid species of four subfamilies (from Table 1) per site that were captured by traps baited with different pheromones during the 2009 season. Chemical abbreviations as in Table 4. Means not significantly different for All Species (Friedman’s Q 2,75 = 0.74, P = 0.7), but significantly different for Cerambycinae (Q 2,75 = 14.0, P = 0.0009), Lamiinae (Q 2,74 = 6.04, P = 0.049), and Spondylidinae (Q 2,75 = 6.2, P = 0.044). Bottom mean (±SE) number of specimens per cerambycid species and site captured during 2009. Means significantly different for All Species (Q 2,73 = 19.8, P < 0.0001) and Cerambycinae (Q 2,67 = 29.9, P < 0.0001), but not for Lamiinae (Q 2,43 = 5.2, P = 0.051). Means within subfamilies with the same letters are not significantly different (REGWQ means-separation test) at P < 0.05
Considering the numbers of beetles that were captured per species in 2009 (Fig. 2b), traps baited with 3R*-ketone captured the greatest number of cerambycids in general, and cerambycines in particular. Treatment means for the lamiines were not significantly different from one another (Fig. 2b). Too few lepturine and spondylidine specimens had been captured to test treatment effects on numbers of specimens per species.
During 2010, traps in the ten counties captured a total of 7,709 cerambycid beetles (range per county 141–1,777 beetles, mean 771 ± 521) of 111 species (range 27–55 species, mean 41.2 ± 8.6). The trends seen in 2009 were repeated to some extent. Thus, the cerambycines were most strongly attracted to 3R*-ketone and the diols, whereas lamiines were most strongly attracted to fuscumol + acetate (Fig. 3a). For the few spondylidines, the plant volatile treatment attracted the greatest number of species (Fig. 3a). Treatment means for lepturines were not significantly different (means 0.5–1.1 species per site; P > 0.05).

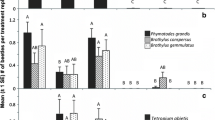
Top mean (±1 SE) number of cerambycid species of four subfamilies (from Table 1) per site that were captured by traps baited with different pheromones during the 2010 season. Chemical abbreviations as in Table 5. Means not significantly different for All Species (Friedman’s Q 6,70 = 12.0, P < 0.061), but significantly different for Cerambycinae (Q 6,70 = 33.9, P < 0.0001), Lamiinae (Q 6,70 = 19.5, P = 0.0035), and Spondylidinae (Q 6,70 = 17.8, P = 0.0067). Bottom mean (±SE) number of specimens per cerambycid species and site captured during 2010. Means significantly different for All Species (Q 6,70 = 32.7, P < 0.0001), Cerambycinae (Q 6,70 = 34.6, P < 0.0001), and Lamiinae (Q 6,69 = 14.7, P 0.022). Means within subfamilies with the same letters are not significantly different (REGWQ means-separation test) at P < 0.05
In 2010, R*R*-diol attracted the greatest number of beetles per species, particularly for the cerambycines (Fig. 3b), due primarily to the large number of N. a. acuminatus that were captured (Table 1, and see below). Lamiines were most abundant in the fuscumol + acetate treatment (Fig. 3b). Sample sizes for lepturines and spondylidines again were too low to compare treatments.
During the 2011 field season, traps in the 12 counties captured a total of 6,953 cerambycid beetles (range per county 151–969 beetles, mean 579 ± 241) of 102 species (range 29–53 species, mean 36.8 ± 2.5). The six-component blend of pheromones, with or without the plant volatiles ethanol and α- and β-pinene, attracted the greatest number of cerambycid species overall (Fig. 4a). The pheromone blend attracted the greatest numbers of cerambycines, and with and without plant volatiles also attracted the greatest number of lamiines (Fig. 4a). Spondylidine species were most numerous in the two treatments that included host plant volatiles (Fig. 4a). The number of lepturine species was not influenced by treatment (means 0.75–1.8 species per site; P > 0.05).

Top mean (±1 SE) number of cerambycid species of four subfamilies (from Table 1) per site that were captured by traps baited with different attractant blends during the 2011 season. Chemical abbreviations as in Table 6. Means significantly different for All Species (Friedman’s Q 4,60 = 20.1, P = 0.0005), Cerambycinae (Q 4,60 = 18.3, P 0.0011), Lamiinae (Q 4,60 = 27.1, P < 0.0001), and Spondylidinae (Q 4,60 = 30.1, P < 0.0001). Bottom mean (±SE) number of specimens per cerambycid species and site captured during 2011. Means significantly different for All Species (Q 4,60 = 18.2, P = 0.0011), Cerambycinae (Q 4,59 = 20.7, P = 0.0004), Lamiinae (Q 4,59 = 19.2, P = 0.0007), and Spondylidinae (ethanol and isopropanol treatments not included because sample sizes too low; Q 2,27 = 14.4, P = 0.0007). Means within subfamilies with the same letters are not significantly different (REGWQ means-separation test) at P < 0.05
During 2011, the two treatments containing the cerambycid pheromone blend (with and without plant volatiles) attracted significantly more individual beetles per species than any of the other treatments (Fig. 4b). Addition of plant volatiles did not affect the number of beetles per species captured overall. Broken down by subfamily, however, cerambycines were most strongly attracted by the pheromone blend alone. In contrast, addition of host plant volatiles to traps baited with the pheromone blend was critically important for the lamiines, with more beetles per species being attracted by that combination than by any other treatment. A similar trend was seen in the spondylidines, with the greatest number of beetles per species being attracted by the combination of the pheromone blend plus plant volatiles (Fig. 4b; ethanol and isopropanol treatments not included due to low sample size). The significant blend plus plant volatile treatment for the spondylidines was due primarily to attraction of Asemum striatum (L.) (753 individuals; Table 1, and see below). Too few lepturine specimens had been captured to test treatment effects on numbers of specimens per species.
Considering now the response of individual species to experimental treatments during 2009, eight species showed significant treatment effects (Table 4), and those effects were clearly in accord with the known semiochemistry for the cerambycines N. a. acuminatus, Neoclytus m. mucronatus (F.), Sarosesthes fulminans (F.), and X. colonus, and the lamiine Astyleiopus variegatus (Haldeman) (see Table 2). Attraction of N. a. acuminatus is inhibited by one or both of the unnatural diastereomers of its pheromone (Lacey et al. 2004; Hanks et al. 2012; Wong et al. 2012). However, the significant mean for the 2,3-hexanediol treatment in the present study (Table 4) suggests that this inhibition did not entirely preclude attraction to the blend of all four stereoisomers because this blend was not out-competed by traps baited with the highly attractive R*R*-diol (see Hanks et al. 2012). The present study also provides the first field data to confirm attraction of S. fulminans to its dominant pheromone component, 3R*-ketone (Lacey et al. 2009). The cerambycine P. testaceus also was significantly and specifically attracted to 3R*-ketone, even though males of that species apparently do not produce this compound (Table 2).
Attraction of the spondylidine Tetropium cinnamopterum (Kirby in Richardson) to fuscumol + acetate was unexpected because previous work had concluded that the pheromone (S)-fuscumol, one of the two enantiomers in the racemic fuscumol used in the present study, must be paired with host plant volatiles to attract this species (Sweeney et al. 2010). Significant attraction in the present study again may have been due to lack of competition among trap treatments, because no traps in the vicinity were baited with the more attractive combination of pheromone plus host plant volatiles. Attraction of the congener Tetropium schwarzianum Casey to fuscumol + acetate was the first indirect evidence, to our knowledge, that males of this species also produce one or both of these compounds as part of their pheromone blend.
The 2010 data set (Table 5) contains new information about the likely pheromone chemistry of the cerambycines Anelaphus parallelus (Newman) and Anelaphus villosus (F.) (both attracted by R*R*-diol; see Table 2). Other cerambycine species for which 3R-ketone was known to be the primary or sole pheromone component were indeed most strongly attracted to that compound or the pheromone blend that contained it (Tables 2, 5), including Cyrtophorus verrucosus (Olivier), N. caprea, N. m. mucronatus, Phymatodes aereus (Newman), and X. colonus. P. testaceus was significantly attracted by the ketone-diol blend, again despite not producing any of the compounds in that blend (Table 2). However, deployment of 3R*-ketone as a single component and in a blend with 2,3-hexanediols showed that N. m. mucronatus and X. colonus apparently were inhibited by 2,3-diols, with lower numbers captured by traps baited with the blend than in traps baited with 3R*-ketone alone (Tables 2, 5). Because previous research has confirmed that R*R*-diol is part of the pheromone of X. colonus, and enhances attraction to the ketone, and that (2S,3R)-2,3-hexanediol is not inhibitory (Lacey et al. 2009), these data implicate (2R,3S)-2,3-hexanediol as the inhibitory stereoisomer.
As expected, N. a. acuminatus was strongly attracted to R*R*-diol, whereas few individuals of this species were attracted to the blend of all four 2,3-hexanediol stereoisomers plus 3R*-ketone (Table 5), presumably due to inhibition by the unnatural diastereomer, as previously noted, and by 3R*-ketone (Hanks et al. 2012). In contrast, Curius dentatus Newman was attracted in similar numbers to both that same blend and R*S*-diol, as reported by Lacey et al. (2004), suggesting that there was no interference by the diastereomeric R*R*-diol or by 3R*-ketone.
The 2010 results for S. fulminans showed it to be significantly attracted only to R*S*-diol, which contained (2S,3R)-2,3-hexanediol, a minor component of its pheromone (Table 2). The lack of attraction to 3R*-ketone alone, to which it had been attracted during the 2009 experiment (Table 5), apparently was due to that treatment being out-competed by the R*S*-diol treatment. Furthermore, the fact that the blend of 3R*-ketone plus all four 2,3-hexanediol stereoisomers was not significantly attractive suggests that attraction was inhibited by one or both of the R*R*-diol enantiomers.
The 2010 data yielded several other informative results. For example, Obrium maculatum (Olivier) was significantly attracted to fuscumol acetate (Table 5), confirming results from an earlier experiment conducted in central Texas (Mitchell et al. 2011). Attraction of the pine-feeding Xylotrechus s. sagittatus (Germar) to ethanol and α-pinene is consistent with earlier research that evaluated attraction of wood-boring insects to these host compounds (Table 2). Only one cerambycine, Eburia quadrigeminata (L.), was attracted by the fermenting sugar bait traps in significant numbers (2.6 ± 1.0 beetles per trap, compared to <0.4 beetles per trap for other treatments; Friedman’s Q 7,70 = 20.6, P < 0.001). The lack of significant attraction of other species is surprising given that such baits have long been considered an effective attractant for cerambycid species whose hosts are deciduous trees (e.g., Linsley 1961).
An unexpected finding in the 2010 data set was strong attraction of M. caryae to R*R*-diol (Table 5), because studies conducted in east-central Illinois had concluded that males of that species produce both RS- and SR-diols as minor pheromone components that were not active by themselves, but which enhanced attraction to the main pheromone components, geranial and neral (Lacey et al. 2008). Furthermore, the fact that M. caryae was not attracted to the blend of 3R*-ketone and 2,3-hexanediols suggests that 3R*-ketone inhibited the response. Our follow-up field bioassay in Illinois during 2011 confirmed that adult M. caryae indeed were attracted by racemic R*R*-diol, but not R*S*-diol. The mean for the R*R*-diol treatment (4.5 ± 1.3 beetles per sample period), was significantly greater than means for the remaining treatments (0.38 ± 0.2, 1.0 ± 0.4, and 0.13 ± 0.1 for R*S*-diol, 3R*-ketone, and control treatments, respectively; N = 60 beetles; Friedman’s Q 3,32 = 10.2, P < 0.001).
Among the lamiines captured in 2010, most species in the tribes Acanthocinini and Acanthoderini already were known to be attracted to fuscumol and/or fuscumol acetate (Tables 2, 5), but the present study was the first evidence of attraction of Acanthoderes quadrigibba (Say), Astylopsis macula (Say), and Lepturges confluens (Haldeman). Astylopsis sexguttata (Say) was significantly attracted only to the plant volatiles ethanol plus α-pinene, as reported previously (Table 2) and consistent with its being a conifer specialist (see Lingafelter 2007). Of the two Monochamus species, M. carolinensis (Olivier) was attracted only by the ethanol plus α-pinene treatment, whereas M. s. scutellatus (Say) was attracted to monochamol (Table 5). Recent studies have revealed that both species are strongly attracted to monochamol only when it is released along with plant volatiles (Allison et al. 2012; Hanks et al. 2012; Macias-Samano et al. 2012), a treatment that was not included in the 2010 experiment. Unexpected findings included attraction of the lamiine Dorcaschema alternatum (Say) (Dorcaschematini) specifically to R*S*-diol, suggesting that attraction to the blend was inhibited by 3R*-ketone and/or R*R*-diol (Tables 2, 5). Attraction of lamiine species to hydroxyketone or the related 2,3-hexanediol type pheromones apparently is uncommon, the only other report to our knowledge being evidence that Graphisuris fasciatus (Degeer) is attracted by R*R*-diol (Hanks et al. 2012). Consistent with that finding, traps baited with R*R*-diol in the present study captured G. fasciatus in numbers second only to the fuscumol + acetate treatment (Table 5).
The one lepturine species that was captured in significant numbers during the 2010 studies, R. i. inquisitor, and the spondylidine A. striatum, were both attracted to ethanol plus α-pinene in the 2010 experiment, in agreement with earlier studies (Table 2), and consistent with their being conifer specialists. Attraction of the spondylidine T. schwarzianum to fuscumol + acetate confirms the findings of the 2009 experiment.
Many of the most prevalent species in the 2011 data set had also been prevalent in 2010 (Tables 5, 6). The results again agreed with what was known of the semiochemistry of many species (Tables 2, 6), but added another two cerambycine species that produce 3R*-ketone as a dominant pheromone component, P. amoenus and P. varius (F.). The high-release rate ethanol lure was the most attractive treatment in 2011 only for the cerambycine A. villosus (significantly attracted by R*R*-diol in 2010). Despite the fact that R*R*-diol was present in the 2011 pheromone blend, few A. villosus were attracted to that treatment, suggesting either that the high release rate ethanol treatments were more attractive than the pheromones, or that certain components in the blend were inhibiting responses to the pheromone components.
The influence of host plant volatiles (ethanol plus α- and β-pinene) on attraction of beetles to the pheromone blend varied with species, and the direction of the effect usually was inconsistent even among closely related species (Table 6). For example, plant volatiles inhibited attraction of P. testaceus and P. varius (F.) (tribe Callidiini) to the pheromone blend, but among species in the tribe Clytini they inhibited attraction of M. caryae and N. caprea, but had no apparent effect on N. a. acuminatus, N. m. mucronatus, and X. colonus. A recent study by Hanks et al. (2012), conducted in east-central Illinois, reported similar patterns in the influence of plant volatiles on attraction by the same pheromone blend for the species that were in common with the present study, N. a. acuminatus, N. m. mucronatus, and X. colonus. Xylotrechus s. sagittatus was not at all attracted to the pheromone blend, but significantly attracted by plant volatiles, as was the case in 2010 (Table 5).
The plant volatiles ethanol plus α- and β-pinene had similar variable effects among lamiines in the tribes Acanthocinini and Acanthoderini (Table 6). Thus, they appeared to inhibit attraction of A. quadrigibba and Sternidius alpha (Say) to fuscumol and/or fuscumol acetate in the blend, while enhancing attraction of the congeners Lepturges angulatus (LeConte) and L. confluens, but having no significant effect on Aegomorphus modestus (Gyllenhal in Schoenherr), A. variegatus, G. fasciatus, and Styloleptus biustus (LeConte). Also, only the combination of plant volatiles and monochamol (in the blend), rather than either alone, attracted Monochamus carolinensis, M. notatus (Drury), and M. s. scutellatus. Similarly, the combination of plant volatiles and fuscumol, rather than either alone, was required to attract the spondylidine species A. striatum, T. cinnamopterum, and T. schwarzianum (Table 6). The lepturine R. i. inquisitor was strongly attracted to the plant volatiles, and in fact the blend of pheromone components apparently inhibited attraction (Table 6). These results again were congruent with those of Hanks et al. (2012) from field trials in Illinois for the lamiine species that were common to both studies (A. variegatus, G. fasciatus, L. angulatus, and M. carolinensis).
During the 3-year study, several species were captured in relatively large numbers, but nevertheless showed no significant treatment effects (see Online Resource 1). In many cases, statistical power was weak because specimens were distributed across study sites and dates to the extent that sample size per replicate was low (e.g., P. amoenus in 2010, Euderces pini [Olivier] in 2011). However, the lack of significant treatment effects for the cerambycine Elaphidion mucronatum (Say), the parandrine Neandra brunnea (F.), and the prionine Orthosoma brunneum (Forster) is consistent with our earlier findings that adults of those species often are intercepted by traps, regardless of how traps are baited (unpublished data). The same situation is likely the case for the abundant lepturine Gaurotes cyanipennis (Say) (Online Resource 1).
Discussion
Our results from three consecutive years of field trials, in which small numbers of semiochemical compounds or combinations (3 in 2009, 6 in 2010, 3 in 2011) attracted numerous cerambycid species, strongly reinforced the notion that pheromone structures are highly conserved within and among at least some of the cerambycid subfamilies. For example, the 3-hydroxyhexan-2-ones and 2,3-hexanediols are important pheromone components for cerambycines, whereas fuscumol and/or fuscumol acetate play a similar role for lamiines in the tribes Acanthocinini and Acanthoderini, but perhaps not for species in other tribes (see Fonseca et al. 2010; Mitchell et al. 2011; Hanks et al. 2012). Capture of the spondylidine species A. striatum and T. schwarzianum provides further evidence that fuscumol and/or fuscumol acetate also serve as pheromones for this subfamily as well.
Exceptions to these general trends included attraction of the cerambycine O. maculatum to fuscumol + acetate, and of the lamiine D. alternatum to R*S*-diol. We have yet to confirm that those species actually produce the compounds to which they were attracted. For D. alternatum, inhibition of the R*S*-diol by 3R*-ketone and/or R*R*-diol may serve to prevent cross attraction to male X. colonus, which produces both of the latter compounds and broadly overlaps in seasonal activity with D. alternatum. At first glance, attraction of P. testaceus to 3R*-ketone in 2009, and that compound blended with 2,3-hexanediols in 2010, seems anomalous because this species does not appear to produce either of these compounds (Table 2). However, both sexes of the western congener P. grandis Casey also are attracted by (R)- and (S)-3-hydroxyhexan-2-ones, but males produced only (R)-2-methylbutan-1-ol as their pheromone (Hanks et al. 2007; species formerly known as P. lecontei, Swift and Ray 2010). As is the case with P. testaceus, P. grandis overlaps in distribution and seasonal phenology with a complex of cerambycine species that use 3R- and 3S-ketones as their dominant or sole pheromone components (Hanks et al. 2007). Also unexpected was attraction of M. caryae to racemic R*R*-diol, but these results were confirmed with the follow-up study in Illinois. These species may eavesdrop on the pheromone communication of other guild members, which may serve as an efficient method of finding suitable hosts for mating and oviposition.
Despite our limited understanding of the pheromones used by the full panoply of species of the subfamily Cerambycinae that were trapped during these studies, patterns are beginning to emerge that may explain how cross attraction is avoided despite the extensive parsimony in pheromone components of many sympatric and synchronic species (see Table 2). For example, the following scenarios could explain how cross attraction is averted among species for which 3R*-ketone is a key pheromone component (species in order of their appearance over the season; Table 3; see Table 2 for summary information about semiochemistry):
-
1.
N. caprea is temporally isolated from other species that produce 3R*-ketone;
-
2.
Attraction of both C. verrucosus and P. amoenus to 3R*-ketone is enhanced by the components 2-nonanone and (R)-2-methylbutan-1-ol, respectively (unpublished data), reducing attraction of these species to other species that produce only the ketone;
-
3.
S. fulminans produces 3R*-ketone and R*S*-diol, and although it is attracted to 3R*-ketone alone when no other attractants are present, it responds to the diol much more strongly and thus would be unlikely to respond to heterospecific males that produce only 3R*-ketone, such as X. colonus;
-
4.
Attraction of X. colonus to 3R*-ketone is enhanced by R*R*-diol, but inhibited by R*S*-diol, preventing attraction to the 3R*-ketone produced by male S. fulminans;
-
5.
N. mucronatus is active later in the season than many species that are attracted by 3R*-ketone, but because it is inhibited by diols, it may not respond to the blend of 3R*-ketone and R*R*-diol produced by male X. colonus. Similarly, it would appear that C. dentatus could potentially be attracted by R*S*-diol emitted by male S. fulminans, but in fact C. dentatus is active much later in the season. Although the lamiine D. alternatum also is attracted by R*S*-diol, it overlaps only briefly with C. dentatus, and attraction to the diols of male S. fulminans may be inhibited by the 3R*-ketone component.
It must be emphasized that the above scenarios are speculative, given our current limited understanding of the role of pheromone stereochemistry for the species involved. Further research will be essential to fully characterize the mechanisms that minimize cross attraction between heterospecifics and allow these species to coexist despite similarities in their pheromones. Nevertheless, because this study encompassed a large number of species, it clearly illustrates that, as with many other insect taxa, cerambycid beetles often use pheromone blends rather than individual pheromone chemicals, resulting in species-specific signals, in addition to having opportunities for spatial and temporal separation. We found good evidence that several species use blends of two or more components to achieve optimal attraction. Conversely, and equally important, there were examples of components of blends that specifically inhibited cross attraction between sympatric species. This evidence for the widespread use of blends also will help to inform full identification of pheromones for additional species. For example, further screening studies could be carried out with specifically formulated blends of likely pheromone components (e.g., 3R*-ketone plus R*R*-diol, or 3R*-ketone plus R*S*-diol, in various proportions).
Less clear are possible mechanisms that might serve to maintain reproductive isolation among species of the subfamily Lamiinae. Many lamiine species are now known to be attracted by fuscumol and/or its acetate (Table 2), although for most of these species, it has yet to be confirmed that either sex produces those compounds. Moreover, many of the lamiine species captured in this study overlapped broadly in seasonal phenology, raising the question of how species specificity is maintained if these species do indeed share pheromone components. The simultaneous attraction of several lamiine species in the genus Monochamus to monochamol or monochamol plus host volatiles (this study; Allison et al. 2012; Fierke et al. 2012; Macias-Samano et al. 2012) represents another example in which it is not yet clear how species specificity of the signal is maintained. The same question also remains for the spondylidines T. cinnamopterum and T. schwarzianum, both of which were attracted by fuscumol + acetate. For these species, cross attraction may be averted by additional mechanisms, such as differences in circadian activity period.
Our trap lures were particularly effective for trapping species in the subfamilies Cerambycinae, Lamiinae, and to a lesser extent Spondylidinae, but had limited or no effect as attractants for lepturines, prionines, and species in the minor subfamilies. These findings are consistent with the current state of knowledge (summarized in Table 2), given that the semiochemicals that we tested, alone and in combinations, were known to have broad activity as aggregation pheromones only for cerambycines, lamiines, and spondylidines. In contrast, female-produced sex pheromones recently have been identified for two lepturine species, cis-vaccenyl acetate for Ortholeptura valida (LeConte) (Ray et al. 2011), and (Z)-hexadec-9-en-4-olide for Desmocerus californicus californicus Horn (Ray et al. 2012a). However, in contrast to data sets reported here, in which individual compounds usually were attractive to a number of related species, the preliminary results for these two lepturine species suggest that their pheromones may be more species specific (JGM and A.M. Ray, unpublished data). Furthermore, the structures of these pheromones are entirely different from the male-produced pheromones currently known from cerambycines and lamiines (see Table 2). Consequently, there was no reason to suspect that lepturines would be strongly attracted to any of the pheromone lures deployed in the studies described here, and most of the lepturine species in the present data set probably were captured by random encounters with traps. Similarly, the small number of prionines that were trapped suggests that none of the compounds that were tested are semiochemicals for prionine species native to Pennsylvania. We certainly could have captured prionines in substantial numbers had we included 3,5-dimethyldodecanoic acid in field bioassays, because it is a powerful sex attractant pheromone for many Prionus species (Barbour et al. 2011), including species native to the northeastern United States (unpublished data; see Monné and Bezark).
Data from 2011 also demonstrated the effectiveness of the host plant volatiles α- and β-pinene, in some cases acting in concert with pheromones, for conifer specialists in several subfamilies (see Lingafelter 2007), including the cerambycine X. s. sagittatus, the lamiines M. carolinensis, M. notatus, and M. s. scutellatus, the lepturine R. i. inquisitor, and all of the spondylidine species. Ethanol alone attracted the cerambycine A. villosus, a hardwood feeder, consistent with earlier research (Table 2). Conversely, it was noteworthy that at least one of the host plant volatiles (probably α- and/or β-pinene) strongly inhibited attraction of a number of hardwood feeders, as might be expected given their host requirements (see Lingafelter 2007), including the cerambycines M. caryae, N. caprea, P. testaceus, and P. varius, and the lamiines A. quadrigibba and S. alpha (Table 6). This situation appears to be analogous to the well-documented repellent effect of angiosperm volatiles for conifer-feeding bark beetles (reviewed by Zhang and Schlyter 2004).
Previous field studies in Illinois had shown that adult E. quadrigeminata, the only species that was significantly attracted to fermenting sugar traps in Pennsylvania, were attracted by traps baited with diluted molasses that had fermented to the point that ethyl acetate was the dominant volatile (unpublished data). These findings suggest that this, and probably other cerambycid species, use fermentation products to find the dead and decaying trees that are their larval hosts (Craighead 1923).
These experiments reaffirmed the roles of semiochemicals as mediators of behavior for many cerambycid species that previously have been studied. In addition to the species that were attracted in large numbers, many other species were caught in smaller numbers, with no statistically significant treatment effects (Online Resource 1). It is entirely possible that attraction of some species may have been weak because their “true” pheromone consists of a blend of compounds, or is enhanced by specific host plant volatiles, or the attractive signal may be multimodal, consisting of a chemical signal in tandem with nonchemical signals.
It must be emphasized, however, that it would be risky, if not impossible, to draw conclusions about the relative biological activities of these semiochemicals based only on the numbers of species, or numbers of individual beetles that were captured, because we have no information about population densities of species at our study sites. Thus, beetles of some species may have been strongly attracted to certain lures, even though statistical analyses identified no significant treatment effects, because those species were rare. In such cases, even weak bioassay data may provide valuable insights into pheromone chemistry, and may be useful in guiding future research to identify pheromones. In support of this notion, the weak 2009 data suggesting attraction of P. aereus to 3R*-ketone, and A. quadrigibba, A. modestus, and S. alpha to fuscumol + acetate (Online Resource 1; treatment means not significant, P > 0.05), could indeed have provided an accurate prediction of compounds that were proven to be significant attractants in 2010 (Table 5). Similarly, mere indications of attraction of other species were consistent with known semiochemistry. For example, the greatest numbers of Neoclytus scutellaris (Olivier) were attracted to the 3R*-ketone treatment in 2009 and 2010, and 3R-ketone is indeed the sole pheromone component of that species (unpublished data). Similarly, most specimens of Astylidius parvus Casey were attracted to the fuscumol + acetate treatment in 2009 and 2010, and fuscumol was documented as an attractant for this species in an earlier study (Mitchell et al. 2011). In fact, given that fuscumol and/or its acetate apparently are common attractants of lamiine species (Mitchell et al. 2011), it is likely that even weak indications of attraction to those compounds in the present study (Online Resource 1) will prove to be accurate predictors of the pheromone chemistry for species such as Ecyrus d. dasycerus (Say), Lepturges pictus (LeConte), Lepturges symmetricus (Haldeman), and Sternidius misellus (LeConte).
In summary, this study has unequivocally demonstrated that the use of volatile semiochemicals is widespread within the Cerambycidae, and that these semiochemicals can be powerful attractants. Traps baited with pheromones and related attractants have proven very effective in quarantine monitoring for other taxa of invasive insects, and often are key components of eradication efforts once such species have colonized (e.g., Brockerhoff et al. 2006a). Based on the large numbers and species diversity of beetles that were captured during this 3-year study, we suggest that traps baited with semiochemicals could be used in analogous fashion to provide a sensitive and straightforward surveillance method for cerambycids, thereby rectifying a serious gap in the ability of regulatory agencies to effectively detect and monitor these insects.
References
Allison JD, Borden JH, McIntosh RL, de Groot P, Gries R (2001) Kairomonal response by four Monochamus species (Coleoptera: Cerambycidae) to bark beetle pheromones. J Chem Ecol 27:633–646
Allison JD, McKenna JL, Millar JG, McElfresh JS, Mitchell RF, Hanks LH (2012) Response of the woodborers Monochamus carolinensis and Monochamus titillator to known cerambycid pheromones in the presence and absence of the host plant volatile alpha-pinene. Environ Entomol (in press)
Barbour JD, Millar JG, Rodstein J, Ray AMB, Alston DG, Rejzek M, Dutcher JD, Hanks LM (2011) Synthetic 3,5-dimethyldodecanoic acid serves as a general attractant for multiple species of Prionus (Coleoptera: Cerambycidae). Ann Entomol Soc Am 104:588–593
Brockerhoff EG, Liebhold AM, Jactel H (2006a) The ecology of forest insect invasions and advances in their management. Can J For Res 36:263–268
Brockerhoff EG, Jones DC, Kimberley MO, Suckling DM, Donaldson T (2006b) Nationwide survey for invasive wood-boring and bark beetles (Coleoptera) using traps baited with pheromones and kairomones. For Ecol Manag 228:234–240
Champlain AB, Knull JN (1932) Fermenting baits for trapping Elateridae and Cerambycidae (Coleop.). Entomol News 43:253–257
Chénier JVR, Philogène BJR (1989) Field responses of certain forest Coleoptera to conifer monoterpenes and ethanol. J Chem Ecol 15:1729–1745
Craighead FC (1923) North American cerambycid larvae: a classification and the biology of North American cerambycid larvae. Can Dep Agric Bull 27, New series (Technical)
Dodds KJ (2011) Effects of habitat type and trap placement on captures of bark (Coleoptera: Scolytidae) and longhorned (Coleoptera: Cerambycidae) beetles in semiochemical-baited traps. J Econ Entomol 104:879–888
Dunn JP, Potter DA (1991) Synergistic effects of oak volatiles with ethanol in the capture of saprophagous wood borers. J Entomol Sci 26:425–429
Fierke MK, Skabeikis DD, Millar JG, Teale SA, McElfresh JS, Hanks LM (2012) Identification of a male-produced pheromone for Monochamus scutellatus scutellatus and Monochamus notatus (Coleoptera: Cerambycidae). J Econ Entomol (in press)
Fonseca MG, Vidal DM, Zarbin PHG (2010) Male-produced sex pheromone of the cerambycid beetle Hedypathes betulinus: chemical identification and biological activity. J Chem Ecol 36:1132–1139
Frost SW, Dietrich H (1929) Coleoptera taken from bait-traps. Ann Entomol Soc Am 22:427–436
Ginzel MD, Hanks LM (2005) Role of host plant volatiles in mate location for three species of longhorned beetles. J Chem Ecol 31:213–217
Graham EE, Mitchell RF, Reagel PF, Barbour JD, Millar JG, Hanks LM (2010) Treating panel traps with a fluoropolymer enhances their efficiency in capturing cerambycid beetles. J Econ Entomol 103:641–647
Hanks LM, Millar JG, Moreira JA, Barbour JD, Lacey ES, McElfresh JS, Reuter FR, Ray AM (2007) Using generic pheromone lures to expedite identification of aggregation pheromones for the cerambycid beetles Xylotrechus nauticus, Phymatodes lecontei, and Neoclytus modestus modestus. J Chem Ecol 33:889–907
Hanks LM, Millar JG, Mongold-Diers JA, Wong JCH, Meier LR, Reagel PF, Mitchell RF (2012) Using blends of cerambycid beetle pheromones and host plant volatiles to simultaneously attract a diversity of cerambycid species. Can J For Res 42:1050–1059
Imrei Z, Millar JG, Janik G, Tóth M (2012) Field screening of known pheromone components of longhorned beetles in the subfamily Cerambycinae (Coleoptera: Cerambycidae) in Hungary. Z Naturforsch C (in press)
Lacey ES, Ginzel MD, Millar JG, Hanks LM (2004) Male-produced aggregation pheromone of the cerambycid beetle Neoclytus acuminatus acuminatus. J Chem Ecol 30:1493–1507
Lacey ES, Moreira JA, Millar JG, Ray AM, Hanks LM (2007) Male-produced aggregation pheromone of the cerambycid beetle Neoclytus mucronatus mucronatus. Entomol Exp Appl 122:171–179
Lacey ES, Moreira JA, Millar JG, Hanks LM (2008) A male-produced aggregation pheromone blend consisting of alkanediols, terpenoids, and an aromatic alcohol from the cerambycid beetle Megacyllene caryae. J Chem Ecol 34:408–417
Lacey ES, Millar JG, Moreira JA, Hanks LM (2009) Male-produced aggregation pheromones of the cerambycid beetles Xylotrechus colonus and Sarosesthes fulminans. J Chem Ecol 35:733–740
Lingafelter SW (2007) Illustrated key to the longhorned wood-boring beetles of the eastern United States. Coleopterists Society, North Potomac
Linsley EG (1961) The Cerambycidae of North America Part 1. Introduction. Univ Calif Publ Entomol 18:1–135
Macias-Samano JE, Wakerchuk D, Millar JG, Hanks LM (2012) 2-Undecyloxy-1-ethanol in combination with other semiochemicals attracts three Monochamus species (Coleoptera: Cerambycidae) in British Columbia. Can Entomol (in press)
Maier CT (2007) Distribution and hosts of Callidiellum rufipenne (Coleoptera: Cerambycidae), an Asian cedar borer established in the Eastern United States. J Econ Entomol 100:1291–1297
Millar JG, Hanks LM, Moreira JA, Barbour JD, Lacey ES (2009) Pheromone chemistry of cerambycid beetles. In: Nakamuta K, Millar JG (eds) Chemical ecology of wood-boring insects. Forestry and Forest Products Research Institute, Ibaraki, pp 52–79
Miller DR, Asaro C, Crowe CM, Duerr DA (2011) Bark beetle pheromones and pine volatiles: attractant kairomone lure blend for longhorn beetles (Cerambycidae) in pine stands of the southeastern United States. J Econ Entomol 104:1245–1257
Mitchell RF, Graham EE, Wong JCH, Reagel PF, Striman BL, Hughes GP, Paschen MA, Ginzel MD, Millar JG, Hanks LM (2011) Fuscumol and fuscumol acetate are general attractants for many species of cerambycid beetles in the subfamily Lamiinae. Entomol Exp Appl 141:71–77
Monné MA, Bezark LG (2012) Checklist of the Oxypeltidae, Vesperidae, Disteniidae and Cerambycidae, (Coleoptera) of the Western Hemisphere. Calif Dep Agric. http://plant.cdfa.ca.gov/byciddb/checklists/WestHemiCerambycidae2012.pdf Accessed 9 Sept 2012
Montgomery ME, Wargo PM (1983) Ethanol and other host-derived volatiles as attractants to beetles that bore into hardwoods. J Chem Ecol 9:181–190
Pajares JA, Álvarez G, Ibeas F, Gallego D, Hall DR, Farman DI (2010) Identification and field activity of a male-produced aggregation pheromone in the pine sawyer beetle, Monochamus galloprovincialis. J Chem Ecol 36:570–583
Phillips TW, Wilkening AJ, Atkinson TH, Nation JL, Wilkinson RC, Foltz JL (1988) Synergism of turpentine and ethanol as attractants for certain pine-infesting beetles (Coleoptera). Environ Entomol 17:456–462
Ray AM, Žunič A, Alten RL, McElfresh JS, Hanks LM, Millar JG (2011) cis-Vaccenyl acetate, a sex attractant pheromone of Ortholeptura valida, a longhorned beetle in the subfamily Lepturinae. J Chem Ecol 37:173–178
Ray A, Swift IP, McElfresh JS, Alten RL, Millar JG (2012a) (R)-Desmolactone, a female-produced sex pheromone component of the cerambycid beetle Desmocerus californicus californicus (subfamily Lepturinae. J Chem Ecol 38:157–167
Ray AM, Barbour JD, McElfresh JS, Moreira JA, Swift I, Wright IM, Žunič A, Mitchell RF, Graham EE, Alten RL, Millar JG, Hanks LM (2012b) 2,3-Hexanediols are female-produced sex attractant pheromones of cerambycid beetles in the prionine genus Tragosoma. J Chem Ecol (in press)
Rodstein J, Millar JG, Barbour JD, McElfresh JS, Wright IM, Barbour KS, Ray AM, Hanks LM (2011) Determination of the relative and absolute configurations of the female-produced sex pheromone of the cerambycid beetle Prionus californicus. J Chem Ecol 37:114–124
SAS Institute (2001) SAS/STAT user’s guide, release 9.2. SAS Institute Inc., Cary
Silk PJ, Sweeney J, Wu J, Price J, Gutowski JM, Kettela EG (2007) Evidence for a male-produced pheromone in Tetropium fuscum (F.) and Tetropium cinnamopterum (Kirby) (Coleoptera: Cerambycidae). Naturwissenschaften 94:697–701
Sweeney J, De Groot P, MacDonald L, Smith S, Cocquempot C, Kenis M, Gutowski JM (2004) Host volatile attractants and traps for detection of Tetropium fuscum (F.), Tetropium castaneum L., and other longhorned beetles (Coleoptera: Cerambycidae). Environ Entomol 33:844–854
Sweeney J, Gutowski JM, Price J, de Groot DE (2006) Effect of semiochemical release, killing agent, and trap design on detection of Tetropium fuscum (F.) and other longhorn beetles (Coleoptera: Cerambycidae). Environ Entomol 35:645–654
Sweeney J, Silk PJ, Gutowski JM, Wu J, Lemay MA, Mayo PD, Magee DI (2010) Effect of chirality, release rate, and host volatiles on response of Tetropium fuscum (F.), Tetropium cinnamopterum Kirby, and Tetropium castaneum (L.) to the aggregation pheromone, fuscumol. J Chem Ecol 36:1309–1321
Swift IP, Ray AM (2010) Nomenclatural changes in North American Phymatodes Mulsant (Coleoptera: Cerambycidae). Zootaxa 2448:35–52
Teale SA, Wickham JD, Zhang F, Su J, Chen Y, Xiao W, Hanks LM, Millar JG (2011) A male-produced aggregation pheromone of Monochamus alternatus (Coleoptera: Cerambycidae), a major vector of pine wood nematode. J Econ Entomol 104:1592–1598
Witzgall P, Kirsch P, Cork A (2010) Sex pheromones and their impact on pest management. J Chem Ecol 36:80–100
Wong JCH, Mitchell RF, Striman BL, Millar JG, Hanks LM (2012) Blending synthetic pheromones of cerambycid beetles to develop trap lures that simultaneously attract multiple species. J Econ Entomol 105:906–915
Zhang Q-H, Schlyter F (2004) Olfactory recognition and behavioural avoidance of angiosperm nonhost volatiles by conifer-inhabiting bark beetles. Agric For Entomol 6:1–19
Acknowledgments
We thank Sven-Erik Spichiger for organizing and overseeing the field bioassays and identification of specimens, and the collectors and taxonomists of the Pennsylvania Department of Agriculture for collecting the data. We also thank Robert F. Mitchell, Joseph C.H. Wong, and Peter F. Reagel for technical assistance with field work in Illinois, and for helpful comments on the manuscript by RFM, JCHW, and Ann M. Ray. This research was supported by grants from The Alphawood Foundation, the National Research Initiative of the United States Department of Agriculture Cooperative State Research, Education and Extension Service, grant #2009-35302-05047, and the USDA North-Central IPM Center, grant # AG 2007-51120-18445.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Hanks, L.M., Millar, J.G. Field bioassays of cerambycid pheromones reveal widespread parsimony of pheromone structures, enhancement by host plant volatiles, and antagonism by components from heterospecifics. Chemoecology 23, 21–44 (2013). https://doi.org/10.1007/s00049-012-0116-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00049-012-0116-8