
Abstract
The angiotensin I converting enzyme 2 (ACE2) is a key factor in the maintenance of intestinal homeostasis. Dysregulation of homeostasis can lead to inflammation of the colon (colitis), which can cause life-threatening enfeeblement or even cancer. Animal models are valuable surrogates in deciphering the pathology behind such human conditions and for screening of putative therapeutic targets or treatment paradigms. However, development of disease models can be time-consuming and technical demanding, which might hamper their application-value. In this study, we genetically disrupted the mouse Ace2 gene by direct injection of in vitro transcribed mRNA coding for transcription activator-like effector nucleases (TALENs) into the cytoplasm of outbred Kunming mouse zygotes. Consequently, somatic mutations were induced with an efficiency of 57 %, of which 39 % were frameshift mutations. Moreover, all modifications were stably transferred during germline transmission. In Ace2-knockout male mice (Ace2 −/y), we observed severe chemical induced colitis, characterized by considerable weight loss, diarrhea and a shortened colon length. Histologically, Ace2 mutations resulted in the infiltration of leukocytes and the overt damage of the intestinal mucosal barrier. In addition, we detected an increased expression of inflammatory cytokines in the colon tissue of Ace2 −/y mice. Collectively, the data indicate that high targeting efficiency and heritability can be achieved in an outbred mouse model by zygote injection of TALEN mRNA. Furthermore, the generated Ace2 −/y mice display phenotypic traits reminiscent of colitis and we anticipate that such mice can be of value in studies of the intestinal microbiome or fecal transplantation.
Similar content being viewed by others


Introduction
Gene targeting is an effective approach for deciphering gene function and for creating animal models of genetic disorders. Much of the work in this field has been conducted in inbred mouse strains by gen-specific insertion of selection markers by conventional homologous recombination (HR) in embryonic stem cells. Although widely appreciated the methodology can be labor intensive with modest efficiency and heritability. Furthermore, the retention of an exogenous selection marker and the genetic homogeneity of inbred strains may produce confounding effects. Thus, working with models of human conditions reproducible and significant findings in an outbred strain can be preferable as these are more likely to mimic human biology.
In recent years, several novel genome-editing technologies have emerged from which the four most studied methods count zinc-finger nucleases (ZFNs) (Kim et al. 1996), transcription activator-like effector nucleases (TALENs) (Sun and Zhao 2013), clustered regularly interspaced short palindromic repeat (CRISPR) (Wang et al. 2014) and engineered meganucleases (Epinat et al. 2003). Each technique has demonstrated highly efficient and locus-specific genome editing in numerous species. As to TALENs, their binding site is determined by a central domain composed of tandem 33–35 amino acid (aa) long repeats. In each repeat, the residues at positions 12 and 13, being the ‘repeat-viable di-residues’ (RVD), cipher the binding target (Cermak et al. 2011). Structurally, the catalytic domain of the FokI nuclease has been fused to the TALE protein at the C-terminal providing DNA double-strand breaking capabilities (Miller et al. 2011). Importantly, efficient TALEN-mediated gene targeting has been demonstrated across different animal orders such as in zebrafish (Bedell et al. 2012), mouse (Sung et al. 2013; Qiu et al. 2013), and human cells (Ding et al. 2013; Hockemeyer et al. 2011).
For years, the Renin-Angiotensin System (RAS) has been regarded one of the most important hormone systems regulating blood pressure and water balance, in which post-translational processing of Angiotensin (ANG) II and its derivatives play a key regulatory role (Ramirez-Sanchez et al. 2013). ANGII is the primary effector hormone and catalyzed by the angiotensin-converting enzyme (ACE) in lung capillaries and, conversely, inactivated by ACE2. The latter produces a C-terminal trimmed cleavage product of ANGII (ANG1-7) possessing inverse biological functions to ANGII. The balance of the two enzyme homologues controls the concentration of ANGII, and consequently regulates the RAS cascade (Guyton 1992).
The ACE2 protein is structurally homologous to the ACE at the N-terminal catalytic domain but with Collectin (a component of the innate immune system) at the C-terminal domain, sharing 41.8 and 48 % sequence identity, respectively (Donoghue et al. 2000; Zhang et al. 2001).
Interestingly, ACE2 deficiency has been associated with a more severe colitis pathophysiology in dextran sodium sulphate (DSS) challenged mice (Hashimoto et al. 2012). A RAS-pathway unrelated role in intestinal homeostasis has been suggested in which the uptake of the essential amino acid tryptophan through the luminal surface of the colon is dependent on ACE2. Tryptophan is required for the production of vitamin B3, which on the other hand regulates the expression of antimicrobial peptides through the mTOR pathway. As a consequence can ACE2 deficient mice not cope with the altered intestinal microbiome succeeding and insult and develop colitis. Thus, ACE2 appears to link amino acid malnutrition to intestinal microbial ecology and inflammation (Hashimoto et al. 2012; Izcue and Powrie 2012). Conflicting data, however, exists showing anti-inflammation effects of an ACE2 inhibitor (Byrnes et al. 2009).
A similar pattern of discrepancy is seen in knockout mouse lines intended for research in ACE2 effects on cardiovascular physiology where obvious differences in phenotypic traits were evident (Crackower et al. 2002; Yamamoto et al. 2006; Gurley et al. 2006). These mouse lines were generated by HR, which introduces exogenous cargo such as selection markers and vector backbone to the host genome that potentially could play a role in the divergent results (Brenner et al. 1993; Wu et al. 1998). Genetic background of the animals used and the zygote-stage at the time of genetic manipulation are other important factors and it therefore remains crucial to develop more appropriate genome modifying tools usable in genetically diverse animals.
In this study, we disrupted the murine Ace2 gene using microinjection of TALENs mRNA producing a mutation efficiency of 57 % that mainly consisted of small deletions. Importantly, TALEN-induced modifications were stably inherited vertically. We further found that ACE2-deficiency led to increased susceptibility to DSS induced colitis, enhanced histological damage in the colon and the up-regulated expression of the inflammatory cytokines IL1-β, IL6, IL10, IFN-γ, and TNF-α. Our results demonstrated that translation of in vitro transcribed TALEN mRNA microinjected into murine ES cells was highly effective and a convenient way to develop and explore a model of colitis.
Materials and methods
Ethics statement
Animal experiment procedures were approved by the Institutional Review Board on Bioethics and Biosafety of BGI.
Design and construction of TALENs vector
Candidate TALENs were identified using the online tool Tal Effector Nucleotide Targeter 2.0 (http://tale-nt.cac.cornell.edu/node/add/talen) (Doyle et al. 2012). Construction of a TALEN pair targeting exon 2 of the Ace2 locus (recognition sequence in bold) and encompassing a 16 bp spacer region containing a DraI restriction site (underlined) was pursued (5′-TCACCGAGGAAAATGCCAAGACATTTTTAAACAACTTTAATCAGGAAGCTGA-3′) (Fig. 1a). A spacer region shorter than 20 bp has previously been shown to be preferable to longer spacer regions in mouse ES cells (Qiu et al. 2013). RVD modules were assembled into the pTAL3 vector using the Golden Gate method (Cermak et al. 2011), and subsequently cloned into the modified backbone-vector pTAL-3.1, in which a cassette encoding a 152 aa TALE N-terminal and a 63 aa C-terminal segment fused to an enhanced FokI nuclease flank either side of the insert (Miller et al. 2011). The FokI nuclease sequences were slightly dissimilar each contained three aa substitutions, being Q486E, I499L and N496D in one and E490 K, I538 K and H537R in the other FokI nuclease domain, known to enhance specificity and thus reducing off-target effects (Doyon et al. 2011). For in vitro transcription, a T3 promoter was inserted upstream of the TALEN cassette producing the final construct denoted pTAL-3.1 (Fig. 1b).

Schematic representation of the designed TALENs and binding site in the murine ACE2 gene. a Top panel; genomic structure of the ACE2 gene, where the 19 exons are depicted as boxes, of which non-coding sequences are in light grey and coding sequence in dark grey. Bottom panel; architecture of the designed TALENs and their binding site in exon 2 of the Ace2 gene. Both TALEN arms contain a nuclear localization signal (NLS), a truncated 152 amino acid N-terminus, and a 63 amino acid C-terminus fused to a FokI nuclease. FokIELD and FokIKKR represent the 3-aa substituted versions of the original FokI. A restriction site recognized by the endonuclease DraI is underlined and in italic. b Schematic representation of the final TALEN construct pTAL-3.1 featuring a cytomegalovirus (CMV) promoter, a T7 RNA polymerase promoter (T7), either the FokIELD or FokIKKR TALEN arm juxtaposed to a bovine growth hormone polyadenylation signal (BGH pA) and flanked by a neomycin selection marker driven by an ampicillin resistance promoter (bla) completed with a simian virus 40 polyadenylation signal (SV40 pA)
Microinjection of zygotes
The TALEN expression vector was digested by the restriction endonucleases AflII and XhoI (NEB, England), and the fragment containing the T3 promoter juxtaposed TALEN sequence was recovered after agarose gel electrophoresis separation using a gel extraction kit (Qiagen, Germany). Subsequently, in vitro transcription was performed according to manufacturer’s instructions using the mMESSAGE mMACHINE T3 kit (Life technologies, USA), followed by polyA tailing (Epicentre, USA). Crude mRNA was precipitated by means of lithium chloride and finally dissolved in nuclease-free water (Sigma, USA).
Eight to ten weeks old female Kunming mice [an outbred mouse strain deriving from Swiss Albino mouse (Deng et al. 2013)] were superovulated by injection of 10 IU PMSG (pregnant mare serum gonadotropin) and 10 IU hCG (human chorionic gonadotropin) 48 h later (Ningbo second hormone factory, Ningbo) and then mated with male mice. Zygotes were collected to which 0.5–2 nL (50 ng/μL) mRNA of each TALEN was microinjected into the cytoplasm using a standard microinjection system (Eppendorf, Germany). The dose of each TALEN pair injected was empirically determined, based on the criteria of more than 50 % embryos alive post-injection. After injection, embryos were immediately transferred surgically into pseudo-pregnant female mice. Mice were housed in a specific pathogen-free facility with ad libitum access to food and water.
Mutation identification of the founder mice
DNA was phenol–chloroform extracted from toe clips of newborn mice. PCR was conducted on the extracted DNA using primers spanning the target site, generating a 432 bp amplicon (ACE2-F, 5′-CTTCTCAGTGCCCAACCCA-3′; ACE2-R, 5′-GGATCAGAGCTACAGAGGCAGT-3′) (Table 1). RFLP (Restriction Fragment Length Polymorphism) was carried out to test whether the specific restriction site was disturbed by the TALEN nucleases. Briefly, the PCR product was cut by 2U of DraI restriction endonucleases (NEB, England) at 37 °C overnight. Afterwards, 4 μL of the digest-product was subjected to electrophoresis on a 2.0 % agarose gel. The PCR product was additionally, cloned into the pMD18-T vector (Takara, China) and Sanger sequencing in order to confirm the mutations.
Mutation detection by T7 endonuclease I assay
Interrogation of potential off-target mutations was performed using T7 endonuclease I (T7EI) assay according to Sakurai’s description with minor modifications (Sakurai et al. 2014). Briefly, PCR was performed on DNA extracted from the 49 founder mice using the OT1 and OT2 primer pairs (Table 1). Subsequently, a mix containing 18 μL PCR product and 2 μL of 10 × NEB buffer 2 was incubated at 95 °C for 10 min and then incubated at room temperature for ~30 min. After the annealing reaction, 0.5 μL of T7 endonuclease I (2.5 U/μL, NEB) was added to the sample and the mixture was incubated at 37 °C for 1 h. Finally, the digested product was electrophoresed on a 2.0 % agarose gel.
Induction of acute colitis
Colitis was induced with 5 % (w/v) DSS (molecular mass 36–50 kDa, Sigma) dissolved in drinking water provided for the first through the seventh experimental day. The DSS solution was freshly prepared every day. Body weight and diarrhea scores (0, no diarrhea; 1, mild diarrhea; 2, severe watery diarrhea; 3, mild diarrhea with blood; 4, severe watery diarrhea with blood.) were documented daily. Colon length was measured at the end of the experiment. A fraction of the colon was soaked in 4 % paraformaldehyde for histological imaging and a fraction was immediately stored in liquid nitrogen for RNA isolation. For this experiment, male mice were divided into four groups: matched littermate wild-type mice with normal water (wt, control), wild-type mice with DSS water (wt, DSS), Ace2-knockout mice with normal water (Ace2 −/y, control; #3*, #5*, #11*, #30*, #35* and #25*), and Ace2-knockout mice with DSS water (Ace2 −/y, DSS; #4*, #9*, #28*, #31*, #24* and #27*).
Histological analysis
Colon fractions from DSS treated or untreated Ace2−/y mice (#3*, #5*, #11* and #4*, #9*, #28*, respectively) as well as DSS treated or untreated wt mice (#12*, #27*, #38* and #6*, #31*, #76*, respectively) were embedded in paraffin and then cut into 5–6 μm thick sections. Haematoxylin and eosin staining was performed in order to visualize histological changes. Procedures were carried out according to Fischer’s description with few modifications (Fischer et al. 2008).
Transcription analysis of inflammatory cytokines
Total RNA was extracted from the colon of three DSS treated (#3*, #5*, #11*) and three untreated (#4*, #9*, #28*) Ace2 −/y animals as well as from matched wt controls by the use of the Trizol reagent according to manufacturer’s instructions. Subsequently, 1 μg of RNA was reverse-transcribed into cDNA using a first-strand cDNA synthesis kit (Takara, China). One micro litre of cDNA was used for quantification on an ABI 7500 real-time machine (ABI, CA, USA) using SYBR Green (Takara, China) and gen expression primer-pairs for the inflammatory cytokines IL1B, IL6, IN10, IFNG, and TNF-α. GAPDH was employed as internal control and used for normalization of Ct values.
Statistical analysis
All the data were presented as mean ± SD. Two-tailed T test was performed to evaluate the statistical significance between two means of equal variance. One asterisk (*) indicates statistical difference complying with the criteria P < 0.05, and two asterisks (**) P < 0.01.
Results
Generation of TALEN induced Ace2 knockout mice
Murine Ace2 produces three transcripts, exon 1–19, 2–19 and 7–18. The generated TALENs target exon 2 immediately downstream of the start codon, thereby introducing frameshift mutations (Fig. 1a). Disruption of the full-length cellular membrane bound ACE2 has been shown sufficient for severe decrease in enzymatic activity with physiological implications (Gurley et al. 2006; Wysocki et al. 2006). Residual activity may stem from the putative secreted short isoform.
In all, 240 injected embryos were transferred into ten pseudo-pregnant mice, of which eight were eventually pregnant and gave birth to 49 live founder mice. Results from a restriction fragment length polymorphism (RFLP) analysis showed that 28 of the newborn mice (57 %) completely or partially lost the spacer containing the DraI site (Fig. 2a), which was confirmed by TA clone sequencing (Fig. 2b). As the Ace2 gene locates to the X chromosome, male mice have only one copy of this gene. Hence, in male mice, the Ace2 gene is completely knocked out more readily, compared to female mice, where a wild-type allele is usually maintained. Among the females, four mice (#17, #20, #26, #27) were biallelic knockouts (Ace2 −/−). Mosaic founder mice having more than two allele types were also identified, such as mouse #8, #22, #23, #38 and particularly mouse 24#, which was shown to harbor four different alleles (Fig. 2a, Supplementary Figure S1). In total, 11 animals with frameshift mutation(s) were identified.

Identification and subdivision of TALEN-induced deletions in founder mice. a Genotyping of all newborn mice by gel electrophoresis of DraI digested PCR products spanning the Ace2 gene target site. The uncut bands (top) signify the mutated alleles with a deleted DraI site. The lower two bands indicate a wt allele whereas a combination is indicative of a mosaicism. Male mice IDs are in red and female IDs in black. The encircled IDs correspond to the founder animals giving rise to the F1 population. b Animals are stratified into a male and female group, constituting 54 and 46 % of the whole population, respectively. Sequencing identified deletions are binned according to size and the percent-wise constitution of these in the two groups is depicted. c Cumulative frequency of base loss around the Ace2 target site. Deletions in male or heterologous female mice produce a count of one whereas biallelic deletions give a count of two. Mosaic animals are excluded. TALEN binding sites are shown in red. (Color figure online)
The herein used TALENs induced a wide range of deletions, from 3 to 102 bp. Most of the knockout (KO) founders lost small fragments (≤20 bp), where only one founder lost 102 bp (Supplementary Figure S1). This suggests that repair of small deletions was the preferable response to TALEN-induced DNA breaks. The cumulative loss frequency was calculated as the sum of a specific base deletion in all the mutant Ace2 alleles except the mosaic ones, as the genotype distribution here is not known. The resulting graph showed that bases within the spacer region displayed a relatively higher loss frequency, presumably because FokI dimerization occurs at this region (Fig. 2c). Importantly, there was a peak at the third “T” and not the last “T” at which FokI was expected to cut preferably given the TALEN design parameters.
Taken together, the result implied that bases in the middle of the spacer were the most likely to be deleted leading to down-regulation of expression.
No detectable off-target cleavage in any of the 49 founder mice
Although inherently site-specific, unsolicited cleavage is a perpetual issue of genome-editing tools. To interrogate possible off-target cleavage by the Ace2 TALENs, we blasted the targeting sequence against the mouse reference genome, and picked the second and third most complimentary hit to be analyzed for mutations.
DNA samples from all the 49 founder mice were divided into four groups, each consisting of 12–13 samples. The pooled DNA was amplified by two pairs of primers spanning the two off-target sites, respectively. Off-target mutations were then detected by T7 endonuclease I (T7EI) assay as previously described (Sakurai et al. 2014). No cleaved bands were detected, indicating that the TALENs at least did not target these two sites and presumably no other site predicted by the primary sequence alone (Fig. 3a, b).

T7EI analysis of the two potential off-target sites hit by TALENs. a Off-target site 1. b Off-target site 2. Pool 1–4: T7EI analysis of the PCR product from the DNA pool of founder mice #1–#12 (pool 1), #13–#24 (pool 2), #25–#36 (pool 3), and #37–#49 (pool 4). PCR: the PCR product from wt mouse DNA. PCR T7EI: T7EI analysis of the PCR product from a wild-type mouse. “*”: unspecific amplification. M: 100 bp DNA ladder
Germline transmission of TALEN-induced mutations
To test the efficiency and stability of the induced mutations during germline transmission, three male mice (#2, #3 and #5) were backcrossed with wild-type mice. The paternal X chromosome is incontestably passed on to female offspring, therefore we genotyped all female mice in the F1 generation. The three founders gave birth to a total of 23 mice, including 14 females. As expected, the analyzed animals all displayed a heterozygous genotype in the RFLP assay (Supplementary Figure S2), implying that the mutant alleles were successfully transferred vertically. These results indicated that the TALEN-induced modification was highly heritable.
Generation of F1 Ace2 KO mice
Female founders with frameshift mutations (#17wt/Δ4, #20 wt/Δ10 and #24 wt/Δ4/Δ20+1/+1) were backcrossed with wild-type mice to generate the F1 population. F1 mice were genotyped by the RFLP-DraI method and in total 37 genetically modified mice were obtained equally distributed between the two genders (Table 2). All listed male mice are Ace2 −/y whereas all female mice are mono-allelic knockouts, Ace2 −/+. To assess if the introduced frameshift mutations in F1 Ace2−/y mice disrupted gene-expression already at the level of mRNA, potentially due to non-sense mediated decay, and to clarify if it affected transcription of shorter isoform, we performed reverse transcription semi-quantitative PCR on total RNA extracted from three Ace2 −/y mice (#5*, #11* and #36*) and three male wt controls (#12, #31, #38). The two longer Ace2 transcripts where identifying with exon 7–9 spanning primers, whereas for the short putative isoform exon 1 and 3 spanning primers (corresponding to exon 7 and 9 in the longer isoforms) were used (Fig. 4a–c). The longer isoforms are present in all analyzed animals although the band-intensity is weaker in the KO animals hinting at a lower mRNA level plausible due to non-sense mediated decay of the out of frame transcript. Importantly, no short Ace2 mRNA was present in any of the investigated animals suggesting that this particular isoform is not expressed in the colon, which could otherwise be a source of confounding effects. On the other hand, even though immunodetection of the shorter ACE2 variant has been reported in renal cortex, the overall activity of ACE2 was below 8 % (Wysocki et al. 2006).

Expression profiling of the long and short Ace2 transcript. The presence of the a long transcripts and b short putative transcript in the colon of three KO mice (#5*, #11*, #36*) and three wt mice (#12, #31, #38) was assessed by RT-qPCR. Primers spanning exon 7–9 produce a fragment of 564 bp, whereas primers spanning 1–3 of the shorter mRNA theoretically produces a 412 bp amplicon. Gapdh was included as an internal control. M: 100 bp DNA ladder. W: water
Increased susceptibility to DSS-induced colitis in Ace2-KO mouse
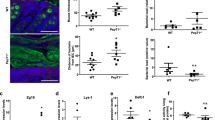
DSS is a sulfated polysaccharide known for its intestinal epithelial barrier disrupting potential and has been used widely to induce murine models of intestinal inflammation. The colitis responses have been reported to be dependent on DSS concentration, molecular weight, duration of the exposure, and also genetic background (Perse and Cerar 2012). In this study, clinical problems were eventually observed in all mice challenged with DSS, including body weight loss (Fig. 5a) and severe diarrhea (data not shown). More importantly, challenged ACE2-deficient mice suffered from more severe diarrhea and lost much more body weight compared to age- and gender-matched wild-type littermates that were equally exposed to the drug. After 7 days DSS feeding, the colon length of mice from the challenged groups was shorter compared to the control groups (wt, DSS vs. wt control and Ace2-KO, DSS vs. Ace2-KO, control; Fig. 5b). Moreover, we detected significant shortening of the colon in DSS treated Ace2-KO mice compared to the wt counterparts, a phenotypic trait characteristic for severe colitis.

ACE2 deficiency increased susceptibility to DSS-induced loss of body weight and colon length. a Daily recording of body weight loss during the DSS feeding period, where body weight loss was calculated as: 1—(body weight at day i)/(body weight at day 0), i = 0, 1,2, 3, 4, 5, 6, 7. b Colon length measurement at the end of the DSS induction period; n = 6 mice in each group
Enhanced infiltration of inflammatory cells in Ace2 −/y mice
As intestinal injury causes a gut epithelium elicited inflammatory response (Karhausen et al. 2013), we carried out haematoxylin and eosin staining to assess the histopathologic alterations in the colon. Infiltration of inflammatory cells was observed in two groups of mice treated with DSS (Fig. 6 and Supplementary Figure S3). In contrast to treated wild-type mice, colons of treated Ace2 −/y mice showed a higher degree of leukocyte infiltration. Overt damage to the intestinal mucosae could be observed, including shedding of epithelium and the breakdown of the lamina propria. Furthermore, a large number of leukocytes entered into the lamina propria. However, severe histologic changes could be observed in one of the three DSS treated control mice. In patients with lymphoplasmacytic colitis, inflammation of the lamina propria is interspersed between regions of normal mucosa making it conceivable that a patchy lymphoplasmacytic inflammation is present in all DSS treated mice but with a higher occurrence in Ace2 −/y mice. In previous studies, the recruitment and activation of neutrophils and macrophages at the site of inflammation has been recognized as key steps in the intestinal immune response (Knutson et al. 2013; Mangerich et al. 2012). Furthermore, evidence suggests a causal link between infiltration of leukocytes and colitis severity (McBee et al. 2010). Together, this suggests that the inactivation of Ace2 potentially leads to a more severe form of DSS induced colitis in the herein described mice through lymphoplasmacytic infiltration and mucosal barrier damage.

Cross-sectional H&E staining of the colon. Haematoxylin and eosin staining of the colon after 7 days of DSS challenge (mouse #12, #9*) or untreated (mouse #6, #3*). Asterisks mark infiltration of leukocytes in DSS-treated mice. 1: Gastric duct; 2: Epithelium; 3: Lamina propria; 4: Muscularis mucosae; 5: Submucosa; 6: Circular muscle. The pictures were magnified 200x
Up-regulated expression of inflammatory cytokines in Ace2 −/y mice
Inappropriate activation of the mucosal immune system can trigger the onset of colitis. Numerous studies have shown that the expression and secretion of various cytokines is altered in inflammatory conditions such as inflammatory bowel disease (Nanau and Neuman 2012; Wittkopf et al. 2014; Simoes e Silva et al. 2013). The elevated expression of cytokines belonging to the IL-17 and IL-22 pathways such as IL1-β (interleukin 1 beta), IL6 (interleukin 6), IL10 (interleukin 10), IFN-γ (interferon gamma) and TNF-α (tumor necrosis factor alpha) point to the recruitment of neutrophils and possibly also natural killer (NK) cells and macrophages at the site of inflammation (Knutson et al. 2013). We therefore evaluated the transcription profile of several inflammatory cytokines presumably changed in mice with DSS-induced colitis. By use of reverse transcription quantitative (RT-qPCR) we quantified the expression level of IL1-β, IL6, IFN-γ, Il10 and TNF-α (Fig. 7a–e). A detectable increase in mRNA expression of IL1-β, IL6 and TNF-α was exhibited in DSS-fed wild-type mice in contrast to controls, implying a modest inflammation. Notably, Ace2 −/y mice consistently displayed a robust up-regulated expression of the pro-inflammatory cytokines (IL1-β, IL6, IFN-γ, and TNF-α) by 20-fold to 270-fold compared to their genetic counterparts. In contrast, the anti-inflammatory cytokine IL10 was only modestly up-regulated in treated Ace2 −/y mice and not in treated wt mice. A plausible explanation for this observation is the IL10 secretion stimuli of IL6 and the presence of NK cells, which activity is known to be inhibited by IL10 (Conti et al. 2003; Mauer et al. 2014). More importantly, this dramatic increase in cytokine transcription was also evident when compared to DSS-fed wild-type mice and thus in line with the more severe pathological alterations identified in the knockout mice. Taken together, these molecular clues suggested that Ace2 gene disruption conceivably is the driver of the colitis progression in the herein studied mice.

Cytokine profiling of the colon: Up-regulated expression of inflammatory cytokines in the colon of Ace2 −/y mice in response to the DSS challenge relative to the wt control. Colon extracted total RNA was used for RT-qPCR employing exon–exon primers specific for murine a Il1b, b Il6, c Il10, d Ifn-γ, and e Tnf-α. Gapdh was chosen as endogenous reference. Data is displayed as mean ± standard deviation; * P < 0.05, ** P < 0.01
Discussion
The Kunming mouse strain is widely used in China comprising about 70 % of the mice used in research, as it is easy to handle and produces large litter sizes. A major drawback, however, is the problematic ES cell isolation and culturing which has restricted its use for genetic manipulation. Only recently was it shown that ES cells could be isolated from hybrid Kunming embryos and so no genetic engineering in Kunming mice has so far been reported (Yu et al. 2014). Being an outbred mouse strain, efficient and stable genetic manipulation would besides heighten the strains usability also provide proof of concept for a tool functional in a heterogeneous genetic makeup. If this in addition is achieved without leaving footprints such as genomic integration of selection markers this could likely circumvent some of the disparities observed between otherwise identical animal disease models.
Dysfunction of the X-linked Ace2 has been associated with colitis, a disease with many causes that can have devastating effects if untreated (Hashimoto et al. 2012). We thus wanted to disrupt the Ace2 gene by microinjection of transcription activator-like effector nucleases (TALENs) mRNA into zygotes of an outbred mice strain (Kunming) serving as a proof of concept for readily and footprint-less creation of murine disease models.
TALENs have in recent years enjoyed much attention due to their simplicity and efficiency across species. Optimization of the expression system is critical for a high and specific cleavage activity and as described in previous studies, truncated amimo- and carboxy-terminals of the TALE protein, and heterodimerizing of FokI nuclease showed reduced off-target effects (Bedell et al. 2012; Doyon et al. 2011). As observed in our work, scaffold expressed TALENs composed of a 152-aa N-terminal, 63-aa C-terminal and a FokIELD/FokIKKR heterodimeric nuclease generated 28 (57 %) Ace2 modified specimens among the newborn pups. Repair of TALEN-induced DNA breaks in the Ace2 gene resulted in 19 alleles featuring small deletions (<20 bp) and only one allele containing an insertion in all the 21 modified alleles investigated. This finding is in coherence with previous findings in which murine cells were microinjected with TALEN mRNA or even whole-genome sequencing of human cells following CRISPR/Cas9-induced repair (Wang et al. 2014; Qiu et al. 2013; Li et al. 2014). Given that the Ace2 gene locates to the X chromosome, male mice displayed higher overall modification efficiency, while females are more prone to become mosaics.
The cargo delivering system is a crucial factor affecting editing efficiency. In our first attempt, we microinjected plasmids holding the TALEN pairs into the nucleus of mouse zygotes at a dosage of 5 ng/uL each. In the end no mutant mouse was identified out of the total 101 founders (data not shown). As an alternative approach, we delivered in vitro transcribed TALEN mRNA into the cytoplasm, generating the 28 genetically modified founder mice. The significant improvement of efficiency is likely due to the fact that cytoplasmic injection of abundant mRNA will result in more robust transcription than nucleic injection of DNA cassettes (Farrow et al. 2006). The rapid onset of transcription furthermore increases the likelihood of manipulating totipotent cells and thus what are to become primordial germ cells, enhancing transmission efficiency. Equally important, mRNA injections preclude the possibility of random genetic integration of the introduced cargo and therefore represent a more “clean” knockout.
Mosaicism is a quite universal phenomenon in TALEN-mediated gene knockout given the continuous expressional activity from TALEN mRNA during early cell-divisions or re-cleavage of already modified alleles (Sung et al. 2013; Tesson et al. 2011). Using fluorescence in situ hybridization it was shown that TALEN induced double-strand disruption commences before 6 h post-injection and although activity is waning, it is still observed 24 h post-injection (Li et al. 2014). In the current study, we noted that 5 out of 49 (10 %) mice were indeed mosaics. Along the same line, mosaicism has also been reported in CRISPR/Cas9-modified mice with a ratio up to 40 % (Yang et al. 2013).
In order to look for undesired cleavage, the sequence recognized by the candidate TALEN-pair was aligned to the host genome and the two most likely sites harboring off-target effects were chosen and analyzed. In this workflow, we found that the binding of the TALENs appeared highly specific and did not disturb any of the two “pseudo-targets”. Notably, the herein used TALENs had not been screened or optimized in any way.
Three F0 mice (#17, #20, #24) were used in the generation of F1 Ace2 −/y mice, which were subjected to the well-established colitis inducing DSS challenge. All Ace2 −/y mice exhibited a higher loss of body weight compared to matched wild-type animals after the challenge, which is likely due to severe diarrhea. Plausible this can be ascribed to the excessive inflammatory response and colon injury observed in the knockouts. Elevated expression of pro-inflammatory cytokines fitting a neutrophil profile such as TNF-α and IFN-γ were present in an exceedingly high level in the DSS challenged Ace2 −/y mice compared to the wt counterparts, pointing to an immense acute phase inflammation in the knockouts. Furthermore, an IL10 mediated anti-inflammatory response appeared to be activated in the induced Ace2 −/y mice. Notably, IL10 deficient mice develop spontaneous colitis (Sellon et al. 1998). Colon histopathology and the examination of anatomy substantiated these findings by showing damage to the mucosae layer of the intestine, infiltration of leukocytes in the epithelium and a shortening of length.
In conclusion, our data demonstrated that TALEN-nucleases delivered as mRNA constitutes a highly effective, clean and inheritable way to inactivate a gene of interest and to develop mouse models. It circumvents the challenge of low transgenerational transmission otherwise seen in blastocysts injected with manipulated ES cells. More so, this can be achieved in genetically heterozygous animals. The inhere described result made in the Ace2 −/y mice are consistent with findings in a similar mouse model produced by the conventional HR-based approach (Hashimoto et al. 2012), thereby underpinning the usability of TALENs as an alternative to ES cell-based gene targeting.
References
Bedell VM, Wang Y, Campbell JM, Poshusta TL, Starker CG, Krug RG 2nd, Tan W, Penheiter SG, Ma AC, Leung AY, Fahrenkrug SC, Carlson DF, Voytas DF, Clark KJ, Essner JJ, Ekker SC (2012) In vivo genome editing using a high-efficiency TALEN system. Nature 491:114–118
Brenner MK, Rill DR, Moen RC, Krance RA, Mirro J Jr, Anderson WF, Ihle JN (1993) Gene-marking to trace origin of relapse after autologous bone-marrow transplantation. Lancet 341:85–86
Byrnes JJ, Gross S, Ellard C, Connolly K, Donahue S, Picarella D (2009) Effects of the ACE2 inhibitor GL1001 on acute dextran sodium sulfate-induced colitis in mice. Inflamm Res 58:819–827
Cermak T, Doyle EL, Christian M, Wang L, Zhang Y, Schmidt C, Baller JA, Somia NV, Bogdanove AJ, Voytas DF (2011) Efficient design and assembly of custom TALEN and other TAL effector-based constructs for DNA targeting. Nucleic Acids Res 39:e82
Conti P, Kempuraj D, Kandere K, Di Gioacchino M, Barbacane RC, Castellani ML, Felaco M, Boucher W, Letourneau R, Theoharides TC (2003) IL-10, an inflammatory/inhibitory cytokine, but not always. Immunol Lett 86:123–129
Crackower MA, Sarao R, Oudit GY, Yagil C, Kozieradzki I, Scanga SE, Oliveira-dos-Santos AJ, da Costa J, Zhang L, Pei Y, Scholey J, Ferrario CM, Manoukian AS, Chappell MC, Backx PH, Yagil Y, Penninger JM (2002) Angiotensin-converting enzyme 2 is an essential regulator of heart function. Nature 417:822–828
Deng ZB, Yuan AW, Luo W, Wang ND, Gong QL, Yu XL, Xue LQ (2013) Transmission of porcine circovirus type 2b (PCV2b) in Kunming mice. Acta Veterinaria Hung 61:234–243
Ding Q, Lee YK, Schaefer EA, Peters DT, Veres A, Kim K, Kuperwasser N, Motola DL, Meissner TB, Hendriks WT, Trevisan M, Gupta RM, Moisan A, Banks E, Friesen M, Schinzel RT, Xia F, Tang A, Xia Y, Figueroa E, Wann A, Ahfeldt T, Daheron L, Zhang F, Rubin LL, Peng LF, Chung RT, Musunuru K, Cowan CA (2013) A TALEN genome-editing system for generating human stem cell-based disease models. Cell Stem Cell 12:238–251
Donoghue M, Hsieh F, Baronas E, Godbout K, Gosselin M, Stagliano N, Donovan M, Woolf B, Robison K, Jeyaseelan R, Breitbart RE, Acton S (2000) A novel angiotensin-converting enzyme-related carboxypeptidase (ACE2) converts angiotensin I to angiotensin 1–9. Circ Res 87:E1–E9
Doyle EL, Booher NJ, Standage DS, Voytas DF, Brendel VP, Vandyk JK, Bogdanove AJ (2012) TAL Effector-Nucleotide Targeter (TALE-NT) 2.0: tools for TAL effector design and target prediction. Nucleic Acids Res 40:W117–W122
Doyon Y, Vo TD, Mendel MC, Greenberg SG, Wang J, Xia DF, Miller JC, Urnov FD, Gregory PD, Holmes MC (2011) Enhancing zinc-finger-nuclease activity with improved obligate heterodimeric architectures. Nat Methods 8:74–79
Epinat JC, Arnould S, Chames P, Rochaix P, Desfontaines D, Puzin C, Patin A, Zanghellini A, Paques F, Lacroix E (2003) A novel engineered meganuclease induces homologous recombination in yeast and mammalian cells. Nucleic Acids Res 31:2952–2962
Farrow PJ, Barrett LB, Stevenson M, Fisher KD, Finn J, Spice R, Allan MA, Berry M, Logan A, Seymour LW, Read ML (2006) Cytoplasmic expression systems triggered by mRNA yield increased gene expression in post-mitotic neurons. Nucleic Acids Res 34:e80
Fischer AH, Jacobson KA, Rose J & Zeller R (2008) Hematoxylin and eosin staining of tissue and cell sections, CSH protocols 2008(5):pdb–prot4986. doi: 10.1101/pdb.prot4986
Gurley SB, Allred A, Le TH, Griffiths R, Mao L, Philip N, Haystead TA, Donoghue M, Breitbart RE, Acton SL, Rockman HA, Coffman TM (2006) Altered blood pressure responses and normal cardiac phenotype in ACE2-null mice. J Clin Investig 116:2218–2225
Guyton AC (1992) Kidneys and fluids in pressure regulation. Small volume but large pressure changes. Hypertension 19:I2–I8
Hashimoto T, Perlot T, Rehman A, Trichereau J, Ishiguro H, Paolino M, Sigl V, Hanada T, Hanada R, Lipinski S, Wild B, Camargo SM, Singer D, Richter A, Kuba K, Fukamizu A, Schreiber S, Clevers H, Verrey F, Rosenstiel P, Penninger JM (2012) ACE2 links amino acid malnutrition to microbial ecology and intestinal inflammation. Nature 487:477–481
Hockemeyer D, Wang H, Kiani S, Lai CS, Gao Q, Cassady JP, Cost GJ, Zhang L, Santiago Y, Miller JC, Zeitler B, Cherone JM, Meng X, Hinkley SJ, Rebar EJ, Gregory PD, Urnov FD, Jaenisch R (2011) Genetic engineering of human pluripotent cells using TALE nucleases. Nat Biotechnol 29:731–734
Izcue A, Powrie F (2012) Immunology: malnutrition promotes rogue bacteria. Nature 487:437–439
Karhausen J, Qing M, Gibson A, Moeser AJ, Griefingholt H, Hale LP, Abraham SN, Mackensen GB (2013) Intestinal mast cells mediate gut injury and systemic inflammation in a rat model of deep hypothermic circulatory arrest. Crit Care Med 41:e200–e210
Kim YG, Cha J, Chandrasegaran S (1996) Hybrid restriction enzymes: zinc finger fusions to Fok I cleavage domain. Proc Natl Acad Sci USA 93:1156–1160
Knutson CG, Mangerich A, Zeng Y, Raczynski AR, Liberman RG, Kang P, Ye W, Prestwich EG, Lu K, Wishnok JS, Korzenik JR, Wogan GN, Fox JG, Dedon PC, Tannenbaum SR (2013) Chemical and cytokine features of innate immunity characterize serum and tissue profiles in inflammatory bowel disease. Proc Natl Acad Sci USA 110:E2332–E2341
Li C, Qi R, Singleterry R, Hyle J, Balch A, Li X, Sublett J, Berns H, Valentine M, Valentine V, Sherr CJ (2014) Simultaneous gene editing by injection of mRNAs encoding transcription activator-like effector nucleases into mouse zygotes. Mol Cell Biol 34:1649–1658
Mangerich A, Knutson CG, Parry NM, Muthupalani S, Ye W, Prestwich E, Cui L, McFaline JL, Mobley M, Ge Z, Taghizadeh K, Wishnok JS, Wogan GN, Fox JG, Tannenbaum SR, Dedon PC (2012) Infection-induced colitis in mice causes dynamic and tissue-specific changes in stress response and DNA damage leading to colon cancer. Proc Natl Acad Sci USA 109:E1820–E1829
Mauer J, Chaurasia B, Goldau J, Vogt MC, Ruud J, Nguyen KD, Theurich S, Hausen AC, Schmitz J, Bronneke HS, Estevez E, Allen TL, Mesaros A, Partridge L, Febbraio MA, Chawla A, Wunderlich FT, Bruning JC (2014) Signaling by IL-6 promotes alternative activation of macrophages to limit endotoxemia and obesity-associated resistance to insulin. Nat Immunol 15:423–430
McBee ME, Zeng Y, Parry N, Nagler CR, Tannenbaum SR, Schauer DB (2010) Multivariate modeling identifies neutrophil- and Th17-related factors as differential serum biomarkers of chronic murine colitis. PLoS One 5:e13277
Miller JC, Tan S, Qiao G, Barlow KA, Wang J, Xia DF, Meng X, Paschon DE, Leung E, Hinkley SJ, Dulay GP, Hua KL, Ankoudinova I, Cost GJ, Urnov FD, Zhang HS, Holmes MC, Zhang L, Gregory PD, Rebar EJ (2011) A TALE nuclease architecture for efficient genome editing. Nat Biotechnol 29:143–148
Nanau RM, Neuman MG (2012) Metabolome and inflammasome in inflammatory bowel disease. Transl Res 160:1–28
Perse M, Cerar A (2012) Dextran sodium sulphate colitis mouse model: traps and tricks. J Biomed Biotechnol 2012:718617
Qiu Z, Liu M, Chen Z, Shao Y, Pan H, Wei G, Yu C, Zhang L, Li X, Wang P, Fan HY, Du B, Liu B, Liu M, Li D (2013) High-efficiency and heritable gene targeting in mouse by transcription activator-like effector nucleases. Nucleic Acids Res 41:e120
Ramirez-Sanchez M, Prieto I, Wangensteen R, Banegas I, Segarra AB, Villarejo AB, Vives F, Cobo J, de Gasparo M (2013) The renin-angiotensin system: new insight into old therapies. Curr Med Chem 20:1313–1322
Sakurai T, Watanabe S, Kamiyoshi A, Sato M, Shindo T (2014) A single blastocyst assay optimized for detecting CRISPR/Cas9 system-induced indel mutations in mice. BMC Biotechnol 14:69
Sellon RK, Tonkonogy S, Schultz M, Dieleman LA, Grenther W, Balish E, Rennick DM, Sartor RB (1998) Resident enteric bacteria are necessary for development of spontaneous colitis and immune system activation in interleukin-10-deficient mice. Infect Immun 66:5224–5231
Simoes e Silva AC, Silveira KD, Ferreira AJ, Teixeira MM (2013) ACE2, angiotensin-(1-7) and Mas receptor axis in inflammation and fibrosis. Br J Pharmacol 169:477–492
Sun N, Zhao H (2013) Transcription activator-like effector nucleases (TALENs): a highly efficient and versatile tool for genome editing. Biotechnol Bioeng 110:1811–1821
Sung YH, Baek IJ, Kim DH, Jeon J, Lee J, Lee K, Jeong D, Kim JS, Lee HW (2013) Knockout mice created by TALEN-mediated gene targeting. Nat Biotechnol 31:23–24
Tesson L, Usal C, Menoret S, Leung E, Niles BJ, Remy S, Santiago Y, Vincent AI, Meng X, Zhang L, Gregory PD, Anegon I, Cost GJ (2011) Knockout rats generated by embryo microinjection of TALENs. Nat Biotechnol 29:695–696
Wang T, Wei JJ, Sabatini DM, Lander ES (2014) Genetic screens in human cells using the CRISPR-Cas9 system. Science 343:80–84
Wittkopf N, Neurath MF, Becker C (2014) Immune-epithelial crosstalk at the intestinal surface. J Gastroenterol 49:375–387
Wu T, Bloom ML, Yu JM, Tisdale JF, Dunbar CE (1998) Murine bone marrow expressing the neomycin resistance gene has no competitive disadvantage assessed in vivo. Hum Gene Ther 9:1157–1164
Wysocki J, Ye M, Soler MJ, Gurley SB, Xiao HD, Bernstein KE, Coffman TM, Chen S, Batlle D (2006) ACE and ACE2 activity in diabetic mice. Diabetes 55:2132–2139
Yamamoto K, Ohishi M, Katsuya T, Ito N, Ikushima M, Kaibe M, Tatara Y, Shiota A, Sugano S, Takeda S, Rakugi H, Ogihara T (2006) Deletion of angiotensin-converting enzyme 2 accelerates pressure overload-induced cardiac dysfunction by increasing local angiotensin II. Hypertension 47:718–726
Yang H, Wang H, Shivalila CS, Cheng AW, Shi L, Jaenisch R (2013) One-step generation of mice carrying reporter and conditional alleles by CRISPR/Cas-mediated genome engineering. Cell 154:1370–1379
Yu S, Yan X, Liu H, Cai X, Cao S, Shen L, Zuo Z, Deng J, Ma X, Wang Y, Ren Z (2014) Improved establishment of embryonic stem (ES) cell lines from the Chinese Kunming mice by hybridization with 129 mice. Int J Mol Sci 15:3389–3402
Zhang H, Wada J, Hida K, Tsuchiyama Y, Hiragushi K, Shikata K, Wang H, Lin S, Kanwar YS, Makino H (2001) Collectrin, a collecting duct-specific transmembrane glycoprotein, is a novel homolog of ACE2 and is developmentally regulated in embryonic kidneys. J Biol Chem 276:17132–17139
Acknowledgments
We are grateful to all members of the transgenic group at BGI. This work was supported by research grants from National Transgenic Project of China (2014ZX0801007B), National Key Basic Research Program of China—973 program (No. 2011CB944201) of China, Shenzhen Engineering Laboratory for Genomics-Assisted Animal Breeding, the Shenzhen Municipal Government of China (No. CXZZ20120615141322654).
Author information
Authors and Affiliations
Corresponding authors
Additional information
Chuxin Liu, Liping Xiao and Feida Li have contributed equally to this article.
Electronic supplementary material
Below is the link to the electronic supplementary material.
11248_2014_9855_MOESM1_ESM.pptx
Supplementary material 1 Sequencing across indels in the Ace2 target site of founder mice. Binding sites of the TALEN pair are shown in red, and the DraI site is underlined. Dashed lines represent base deletion whereas inserted bases are indicated by blue letters. The ID numbers of five mosaic specimens are underlined. Type of mutation is indicated on the right side of the figure (PPTX 68 kb)
11248_2014_9855_MOESM2_ESM.pptx
Supplementary material 2 Genotyping of female offspring from three male founders. DNA from 14 female offspring, progeny of three male founders (#2, #3 and #5) backcrossed with wt mice, was subjected to an RFLP-DraI assay and visualized on 2 % agarose gel. M: 200 bp DNA ladder (PPTX 682 kb)
11248_2014_9855_MOESM3_ESM.pptx
Supplementary material 3 Additional cross-sectional H&E staining pictures of the colon. Haematoxylin and eosin staining of the colon from seven days untreated or DSS challenged mice. The pictures were magnified 200x (PPTX 1955 kb)
Rights and permissions
About this article

Cite this article
Liu, C., Xiao, L., Li, F. et al. Generation of outbred Ace2 knockout mice by RNA transfection of TALENs displaying colitis reminiscent pathophysiology and inflammation. Transgenic Res 24, 433–446 (2015). https://doi.org/10.1007/s11248-014-9855-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11248-014-9855-3