
Abstract
New species of a gecko of the genus Euleptes is described here—E. klembarai. The material comes from the middle Miocene (Astaracian, MN 6) of Slovakia, more precisely from the well-known locality called Zapfe`s fissure fillings (Devínska Nová Ves, Bratislava). The fossil material consists of isolated left maxilla, right dentary, right pterygoid and cervical and dorsal vertebrae. The currently known fossil record suggests that isolation of environment of the Zapfe`s fissure site, created a refugium for the genus Euleptes in Central Europe (today, this taxon still inhabits southern part of Europe and North Africa—E. europea), probably resulting from the island geography of this area during the middle Miocene. The isolation of this territory might have facilitated allopatric speciation.
Similar content being viewed by others


1 Introduction
Gekkota (geckos and pygopods) is a speciose clade of lepidosaurs, comprising more than 1600 extant species (Bauer 2013; Uetz and Freed 2017). However, the fossil record of these successful and cosmopolitan lizards is generally poorly represented, chiefly by isolated skull elements and vertebrae (Estes 1983; Müller and Mödden 2001; Augé 2005; Daza et al. 2014), although some of the oldest stem-gekkotans (Norellius and Gobekko Daza et al. 2013a; Conrad and Daza 2015) include articulated and complete skulls, and gekkotans in amber from the Cretaceous of Myanmar and the Miocene of Dominican Republic represent some astonishing specimens with superb preservation of skeletal and soft tissue (Böhme 1984; Daza and Bauer 2012; Daza et al. 2013b, 2016). Very important and superbly preserved find in Baltic amber is represented by Yantarogecko balticus from the Early Eocene of north–western Russia (Bauer et al. 2005). The bias towards selectively preserving some bones (frontal, maxilla, dentary, presacral vertebrae) most likely reflects the lightly built skeleton (Evans 2003, 2008) and also seems related to the propensity of some bones to resist decomposition or digestion (Kupriyanov et al. 2012). Unfortunately, this results in significant gaps in our knowledge of the evolution of this clade. This is especially true of material from the Paleogene, which is especially fragmentary (Estes 1983; Daza et al. 2014).
In Europe the Tertiary gekkotan fossil record is more complete than in most other areas of the world (Daza et al. 2014), yet it is represented almost exclusively by isolated skull bones and was studied only occasionally. Among published reports, only several specimens were assigned to the sphaerodactylid genus Euleptes from the Miocene of Europe, including the species E. gallica (Müller 2001; Čerňanský and Bauer 2010) and unnamed material from Germany and Slovakia (Estes 1969; Müller and Mödden 2001). Extinct genera are represented by Gerandogekko which has been identified as closely related to Euleptes (Daza et al. 2014) [including two species: G. arambourgi and G. gaillardi (Hoffstetter 1946; Kluge 1967; Schleich 1985)], and Palaeogekko risgovienis from Germany (Schleich 1987). Besides, there is also material from several sites from the Oligocene and Miocene of Germany and Miocene of France and Austria (Augé and Rage 2000; Čerňanský 2016; Čerňanský et al. 2015, 2016; Colombero et al. 2017), that might be part of Euleptes.
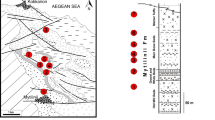
One of the best morphological descriptions of Euleptes material is based on specimens from the Neogene of Devínska Nová Ves (now a part of Bratislava, Slovakia; the site is also known as Neudorf an der March). The material was identified as a gecko by Estes (1969; Fig. 1a–c) who described it as cf. Phyllodactylus sp. A subsequent review of the genus Phyllodactylus restricted the species from Europe to the genus Euleptes (Bauer et al. 1997). The fossil comes from the locality called Zapfe`s fissure fillings, well known because of its rich mammal fauna, e.g. Pliopithecus vindobonensis or Chalicotherium (see e.g., Zapfe 1958, 1979). The fissures are often filled by the sinter (cave sediments) “terra rossa” and “terra fusca” (Mišík 1976). The locality is situated at the northern margin of the Vienna Basin on the northern slope of Devínska Kobyla Hill in the Stockerau limestone pit and it is dated to the middle Miocene, more precisely the Astaracian, i.e. the uppermost part of the Middle Badenian (Neogene Mammal Zone MN 6; see e.g., Cícha et al. 1972; Fejfar 1974). The previously known material includes a left maxilla and fragment of the anterior portion of the left dentary (Estes 1983). In the description of this material, Estes (1969) recognized a slightly taller and more posteriorly extended facial process of the maxilla than that in the extant Euleptes europaea. Daza et al. (2014) mentioned the allocation of this material to the genus Euleptes, however, they called for a careful revision of this material in order to resolve its species allocation. In this paper we describe new material from this locality and review the material previously reported by Estes (1969). The new information indicates that the gecko from Devínska Nová Ves represents a species distinct from all known species of Euleptes and closely related genera.

Euleptes klembarai sp. nov.: the holotypic maxilla NHMW 1977/1865; a actual damaged specimen, b detail of the anterior preserved region. Reconstruction of the maxilla in c lateral and d medial aspect
2 Materials and methods
The material studied here is housed in the Natural History Museum Vienna in Austria. The material consists of isolated left maxilla, right dentary, right pterygoid and cervical and dorsal vertebrae collected by screen-washing from the middle Miocene Zapfe`s fissure fillings. The specimens were imaged using a scanning electron microscope (SEM) at the Slovak Academy of Sciences and with a Leica M125 binocular microscope with axially mounted DFC500 camera; software: LAS (Leica Application Suite) version 4.1.0 (build 1264). The image processing program ImageJ (Schneider et al. 2012) was used for measurements. Measurements of bones for ten specimens of E. europaea were taken from digital X-rays and High Resolution X-ray Tomographies. Digital radiographs were obtained at the National Museum of Natural History, Smithsonian Institution in Washington DC, using a Kevex TM PXS10-16W X-ray source and Varian Amorphous Silicon Digital X-Ray Detector PaxScanH 4030R set to 130 kV at 81 mA. For each X-ray, linear and pseudofilm filters were used. High Resolution X-Ray CT scans were obtained from one specimen using an XradiaMicroCT scanner (Pleasanton, CA, USA) at the Department of Geological Sciences of the University of Texas at Austin using a 4 × detectorobjective and an X-ray source set at 80 kV and 10 W. 3D models were generated using Avizo Lite 9.0.0 (Visualization Sciences Group). The standard anatomical orientation system is used throughout this article.
Unfortunately while obtaining SEM images of the maxilla and dentary, due to a failure of the equipment, the maxilla and dentary were severely damaged by a technician from the Slovak Academy of Sciences in Bratislava during the sputter deposition process (the coating head of the machine fell down on the material). In any case we document these images, as they are still the best images available from this material. Diagnostic features of the material are still recognizable in the specimens despite the damage. We have made precise reconstructions based on: (a) previous descriptions including illustrations of maxilla by Estes (1969), (b) the preserved damaged parts of the material, and (c) notes we made on the specimens previous to the incident.
2.1 Codes for institutional collections
NHMW, The Natural History Museum Vienna, Austria.
MCZ, Museum of Comparative Zoology, HarvardUniversity, Cambridge, USA.
USNM, National Museum of Natural History, Smithsonian Institution, Washington DC, USA.
3 Systematic palaeontology
Gekkota Camp, 1923
Sphaerodactylidae Underwood, 1954
Euleptes Fitzinger, 1843
Euleptes klembarai sp. nov. (Figs. 1, 2 and 3)

Euleptes klembarai sp. nov.: right dentary NHMW 1977/1865/0084 in medial view—a real situation, b reconstruction. Right pterygoid NHMW 1977/1865/0113 in c dorsal and d ventral aspect

Euleptes klembarai sp. nov.: cervical vertebra NHMW 1977/1865/0116 (a–e) and dorsal vertebra NHMW 1977/1865/0117 (f–j) in (a, f) lateral, (b, g) dorsal, (c, h) ventral, (d, i) anterior and (e, j) posterior aspects
1969—cf. Phyllodactylus sp. Estes, Fig. 1a–c.
2014—Euleptes sp. Daza et al., Fig. 10f.
Holotype An almost complete left maxilla, NHMW 1977/1865, housed in the Natural History Museum Vienna.
Paratypes right dentary NHMW 1977/1865/0084, right pterygoid NHMW 1977/1865/0113, one cervical vertebra NHMW 1977/1865/0116, four dorsal vertebrae NHMW 1977/1865/0117-20.
Derivation of Specific Name We name this species after the Slovak palaeoherpetologist Jozef Klembara for his valuable contributions to vertebrate palaeontology and squamate morphology.
Locality and Horizon Zapfe`s fissure fillings, Devínska Nová Ves (Bratislava, Slovakia), middle Miocene (Astaracian, MN 6).
Diagnosis A medium sized species of Euleptes (approximated 37.47 mm SVL, see measurements on Table 1) differing from all fossil and extant taxa of this genus in the following combination of features from the maxilla: (1) a rounded rather than angular tip of the anterior margin of the maxilla facial process; (2) a less steeply inclined posterior edge of the facial process; (3) a less marked groove extending posteriorly from the last supralabial foramen than do other Euleptes.
3.1 Description
Maxilla The facial process of the maxilla is subtriangular in shape. The bone is rather elongate, and relatively lightly-built (Fig. 1). It is almost complete, only the ventral mid-region is lacking. The maxilla preserves the complete tooth row, consisting of 29 tooth loci, eight with teeth still attached (four in the anterior and four in the posterior regions). The premaxillary process is bifurcated as in other geckos, having a short and blunt lateral ramus and a slightly more distinct, more medially oriented and slightly more dorsally located medial ramus. Between them, an oval premaxillary notch is present. The supradental shelf is thin, and expands medially. The maxilla attains its highest point at the level of the eleventh tooth position (from the anterior end of the tooth row). In this section, a palatine process is developed. The medial margin of the shelf is slightly convex (bent dorsally). In the anterior region of the facial process, on the medial side there is a fine ridge that runs posterodorsally. This ridge originates from the supradental shelf at the level of the fourth tooth position (from the anterior end of the tooth row). The rounded superior alveolar foramen is located at the level of the thirteenth tooth position (counted from posterior). The facial process is well-preserved, high and trapezoidal in shape. The anterior margin of the facial process gently slopes anteroventrally in a nearly straight line. Its terminus is free and well delimited from the dental portion (pars dentalis) of the maxilla. It forms a triangular tip, typical for Euleptes spp. The end of this tip is rounded and blunt and less acute as in other species.
The lateral surface is pierced by at least six labial foramina of various size in its lower region. There is only a very finely developed, almost indistinct groove running posteriorly from the last preserved foramen. Besides these foramina, two additional small foramina are located above, close to the base of the facial process.
Dentary The dentary is a long and slender bone (Fig. 2a, b), with a slight medial curvature at its anterior end. The alveolar crest is tall, being almost half of tooth height. Teeth rest on the alveolar crest and there are 24 preserved tooth loci, (three teeth are still attached; the total tooth number was probably slightly higher, because the posterior region is broken). The Meckelian canal is closed, and opens into a V shaped notch near the end of the bone. The notch reaches the level of the third preserved tooth position (counted from the posteriormost preserved locus). The dentary is broken posteriorly and both the coronoid and angular processes are missing. The subdental shelf gradually thins anteriorly. The external surface is pierced by five mental foramina. In the posterior region, foramina are antero-posteriorly elongated, whereas more anterior ones are more rounded.
Dentition There are eight functional teeth preserved in the maxilla and three in the lower jaw. The teeth have pleurodont implantation, are conical, slender, straight and distinctly pointed. Resorption pits are located at the tooth bases on the lingual side. The anterior maxillary teeth are larger than the posterior ones. The intervening teeth are missing, therefore it is impossible to know their size, although broken bases of middle teeth indicate that these might have been comparable in size to the anterior teeth.
Pterygoid There is a complete right pterygoid (Fig. 2c, d). The pterygoid is a tri-radiate, nearly “y” shaped element. The palatine process is broad and blunt, and defines a sigmoid anterior border that would form a lateral extension of the suborbital fenestra. The ectopterygoid process is narrow with a pointed anterior end. The process is almost straight. On the ventral surface, there is a transverse crest that runs along the central portion of the ectopterygoid process. The anterior border between the ectopterygoid and palatine processes is laminar and sinuously indented. This ventral surface is very flattened and forms a well-marked depression. Along the margin of this depression, four small foramina are located (three on the medial and one on the lateral side), these foramina might be connected to two foramina that pierce the dorsal surface. The obtuse process (Sensu Klembara 2015) is short and triangular in shape, forming a distinct medially oriented flange. The quadrate process is long and blade-like and curves posterolaterally. Its posterior termination is blunt-ended. The fossa columellae (= epipterygoid fossa) is small and rounded, and it is prolonged posteriorly by a groove along the dorsal surface of the quadrate process.
Cervical vertebra Only one isolated cervical vertebra is preserved (Fig. 3a–e). It is a very small, lightly built, amphicoelous vertebra with the centrum pierced by a notochordal canal. It is a short element (i.e., taller than long). The neural canal is large and heart-shaped. In dorsal aspect, a small notch is present in the anterior mid-region of the arch. Neural spine is low, trapezoidal in shape in lateral aspect. Prezygapophyses are more or less directed anteriorly. They possess roughly triangular articulation facets that are inclined from the horizontal plane at an angle 42°.
Dorsal vertebrae Four vertebrae are preserved. They are small, anteroposteriorly elongated and amphicoelous (Fig. 3f–j). The pre- and postzygapophyses are almost anteriorly and posteriorly directed, bearing roughly triangular articulation facets. The prezygapophyses are more inclined ventrally (30°) relative to those in the cervical vertebra. In dorsal view, the anterior margin of the neural arch between the prezygapophyses develops a deep triangular notch. The neural canal is large, and bean shaped. On the dorsal surface of the low neural arch, a straight dorsal and longitudinal crest is present along the surface of the neural spine. The neural spine does not rise dorsally, but is markedly expanded posteriorly, and extends posteriorly to the same level as the postzygapohyses. In lateral view, there are well-defined synapophyses, which are elliptically shaped.
4 Discussion
The Slovak material described here is allocated to Euleptes on the basis of the following features: (1) the presence of the triangular tip of the anterior margin of the facial process; (2) the dorsal margin of this facial process is smooth, mound shaped, and lacking a pronounced dorsal process (Bauer et al. 1997; Müller and Mödden 2001). This morphology contrasts sharply with the prominent, often pointed process typical of many gekkotans (Daza 2008), including the middle Miocene Palaeogekko risgoviensis (Schleich 1987). Moreover, the material described here shows several unique features (see Diagnosis) and we have decided to erect a new species name E. klembarai; (3) The size of the maxilla is also larger than in the specimens of E. europaea, producing slightly larger estimates for skull length than in the material reviewed of Euleptes (Table 1). The size of this fossil is intermediate between E. europaea and fossil material attributed to E. gallica from the early Miocene of Europe (Daza et al. 2014). There are also several other differences between E. gallica and E. klembarai (for character states in E. gallica, see Müller 2001): (1) 29 tooth positions in E. klembarai, whereas 31–34 are observed in E. gallica; (2) the nasal process is stouter in E. gallica; (3) a more prominently built and markedly more pointed triangular tip of the maxilla in E. gallica; (4) a more steeply inclined posterior edge of the facial process in E. gallica; and (5) the lateral surface of the facial process is clearly more vascularized in E. gallica and E. europaea than in E. klembarai (see Daza et al. 2014). In the other species of Euleptes, a large foramen pierces the posterior surface of the postnarial region of the maxilla, this foramen opens medially high above the medial maxillary ridge, but in E. klembarai this foramen is considerably lower, and opens medially just above the maxillary ridge.
All bones described here are assigned to one species on the basis of great similarity with the morphology presented by the taxon Euleptes, moreover being comparable in size and coming from the same locality. During the middle Miocene, the area of Devínska Nová Ves was part of an archipelago in the western part of the Central Paratethys (Kvaček et al. 2006). Zapfe`s fissure is located in the territory of Devínska Kobyla Hill. This area was an island during this time, isolated from the nearest land by a shallow strait (Sabol and Kováč 2006). The isolation of this territory might have facilitated allopatric speciation. Except for the middle Miocene of Steinberg in Germany (Palaeogekko risgoviensis; see Schleich 1987), the majority of localities of similar age in Central Europe lack Gekkotan material (e.g., Sandelzhausen, Hambach, Litke, Griesbeckerzell; see Böhme 2010; Ivanov and Böhme 2011; Venczel and Hír 2015; Čerňanský et al. 2017). This is in contrast to early Miocene deposits where such material is more abundant (it should be noted that in the middle Miocene of Western Europe, Gerandogekko gaillardi is present, see Daza et al. 2014). The middle Miocene is particularly interesting because of the abrupt end of the Miocene Climatic Optimum (MN 6–MN 7 + 8; Böhme 2003). The squamate fauna of the middle Miocene remained rich, diverse and of tropical type, however many taxa began to progressively disappear from Europe (see Rage 2013). The currently known fossil record suggests that separation of environment of the Zapfe`s fissure site, created a refugium for members of the genus Euleptes in Central Europe, probably resulting from the island geography of this area during the middle Miocene. According to Fejfar and Sabol (2009), the fossil record from the site contains some faunal elements (e.g., Pliopithecus vindobonensis, Keramidomys carpathicus, or Democricetodon vindobonensis) which indicate an isolated character of the assemblage over a critical time period (necessary for the evolution of endemic forms).
References
Augé, M. (2005). Évolution des lézards du Paléogène en Europe. Mémoires du Muséum national d’Histoire naturelle, 192, 1–369.
Augé, M., & Rage, J.-C. (2000). Les Squamates (Reptilia) du Miocène moyen de Sansan. Mémoires du Muséum National d’Histoire Naturelle, 183, 263–313.
Bauer, A. M. (2013). Geckos—the animal answer guide. Baltimore: The Johns Hopkins University Press.
Bauer, A. M., Good, D. A., & Branch, W. R. (1997). The taxonomy of the Southern African leaf-toed geckos (Squamata: Gekkonidae) with a review of old World “Phyllodactylus” and the description of five new genera. Proceedings of the California Academy of Sciences, 49, 447–497.
Bauer, A. M., Böhme, W., & Weitschat, W. (2005). An Early Eocene gecko from Baltic amber and its implications for the evolution of gecko adhesion. Journal of Zoology, 265, 327–332.
Böhme, W. (1984). Erstfund eines fossilien Kugelfingergeckos (Sauria: Gekkonidae: Sphaerodactylinae) aus Dominikanischem Bernstein (Oligozän von Hispaniola, Antillen). Salamandra, 20, 212–220.
Böhme, M. (2003). The Miocene climatic optimum: evidence from ectothermic vertebrates of Central Europe. Palaeogeography Palaeoclimatology Palaeoecology, 195, 389–401.
Böhme, M. (2010). Ectothermic vertebrates (Actinopterygii, Allocaudata, Urodela, Anura, Crocodylia, Squamata) from the Miocene of Sandelzhausen (Germany, Bavaria) and their implications for environment reconstruction and palaeoclimate. Paläontologische Zeitschrift, 8, 3–41.
Čerňanský, A. (2016). Another piece of the puzzle: the first report on the Early Miocene lizard fauna from Austria (Ottnangian, MN 4; Oberdorf locality). Pal Z, 90, 723–746.
Čerňanský, A., & Bauer, A. M. (2010). Euleptes gallica Müller (Squamata: Gekkota: Sphaerodactylidae) from the Lower Miocene of North-West Bohemia, Czech Republic. Folia Zoologica, 59, 323–328.
Čerňanský, A., Rage, J.-C., & Klembara, J. (2015). The early Miocene squamates of Amöneburg (Germany): the first stages of modern squamates in Europe. Journal of Systematic Palaeontology, 13, 97–128.
Čerňanský, A., Klembara, J., & Müller, J. (2016). The new rare record of the late Oligocene lizards and amphisbaenians from Germany and its impact on our knowledge of the European terminal Palaeogene. Palaeobiodiversity and Palaeoenvironments, 96, 559–587.
Čerňanský, A., Szyndlar, Z., & Mörs, T. (2017). Fossil squamate faunas from the Neogene of Hambach (northwestern Germany). Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-016-0252-1.
Cícha, I., Fahlbusch, V., & Fejfar, O. (1972). Die biostratigraphischen Korrelation einiger jungtertiärer Wirbeltierfaunen Mitteleuropas. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 140, 129–145.
Colombero, S., D’Amico, C., Alba, M., Delfino, M., Esu, D., Giuntelli, P., et al. (2017). Late Messinian mollusks and vertebrates from Moncucco Torinese, north-western Italy. Paleoecological and paleoclimatological implications. Palaeontologia Electronica, 20, 1–66.
Conrad, J. L., & Daza, J. D. (2015). Naming and Rediagnosing the Cretaceous Gekkonomorph (Reptilia, Squamata) from Oosh (Ovorkhangai, Mongolia). Journal of Vertebrate Paleontology, 35, e980891.
Daza, J.D. (2008). Cladistic analysis of the Gekkota (Reptilia) by means of craniological data. Ph.D. dissertation, University of Puerto Rico, Rio Piedras.
Daza, J. D., & Bauer, A. M. (2012). A new amber-embedded sphaerodactyl gecko from Hispaniola, with comments on the morphological synapomorphies of the Sphaerodactylidae. Breviora, 529, 1–28.
Daza, J. D., Bauer, A. M., & Snively, E. (2013a). Gobekko cretacicus (Reptilia: Squamata) and its bearing on the interpretation of gekkotan affinities. Zoological Journal of the Linnean Society, 167, 430–448.
Daza, J. D., Bauer, A. M., Wagner, P., & Böhme, W. (2013b). A reconsideration of Sphaerodactylus dommeli Böhme, 1984 (Squamata: Gekkota: Sphaerodactylidae), a Miocene lizard in amber. Journal of Zoological Systematics and Evolutionary Research, 51, 55–63.
Daza, J. D., Bauer, A. M., & Snively, E. (2014). On the Gekkotan fossil record. The Anatomical Record, 297, 433–462.
Daza, J. D., Stanley, E. L., Wagner, P., Bauer, A. M., & Grimaldi, D. A. (2016). Mid-Cretaceous amber fossils illuminate the past diversity of tropical lizards. Science Advances, 2, e1501080.
Estes, R. (1969). Die Fauna der miozänen Spaltenfühlung von Neudorf an der March (ČSSR) (Reptilia, Lacertilia). Österreichische Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Klasse Abteilung I Sitzungsberichte, 178, 77–82.
Estes, R. (1983). Encyclopedia of Paleoherpetology, Part 10 A-Sauria terrestria (p. 249). Amphisbaenia: Gustav Fischer Verlag.
Evans, S. E. (2003). At the feet of the dinosaurs: the early history and radiation of lizards. Biological Reviews, 78, 513–551.
Evans, S. (2008). The skull of Lepidosauria. In C. Gans, A. S. Gaunt, & K. Andler (Eds.), Biology of the Reptilia (Vol. 20, pp. 1–347). Sirsi: Society for the Study of Amphibians and Reptiles.
Fejfar, O. (1974). Die Eomyiden und Cricetiden (Rodentia, Mammalia) des Mioza¨ns der Tschechoslowakei. Palaeontographica, Abteilungen A, 146, 100–180.
Fejfar, O., & Sabol, M. (2009). Middle Miocene Plesiodimylus from the Devínska Nová Ves-Fissures site (western Slovakia). Bulletin of Geosciences, 84, 611–624.
Hoffstetter, P. (1946). Sur les Gekkonidae fossils. Bulletin du Muséum national d’histoire naturelle, 18, 195–203.
Ivanov, M., & Böhme, M. (2011). Snakes from Griesbeckerzell (Langhian, Early Badenian), North Alpine Foreland Basin (Germany), with comments on the evolution of snake faunas in Central Europe during the Miocene Climatic Optimum. Geodiversitas, 33, 411–449.
Klembara, J. (2015). New finds of anguines (Squamata, Anguidae) from the early Miocene of Northwest Bohemia (Czech Republic). Paläontologische Zeitschrift, 89, 171–195.
Kluge, A. G. (1967). Higher taxonomic categories of gekkonid lizards and their evolution. Bulletin of the American Museum of Natural History, 135, 1–60.
Kupriyanov, V. M. S., Daza, J. D., Bauer, A. M., Gaban-Lima, R., Rocha-Brito, G. R., & Höfling, E. (2012). Six species of Amazonian Woodcreepers (Aves: Dendrocolaptidae) preying upon lizards and frogs. Journal of Natural History, 46, 2985–2997.
Kvaček, Z., Kováč, M., Kovar-Eder, J., Doláková, N., Jechorek, H., Parashiv, V., et al. (2006). Miocene evolution of landscape and vegetation in the Central Paratethys. Geologica Carpathica, 57, 295–310.
Mišík, M. (1976). Geological excursions along Slovakia (pp. 1–359). Bratislava: SPN.
Müller, J. (2001). A new fossil species of Euleptes from early Miocene of Montaigu, France (Reptilia, Gekkonidae). Amphibia-Reptilia, 22, 342–347.
Müller, J., & Mödden, C. (2001). A fossil leaf-toed gecko from the Oppenheim/Nierstein Quarry (Lower Miocene, Germany). Journal of Herpetology, 35, 532–537.
Rage, J.-C. (2013). Mesozoic and Cenozoic squamates of Europe. Palaeobiodiversity and Palaeoenvironments, 93, 517–534.
Sabol, M., & Kováč, M. (2006). Badenian palaeoenvironment, faunal succession and biostratigraphy: a case study from northern Vienna Basin, Devínska Nová Ves- Bonanza site (Western Carpathians, Slovakia). Beiträge zur Paläontologie, 30, 415–425.
Schleich, H. H. (1985). Zur Verbreitungtertiärer und quartärer Reptilien und Amphibien: I. Süddeutschland. Münchner Geowissenschaftliche Abhandlungen A Geologie und Paläontologie, 4, 67–149.
Schleich, H. H. (1987). Neue Reptilienfunde aus dem Tertiär Deutschlands 7. Erstnachweis von Geckos aus dem Mittelmiozän Süddeutschlands: Palaeogekko risgoviensisnov. gen. nov. spec. (Reptilia, Sauria, Gekkonidae). Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historischen Geologie, 27, 67–93.
Schneider, C. A., Rasband, W. S., & Eliceiri, K. W. (2012). NIH Image to ImageJ: 25 years of image analysis. Nature Methods, 9, 671–675.
Uetz, P., Freed, P., Jirí Hošek (eds.), The Reptile database, http://www.reptile-database.org. Accessed Jan 2017.
Venczel, M., & Hír, J. (2015). Lissamphibians and squamate reptiles from the early middle Miocene of Litke, Northern Hungary. Geobios, 48, 291–504.
Zapfe, H. (1958). The skeleton of Pliopithecus (Epipliopithecus) vindobonensis Zapfe and Hürzeler. American Journal of Physical Anthropology, 16, 441–455.
Zapfe, H. (1979). Chalicotherium grande (Blainv.) aus der miozanen Spaltenfullung von Neudorfan der March (Devinska Nova Ves), Tschechoslowakei. Neue Denkschriften des Naturhistorischen Museums in Wien, 2, 1–282.
Acknowledgements
The authors are indebted to U. Göhlich (NHMW) for access to material described in this paper. We thank V. Šmatko (Slovak Academy of Sciences) for SEM pictures. We thank K. Tighe, A. Wynn, K. de Queiroz, R. Bell, and R. McDiarmid for access to specimens and equipment at the USNM. We thank M. C. Vallejo and E. Glynne for assistance obtaining digital X-rays. We thank J. Maisano for helping us obtaining HRCT data, and J. Losos and J. Rosado for access to comparative material at MCZ. We also want to thank Jessica A. Maisano (Jackson School of Geosciences, University ofTexas at Austin) for assistance with the CT scans and image processing. We thank J.-C. Rage (Museum national d’Histoirenaturelle Paris) and one anonymous reviewer for their critical reading of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Editorial handling: D. Marty.
Rights and permissions
About this article

Cite this article
Čerňanský, A., Daza, J.D. & Bauer, A.M. Geckos from the middle Miocene of Devínska Nová Ves (Slovakia): new material and a review of the previous record. Swiss J Geosci 111, 183–190 (2018). https://doi.org/10.1007/s00015-017-0292-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00015-017-0292-1