
Abstract
When males become more ornamented and reproduce more successfully as they grow older, phenotypic correlations between ornament exaggeration and reproductive success can be confounded with age effects in cross-sectional studies, and thus say relatively little about sexual selection on these traits. This is exemplified here in a correlative study of male fertilization success in a large colony of American barn swallows (Hirundo rustica erythrogaster). Previous studies of this species have indicated that two sexually dimorphic traits, tail length and ventral plumage coloration, are positively correlated with male fertilization success, and a mechanism of sexual selection by female choice has been invoked. However, these studies did not control for potential age-related variation in trait expression. Here, we show that male fertilization success was positively correlated with male tail length but not with plumage coloration. We also show that 1-year-old males had shorter tails and lower fertilization success than older males. This age effect accounted for much of the covariance between tail length and fertilization success. Still, there was a positive relationship between tail length and fertilization success among older males. But as this group consisted of males from different age classes, an age effect may be hidden in this relationship as well. Our data also revealed a longitudinal increase in both tail length and fertilization success for individual males. We argue that age-dependent ornament expression and reproductive performance in males complicate inferences about female preferences and sexual selection.
Similar content being viewed by others


Introduction
A central theme in the study of animal mating systems is why some males succeed in fertilizing more eggs than others. Such analyses require good measures of fertilization success for individual males in combination with data on their morphological, physiological, behavioral, or genetic traits that might influence mating and fertilization processes. A conceptual framework for the relationship between traits and fertilization success is provided by the theory of sexual selection (Darwin 1871; Andersson 1994), which explains how traits mediate mating and fertilization success through male–male competition or female choice. An additional source of variation in male fertilization success lies within the individual itself. As explained by life-history theory (Stearns 1976, 1992; Lessells 1991), natural selection should shape an individual’s resource allocation over successive breeding seasons so as to maximize lifetime reproductive success. Consequently, male fertilization success may vary with age. Sources of variation in male fertilization success can therefore be found in intrinsic quality-differences among males, as emphasized by sexual selection theory, as well as in seasonal and age-related variation in mating effort within each male, as predicted by life-history theory.
The predominant mating system in birds is social monogamy in combination with extrapair mating (Bennett and Owens 2002). After more than two decades of paternity studies, conducted in hundreds of species, there is still much controversy about interpretations and understanding of avian extrapair mating systems and their variation (Griffith et al. 2002; Westneat and Stewart 2003; Arnqvist and Kirkpatrick 2005; Griffith 2007). While there probably are multiple reasons for the slow conceptual progress in this field, we would like to point to a particular interpretation bias that has become increasingly obvious and that relates specifically to the comprehension of variation in male fertilization success. Many avian paternity studies seem to seek explanations solely from sexual selection theory, and especially in the potential role of female extrapair mating preferences for male traits, while they largely ignore sources of variation caused by male age effects and life-history variation in male effort. Similar concerns have also been expressed in various forms by other authors in recent years (e.g., Westneat and Stewart 2003; Westneat 2006; Schmoll et al. 2007). The distinction between sexual selection and life-history explanations becomes especially important when male fertilization success is correlated with sex traits that also show age-dependent expression, which is in fact quite common (Manning 1985; Andersson 1994). How can sexual selection be inferred from such phenotypic correlations when both the male trait and male fertilization success show age-dependent expression?
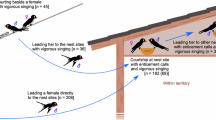
A simple graphical model (Fig. 1) can illustrate the rationale of sexual selection and life-history theories in the context of age-related correlations between a male sexual trait and fertilization success. Sexual selection theory assumes that males with elaborate traits achieve higher fertilization success, either because females use trait exaggeration as a signal in their mate choice or because males with elaborate traits compete more successfully against rivals for access to females and matings. In either case, the sex trait is instrumental for the individual’s mating success, i.e., a causal relationship. When males develop larger sex traits as they grow older, they will also achieve higher fertilization success as a consequence. When males of the same age vary in ornament expression, the sexual selection model will similarly predict a positive correlation between trait elaboration and fertilization success within age groups. The life-history model, on the other hand, assumes no causal relationship between the trait per se and fertilization success and will therefore not predict any positive correlation within age groups. However, since both variables show age-dependent increases as a consequence of more resources being allocated to reproduction and mating effort with age, there will be an overall correlation between trait expression and fertilization success across all males and age groups in the population. Hence, the two models can only be distinguished in correlational studies where correlations are broken down by age groups; a positive correlation between trait expression and fertilization success within age groups can provide unique support for sexual selection.

Two alternative explanations for phenotypic correlations between ornament size and mating success when both variables are age-related. In the sexual selection model, males with larger ornaments have higher mating success, either through a female preference or improved competitive ability against rivals. In the life-history model, there is no causal relationship between ornament and mating success, but males increase their mating effort over successive breeding seasons. Both models assume an age-related increase in ornament size and mating success across all males in the population, but only the sexual selection model predicts a positive relationship within each age class. There is also a theoretical possibility of a combination of the two processes, for example, that sexual selection effects only apply to certain age classes
Here, we present a correlative analysis of male fertilization success and two ornamental sex traits—tail streamer length and rufous ventral coloration—in a Canadian population of the barn swallow Hirundo rustica. Our aim is to examine whether these traits show age-dependent expression, and if so, whether variation in male fertilization success is better predicted by their age than their ornaments. The barn swallow has a typical passerine mating system with socially monogamous pair bonds and extensive extrapair mating (Møller 1994; Turner 2006). It is widely distributed across North America and Eurasia, and currently, six subspecies are recognized (Dor et al. 2010). Extensive studies of the nominate subspecies in Europe have made it an icon for sexual selection (Møller 1994), as both experimental and correlational evidence has indicated that the long tail in males is under current sexual selection by female choice in Denmark (Møller 1988; Møller and Tegelström 1997), Italy (Saino et al. 1997; Møller et al. 1998), and Spain (Møller et al. 2003). Similar studies in North America (ssp. erythrogaster) are less conclusive. Experimental manipulation of male tail length in a population in Ontario, Canada, revealed that males with elongated tails lost more paternity than males with shortened tails, i.e., opposite to what would be expected from a female within-pair preference for long tails (Smith et al. 1991). However, there was a significant negative relationship between the original, unmanipulated tail length of males and their paternity loss, which suggests that a long tail is an indicator of male superiority (Smith et al. 1991). A more recent study from the same area showed that male tail length correlated positively with both extrapair and total (i.e., sum of within-pair and extrapair) fertilization success (Kleven et al. 2006a). In contrast, two correlative studies from New York State, using the number of fledged young or within-pair paternity as measures of reproductive success, failed to find reproductive advantages in males with long tails (Safran and McGraw 2004; Neuman et al. 2007). A recent study of the gutturalis subspecies in Japan also found no evidence for male tail length being under current sexual selection (Kojima et al. 2009).
Two recent studies have indicated that the rufous ventral coloration is a sexually selected trait in the American barn swallow (Safran and McGraw 2004; Safran et al. 2005). Through a sophisticated experimental design, Safran et al. (2005) showed that manipulation of ventral coloration affected within-pair paternity; i.e., enhancement in male ventral coloration resulted in a higher proportion of offspring sired in the next breeding attempt.
While there are now quite a number of paternity studies conducted on barn swallows, with variable support for correlations between male fertilization success and different male traits (e.g., tail length and coloration), it is not yet known how male fertilization success specifically changes with age in this species. But long-term studies of ringed barn swallows in Denmark and Spain have indicated that males increase their reproductive performance over their first three breeding seasons, with a parallel increase in tail length and earlier spring arrival, and then show a subsequent reproductive decline (senescence) in their final years (Møller and de Lope 1999; Balbontín et al. 2007). This age effect seems to be due to a within-individual change in competence rather than selection (Balbontín et al. 2007). Paternity studies with information on male age (e.g., Møller et al. 2003, 2009) have focused primarily on the effects of ornamental traits, arrival date, and condition, controlling for male age, and not analyzed directly the effect of male age per se on paternity. Among socially monogamous passerines, male age generally seems to be an important factor affecting within-pair paternity, extrapair paternity, or both (cf. meta-analysis by Møller and Ninni 1998 and reviews by Griffith et al. 2002 and Schmoll et al. 2007). It is therefore of general importance to analyze the separate effects of male age and ornamental traits on male fertilization success in barn swallows.
With the aim of contrasting the ability of sexual selection and life-history models to predict variation in male fertilization success (Fig. 1), we analyzed how male fertilization success varied between male age groups and with respect to tail length and ventral coloration. If the two ornamental traits are under current sexual selection, we would expect correlations between trait expression and fertilization success to be manifested across all males in the population as well as within each age group. Alternatively, if male ornaments are not causally related to fertilization success but there is a strong age dependency in male reproduction and mating effort, we predict that older males should have more elaborate traits and higher fertilization success than young males, that age should be a better predictor of male fertilization success than ornament expression, and that there should be no correlation between ornaments and fertilization success within age classes.
Methods
We studied North American barn swallows from late April to mid-July in 2003, and from late April to early August in 2004, in a single breeding colony situated under a road bridge (44°28′05" N, 76°28′40" W), approximately 20 km from the Queen’s University Biological Station, Ontario, Canada. This is close to the study area of Smith et al. (1991). Paternity analyses from this colony were conducted as part of previous studies that also included other colonies (Kleven et al. 2005; Kleven et al. 2006a, b). Our general field procedures are detailed in these papers, so we only briefly outline them here. Adults were captured with mist nets, ringed, and given a unique color code of acrylic paint on a small spot on their right wing for field identification of birds on their nests. We captured and marked nearly all of the breeding individuals (98% in 2003 and 99% in 2004) in our study colony. Barn swallows show strong breeding site tenacity, with only exceptional cases of adults dispersing to a new colony within or between seasons (Shields 1984; Møller 1994; Safran 2004). In our population, we recorded a 53% return rate in 2004 of adults breeding in 2003 (Kleven et al. 2005), which is even higher than the annual survival rate for adults calculated in a long-term study in Denmark (~34% for both sexes; Møller and Szép 2002). It is thus reasonable to assume that new individuals in 2004 were first-time breeders. In fact, six of 77 new male breeders in 2004 (data from some additional smaller colonies included) were hatched in the same colony the year before (Kleven et al. 2005). Hence, we classified first-time breeders in 2004 as “young,” and returned breeders as “older.” Cross-sectional analyses of age differences could therefore be conducted only on the data set from 2004, whereas longitudinal effects could be analyzed from males breeding in both 2003 and 2004.
The length of the outermost tail feathers on both sides was measured to the nearest 0.5 mm using a ruler with a pin-zero-stop. All tail length measurements were conducted by one person (O.K.), and repeatability of the measurements was high (r = 0.99; Kleven et al. 2006a). In the following analyses, we have used the mean of the right and left outermost tail feathers in statistical analyses of tail length, but for three males, tail length was based on one tail feather due to the other being broken.
We collected 5–15 feathers from each of four ventral plumage regions (throat, breast, belly, and vent) for later analyses of coloration. Sampling and color measurement procedures were similar to those described by Safran and McGraw (2004). A single color measurement was made for each of the four regions, and each measurement included a score of three color metrics, i.e., hue, saturation, and brightness. We averaged the three color metrics over the four body regions for each bird. As average, hue, saturation, and brightness were highly intercorrelated (all r > 0.71 and all p values < 0.001, n = 98 birds), we used principal component analysis to collapse the three variables. A single component (PC1) was extracted, and it explained 87% of the variance in the three color components. A low PC1 score indicates a browner, more saturated, and darker ventral plumage.
Data on both tail streamer length and ventral coloration were obtained from 56 males (18 young and 38 older) out of 66 total males that attempted to breed in the colony in 2004, and all analyses of age effects are restricted to this subset of males. Tail length was also measured for their social mates (n = 56), but feathers were collected only from a subset of them (n = 42), which accounts for the reduced sample size in the analyses of female ventral coloration. No color data were collected in the 2003 breeding season.
Blood samples (2–25 μL) were collected by brachial venipuncture from adults at first capture, and from nestlings 3 days after hatching, and stored in a lysis buffer for subsequent parentage analysis. We assigned paternity using nine polymorphic microsatellite markers. Paternity data in this study represent a subsample of the data reported in Kleven et al. (2005), in which detailed information about the markers, their variability, and paternity determination are presented. Briefly, we extracted DNA with QIAamp® DNA Blood Kit and DNeasy® Tissue Kit (Qiagen, Venlo, the Netherlands) and amplified the microsatellites by polymerase chain reaction (PCR). PCR products were run on an ABI 3100 sequencer (Applied Biosystems, Foster City, USA), and the fragment sizes (alleles) were analyzed with GeneMapper v3.0 (Applied Biosystems). The combined exclusion probability (Jamieson 1994) for the nine markers was higher than 0.9999. We were able to identify the sire (within-pair or extrapair) of all young (249 chicks from 57 broods in 2003; 354 from 82 broods in 2004) in the focal colony from a complete match with the paternally inherited allele at all markers, allowing for a few cases of single slippage mutations at two hypermutable loci (HrU6 and HrU10; Brohede et al. 2002; Anmarkrud et al. 2008).
Paternity was assigned to all young in 53 first broods and 4 renestings in 2003, and to all young in 58 first broods and 24 renestings or second broods in 2004. The two study years were very similar with respect to brood sizes (mean brood sizes of 4.5 ± 1.2 and 4.6 ± 1.1 young for first broods in 2003 and 2004, respectively) and the proportion of young sired by extrapair males (33% and 30%, respectively). Hence, our longitudinal analysis of male fertilization success across the study of 2 years should not be confounded by particular year effects.
Means ± 1SD are reported, unless otherwise stated. In comparisons of means, we applied the unequal variance t test (Ruxton 2006) when variances differed significantly.
Results
Sex and age differences in tail length and ventral coloration
In 2004, males had longer tails than females (87.7 ± 7.3 mm versus 76.5 ± 4.8 mm) and a browner ventral plumage (PC1: −0.39 ± 0.90 versus 0.53 ± 0.90). In both sexes, older individuals had longer tail streamers and a browner ventral plumage than young ones (Table 1; see also mean values for males in Table 2). Hence, both traits were sexually dimorphic and showed age-related expression. However, there was no significant correlation between the two traits, in either males (Pearson r = −0.09, n = 56, p = 0.63) or females (Pearson r = −0.08, n = 42, p = 0.60). This means that birds with long tails did not differ in ventral coloration from those with shorter tails, and that there was considerable overlap between age groups in both traits.
A within-individual increase in tail length was also evident in the longitudinal data set of 40 males that were present in both the 2003 and the 2004 breeding seasons. Their mean tail length increased from 87.1 ± 6.5 mm in 2003 to 89.4 ± 7.4 mm the following year (paired t test: t 39 = −3.74, p = 0.0006). There was no significant differences in tail length between breeders in the 2003 season that returned in 2004 and those that did not (males: 87.4 ± 6.6 mm vs. 86.8 ± 7.1 mm, Student’s t 57 = 0.33, p = 0.75; females: 76.3 ± 3.9 mm versus 75.6 ± 4.3 mm, Student’s t 53 = 0.62, p = 0.53).
Male age and fertilization success
The range in first egg dates (breeding onset) spanned from 9 May to 5 July (26 May ± 13.5 days, n = 56 males) in 2004. Older males started to breed almost 2 weeks before young males (Table 2). They also tended to pair with older females, and young males similarly paired with young females (χ 21 = 6.13, n = 56, p = 0.013). However, there was no evidence of assortative pairing for tail length (Pearson r = −0.01, n = 56, p = 0.96) or ventral coloration (Pearson r = −0.12, n = 42, p = 0.44). Earlier breeding onset for older males was also associated with a higher frequency of second broods, as 22 of 38 older males initiated a second nesting attempt, but only 2 of 18 young males did so (χ 21 = 10.9, p < 0.001).
The number of offspring sired in the colony varied considerably among individual males. In the 2004 season, male total fertilization success, i.e., offspring sired in own nests and through extrapair fertilizations, ranged from 0 to 20 offspring (mean = 5.8 ± 4.2 offspring, n = 6). Older males sired more than twice as many offspring as young males (Table 2). The difference was statistically significant both for the number of within-pair offspring and for the number of extrapair offspring analyzed separately (Table 2).
The increase in fertilization success with age was also true in the longitudinal data set of 30 males that bred in both years (Table 3). Mean number of young sired per male increased from 3.9 in 2003 to 5.8 in 2004. When the number of offspring was broken down into within-pair and extrapair young, the increase was significant for within-pair young, and nearly so for extrapair young (Table 3). These comparisons were restricted to first broods only since paternity data from 2003 was only available for first broods. The 30 males also started to breed on average of 3 days earlier in 2004 than in 2003, but this difference was not statistically significant (Table 3).
Male traits, age, and fertilization success
Because older males had longer tails and a browner and darker ventral plumage than young males, and older males also sired more offspring than young males, a correlation between trait expression and number of fertilizations would be expected for both traits when analyzed across all males. This was true for tail length, which correlated positively with total fertilization success (Pearson r = 0.40, n = 56, p = 0.02), but not for ventral coloration (r = −0.16, n = 56, p = 0.24). When the correlations were broken down by age groups, there was a positive correlation between tail length and fertilization success for older males, but not for young males (Fig. 2e). The same contrast between age groups was evident for within-pair (Fig. 2a) and extrapair success (Fig 2c) analyzed separately, though the positive correlation between tail length and number of extrapair offspring was not statistically significant for older males. For the relationship between ventral coloration and total number of fertilizations, there were no significant correlations within either age group (Fig. 2f), nor were any age-specific correlations significant when within-pair and extrapair fertilizations were analyzed separately (Fig. 2b, d).

Tail length and ventral coloration in relation to fertilization success throughout the breeding season (i.e., including second broods) of young (n = 18 males) and older (n = 38 males) barn swallows. a Tail length and the number of within-pair young sired (young: r = −0.05, p = 0.85; older: r = 0.36, p = 0.026). b Ventral coloration and number of within-pair young sired (young: r = 0.22, p = 0.38; older: r = 0.05, p = .76). c Tail length and number of extrapair young sired (young: r s = 0.12, p = 0.64; older: r s = 0.25, p = 0.14). d Ventral coloration and number of extrapair young sired (young: r s = −0.35, p = 0.16; older: r s = −0.24, p = 0.15). e Tail length and total number of young sired (young: r = −0.01, p = 0.97; older: r = 0.34, p = 0.035). f Ventral coloration and total number of young sired (young: r = 0.14, p = 0.58; older: r = −0.01, p = 0.56). Low color scores indicate a browner, more saturated, and darker plumage. Male age is shown by different symbols (open circles young; closed circles older), and a regression line is indicated for young (dashed line) and older males (solid line) separately
In a multivariate analysis of total fertilization success, in which both ornamental traits and male age were included as predictor variables, male age and tail length, but not ventral coloration, significantly explained variation in male total fertilization success (Table 4).
Discussion
Our results can be summarized in three major findings. First, age was a strong predictor of male fertilization success, as older males bred earlier in the season, were more likely to have a second brood, were better at securing paternity, and gained more extrapair paternity throughout the breeding season than yearlings. These age effects were also evident for individual males from 1 year to the next. Second, tail length and ventral coloration showed age-related expression in both sexes, but only tail length was correlated with fertilization success. Third, tail length was a significant predictor of male fertilization success in older males, but not in young males. Collectively, these results are consistent with an age-related, life-history effect on male fertilization success, but provide no support for sexual selection on ventral coloration and only mixed support for sexual selection on tail length.
Our finding that male age was positively correlated with within-pair and extrapair fertilization success in a species with an extensive extrapair mating system is neither novel nor surprising, as several previous studies of other species have shown such a relationship for either one or both components of total fertilization success (reviewed in Møller and Ninni 1998; Griffith et al. 2002; Schmoll et al. 2007). Yet, it is worth noting that the age effect was particularly strong for extrapair fertilization success. Young males had almost no success in extrapair mating; nearly all extrapair offspring were sired by older males. A similar tendency for a stronger age effect on extrapair paternity than on within-pair paternity has been reported in several passerines (e.g., Wetton et al. 1995; Johnsen et al. 2001; Kleven et al. 2006c; Schmoll et al. 2007) and may be a general phenomenon in socially monogamous species with frequent extrapair mating. This differential age effect is most likely explained by male factors, at least at a proximate level. In a number of species, older males seem to prioritize extrapair mating activities over mate guarding their females, while young males do the opposite (Wagner et al. 1996; Johnsen et al. 2003; Kleven et al. 2006c). The reason why older males still suffer relatively little paternity loss despite relaxed mate guarding may be explained by a better performance in sperm competition due to higher sperm production capacity (Laskemoen et al. 2010). Analyses of individual variation in testis size in many passerine species have revealed that young males generally have smaller testes than older males (Laskemoen et al. 2008, and references therein). This is also true for barn swallows (own unpublished data). Anyway, the strong age effect in extrapair fertilization success emphasizes the value of identifying extrapair sires in parentage studies. Many parentage studies suffer from incomplete sampling of potential extrapair sires, or the genotyping system (e.g., multilocus minisatellite fingerprinting or too few microsatellite markers) is not very powerful in detecting the true sire among many candidates and can only be used for paternity exclusion purposes. Such studies may easily have underestimated male age effects in extrapair mating systems.
An increase in tail length with age is well-known from several studies of European barn swallows (Møller 1991; Møller and de Lope 1999; Turner 2006). Tail length is shorter, on average, in American than in European barn swallows (Turner 2006), but the same general patterns of sexual dimorphism and age dependency exist. For ventral coloration, the situation is somewhat different, as European barn swallows have a largely whitish ventral plumage that is apparently not as sexually dichromatic as the rufous belly and vent in American barn swallows (Safran and McGraw 2004). However, European barn swallows do have a reddish throat, and this trait is darker (lower brightness) in males than in females, and becomes darker with age in both sexes (Galván and Møller 2009). This is basically the same pattern that we have found for the entire ventral coloration (throat, breast, belly, and vent) in our study of North-American birds. Interestingly, color expression did not correlate with tail length, as also found by Safran and McGraw (2004). This is surprising, especially since both traits show age-dependent expression (this study) and patterns of condition dependency (tail length: Møller 1994; Møller and de Lope 1999, ventral coloration: Safran et al. 2008). Hence, they must signal fundamentally different qualities. Clearly, the two traits have different metabolic pathways and may well serve different adaptive functions (cf. Andersson et al. 2002).
A more fundamental question underlying our analysis and interpretation is how the two sexually dimorphic traits have evolved and are maintained in the population. More precisely, the question is whether the sexual dimorphisms are due to current sexual selection (Westneat 2006). Here, we arrive at some interesting contrasts with previous studies. We found no evidence that ventral plumage coloration is under current sexual selection. However, experimental color enhancement of the plumage has clearly demonstrated a positive effect on male fertilization success in a New York population (Safran et al. 2005), presumably mediated through elevated androgen levels (Safran et al. 2008). We are currently unable to explain this apparent paradox. One possibility is that the plumage manipulation caused males to alter their reproductive behavior and thereby secure more fertilizations (Saino et al. 2003). Then, male behavior is the key trait under sexual selection, and plumage color per se may not be a trait preferred by females. Alternatively, there are temporal or spatial variations in the intensity of sexual selection on this trait (Westneat 2006; Chaine and Lyon 2009), which means that our negative results from one season and one colony may not be representative for the North American population at large. Clearly, more comprehensive studies are needed to resolve this enigma.
For tail length, there was an overall positive correlation with fertilization success, as well as within the group of older males. At first glance, this may be taken as evidence in support of sexual selection. However, exclusive support for sexual selection requires a positive correlation between the trait and fertilization success also within separate age classes (Fig. 1). The group of older males (defined as return breeders from the previous year) presumably consists of several age classes, as barn swallows may breed for up to five or six seasons (Møller and de Lope 1999; Turner 2006). If both tail length and fertilization success increase beyond the second year of breeding, then the correlation observed for older males may be fully consistent with the life-history model for separate age classes (cf. Fig. 1). In European barn swallows, tail length continues to increase with age after the second breeding year (Møller 1994; Møller and de Lope 1999). To our knowledge, there is currently no evidence to indicate whether fertilization success shows a similar increase. The lack of correlation between tail length and fertilization success in young males might be interpreted more directly as lack of support for the sexual selection model. This group is probably a more homogenous age class, consisting of 1-year-old, first-time breeders. Barn swallows normally start breeding at the age of 1 year, away from their natal colony, but show strong breeding site fidelity once they have settled as breeders (Møller 1994; Turner 2006; Galván and Møller 2009). Because we had ringed nearly all breeding adults in the colony in the previous year, newcomers in 2004 must have been almost exclusively 1 year olds.
The results of the tail manipulation experiment on American barn swallows in the same study area by Smith et al. (1991) can also be taken as evidence against a role of sexual selection. They found that male fertilization success was not affected by tail elongation or shortening, but correlated significantly with the male’s tail length before manipulation. Thus, tail length did not seem to be a signal for female choice, and males with an originally long tail may have had higher fertilization success because they were older. There are also experimental studies of the aerodynamic properties of the long tail streamers in European barn swallows, which conclude that the trait is only under natural selection for optimal flight performance (e.g. Bro-Jørgensen et al. 2007). At present, we are not able to confirm or reject a role of sexual selection for the evolution of male tail length in our study population. For a full evaluation of the issue, we need more age-specific data on tail length and fertilization success. More direct evidence for a role of female preferences, for example, through experimental trait manipulation combined with observations of sexual behavior in the two sexes, should also be sought.
Regardless of the role of ornamental traits, our study has clearly shown that male age predicts fertilization success in American barn swallows. This pattern also demands an adaptive explanation. Does natural selection alone shape male allocation of reproductive effort between breeding seasons, or does female choice somehow also play a role? Females could choose older males in social mate choice, in sexual (extrapair) mate choice, or both. It is possible that females obtain direct reproductive benefits by choosing an older male as a breeding partner, because older males are superior in many phenotypic respects, at least up to 5 years of age (Møller and de Lope 1999), and thus might be better parents. In extrapair mate choice, however, females receive only sperm, and direct reproductive benefits seem therefore less likely. Instead, older males may deliver better genes (Brooks and Kemp 2001). If so, offspring sired by older males should perform better than offspring sired by young males. We are not aware of any studies that have conclusively shown a paternal genetic benefit from a female extrapair mating preference for older males.
We instead argue that male age-related fertilization success follows a common pattern in birds of age-related increases in reproductive effort in both sexes (Sæther 1990). For females, this is often expressed in elevated clutch size after the first year of breeding (Klomp 1970). For males, it is manifested in delayed plumage maturation (Selander 1972), larger testes (Laskemoen et al. 2008), earlier arrival on the breeding grounds (Ketterson and Nolan 1983; Stewart et al. 2002), and increased fertilization success (Griffith et al. 2002, this study). The phenomenon can be seen as a life-history strategy of optimal allocation of reproductive effort between breeding events (Stearns 1976; Lessells 1991), where reduced effort the first year can be adaptive through enhanced success in the future. Increased efforts later in life can also be associated with more experience and higher capacity for reproduction. Hence, age-related male fertilization success may be viewed as a naturally selected reproductive trait in males, like clutch size in females. Our results of age-dependent fertilization success also suggest that age effects should be taken into account when estimating the opportunity for sexual selection (e.g., Freeman-Gallant et al. 2005; Whittingham and Dunn 2005; Kleven et al. 2006a), because sexual selection can only result from variation in lifetime fitness (Schmoll et al. 2007).
We fully agree with Westneat and Stewart (2003) in their claim that many studies on avian extrapair mating behavior jump to conclusions about female choice without a proper consideration of viable alternatives based on male factors. It is principally wrong to assume by default that male fertilization success results from mechanisms of sexual selection, like female preferences, since there is also intraindividual variation in male fertilization success related to age, as explained by life-history theory. It is quite striking and thought-provoking that there is not yet any published information on longitudinal variation in male fertilization success in such a well-studied species like the barn swallow. Likewise, many paternity studies in birds focus exclusively on sex traits and sexual selection, and ignore life-history theory. Our study may serve as an illustration of the complex task of understanding the evolution of avian extrapair mating systems and sexually dimorphic traits from purely correlative evidence.
References
Andersson M (1994) Sexual selection. Princeton University Press, Princeton
Andersson S, Pryke SR, Ornborg J, Lawes MJ, Andersson M (2002) Multiple receivers, multiple ornaments, and a trade-off between agonistic and epigamic signaling in a widowbird. Am Nat 160:683–691
Anmarkrud J, Kleven O, Bachmann L, Lifjeld JT (2008) Microsatellite evolution: mutations, sequence variation, and homoplasy in the hypervariable avian microsatellite locus HrU10. BMC Evol Biol 8:138
Arnqvist G, Kirkpatrick M (2005) The evolution of infidelity in socially monogamous passerines: the strength of direct and indirect selection on extra-pair copulation behavior in females. Am Nat 165:S26–S37
Balbontín J, Hermosell IG, Marzal A, Reviriego M, De Lope F, Møller AP (2007) Age-related change in breeding performance in early life is associated with an increase in competence in the migratory barn swallow Hirundo rustica. J Anim Ecol 76:915–925
Bennett PM, Owens IPF (2002) Evolutionary ecology of birds: life histories, mating systems and extinction. Oxford University Press, Oxford
Brohede J, Primmer CR, Moller A, Ellegren H (2002) Heterogeneity in the rate and pattern of germline mutation at individual microsatellite loci. Nucl Acids Res 30:1997–2003
Bro-Jørgensen J, Johnstone RA, Evans MR (2007) Uninformative exaggeration of male sexual ornaments in barn swallows. Curr Biol 17:850–855
Brooks R, Kemp DJ (2001) Can older males deliver the good genes? Trends Ecol Evol 16:308–313
Chaine AS, Lyon BE (2009) Adaptive plasticity in female mate choice dampens sexual selection on male ornaments in the lark bunting. Science 324:1143–1143
Darwin C (1871) The descent of man, and selection in relation to sex. John Murray, London
Dor R, Safran RJ, Sheldon FH, Winkler DW, Lovette IJ (2010) Phylogeny of the genus Hirundo and the barn swallow subspecies complex. Mol Phyl Evol 56:409–418
Freeman-Gallant CR, Wheelwright NT, Meiklejohn KE, States SL, Sollecito SV (2005) Little effect of extrapair paternity on the opportunity for sexual selection in savannah sparrows (Passerculus sandwichensis). Evolution 59:422–430
Galván I, Møller AP (2009) Different roles of natural and sexual selection on senescence of plumage colour in the barn swallow. Funct Ecol 23:302–309
Griffith SC (2007) The evolution of infidelity in socially monogamous passerines: neglected components of direct and indirect selection. Am Nat 169:274–281
Griffith SC, Owens IPF, Thuman KA (2002) Extrapair paternity in birds: a review of interspecific variation and adaptive function. Mol Ecol 11:2195–2212
Jamieson A (1994) The effectiveness of using co-dominant polymorphic allelic series for (1) checking pedigrees and (2) distinguishing full-sib pair members. Anim Genet 25:37–44
Johnsen A, Lifjeld JT, Andersson S, Örnborg J, Amundsen T (2001) Male characteristics and fertilisation success in bluethroats. Behaviour 138:1371–1390
Johnsen A, Lifjeld JT, Krokene C (2003) Age-related variation in mate-guarding intensity in the bluethroat (Luscinia s. svecica). Ethology 109:147–158
Ketterson ED, Nolan JV (1983) The evolution of differential bird migration. Curr Ornithol 1:357–402
Kleven O, Jacobsen F, Robertson RJ, Lifjeld JT (2005) Extrapair mating between relatives in the barn swallow: a role for kin selection? Biol Lett 1:389–392
Kleven O, Jacobsen F, Izadnegahdar R, Robertson R, Lifjeld J (2006a) Male tail streamer length predicts fertilization success in the North American barn swallow (Hirundo rustica erythrogaster). Behav Ecol Sociobiol 59:412–418
Kleven O, Jacobsen F, Izadnegahdar R, Robertson RJ, Lifjeld JT (2006b) No evidence of paternal genetic contribution to nestling cell-mediated immunity in the North American barn swallow. Anim Behav 71:839–845
Kleven O, Marthinsen G, Lifjeld J (2006c) Male extraterritorial forays, age and paternity in the socially monogamous reed bunting (Emberiza schoeniclus). J Ornithol 147:468–473
Klomp H (1970) The determination of clutch size in birds: a review. Ardea 58:1–124
Kojima W, Kitamura W, Kitajima S, Ito Y, Ueda K, Fujita G, Higuchi H (2009) Female barn swallows gain indirect but not direct benefits through social mate choice. Ethology 115:939–947
Laskemoen T, Fossøy F, Rudolfsen G, Lifjeld JT (2008) Age-related variation in primary sexual characters in a passerine with male age-related fertilization success, the bluethroat Luscinia svecica. J Avian Biol 39:322–328
Laskemoen T, Kleven O, Fossøy F, Robertson R, Rudolfsen G, Lifjeld J (2010) Sperm quantity and quality effects on fertilization success in a highly promiscuous passerine, the tree swallow Tachycineta bicolor. Behav Ecol Sociobiol 64:1473–1483
Lessells CM (1991) The evolution of life histories. In: Krebs JR, Davies NB (eds) Behavioral ecology. An evolutionary approach, 3rd edn. Blackwell, Oxford, pp 32–68
Manning JT (1985) Choosy females and correlates of male age. J Theor Biol 116:349–354
Møller AP (1988) Female choice selects for male sexual tail ornaments in the monogamous swallow. Nature 332:640–642
Møller AP (1991) Sexual selection in the monogamous barn swallow (Hirundo rustica): I. Determinants of tail ornament size. Evolution 45:1823–1836
Møller AP (1994) Sexual selection and the barn swallow. Oxford University Press, Oxford
Møller AP, Balbontín J, Cuervo JJ, Hermosell IG, de Lope F (2009) Individual differences in protandry, sexual selection, and fitness. Behav Ecol 20:433–440
Møller AP, Brohede J, Cuervo JJ, de Lope F, Primmer CR (2003) Extrapair paternity in relation to sexual ornamentation, arrival date, and condition in a migratory bird. Behav Ecol 14:707–712
Møller AP, de Lope F (1999) Senescence in a short-lived migratory bird: age-dependent morphology, migration, reproduction and parasitism. J Anim Ecol 68:163–171
Møller AP, Ninni P (1998) Sperm competition and sexual selection: a meta-analysis of paternity studies of birds. Behav Ecol Sociobiol 43:345–358
Møller AP, Saino N, Taramino G, Galeotti P, Ferrario S (1998) Paternity and multiple signaling: effects of a secondary sexual character and song on paternity in the barn swallow. Am Nat 151:236–242
Møller AP, Szép T (2002) Survival rate of adult barn swallows Hirundo rustica in relation to sexual selection and reproduction. Ecology 83:2220–2228
Møller AP, Tegelström H (1997) Extra-pair paternity and tail ornamentation in the barn swallow Hirundo rustica. Behav Ecol Sociobiol 41:353–360
Neuman CR, Safran RJ, Lovette IJ (2007) Male tail streamer length does not predict apparent or genetic reproductive success in North American barn swallows Hirundo rustica erythrogaster. J Avian Biol 38:28–36
Ruxton GD (2006) The unequal variance t-test is an underused alternative to Student's t-test and the Mann-Whitney U test. Behav Ecol 17:688–690
Safran RJ (2004) Adaptive site selection rules and variation in group size of barn swallows: individual decisions predict population patterns. Amer Natur 164:121–131
Safran RJ, Adelman JS, McGraw KJ, Hau M (2008) Sexual signal exaggeration affects physiological state in male barn swallows. Curr Biol 18:R461–R462
Safran RJ, McGraw KJ (2004) Plumage coloration, not length or symmetry of tail-streamers is a sexually selected trait in North American barn swallows. Behav Ecol 15:455–461
Safran RJ, Neuman CR, McGraw KJ, Lovette IJ (2005) Dynamic paternity allocation as a function of male plumage color in barn swallows. Science 309:2210–2212
Saino N, Primmer CR, Ellegren H, Møller AP (1997) An experimental study of paternity and tail ornamentation in the barn swallow (Hirundo rustica). Evolution 51:562–570
Saino N, Romano M, Sacchi R, Ninni P, Galeotti P, Møller AP (2003) Do male barn swallows (Hirundo rustica) experience a trade-off between the expression of multiple sexual signals? Behav Ecol Sociobiol 54:465–471
Sæther BE (1990) Age-specific variation in reproductive performance of birds. Curr Ornithol 7:251–283
Schmoll T, Mund V, Dietrich-Bischoff V, Winkel W, Lubjuhn T (2007) Male age predicts extrapair and total fertilization success in the socially monogamous coal tit. Behav Ecol 18:1073–1081
Selander RK (1972) Sexual selection and dimorphism in birds. In: Campbell B (ed) Sexual selection and the descent of man 1871–1971. Aldine, Chicago, pp 180–230
Shields WM (1984) Factors affecting nest and site fidelity in Adirondack barn swallows (Hirundo rustica). Auk 101:780–789
Smith HG, Montgomerie R, Põldmaa T, White BN, Boag PT (1991) DNA fingerprinting reveals relation between tail ornaments and cuckoldry in barn swallows, Hirundo rustica. Behav Ecol 2:90–98
Stearns SC (1976) Life-history tactics—review of ideas. Q Rev Biol 51:3–47
Stearns SC (1992) The evolution of life histories. Oxford University Press, Oxford
Stewart RLM, Francis CM, Massey C (2002) Age-related differential timing of spring migration within sexes in passerines. Wilson Bull 114:264–271
Turner A (2006) The barn swallow. T & AD Poyser, London
Wagner RH, Shug MD, Morton ES (1996) Condition-dependent control of paternity by female purple martins: implications for coloniality. Behav Ecol Sociobiol 38:379–389
Westneat DF (2006) No evidence of current sexual selection on sexually dimorphic traits in a bird with high variance in mating success. Am Nat 167:E171–E189
Westneat DF, Stewart IK (2003) Extra-pair paternity in birds: causes, correlates and conflict. Annu Rev Ecol Evol Syst 34:365–396
Wetton JH, Burke T, Parkin DT, Cairns E (1995) Single-locus DNA fingerprinting reveals that male reproductive success increases with age through extra-pair paternity in the house sparrow (Passer domesticus). Proc R Soc B 260:91–98
Whittingham LA, Dunn PO (2005) Effects of extra-pair and within-pair reproductive success on the opportunity for selection in birds. Behav Ecol 16:138–144
Acknowledgments
We thank Diana and Bob Trainor for kindly giving us access to their property, the staff at Queen’s University Biological Station for all help and support, Rasa Izadnegahdar and Sylvia Wood for assistance with the fieldwork, and Arild Johnsen and Neil Metcalfe for helpful comments on the manuscript. This research was funded by the Research Council of Norway, a grant from Queen’s University, a grant from Natural Sciences and Engineering Research Council of Canada, and a grant from William C. Brown Research Fund.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by J. Lindström
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Lifjeld, J.T., Kleven, O., Jacobsen, F. et al. Age before beauty? Relationships between fertilization success and age-dependent ornaments in barn swallows. Behav Ecol Sociobiol 65, 1687–1697 (2011). https://doi.org/10.1007/s00265-011-1176-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00265-011-1176-4