
Abstract
Acid sphingomyelinase (ASMase) is a key initiator of sphingomyelin/ceramide signal transduction activated by many stress stimuli. Over the past two decades, much progress has been made in defining the clinical relevance of sphingomyelin/ceramide signaling in numerous diseases using ASMase knockout mice. Organs that operate this pathway are numerous and the disease states regulated are diverse, with ceramide generation governing injury in tumor, gut, ovary, brain, lung, heart, liver, and during infection. This chapter emphasizes evolutionary conservation of sphingolipid stress signaling and mammalian adaptations that permit transduction of organotypic responses. Recognition that the sphingomyelin/ceramide transducer calibrates extent of tissue injury, ultimately acting as a molecular switch that determines organ fate, is driving development of new pharmacologic concepts and tools to intervene therapeutically.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
- ASMase
- Ceramide-rich platforms (CRPs)
- ASMase Knockout
- Single-dose radiotherapy (SDRT)
- Cancer
- Niemann-Pick Disease (NPD)
- Stroke
- Blood-brain barrier (BBB)
- Acute respiratory distress syndrome (ARDS)
- Cystic Fibrosis (CF)
- Diabetes
- Infections
1 Introduction
1.1 ASMase
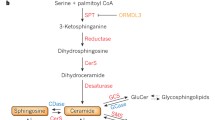
The sphingomyelin pathway is a ubiquitous, evolutionarily conserved signaling system initiated by hydrolysis of sphingomyelin to generate the second messenger ceramide. Sphingomyelinase (SMase) is a specialized form of phospholipase C, which cleaves the phosphodiester bond of sphingomyelin, generating ceramide. Several SMase isoforms can be distinguished by their pH optima, cofactor dependence, and subcellular location. SMases are classified into three groups—acid SMase (ASMase), neutral SMase (NSMase), and alkaline SMase (Alk-SMase) (Kolesnick 2002). While ASMase was originally considered a strictly lysosomal enzyme because of its pH optimum at 4.5–5.0, an ASMase isoform was found within secretory vesicles at the plasma membrane (Liu and Anderson 1995; Schissel et al. 1998a). Subsequent studies showed that ASMase exists in two forms, termed lysosomal ASMase (L-ASMase) and secretory ASMase (S-ASMase), differing in glycosylation pattern and NH2-terminal processing, and consequently in subcellular targeting. ASMase is present in all types of cells, preferentially endowed in endothelium of blood vessels, and cells of the reticuloendothelial system (RES) of liver (Kupffer cells), spleen, bone marrow, lung, as well as macrophages (Otterbach and Stoffel 1995). Activation of ASMase has been demonstrated in response to various unrelated stress stimuli. ASMase is also a key enzyme responsible for ceramide homeostasis.
1.2 Ceramide
Ceramide is a sphingosine-based lipid capable of initiating signaling for numerous stress stimuli, including, but not limited to TNF-α, Fas ligand, ionizing radiation (IR), heat shock, ultraviolet light, and oxidative stress (Fuks et al. 1995; Grassme et al. 2003a; Gulbins 2003; Gulbins et al. 1995; Haimovitz-Friedman et al. 1994; Jarvis et al. 1994; Obeid et al. 1993; Verheij et al. 1996). Structurally, ceramide contains a long-chain sphingoid base backbone, an amide-linked long-chain fatty acid, and a hydroxyl head group. Monounsaturated or saturated fatty acids of various lengths from 2 to 28 carbon atoms are usually found in natural ceramides (Kolesnick et al. 2000). Physical properties of ceramide are determined by length of the fatty acyl chain. Ceramides with long fatty acyl chain (12 carbons or longer) belong to the category of “non-swelling amphiphiles,” as they cannot give rise to micelles or other aggregates in aqueous suspension and hence cannot serve as detergents (Small 1970). In contrast, short-chain ceramides swell in water, a property that limits their utility in research (Stancevic and Kolesnick 2010). In mammalian cells, ceramides usually contain fatty acyl chains of 16–24 carbon atoms. Depending on stimulus and cell type, ceramide can be generated either through SMase-dependent hydrolysis of sphingomyelin or a de novo synthetic pathway. SMase-mediated ceramide generation is usually a rapid event localized in most cells at the plasma membrane, while de novo synthesis of ceramide occurs in a more prolonged fashion, exclusively intracellularly within endoplasmic reticulum or mitochondrial-associated membrane (MAM) (Shimeno et al. 1995; Bionda et al. 2004; Kirschnek et al. 2000; Schissel et al. 1998b). As a second messenger, ceramide plays a ubiquitous role in diverse biologic processes (Kolesnick and Hannun 1999). Accumulating evidence suggests that changes in local membrane structure induced by ceramide elevation are essential for its biological function. Once generated, ceramide may amass or be converted into a variety of metabolites that serve as bioeffector molecules, often inducing proliferation, and except for sphingosine and GD3 usually do not signal cell death. Further, some cells manifest enhanced conversion of ceramide to S1P. Balance between pro-apoptotic properties of ceramide and anti-apoptotic properties of S1P has been referred to as the “S1P Rheostat” (Cuvillier et al. 1996) and may under some circumstances play a crucial role in determining outcome of stress damage in tissues.
1.3 Mechanism of Sphingomyelin/Ceramide Signaling
In 1987, Paley et al. reported that 1,2-diacylglycerols induce SMase activation in GH3 pituitary cells and suggested for the first time that a sphingomyelin-based signaling pathway may be active in this response (Kolesnick and Paley 1987). Subsequently, Okazaki et al. (1989, 1990) confirmed this notion, demonstrating that receptor binding of vitamin D3 can activate a sphingomyelin/ceramide signaling pathway in HL-60 cells. Over the past two decades, extensive literature supports the sphingomyelin/ceramide signaling pathway as an evolutionarily conserved response system for stress (UV, heat, mechanical stress, etc.) (Grassme et al. 2007; Hannun and Obeid 2008). This system is usually in the “off” state under baseline conditions, activated upon contacting stress, and calibrates extent of cellular stress, evoking an adaptive or apoptotic cellular response depending on strength of signal. The most widely studied ceramide-generating mechanism involves translocation of ASMase to the outer plasma membrane, where it hydrolyses its substrate sphingomyelin, generating ceramide therein (Grassme et al. 2001a). This event usually occurs within seconds to minutes of encountering stress, generating ceramide rapidly at the plasma membrane (Rotolo et al. 2012) (Fig. 1). While the mechanism by which diverse stresses are registered at the plasma membrane is uncertain, stress-induced translocation of ASMase to the exoplasmic leaflet of the plasma membrane to initiate stress signaling appears to involve fusion of a subset of S-ASMase-containing acidic vesicles (Kolesnick, Paris, and Rao, unpublished) that abut the inner plasma membrane and upon contacting stress fuse with the bilayer to expose ASMase on the surface (Grassme et al. 2001a). Ceramide, once generated, has a unique biophysical property, that of self-association, forming ceramide-rich platforms (CRPs) on the cell surface, 1–5 μm in diameter, which serve as sites for protein oligomerization and transmembrane signal transduction (Grassme et al. 2001a, 2003a; Rotolo et al. 2005). These CRPs are sites of protein–protein interactions that lead to downstream signaling, and perturbation of CRP formation has been implicated in pathogenesis of a variety of human diseases (Smith and Schuchman 2008).

Ionizing radiation triggers ceramide-rich platform (CRP) formation in BAEC. Clustering of ceramide and ASMase into macrodomains on the outer leaflet of the plasma membrane of BAEC at 1 min post 15 Gy. Cells were fixed 1 min post-irradiation and platforms identified by standard fluorescent microscopy after staining with anti-ceramide IgM MID 15B4 (1:50 dilution, Alexis Biochemicals) or rabbit anti-ASMase Ab 1598 (1:100 dilution), followed by Cy3-conjugated anti-mouse or anti-rabbit IgM (1:500 dilution, Roche Molecular Biochemicals), respectively [Adapted from Rotolo et al. (2012)]
Evidence indicates that ceramide also acts as a classic second messenger in proliferative and stress responses, stoichiometrically activating protein targets (Kolesnick 2002; Hannun and Obeid 2002). Direct targets for ceramide include protein kinase C (PKC) δ, ε and ζ (Kashiwagi et al. 2002; Bourbon et al. 2000) kinase suppressor of Ras (KSR1) (Grassme et al. 2001b), c-Raf-1 (Yu et al. 2000), protein phosphatase 1 and 2a (Chalfant et al. 1999), phospholipase A2 (Huwiler et al. 2001), and cathepsin D (Heinrich et al. 1999). Ceramide binds the C1B lipid-binding domain of KSR1 and c-Raf-1 and via a C1B domain induces PKCε translocation to an intracellular site distinct from the site phorbol esters regulate. A KSR1 C1B-domain polypeptide has been used as a tool to detect surface ceramide on JY B lymphocytes (Grassme et al. 2001b) or as a pharmacologic reagent to inhibit CD95 clustering, hepatocyte apoptosis, and death of mice after intravenous anti-CD95 antibody. This diversity of targets for ceramide interaction is consistent with its pleiotropic involvement in cellular and tissue responses.
1.4 Generation of ASMase Knockout Mice
An inherited deficiency of ASMase activity had been identified as the cause of the lysosomal storage disorder Niemann–Pick disease (NPD) in 1966 by the pioneering work of Brady and colleagues (Brady et al. 1966). To better understand the importance of ASMase in pathogenesis of this disease, two ASMase knockout mouse models were independently developed by Ed Schuchman and Wilhelm Stoffel in 1995 (Otterbach and Stoffel 1995; Horinouchi et al. 1995). These ASMase knockout mice display a phenotype essentially identical to type A NPD (type A NPD is a severe neurodegenerative disease of infancy usually fatal by ages 2–3; see below), including failure to thrive, neurodegeneration, and a shortened life span (Horinouchi et al. 1995). ASMase knockout mice exhibit progressive lipid storage in the RES of liver, lung, and bone marrow and in macrophages of lung, as well as in brain (Otterbach and Stoffel 1995; Horinouchi et al. 1995). The Schuchman ASMase knockout mice are normal at birth and develop routinely until about 12 weeks of age when ataxia and mild tremors became noticeable. At time of death, usually between 6 and 8 months of age, ASMase knockout mice are less than half the weight of wild-type littermates and display an obvious hunched appearance (Fig. 2) (Horinouchi et al. 1995). While the principal lipid accumulating is sphingomyelin, cholesterol and ganglioside storage have also been found. Total blood cholesterol levels are elevated nearly 80 % in ASMase knockout mice compared with wild-type mice. Furthermore, remarkable loss of Purkinje cells in the cerebellum leads to severe impairment of neuromotor coordination. Pulmonary inflammatory disease has also been reported in ASMase knockout mice (Dhami et al. 2001). It should, however, be pointed out that the phenotype of Schuchman’s ASMase knockout mouse line is different from that of Stoffel’s, despite the fact that the technology used to generate these mice was similar. The life expectancy of the Stoffel ASMase-deficient mice was around 4 months, with mice succumbing to advanced NPD (Otterbach and Stoffel 1995). In contrast, mice from the Schuchman group survive to 8 months of age and display a delay in accumulation of sphingomyelin until 12 weeks or so and delay in significant NPD symptomatology until 16 weeks (Horinouchi et al. 1995). The reason for the differences is not clear. It should be emphasized that all studies demonstrating abnormalities in the apoptotic response to various stress should be carried out in ASMase knockout mice before biochemical, histologic, or clinical manifestations of NPD are apparent.

Phenotype of the ASMase knockout mouse. The dramatic size difference and “hunched” appearance of ASMase-deficient mice (top) compared to a control littermate (bottom) at 4 months of age [Adapted from Horinouchi et al. (1995)]
Availability of ASMase knockout mice and development of pharmacologic modulators of SMase function have provided new insight into involvement of the sphingomyelin/ceramide transducer in animal models of human disease. Tissues possess specific cells that represent primary sensors of environmental stress. These sensor cells operate distinct intracellular pathways that detect and calibrate magnitude of stress by converting stress into biochemical signals (Ch’ang et al. 2005). The sphingomyelin/ceramide pathway represents a sensor cell-transduction system operative in select tissues. For instance, IR induces apoptosis of thymocytes via p53 leading to thymus involution, whereas endothelium in the irradiated GI tract uses ASMase, not p53, to initiate apoptosis, conferring GI damage (Paris et al. 2001). Further, while the sensor cell type that utilizes the sphingomyelin/ceramide transducer varies between organs, for instance, germ cells in ovaries and hepatocytes in liver, microvascular endothelium appears as the most common primary sensor in the ASMase knockout mouse model. Endothelial cells are 20-fold enriched in S-ASMase relative to other mammalian cells (Marathe et al. 1998) and are particularly sensitive to stress-induced apoptosis in vitro and in vivo (Kolesnick and Fuks 2003). Endothelium in lung and throughout the central nervous system (CNS) of ASMase knockout mice is almost completely resistant to apoptosis induced by irradiation (Santana et al. 1996; Pena et al. 2000). Furthermore, ASMase knockout mice display defects in hepatocyte apoptosis, liver failure, and animal lethality upon intravenous injection of anti-CD95 antibodies (Lin et al. 2000). ASMase knockout mice manifest a marked defect in the ovarian developmental program (Perez et al. 1997). A failure to normally delete oocytes in ASMase knockout females during embryogenesis leads to ovarian hyperplasia at birth (Morita et al. 2000). Furthermore, the primary cellular sensor may vary between stresses even within the same organ (Ch’ang et al. 2005). ASMase knockout mice provide an invaluable tool to evaluate defective apoptotic signaling and explore the role of the sphingomyelin/ceramide signaling pathway in pathophysiology of various human diseases. The following section defines how this pathway is being actively explored in a number of experimental models of human disease using ASMase knockout mice.
2 ASMase Knockout Mice and Human Disease
2.1 ASMase Knockout Mice for Cancer Radiation Therapy
As stress is known to increase ceramide levels in select mammalian cells, it is not surprising that increased ceramide content has been observed in response to a number of cancer treatments. Interestingly, many tumors seem to have developed strategies to reduce baseline ceramide levels, associated with resistance to chemotherapy or radiotherapy. Several studies showed that ceramide levels were dramatically decreased in some types of human cancer, including colon cancers (Selzner et al. 2001), gliomas (Riboni et al. 2002), and ovarian cancers (Rylova et al. 1998). There is growing evidence that sphingomyelin/ceramide signaling pathway is activated during various cancer treatments, especially during high single-dose radiotherapy (SDRT).
While the classical theory on the cellular effect of IR identifies DNA as the major target for initiating death pathways, a growing body of data now demonstrates that ASMase-mediated ceramide generation regulates apoptosis in response to IR. In 1994, Haimovitz-Friedman et al. reported that irradiation of bovine aortic endothelial cells (BAEC) induced sphingomyelin conversion to ceramide and apoptosis, suggesting that radiation damage to plasma membrane might initiate apoptotic signaling through ceramide. Activation of ASMase was also observed in Rat-1 Myc-ER cells and primary skin fibroblasts (Zundel and Giaccia 1998; Bohler et al. 1997). Definitive evidence for a role of ASMase in radiation-induced apoptosis was derived from studies with ASMase-deficient models. B lymphoblastoid cells immortalized from an NPD type A patient fail to hydrolyze sphingomyelin in response to radiation, generate ceramide, or undergo apoptosis (Santana et al. 1996). Retroviral transfer of normal ASMase cDNA into these cells restored ceramide generation, leading to restoration of apoptotic cell death upon radiation treatment. Mouse embryonic fibroblasts (MEFs) from ASMase knockout mice also failed to generate ceramide and were completely resistant to radiation-induced apoptosis, yet remained sensitive to staurosporine-induced apoptosis, which is not mediated via ceramide signaling (Lozano et al. 2001). Additional studies showed that sensitivity to radiation-induced apoptosis could be restored in MEFs by exogenous addition of nanomolar concentrations of natural ceramide. Rescue of the radiation apoptotic phenotype by adding ceramide provides strong evidence that ceramide is obligate for this form of apoptosis induced by radiation.
IR, delivered as a small daily fraction of 1.8–3.0 Gy, cures approximately 60 % of localized, nonmetastatic cancer. Fractioned radiotherapy has been favored because normal tissue stem cell clonogens repair DNA double-strand breaks (DSBs) more proficiently than their tumor counterparts during interfraction intervals, enabling dose escalation to tumor cure levels with acceptable normal tissue damage. However, in many instances, curative tumor doses cannot be reached due to high radiosensitivity of adjacent normal organs. The recent emergence of image-guided, high-precision targeting of human tumors that effectively excludes normal tissues from the treatment field may obviate this issue, allowing for high SDRT (20–24 Gy), a modality whose success has exceeded all expectations (Song et al. 2004).
The mechanism of tumor response after SDRT may differ from that of conventional fractioned radiotherapy (Moeller et al. 2005; Fuks and Kolesnick 2005). The central role of ASMase-mediated apoptosis in endothelial cells for optimal SDRT response was detailed in two tumor models (melanoma and fibrosarcoma). Studies by Garcia-Barros et al. (2003) indicated that exposure to single radiation doses >8 Gy engages an apoptotic response in tumor microvessels by activating the sphingomyelin/ceramide transducer in endothelium (Fig. 3). MCA129 fibrosarcomas and B16 melanomas transplanted into ASMase or BAX knockout mice, which provide tumors with host-derived apoptosis-resistant microvascular networks, were completely resistant to potentially curative doses of 15–20 Gy SDRT. Hence, high-dose radiation exposure appears to induce primarily sublethal lesions in tumor stem cell clonogens that at these doses are converted into lethal damage upon induction of apoptotic microvascular dysfunction. Discovery of the critical role of the sphingomyelin/ceramide transducer in curative SDRT defines endothelium as a valid pharmacologic target, consistent with early data suggesting that antagonism of VEGF or the VEGFR2 receptor radiosensitizes tumors by derepressing endothelial ASMase (Truman et al. 2010).

MCA/129 fibrosarcomas implanted into ASMase −/− mice display reduced radiation-induced endothelial cell apoptosis. MCA/129 fibrosarcomas, grown to 150–200 mm3, were irradiated with 15 Gy. Tumor specimens were obtained 4 h post-irradiation, fixed in 4 % fresh formaldehyde, embedded in paraffin, and 5-μm sections were evaluated for apoptosis by terminal deoxytransferase-mediated deoxyuridine triphosphate nick end labeling (TUNEL). Thereafter, endothelial cells were identified using an antibody specific for the endothelial cell surface marker CD-34. Apoptotic endothelial cells, identified as a red-brown TUNEL-positive nuclear signal surrounded by dark-blue plasma membrane signal indicative of CD-34 staining, are lacking in sections of tumor grown in ASMase −/− mice. Similar results were obtained using B16F1 melanomas [Adapted from Garcia-Barros et al. (2003)]
2.2 ASMase Knockout Mice for Study of GI Damage
In addition to the potential importance of ASMase in cancer therapy, researchers have been investigating impact of ASMase on normal tissue damage. The radiation GI syndrome is a major toxicity that may occur in a Fukushima-like nuclear accident or upon deliberate radiation exposure. The GI syndrome involves destruction of crypt/villus units, loss of mucosal integrity, and infection by resident enterobacterial flora, characterized clinically as anorexia, vomiting, diarrhea, dehydration, systemic infection, and in extreme cases, septic shock and death. Studies of Paris et al. first reported that microvascular endothelium serves as a primary target for radiation in induction of tissue damage (Paris et al. 2001). Vascular dysfunction in the GI mucosa occurs following a rapid wave of radiation-induced endothelial apoptosis (2–6 h post-IR) regulated by ASMase-mediated ceramide generation within the villus/crypt microvascular network. Genetic inactivation of ASMase inhibits this ceramide-driven mode of endothelial apoptosis, preventing IR-induced animal lethality (Paris et al. 2001). Cho et al. (2004) supported this hypothesis, reporting intravenous injection of the angiopoietin-1 variant COMP-Ang1, which specifically targeted Tie-2 receptors of intestinal microvascular endothelial but not epithelial cells, attenuated radiation-induced endothelial apoptosis, and protected against GI syndrome lethality.
Graft-versus-host disease (GVHD) represents a different process in which ASMase-mediated apoptosis plays a role in morbidity and mortality. GVHD is a frequent complication in leukemia patients receiving allogeneic bone marrow transplantation. In GVHD, donor cytotoxic T lymphocytes (CTLs) attack host tissues leading to organ damage. Recently, Rotolo et al. (2009) showed that CTLs generate CRPs on critical cells within GVHD-responsive target organs (small intestines, skin, liver), signaling apoptosis to initiate GVHD. Evidence indicated cytokines of the TNF superfamily as critical to CTL-induced pathophysiology. Using clinically relevant mouse models of acute GVHD in which allogeneic bone marrow and T cells were transplanted into ASMase+/+ and ASMase−/− mice, host ASMase was identified as critical for full-blown GVHD. Lack of host ASMase reduced apoptosis of relevant GVH target cells, including hepatocytes and intestinal and skin cells, as target cells failed to form CRPs upon contacting activated CTLs and consequently resisted CTL-mediated cell death. Hence, the sphingomyelinase/ceramide signaling pathway may provide targets for pharmacologic modulation of GI and perhaps other damage in GVHD, the major dose-limiting toxicity in leukemia patients receiving allogeneic marrow transplantation (Waselenko et al. 2004).
2.3 ASMase Knockout Mice for Study of Ovarian Damage
The ovary represents another organ whose chemosensitivity/radiosensitivity is regulated by ASMase. Almost 90 % of oocytes die by apoptosis prior to birth, a process that continues throughout adulthood resulting in menopause. Tilly and coworkers showed that both fetal and postpartum oocyte deletion in mice require sphingomyelin/ceramide signaling, with ASMase knockout mice displaying 1.6-fold increased oocytes at birth, maintained throughout adulthood (Morita et al. 2000; Casper and Jurisicova 2000). This group defined a mechanism for ceramide-mediated deletion in aging oocytes (Perez et al. 2005). Young wild-type oocytes isolated with their granulosa cell layer display elevated ceramide in both cell populations and were resistant to spontaneous or ceramide-induced apoptosis. In contrast, aged oocytes manifested high granulosa cell ceramide but low oocyte ceramide. Over 24 h in culture, ceramide was transferred from granulosa cells to oocytes by a process requiring gap junctions and intact rafts, and apoptosis, requiring Bax downstream, ensued. S1P treatment did not prevent ceramide transfer but prevented apoptosis. Similarly, genetic ASMase deletion or binding intracellular ceramide by injecting anti-ceramide antibody lowered available ceramide in young oocytes rendering them susceptible to exogenously added ceramide. These studies indicate that the sphingomyelin/ceramide transducer regulates normal ovarian physiology.
The sphingomyelin/ceramide transducer also mediates lethal effects of drugs and IR on oocytes. Morita et al. showed that apoptosis of isolated murine oocytes ex vivo to 200 nM doxorubicin was prevented by S1P pretreatment. Ceramide generation in response to doxorubicin appeared to occur through sphingomyelin hydrolysis rather than de novo ceramide synthesis as there was a robust apoptotic response in ASMase wild-type oocytes cultured with doxorubicin for 24 h, whereas ASMase knockout oocytes were almost completely resistant to doxorubicin destruction. Further, S1P injection into the bursa surrounding the mouse ovary dose-dependently prevented IR-induced oocyte apoptosis and sterility, preserving oocyte function, enabling successful in vitro and in vivo fertilization. In contrast to unprotected mice, irradiated S1P-protected mice maintained normal fertility for over 1 year, and their F1 and F2 offspring did not display behavioral, hematologic, histologic, biochemical, or genetic abnormalities (Paris et al. 2002). Moreover, Tilly and coworkers recently reported a successful monkey trial to evaluate the effect of S1P and its long-acting mimetic FTY720 on ovarian function and fertility after irradiation. In these studies, S1P and FTY720 protected ovaries of adult female rhesus monkeys from damage caused by 15 Gy targeted radiotherapy, allowing for long-term fertility (Zelinski et al. 2011). These studies indicate that chemotherapy-induced stress and IR-induced stress engage the sphingomyelin/ceramide physiologic mechanism for oocyte deletion, accelerating organ damage. Further, these studies suggest a small molecule approach to this intractable side effect of cancer therapy.
2.4 ASMase Knockout Mice for Study of Brain Diseases
2.4.1 Niemann–Pick Disease
Types A and B NPD result from inherited ASMase deficiency. The first type A NPD patient was described by Albert Niemann in 1914. Type A NPD is a severe neurodegenerative disease of infancy usually fatal by ages 2–3. In contrast, type B NPD patients have minimal or no neurologic involvement and often survive into adulthood but may have severe and progressive visceral organ abnormalities, including hepatosplenomegaly, and pulmonary and cardiovascular disease (Schuchman 2010). Differences between types appear due to the level of residual ASMase activity. ASMase knockout mice show a similar phenotype to type A NPD patients, owing to complete absence of ASMase activity. It has been reported that calcium homeostasis is altered in the cerebellum of ASMase knockout mice, suggesting that calcium dysfunction may cause Purkinje cell degeneration (Ginzburg and Futerman 2005).
Several groups have used these mice for testing of novel therapeutics for NPD, including recombinant enzyme replacement therapy (ERT), gene therapy, and stem cell transplantation. In 1999, large-scale purification of recombinant human ASMase was achieved in Chinese hamster ovary cells (He et al. 1999), allowing for human ERT. Initial studies that evaluated the effect of ERT in ASMase knockout mice were performed by Miranda and coworkers, who demonstrated that the sphingomyelin levels were significantly reduced in RES organs when recombinant ASMase (rASMase) was administered intravenously into young mice (Miranda et al. 2000). However, there was no impact on progression of neurologic disease, and mouse life span was not extended due to failure of rASMase to pass the blood–brain barrier into the CNS (Sly and Vogler 2002). Recent studies showed that intraparenchymal injection of rASMase results in regional reduction in sphingomyelin and cholesterol levels, but re-accumulation is observed at 2 weeks post-injection (Yang et al. 2007). Follow-up studies from the same lab demonstrated that intracerebroventricular infusion of rASMase led to widespread ASMase distribution and significant reduction in lysosomal sphingomyelin accumulation (Dodge et al. 2009).
The effect of stem cell transplantation on progression of neurologic disease also has been extensively evaluated in ASMase knockout mice. Intravenous transplantation of ASMase-expressing bone marrow cells into ASMase knockout mice showed positive effects on RES organs, but effects on neurologic disease were modest (Miranda et al. 1998). Further studies showed that intracerebral transplantation of ASMase-expressing mesenchymal stem cells into ASMase knockout mice delayed onset of neurologic abnormalities and extended life span, but progressive neurologic disease was not prevented (Jin et al. 2002; Jin and Schuchman 2003). Overall, these findings suggest stem cell transplantation as therapeutically promising.
In addition to the therapeutic approaches mentioned above, gene therapy has been intensively investigated during the last decade. Adeno-associated viral (AAV) vectors have been widely used. Several studies demonstrated that intracranial injections of AAV encoding human ASMase are effective in decreasing the burden of sphingolipid storage in the brains of ASMase knockout mice. ASMase activity was found not only within deep cerebellar nuclei but also throughout the CNS. Moreover, progressive ataxia was prevented and life span normalized (Dodge et al. 2005; Passini et al. 2005, 2007). These data suggest AAV-based therapy as a promising therapeutic modality for treating NPD.
2.4.2 Stroke
Stroke, the third leading cause of death in the industrialized world, in the large majority of cases results from occlusion of arterial blood flow into the brain. While pathogenesis of the ischemic lesion is complex, excess excitotoxicity, peri-infarct depolarization, inflammation, and apoptosis are factors contributing to evolution of tissue damage. These events do not impact the ischemic region homogenously. Rather, a perfusion-occluded anoxic core, manifesting necrosis, is surrounded by a rim of restricted blood flow, termed the penumbra. Although penumbra ischemic cells, and especially neurons, are at high risk for apoptotic death, this region is potentially salvageable. Hence, a recent emphasis of pharmacologic intervention in stroke is in apoptosis.
Nakane and coworkers reported in gerbils (Nakane et al. 2000) and rats (Kubota et al. 2000) that lethal forebrain ischemia (5 min) induced by bilateral carotid occlusion induced rapid sphingomyelin hydrolysis to ceramide (30 min), preceding neuronal apoptotic death. Debatin and colleagues (Herr et al. 1999) subsequently showed that rat middle cerebral artery (MCA) occlusion leads to twofold increased ceramide levels compared to the ipsilateral non-occluded brain hemisphere at 6 h after reperfusion. ASMase-generated ceramide purportedly initiated neuronal apoptosis in this model by upregulating death-inducing ligands (Fas ligand, TRAIL, and TNF-α), an effect prevented by the neuroprotector FK506 (Herr et al. 1999). Genetic evidence supporting this notion was provided by Mattson and coworkers (Yu et al. 2000) reporting that ASMase knockout mice neither generated ceramide nor upregulated inflammatory cytokines upon MCA occlusion and displayed markedly reduced neuronal apoptosis in the penumbra. Infarct size was reduced 30 %, and neurologic (primarily motor) deficiencies improved dramatically. Primary cultures of ASMase−/− cortical neurons displayed markedly reduced excitotoxicity upon glutamate treatment and chemical hypoxia induced by cyanide, accompanied by protection from intracellular Ca2+ elevation, and generation of reactive oxygen species (note: ischemic and excitotoxic neuronal death is considered mediated by calcium overload and oxyradical production). These studies indicate cell autonomous utilization of the sphingomyelin/ceramide transducer by cortical neurons to discriminate death signals.
2.4.3 Blood–Brain Barrier Dysfunction
The blood–brain barrier (BBB) is a highly specialized microvascular network characterized by tight cell–cell junctions lacking fenestrations, which restricts transcapillary flux of water-soluble compounds into the CNS (Neuwelt 2004). The dysfunctions of BBB is observed during CNS bacterial and viral infections, inflammatory and degenerative CNS disorders, cerebrovascular disease, trauma, primary and metastatic brain tumors, and after IR. BBB disruption often results in vasogenic edema that contributes significantly to disease-associated symptoms. Concomitantly, however, it facilitates transport of cytokines, antibodies, inflammatory and immune cells, as well as drugs, to disease-stricken areas.
Recent studies demonstrate a role for sphingomyelin/ceramide transduction in IR-induced BBB dysfunction. Pena et al. (2000) showed that irradiated murine CNS endothelium undergoes dose-dependent apoptosis at 5–100 Gy SDRT, peaking after 12 h. After 50 Gy, which induces subacute and chronic damage, approximately 20 % of endothelium underwent apoptosis at 12 h, inhibitable by genetic ASMase inactivation or pretreatment with the endothelial survival factor bFGF. Li et al. (2003) showed that this level of endothelial apoptosis resulted in a 40–60 % reduction in endothelial cell density 24 h after 50 Gy, an event abrogated in ASMase knockout mice, but not in p53 knockout mice. Further, BBB dysfunction, assessed by leakage of albumin or Evans blue dye into the CNS, was abrogated in ASMase−/− mice. These observations suggest that pharmacologic activation of the sphingomyelin/ceramide transducer might be used therapeutically to produce transient BBB disruption, designed to facilitate drug delivery to disease sanctuary regions within the CNS (van Vulpen et al. 2002).
2.5 ASMase Knockout Mice for Study of Lung Diseases
ASMase deficiency in both type B NPD patients and in ASMase knockout mice alters pulmonary cellular and organ structure secondary to accumulation of sphingomyelin, leading to lung abnormalities and decreased pulmonary function. Type B NPD is linked to progressive pulmonary function decline and frequent respiratory infection. X-ray and CT examination in type B NPD patients revealed that over 90 % displayed evidence of interstitial lung disease (Mendelson et al. 2006). ASMase knockout mice begin at 10 weeks of age to have a significantly higher number of cells in their pulmonary airspaces than normal mice, consisting primarily of enlarged and often multinucleated macrophages. In mice and humans, the alveolar macrophage serves as the first line of host defense to clear extracellular bacteria from the lung, implying an important role of ASMase in lung host defense against pathogens. Further, mechanical research indicates that ASMase is required for normal surfactant catabolism by alveolar macrophages. In this regard, ASMase knockout mice exhibit elevated levels of total surfactant lipid and protein (Buccoliero et al. 2004) and alterations in surfactant composition, including increased sphingomyelin content, that appear to contribute to abnormal surfactant function observed in ASMase knockout mice (Buccoliero et al. 2004; Tuder et al. 2003). In principle, these studies suggest that ASMase might regulate fundamental aspects of lung disease pathologies.
2.5.1 Acute Respiratory Distress Syndrome
Acute respiratory distress syndrome (ARDS) is defined by severe noncardiac respiratory distress, impaired arterial oxygenation (hypoxemia), and bilateral pulmonary infiltrates. It has diverse origins, including sepsis, pneumonia, aspiration, trauma, smoke or toxic gas inhalation, and some drugs (Piantadosi and Schwartz 2004). Pathogenesis involves primary endothelial dysfunction, leakage of fluid across the alveolar-capillary barrier, and fluid accumulation in the alveolar space (edema), causing refractory hypoxemia. Cytokines and chemokines may be crucial in endothelial cell dysfunction. Recent attention has focused on platelet-activating factor (PAF) that mediates acute lung injury (ALI) that progresses to ARDS in part through prostaglandin E2 production.
Goggel et al. (2004) demonstrated a role for ceramide in PAF-induced pulmonary edema in mouse and rat models (Barnes 2004). PAF injection into mice or perfusion of isolated intact rat lungs with PAF resulted in rapid serum ASMase and pulmonary ceramide elevation. PAF-induced pulmonary edema was reduced in ASMase knockout mice. Rat pulmonary edema was mimicked by perfusion of short-chain C2-ceramide, but not C2-dihydroceramide, which differs only in the trans double bond at position 4–5. Further, injection of anti-ceramide antibodies antagonized mouse pulmonary edema by about 50 %. Other less specific ASMase inhibitors (desipramine and D609) acted similarly and, when combined with a cyclooxygenase inhibitor to prevent prostaglandin E2 production, abolished edema. These studies suggest that concomitant inhibition of two intracellular lipid signaling pathways might serve as a new approach to ARDS therapy (Barnes 2004).
2.5.2 Pulmonary Fibrosis
Pulmonary fibrosis is another common form of interstitial lung disease associated with inflammation and apoptosis. Recently, the Schuchman group found that intratracheal instillation of bleomycin which can cause pulmonary fibrosis in normal mice leads to increase in lung ASMase activity, while bleomycin instillation in ASMase knockout mice does not cause fibrosis. This suggests that inhibition of ASMase transiently might be a useful way to prevent toxicity of some cancer drugs (Dhami et al. 2010).
Since ASMase plays an important role in surfactant clearance in lung where pathogens interact closely with target cell membranes, there has been an expanding literature regarding ASMase regulation of pathogen infection using ASMase knockout mice. Cystic fibrosis (CF) is a genetic disorder caused by the mutation of cystic fibrosis transmembrane conductance regulator (cftr) gene. Growing evidence shows that sphingolipids, in particular ceramide, play an important role in CF and the bacterial infections associated with this disease (Teichgraber et al. 2008; Becker et al. 2010).
According to the CF registry, chronic lung infection with Pseudomonas aeruginosa contributes to death in over three-quarter of patients (Pier 2002). Gulbins and coworkers defined a pathophysiology for Pseudomonas pulmonary infection involving ASMase/ceramide dysregulation in CF. This laboratory discovered an age-dependent ceramide accumulation in the respiratory tracts of cftr-deficient mice that might be caused by an imbalance between ASMase cleavage of sphingomyelin to ceramide and acid ceramidase consumption of ceramide, resulting in higher ceramide levels (Teichgraber et al. 2008). Ceramide accumulation found in the lungs of CF mice enhanced age-dependent pulmonary inflammation, death of respiratory epithelial cells, and resulted in high susceptibility to severe P. aeruginosa infections, events corrected in ASMase+/− mice or by amitriptyline inactivation of ASMase. Based on these results, a successful human clinical trial was initiated by these investigators, the outcome of which suggests that amitriptyline treatment may improve lung function in CF patients (Riethmuller et al. 2009).
2.6 ASMase Knockout Mice for Study of Heart Disease and Diabetes
Several reports suggest that lipid abnormalities are part of the phenotype of patients with type A and B NPD, associated with early atherosclerotic heart disease. Moreover, reduced high-density lipoprotein (HDL) cholesterol, hypertriglyceridemia, and elevated low-density lipoprotein (LDL) cholesterol were found in type A and B patients, coinciding with presence of early atherosclerotic plaques (McGovern et al. 2004). Subendothelial retention of atherogenic lipoprotein, especially LDL, is a critical early event in atherogenesis. Tabas and coworkers demonstrated that S-ASMase hydrolyzes sphingomyelin present in LDL at physiologic pH, leading to formation of aggregated LDL in the subendothelial space, stimulating macrophage foam cell formation (Schissel et al. 1998b) and accelerating plaque formation. Recently, studies from the same group showed 50 % decrease in early foam cell aortic root lesional area and 87 % reduction in lipoprotein trapping in ASMase-deficient Apoe−/− mice compared with ASMase wild-type Apoe−/− mice (Devlin et al. 2008). Elevated S-ASMase activity was also observed in chronic heart failure (Doehner et al. 2007). Moreover, macrophages from ASMase knockout mice have defective cholesterol trafficking and efflux (Leventhal et al. 2001). All these data suggest that ASMase might play an important role in atherosclerosis development and serve as target for therapeutics.
In addition to its role in heart disease, ASMase has been proposed as linked with diabetes. ASMase knockout mice on a hypercholesterolemic genetic background show resistance to diet-induced hepatic triacylglycerol accumulation and hyperglycemia (Deevska et al. 2009). Interestingly, deletion of ASMase resolved diet-induced hepatic steatosis and improved insulin sensitivity. The improvement in steatosis and insulin sensitivity was associated with elevation in sphingolipids (including dihydroceramides, ceramides, sphingomyelin, and sphingosine) and marked decrease in triacylglycerol accumulation (Deevska et al. 2009). These data imply that ASMase might play a significant role in progression of diabetes.
2.7 ASMase Knockout Mice for Study of Liver Diseases
A number of distinct disease models provide strong evidence for a potential role of ASMase-mediated apoptosis in progression of liver disorders. The first model is that of T cell-mediated auto-aggressive liver disease. In 2000, Kirschnek et al. reported that intracenous injections of phytohemagglutinin (PHA) results in a Fas-dependent autoimmune hepatitis (Kirschnek et al. 2000). Further studies revealed that this syndrome is caused by induction of Fas ligand on lymphocytes upon PHA stimulation and migration of these lymphocytes to liver. Hepatocytes are killed by apoptosis upon contact with the Fas ligand-positive T cells, leading to autoimmune hepatitis. Deletion of ASMase in mice protected hepatocytes from T cell-induced apoptosis (Kirschnek et al. 2000) and protected mice against autoimmune-like hepatitis. These results provide in vivo evidence for an important physiologic function of ASMase in PHA-induced hepatitis.
A second hepatic disease model is that of Wilson’s disease. Inactivating mutations of ATP7B, a key enzyme of liver copper secretion, causes inappropriate accumulation of Cu2+ in liver parenchyma, resulting in cirrhosis and hemolytic anemia. In this disease model, Cu2+-triggered hepatocyte apoptosis is mediated by ASMase activation and ceramide release. Moreover, elevated ASMase plasma activity was observed in Wilson’s disease patients. Genetic deficiency or pharmacologic inhibition of ASMase by amitriptyline prevented Cu2+-induced hepatocyte apoptosis and protected rats from acute hepatocyte death, liver failure, and early death (Lang et al. 2007). Cu2+-induced ASMase secretion from leukocytes leads to ceramide generation in erythrocytes and subsequent phosphatidylserine exposure at the cell surface. The functional consequence of this series of events is that affected erythrocytes are deleted immediately from blood by macrophages in mice resulting in anemia, whereas ASMase-deficient erythrocytes were resistant to Cu2+ treatment. These data indicate the significance of ASMase and ceramide for pathogenesis of Wilson’s disease and provide a basis for treatment by pharmacologic inhibition of ASMase.
In addition to these two disease models, studies showed that hepatocytes from ASMase knockout mice are resistant to apoptosis mediated by TNF-α (Garcia-Ruiz et al. 2003). A consensus has developed that acute and chronic liver injury are largely mediated by hepatocyte apoptosis, with TNF-superfamily members, particularly Fas/CD95, TNFR, and DR5 playing critical roles (Guicciardi and Gores 2005). Biochemical and histochemical elevation of cognate cytokine ligands (Fas ligand, TNF-α, and TRAIL) correlates with extent of disease and outcome, and their pharmacologic and genetic inhibition attenuate liver damage. Furthermore, TNF-α administration to mice recapitulates acute hepatic injury syndromes (Ding and Yin 2004). Fernandez-Checa and colleagues showed that wild-type hepatocytes respond to TNF-α with rapid ASMase activation, ceramide generation, and massive apoptosis (Garcia-Ruiz et al. 2003), while ASMase-deficient hepatocytes were TNF-resistant. Consistent with these observations, ASMase knockout mice were resistant to TNF-α-induced hepatitis and death from liver failure. Similarly, ASMase knockout mice were resistant to anti-Fas-induced ceramide elevation and hepatic injury (Lin et al. 2000), and attenuation of ceramide elevation via adenoviral overexpression of neutral ceramidase prevented TNF-induced hepatitis (Osawa et al. 2005). Furthermore, knockout of the mouse adaptor protein FAN (Malagarie-Cazenave et al. 2004), which binds TNFR1 to transduce NSMase activation, displayed reduced IL-6 production and TNF-induced hepatic injury and improved survival. Evidence suggests that ASMase-generated ceramide may act by inducing downregulation of methionine adenosyltransferase 1A (MAT1A) (Mari et al. 2004), an enzyme required for maintaining levels of the one-carbon donor S-adenosyl-methionine (SAM). MAT1A impairment lowers SAM, contributing to injury in experimental models of liver damage and in patients with alcohol- and viral-mediated cirrhosis. Intraperitoneal SAM injection into wild-type mice mimicked the ASMase−/− phenotype, preventing TNF- and lipopolysaccharide-induced hepatitis, improving survival. Altogether, these studies indicate that elevating ceramide enhances liver damage, while preventing elevation delivers protection, legitimizing this target for pharmacologic intervention in hepatic injury.
2.8 ASMase Knockout Mice for Study of Infection
Several studies support ASMase as having a central role in infection of mammalian cells with diverse microbial pathogens. CRPs are co-opted by a wide range of pathogens including bacteria, viruses, and parasites to infect mammalian cells. Initial studies with N. gonorrhoeae showed ASMase activation and release of ceramide upon infection of human epithelial cells and macrophages. Moreover, pharmacologic inhibition or genetic knockout of ASMase prevented infection of human epithelial cells with N. gonorrhoeae (Grassme et al. 1997; Hauck et al. 2000). These data suggested that ASMase is required for internalization of N. gonorrhoeae, confirmed in subsequent studies (Grassme et al. 2003b). P. aeruginosa also activates ASMase and triggers ceramide release within minutes of contacting target cells. Pseudomonas-induced CRP formation on the surface of the target cell appears required for acute infection (Fig. 4), as ASMase knockout prevented CRP formation, internalization of P. aeruginosa by lung epithelial cells, and subsequent epithelial cell apoptosis (Grassme et al. 2003b). Evidence also indicates ASMase involvement in infection by Staphylococcus aureus, Salmonella typhimurium, Escherichia coli, and Mycobacterium (for further details please see Grassmé and Becker 2013).

P. aeruginosa triggers platform formation by ASMase-mediated release of ceramide. Infection (20 min) of murine tracheal epithelial cells with P. aeruginosa strain ATCC 27853 induced formation of CRPs that contain ASMase. Cells were infected for 20 min with P. aeruginosa, fixed in PFA, and isolated and stained with FITC-labeled anti-ASMase and Cy3-coupled anti-ceramide antibodies. The cells were analyzed by fluorescence microscopy [Adapted from Grassme et al. (2003b)]
In addition to bacteria, virus–host cell interaction may require ASMase activity. Griffin and coworkers reported ASMase knockout mice more susceptible to sindbis virus (SV) infection than wild-type or heterozygous (Het) littermates (Ng and Griffin 2006). In SV infection, sphingomyelin is required for virus entry, an event impacted by altered sphingomyelin storage in ASMase-deficient mice. Accumulation of sphingomyelin in brains of ASMase knockout mice allows more rapid entry and spread of SV, leading to higher viral titers in the nervous system, and subsequent increased neuronal death compared with wild-type mice. Several cell-based studies also confirm a fundamental role of ASMase in virus infection. For instance, infection of human epithelial cells with rhinovirus induces ASMase activation, release of ceramide, and CRP formation, events blocked in human fibroblasts genetically deficient for ASMase or by pharmacologic inhibitors of ASMase (Grassme et al. 2005). Inhibition of SMase by N-palmitoyl-dl-dihydrosphingosine similarly decreased yield of infectious herpes virus, suggesting ASMase involvement in herpes infection or replication (Steinhart et al. 1984). Moreover, several studies suggest that the sphingomyelin/ceramide signaling pathway may influence the course of HIV-1 infection. HIV-cell infection involves interaction of HIV receptor molecule gp120 with CD4 on the T cell surface. CD4 ligation initiates T cell depletion by clustering of endogenous Fas/Fas ligand (Wang et al. 1994), an event requiring CRPs. Altogether, these data suggest that manipulation of sphingolipids might be strategic for treatment of virus infection.
In addition to a role of ASMase in virus uptake, a recent study showed that ASMase activity might be required by host immune system to eliminate virus-infected cells. CD8+ CTLs and natural killer cells lyse virus-infected target cells through exocytosis of cytotoxic granules. CTLs from ASMase knockout mice are defective in exocytosis of cytolytic effector molecules, leading to attenuated cytotoxic activity of CTLs and delayed elimination of lymphocytic choriomeningitis virus (Herz et al. 2009). Moreover, macrophages from ASMase knockout mice are defective in killing of the bacterium L. monocytogenes, resulting in unrestricted spread of infection (Utermohlen et al. 2003). Given the complexity of the impact of ASMase deficiency on the course of infection, full molecular characterization of sphingolipid involvement in specific infectious states would appear warranted before clinical pharmacologic manipulation is attempted.
Finally, at least one parasitic infection is influenced by SMase-induced ceramide generation in a unique way. Plasmodium seems to have developed a strategy to circumvent a requirement for mammalian target cell ASMase. In fact, Plasmodium expresses its own SMase for generation of ceramide in trans in target cells, an effect required for erythrocyte infection with P. falciparum or P. berghei. Therefore, only a slightly different course of malarial infection was observed between ASMase wild-type and knockout mice (Brand et al. 2008). Thus, in addition to host ASMase, pathogen SMase might represent a novel target for malaria, defined using the ASMase mouse knockout.
2.9 Perspective
ASMase knockout mice provide an invaluable tool to explore the role of the sphingomyelin/ceramide signaling pathway in pathophysiology of various human diseases (Table 1). Accumulating evidence indicates a crucial involvement of sphingomyelin/ceramide in a number of disease initiation states, as well as disease treatments, making the pathway an attractive target for therapy. In particular, manipulation of CRPs might be strategic for treatment of various human diseases. Further studies, however, are needed to more fully molecularly characterize the enzymes involved in generation and metabolism of ceramide in these disease states to develop a more comprehensive understanding of the signaling network that regulates their individual sphingomyelin/ceramide transduction. Nonetheless, human diseases regulated through sphingomyelin/ceramide signal transduction represent a newly defined class of clinical entities that provides unique targets of opportunity for pharmacologic intervention. This developmental process undoubtedly will continue to rely on use of the original ASMase knockout mice strain, and on newer versions currently in development, in the process of disease discovery and therapeutics.
References
Barnes P (2004) Ceramide lances the lungs. Nat Med 10:130–131
Becker K, Grassme H, Zhang Y, Gulbins E (2010) Ceramide in Pseudomonas aeruginosa infections and cystic fibrosis. Cell Physiol Biochem 26:57–66
Bionda C, Portoukalian J, Schmitt D, Rodriguez-Lafrasse C, Ardail D (2004) Subcellular compartmentalization of ceramide metabolism: MAM (mitochondria-associated membrane) and/or mitochondria? Biochem J 382:527–533
Bohler T, Baumler C, Herr I, Groll A, Kurz M, Debatin K (1997) Activation of the CD95 system increases with disease progression in human immunodeficiency virus type 1-infected children and adolescents. Pediatr Infect Dis J 16:754–759
Bourbon N, Yun J, Kester M (2000) Ceramide directly activates protein kinase C zeta to regulate a stress-activated protein kinase signaling complex. J Biol Chem 275:35617–35623
Brady R, Kanfer J, Mock M, Fredrickson D (1966) The metabolism of sphingomyelin. II. Evidence of an enzymatic deficiency in Niemann-Pick disease. Proc Natl Acad Sci USA 55:366–369
Brand V, Koka S, Lang C, Jendrossek V, Huber S, Gulbins E, Lang F (2008) Influence of amitriptyline on eryptosis, parasitemia and survival of Plasmodium berghei-infected mice. Cell Physiol Biochem 22:405–412
Buccoliero R, Ginzburg L, Futerman A (2004) Elevation of lung surfactant phosphatidylcholine in mouse models of Sandhoff and of Niemann-Pick A disease. J Inherit Metab Dis 27:641–648
Casper R, Jurisicova A (2000) Protecting the female germ line from cancer therapy. Nat Med 6:1100–1101
Ch’ang H, Maj J, Paris F, Xing H, Zhang J, Truman J, Cardon-Cardo C, Haimovitz-Friedman A, Kolesnick R, Fuks Z (2005) ATM regulates target switching to escalating doses of radiation in the intestines. Nat Med 11:484–490
Chalfant C, Kishikawa K, Mumby M, Kamibayashi C, Bielawska A, Hannun Y (1999) Long chain ceramides activate protein phosphatase-1 and protein phosphatase-2A. Activation is stereospecific and regulated by phosphatidic acid. J Biol Chem 274:20313–20317
Cho C, Kammerer R, Lee H, Yasunaga K, Kim K, Choi H, Kim W, Kim S, Park S, Lee G, Koh G (2004) Designed angiopoietin-1 variant, COMP-Ang1, protects against radiation-induced endothelial cell apoptosis. Proc Natl Acad Sci USA 101:5553–5558
Cuvillier O, Pirianov G, Kleuser B, Vanek P, Coso O, Gutkind S, Spiegel S (1996) Suppression of ceramide-mediated programmed cell death by sphingosine-1-phosphate. Nature 381:800–803
Deevska G, Rozenova K, Giltiay N, Chambers M, White J, Boyanovsky B, Wei J, Daugherty A, Smart E, Reid M, Merrill A, Nikolova-Karakashian M (2009) Acid sphingomyelinase deficiency prevents diet-induced hepatic triacylglycerol accumulation and hyperglycemia in mice. J Biol Chem 284:8359–8368
Devlin C, Leventhal A, Kuriakose G, Schuchman E, Williams K, Tabas I (2008) Acid sphingomyelinase promotes lipoprotein retention within early atheromata and accelerates lesion progression. Arterioscler Thromb Vasc Biol 28:1723–1730
Dhami R, He X, Gordon R, Schuchman E (2001) Analysis of the lung pathology and alveolar macrophage function in the acid sphingomyelinase–deficient mouse model of Niemann-Pick disease. Lab Invest 81:987–999
Dhami R, He X, Schuchman E (2010) Acid sphingomyelinase deficiency attenuates bleomycin-induced lung inflammation and fibrosis in mice. Cell Physiol Biochem 26:749–760
Ding W, Yin X (2004) Dissection of the multiple mechanisms of TNF-alpha-induced apoptosis in liver injury. J Cell Mol Med 8:445–454
Dodge J, Clarke J, Song A, Bu J, Yang W, Taksir T, Griffiths D, Zhao M, Schuchman E, Cheng S, O’Riordan C, Shihabuddin L, Passini M, Stewart G (2005) Gene transfer of human acid sphingomyelinase corrects neuropathology and motor deficits in a mouse model of Niemann-Pick type A disease. Proc Natl Acad Sci USA 102:17822–17827
Dodge J, Clarke J, Treleaven C, Taksir T, Griffiths D, Yang W, Fidler J, Passini M, Karey K, Schuchman E, Cheng S, Shihabuddin L (2009) Intracerebroventricular infusion of acid sphingomyelinase corrects CNS manifestations in a mouse model of Niemann-Pick A disease. Exp Neurol 215:349–357
Doehner W, Bunck A, Rauchhaus M, von Haehling S, Brunkhorst F, Cicoira M, Tschope C, Ponikowski P, Claus R, Anker S (2007) Secretory sphingomyelinase is upregulated in chronic heart failure: a second messenger system of immune activation relates to body composition, muscular functional capacity, and peripheral blood flow. Eur Heart J 28:821–828
Fuks Z, Kolesnick R (2005) Engaging the vascular component of the tumor response. Cancer Cell 8:89–91
Fuks Z, Alfieri A, Haimovitz-Friedman A, Seddon A, Cordon-Cardo C (1995) Intravenous basic fibroblast growth factor protects the lung but not mediastinal organs against radiation-induced apoptosis in vivo. Cancer J Sci Am 1:62–72
Garcia-Barros M, Paris F, Cordon-Cardo C, Lyden D, Rafii S, Haimovitz-Friedman A, Fuks Z, Kolesnick R (2003) Tumor response to radiotherapy regulated by endothelial cell apoptosis. Science 300:1155–1159
Garcia-Ruiz C, Colell A, Mari M, Morales A, Calvo M, Enrich C, Fernandez-Checa J (2003) Defective TNF-alpha-mediated hepatocellular apoptosis and liver damage in acidic sphingomyelinase knockout mice. J Clin Invest 111:197–208
Ginzburg L, Futerman A (2005) Defective calcium homeostasis in the cerebellum in a mouse model of Niemann-Pick A disease. J Neurochem 95:1619–1628
Goggel R, Winoto-Morbach S, Vielhaber G, Imai Y, Lindner K, Brade L, Brade H, Ehlers S, Slutsky A, Schutze S, Gulbins E, Uhlig S (2004) PAF-mediated pulmonary edema: a new role for acid sphingomyelinase and ceramide. Nat Med 10:155–160
Grassmé H, Becker KA (2013) Bacterial infections and ceramide. In: Gulbins E, Petrache I (eds) Sphingolipids in disease, vol 216, Handbook of experimental pharmacology. Springer, Heidelberg
Grassme H, Gulbins E, Brenner B, Ferlinz K, Sandhoff K, Harzer K, Lang F, Meyer T (1997) Acidic sphingomyelinase mediates entry of N. gonorrhoeae into nonphagocytic cells. Cell 91:605–615
Grassme H, Jekle A, Riehle A, Schwarz H, Berger J, Sandhoff K, Kolesnick R, Gulbins E (2001a) CD95 signaling via ceramide-rich membrane rafts. J Biol Chem 276:20589–20596
Grassme H, Schwarz H, Gulbins E (2001b) Molecular mechanisms of ceramide-mediated CD95 clustering. Biochem Biophys Res Commun 284:1016–1030
Grassme H, Cremesti A, Kolesnick R, Gulbins E (2003a) Ceramide-mediated clustering is required for CD95-DISC formation. Oncogene 22:5457–5470
Grassme H, Jendrossek V, Riehle A, von Kurthy G, Berger J, Schwarz H, Weller M, Kolesnick R, Gulbins E (2003b) Host defense against Pseudomonas aeruginosa requires ceramide-rich membrane rafts. Nat Med 9:322–330
Grassme H, Riehle A, Wilker B, Gulbins E (2005) Rhinoviruses infect human epithelial cells via ceramide-enriched membrane platforms. J Biol Chem 280:26256–26262
Grassme H, Riethmuller J, Gulbins E (2007) Biological aspects of ceramide-enriched membrane domains. Prog Lipid Res 46:161–170
Guicciardi M, Gores G (2005) Apoptosis: a mechanism of acute and chronic liver injury. Gut 54:1024–1033
Gulbins E (2003) Regulation of death receptor signaling and apoptosis by ceramide. Pharmacol Res 47:393–399
Gulbins E, Bissonnette R, Mahboubi A, Martin S, Nishioka W, Brunner T, Baier G, Baier-Bitterlich G, Byrd C, Lang F (1995) FAS-induced apoptosis is mediated via a ceramide-initiated RAS signaling pathway. Immunity 2:341–351
Haimovitz-Friedman A, Kan C, Ehleiter D, Persaud R, McLoughlin M, Fuks Z, Kolesnick R (1994) Ionizing radiation acts on cellular membranes to generate ceramide and initiate apoptosis. J Exp Med 180:525–535
Hannun Y, Obeid L (2002) The ceramide-centric universe of lipid-mediated cell regulation: stress encounters of the lipid kind. J Biol Chem 277:25847–25850
Hannun Y, Obeid L (2008) Principles of bioactive lipid signalling: lessons from sphingolipids. Nat Rev Mol Cell Biol 9:139–150
Hauck C, Grassme H, Bock J, Jendrossek V, Ferlinz K, Meyer T, Gulbins E (2000) Acid sphingomyelinase is involved in CEACAM receptor-mediated phagocytosis of Neisseria gonorrhoeae. FEBS Lett 478:260–266
He X, Miranda S, Xiong X, Dagan A, Gatt S, Schuchman E (1999) Characterization of human acid sphingomyelinase purified from the media of overexpressing Chinese hamster ovary cells. Biochim Biophys Acta 1432:251–264
Heinrich M, Wickel M, Schneider-Brachert W, Sandberg C, Gahr J, Schwandner R, Weber T, Saftig P, Peters C, Brunner J, Kronke M, Schutze S (1999) Cathepsin D targeted by acid sphingomyelinase-derived ceramide. EMBO J 18:5252–5263
Herr I, Martin-Villalba A, Kurz E, Roncaioli P, Schenkel J, Cifone M, Debatin K (1999) FK506 prevents stroke-induced generation of ceramide and apoptosis signaling. Brain Res 826:210–219
Herz J, Pardo J, Kashkar H, Schramm M, Kuzmenkina E, Bos E, Wiegmann K, Wallich R, Peters P, Herzig S, Schmelzer E, Kronke M, Simon MM, Utermohlen O (2009) Acid sphingomyelinase is a key regulator of cytotoxic granule secretion by primary T lymphocytes. Nat Immunol 10:761–768
Horinouchi K, Erlich S, Perl D, Ferlinz K, Bisgaier C, Sandhoff K, Desnick R, Stewart C, Schuchman E (1995) Acid sphingomyelinase deficient mice: a model of types A and B Niemann-Pick disease. Nat Genet 10:288–293
Huwiler A, Johansen B, Skarstad A, Pfeilschifter J (2001) Ceramide binds to the CaLB domain of cytosolic phospholipase A2 and facilitates its membrane docking and arachidonic acid release. FASEB J 15:7–9
Jarvis W, Kolesnick R, Fornari F, Traylor R, Gewirtz D, Grant S (1994) Induction of apoptotic DNA damage and cell death by activation of the sphingomyelin pathway. Proc Natl Acad Sci USA 91:73–77
Jin H, Schuchman E (2003) Ex vivo gene therapy using bone marrow-derived cells: combined effects of intracerebral and intravenous transplantation in a mouse model of Niemann-Pick disease. Mol Ther 8:876–885
Jin H, Carter J, Huntley G, Schuchman E (2002) Intracerebral transplantation of mesenchymal stem cells into acid sphingomyelinase-deficient mice delays the onset of neurological abnormalities and extends their life span. J Clin Invest 109:1183–1191
Kashiwagi K, Shirai Y, Kuriyama M, Sakai N, Saito N (2002) Importance of C1B domain for lipid messenger-induced targeting of protein kinase C. J Biol Chem 277:18037–18045
Kirschnek S, Paris F, Weller M, Grassme H, Ferlinz K, Riehle A, Fuks Z, Kolesnick R, Gulbins E (2000) CD95-mediated apoptosis in vivo involves acid sphingomyelinase. J Biol Chem 275:27316–27323
Kolesnick R (2002) The therapeutic potential of modulating the ceramide/sphingomyelin pathway. J Clin Invest 110:3–8
Kolesnick R, Fuks Z (2003) Radiation and ceramide-induced apoptosis. Oncogene 22:5897–5906
Kolesnick R, Hannun Y (1999) Ceramide and apoptosis. Trends Biochem Sci 24:224–225
Kolesnick R, Paley A (1987) 1,2-Diacylglycerols and phorbol esters stimulate phosphatidylcholine metabolism in GH3 pituitary cells. Evidence for separate mechanisms of action. J Biol Chem 262:9204–9210
Kolesnick R, Goni F, Alonso A (2000) Compartmentalization of ceramide signaling: physical foundations and biological effects. J Cell Physiol 184:285–300
Kubota M, Nakane M, Nakagomi T, Tamura A, Hisaki H, Ueta N, Inokuchi J, Hirayama A (2000) Sphingolipid biosynthesis by L-PDMP after rat MCA occlusion. Acta Neurochir Suppl 76:339–341
Lang P, Schenck M, Nicolay J, Becker J, Kempe D, Lupescu A, Koka S, Eisele K, Klarl B, Rubben H, Schmid K, Mann K, Hildenbrand S, Hefter H, Huber S, Wieder T, Erhardt A, Haussinger D, Gulbins E, Lang F (2007) Liver cell death and anemia in Wilson disease involve acid sphingomyelinase and ceramide. Nat Med 13:164–170
Leventhal A, Chen W, Tall A, Tabas I (2001) Acid sphingomyelinase-deficient macrophages have defective cholesterol trafficking and efflux. J Biol Chem 276:44976–44983
Li Y, Chen P, Haimovitz-Friedman A, Reilly R, Wong C (2003) Endothelial apoptosis initiates acute blood–brain barrier disruption after ionizing radiation. Cancer Res 63:5950–5956
Lin T, Genestier L, Pinkoski M, Castro A, Nicholas S, Mogil R, Paris F, Fuks Z, Schuchman E, Kolesnick R, Green D (2000) Role of acidic sphingomyelinase in Fas/CD95-mediated cell death. J Biol Chem 275:8657–8663
Liu P, Anderson RG (1995) Compartmentalized production of ceramide at the cell surface. J Biol Chem 270:27179–27185
Lozano J, Menendez S, Morales A, Ehleiter D, Liao W, Wagman R, Haimovitz-Friedman A, Fuks Z, Kolesnick R (2001) Cell autonomous apoptosis defects in acid sphingomyelinase knockout fibroblasts. J Biol Chem 276:442–448
Malagarie-Cazenave S, Segui B, Leveque S, Garcia V, Carpentier S, Altie M, Brouchet A, Gouaze V, Andrieu-Abadie N, Barreira Y, Benoist H, Levade T (2004) Role of FAN in tumor necrosis factor-alpha and lipopolysaccharide-induced interleukin-6 secretion and lethality in D-galactosamine-sensitized mice. J Biol Chem 279:18648–18655
Marathe S, Schissel S, Yellin M, Beatini N, Mintzer R, Williams K, Tabas I (1998) Human vascular endothelial cells are a rich and regulatable source of secretory sphingomyelinase. Implications for early atherogenesis and ceramide-mediated cell signaling. J Biol Chem 273:4081–4088
Mari M, Colell A, Morales A, Paneda C, Varela-Nieto I, Garcia-Ruiz C, Fernandez-Checa J (2004) Acidic sphingomyelinase downregulates the liver-specific methionine adenosyltransferase 1A, contributing to tumor necrosis factor-induced lethal hepatitis. J Clin Invest 113:895–904
McGovern M, Pohl-Worgall T, Deckelbaum R, Simpson W, Mendelson D, Desnick R, Schuchman E, Wasserstein M (2004) Lipid abnormalities in children with types A and B Niemann Pick disease. J Pediatr 145:77–81
Mendelson D, Wasserstein M, Desnick R, Glass R, Simpson W, Skloot G, Vanier M, Bembi B, Giugliani R, Mengel E, Cox G, McGovern M (2006) Type B Niemann-Pick disease: findings at chest radiography, thin-section CT, and pulmonary function testing. Radiology 238:339–345
Miranda S, Erlich S, Friedrich V, Haskins M, Gatt S, Schuchman E (1998) Biochemical, pathological, and clinical response to transplantation of normal bone marrow cells into acid sphingomyelinase-deficient mice. Transplantation 65:884–892
Miranda S, He X, Simonaro C, Gatt S, Dagan A, Desnick R, Schuchman E (2000) Infusion of recombinant human acid sphingomyelinase into niemann-pick disease mice leads to visceral, but not neurological, correction of the pathophysiology. FASEB J 14:1988–1995
Moeller B, Dreher M, Rabbani Z, Schroeder T, Cao Y, Li C, Dewhirst M (2005) Pleiotropic effects of HIF-1 blockade on tumor radiosensitivity. Cancer Cell 8:99–110
Morita Y, Perez G, Paris F, Miranda S, Ehleiter D, Haimovitz-Friedman A, Fuks Z, Xie Z, Reed J, Schuchman E, Kolesnick R, Tilly J (2000) Oocyte apoptosis is suppressed by disruption of the acid sphingomyelinase gene or by sphingosine-1-phosphate therapy. Nat Med 6:1109–1114
Nakane M, Kubota M, Nakagomi T, Tamura A, Hisaki H, Shimasaki H, Ueta N (2000) Lethal forebrain ischemia stimulates sphingomyelin hydrolysis and ceramide generation in the gerbil hippocampus. Neurosci Lett 296:89–92
Neuwelt E (2004) Mechanisms of disease: the blood–brain barrier. Neurosurgery 54:131–140
Ng C, Griffin D (2006) Acid sphingomyelinase deficiency increases susceptibility to fatal alphavirus encephalomyelitis. J Virol 80:10989–10999
Obeid L, Linardic C, Karolak L, Hannun Y (1993) Programmed cell death induced by ceramide. Science 259:1769–1771
Okazaki T, Bell R, Hannun Y (1989) Sphingomyelin turnover induced by vitamin D3 in HL-60 cells. Role in cell differentiation. J Biol Chem 264:19076–19080
Okazaki T, Bielawska A, Bell R, Hannun Y (1990) Role of ceramide as a lipid mediator of 1 alpha,25-dihydroxyvitamin D3-induced HL-60 cell differentiation. J Biol Chem 265:15823–15831
Osawa Y, Uchinami H, Bielawski J, Schwabe R, Hannun Y, Brenner D (2005) Roles for C16-ceramide and sphingosine 1-phosphate in regulating hepatocyte apoptosis in response to tumor necrosis factor-alpha. J Biol Chem 280:27879–27887
Otterbach B, Stoffel W (1995) Acid sphingomyelinase-deficient mice mimic the neurovisceral form of human lysosomal storage disease (Niemann-Pick disease). Cell 81:1053–1061
Paris F, Fuks Z, Kang A, Capodieci P, Juan G, Ehleiter D, Haimovitz-Friedman A, Cordon-Cardo C, Kolesnick R (2001) Endothelial apoptosis as the primary lesion initiating intestinal radiation damage in mice. Science 293:293–297
Paris F, Perez G, Fuks Z, Haimovitz-Friedman A, Nguyen H, Bose M, Ilagan A, Hunt P, Morgan W, Tilly J, Kolesnick R (2002) Sphingosine 1-phosphate preserves fertility in irradiated female mice without propagating genomic damage in offspring. Nat Med 8:901–902
Passini M, Macauley S, Huff M, Taksir T, Bu J, Wu I, Piepenhagen P, Dodge J, Shihabuddin L, O’Riordan C, Schuchman E, Stewart G (2005) AAV vector-mediated correction of brain pathology in a mouse model of Niemann-Pick A disease. Mol Ther 11:754–762
Passini M, Bu J, Fidler J, Ziegler R, Foley J, Dodge J, Yang W, Clarke J, Taksir T, Griffiths D, Zhao M, O’Riordan C, Schuchman E, Shihabuddin L, Cheng S (2007) Combination brain and systemic injections of AAV provide maximal functional and survival benefits in the Niemann-Pick mouse. Proc Natl Acad Sci USA 104:9505–9510
Pena L, Fuks Z, Kolesnick R (2000) Radiation-induced apoptosis of endothelial cells in the murine central nervous system: protection by fibroblast growth factor and sphingomyelinase deficiency. Cancer Res 60:321–327
Perez G, Knudson C, Leykin L, Korsmeyer S, Tilly J (1997) Apoptosis-associated signaling pathways are required for chemotherapy-mediated female germ cell destruction. Nat Med 3:1228–1232
Perez G, Jurisicova A, Matikainen T, Moriyama T, Kim M, Takai Y, Pru J, Kolesnick R, Tilly J (2005) A central role for ceramide in the age-related acceleration of apoptosis in the female germline. FASEB J 19:860–862
Piantadosi C, Schwartz D (2004) The acute respiratory distress syndrome. Ann Intern Med 141:460–470
Pier G (2002) CFTR mutations and host susceptibility to Pseudomonas aeruginosa lung infection. Curr Opin Microbiol 5:81–86
Riboni L, Campanella R, Bassi R, Villani R, Gaini S, Martinelli-Boneschi F, Viani P, Tettamanti G (2002) Ceramide levels are inversely associated with malignant progression of human glial tumors. Glia 39:105–113
Riethmuller J, Anthonysamy J, Serra E, Schwab M, Doring G, Gulbins E (2009) Therapeutic efficacy and safety of amitriptyline in patients with cystic fibrosis. Cell Physiol Biochem 24:65–72
Rotolo J, Zhang J, Donepudi M, Lee H, Fuks Z, Kolesnick R (2005) Caspase-dependent and -independent activation of acid sphingomyelinase signaling. J Biol Chem 280:26425–26434
Rotolo J, Stancevic B, Lu S, Zhang J, Suh D, King C, Kappel L, Murphy G, Liu C, Fuks Z, van den Brink M, Kolesnick R (2009) Cytolytic T cells induce ceramide-rich platforms in target cell membranes to initiate graft-versus-host disease. Blood 114:3693–3706
Rotolo J, Stancevic B, Zhang J, Hua G, Fuller J, Yin X, Haimovitz-Friedman A, Kim K, Qian M, Cardó-Vila M, Fuks Z, Pasqualini R, Arap W, Kolesnick R (2012) Anti-ceramide antibody prevents the radiation GI syndrome in mice. J Clin Invest 122:1786–1790
Rylova S, Somova O, Dyatlovitskaya E (1998) Comparative investigation of sphingoid bases and fatty acids in ceramides and sphingomyelins from human ovarian malignant tumors and normal ovary. Biochemistry 63:1057–1060
Santana P, Pena L, Haimovitz-Friedman A, Martin S, Green D, McLoughlin M, Cordon-Cardo C, Schuchman E, Fuks Z, Kolesnick R (1996) Acid sphingomyelinase-deficient human lymphoblasts and mice are defective in radiation-induced apoptosis. Cell 86:189–199
Schissel SL, Keesle GA, Schuchman EH, Williams KJ, Tabas I (1998a) The cellular trafficking and zinc dependence of secretory and lysosomal sphingomyelinase, two products of the acid sphingomyelinase gene. J Biol Chem 273:18250–18259
Schissel S, Jiang X, Tweedie-Hardman J, Jeong T, Camejo E, Najib J, Rapp J, Williams K, Tabas I (1998b) Secretory sphingomyelinase, a product of the acid sphingomyelinase gene, can hydrolyze atherogenic lipoproteins at neutral pH. Implications for atherosclerotic lesion development. J Biol Chem 273:2738–2746
Schuchman E (2010) Acid sphingomyelinase, cell membranes and human disease: lessons from Niemann-Pick disease. FEBS Lett 584:1895–1900
Seino K, Kayagaki N, Takeda K, Fukao K, Okumura K, Yagita H (1997) Contribution of Fas ligand to T cell-mediated hepatic injury in mice. Gastroenterology 113:1315–1322
Selzner M, Bielawska A, Morse M, Rudiger H, Sindram D, Hannun Y, Clavien P (2001) Induction of apoptotic cell death and prevention of tumor growth by ceramide analogues in metastatic human colon cancer. Cancer Res 61:1233–1240
Shimeno H, Soeda S, Yasukouchi M, Okamura N, Nagamatsu A (1995) Fatty acyl-Co A: sphingosine acyltransferase in bovine brain mitochondria: its solubilization and reconstitution onto the membrane lipid liposomes. Biol Pharm Bull 18:1335–1339
Sly W, Vogler C (2002) Brain-directed gene therapy for lysosomal storage disease: going well beyond the blood-brain barrier. Proc Natl Acad Sci USA 99:5760–5762
Small D (1970) Surface and bulk interactions of lipids and water with a classification of biologically active lipids based on these interactions. Fed Proc 29:1320–1326
Smith E, Schuchman E (2008) The unexpected role of acid sphingomyelinase in cell death and the pathophysiology of common diseases. FASEB J 22:3419–3431
Song D, Kavanagh B, Benedict S, Schefter T (2004) Stereotactic body radiation therapy. Rationale, techniques, applications, and optimization. Oncology 18:1419–1430
Stancevic B, Kolesnick R (2010) Ceramide-rich platforms in transmembrane signaling. FEBS Lett 584:1728–1740
Steinhart W, Busch J, Oettgen J, Howland J (1984) Sphingolipid metabolism during infection of human fibroblasts by herpes simplex virus type 1. Intervirology 21:70–76
Teichgraber V, Ulrich M, Endlich N, Riethmuller J, Wilker B, De Oliveira-Munding C, van Heeckeren A, Barr M, von Kurthy G, Schmid K, Weller M, Tummler B, Lang F, Grassme H, Doring G, Gulbins E (2008) Ceramide accumulation mediates inflammation, cell death and infection susceptibility in cystic fibrosis. Nat Med 14:382–391
Truman J, Garcia-Barros M, Kaag M, Hambardzumyan D, Stancevic B, Chan M, Fuks Z, Kolesnick R, Haimovitz-Friedman A (2010) Endothelial membrane remodeling is obligate for anti-angiogenic radiosensitization during tumor radiosurgery. PLoS One 5(8):e12310
Tuder R, Petrache I, Elias J, Voelkel N, Henson P (2003) Apoptosis and emphysema: the missing link. Am J Respir Cell Mol Biol 28:551–554
Utermohlen O, Karow U, Lohler J, Kronke M (2003) Severe impairment in early host defense against Listeria monocytogenes in mice deficient in acid sphingomyelinase. J Immunol 170:2621–2628
van Vulpen M, Kal H, Taphoorn M, El-Sharouni S (2002) Changes in blood–brain barrier permeability induced by radiotherapy: implications for timing of chemotherapy? (Review). Oncol Rep 9:683–688
Verheij M, Bose R, Lin X, Yao B, Jarvis W, Grant S, Birrer M, Szabo E, Zon L, Kyriakis J, Haimovitz-Friedman A, Fuks Z, Kolesnick R (1996) Requirement for ceramide-initiated SAPK/JNK signalling in stress-induced apoptosis. Nature 380:75–79
Wang Z, Dudhane A, Orlikowsky T, Clarke K, Li X, Darzynkiewicz Z, Hoffmann M (1994) CD4 engagement induces Fas antigen-dependent apoptosis of T cells in vivo. Eur J Immunol 24:1549–1552
Waselenko J, MacVittie T, Blakely W, Pesik N, Wiley A, Dickerson W, Tsu H, Confer D, Coleman C, Seed T, Lowry P, Armitage J, Dainiak N (2004) Medical management of the acute radiation syndrome: recommendations of the Strategic National Stockpile Radiation Working Group. Ann Intern Med 140:1037–1051
Yang W, Dodge J, Passini M, Taksir T, Griffiths D, Schuchman E, Cheng S, Shihabuddin L (2007) Intraparenchymal injections of acid sphingomyelinase results in regional correction of lysosomal storage pathology in the Niemann-Pick A mouse. Exp Neurol 207:258–266
Yu Z, Nikolova-Karakashian M, Zhou D, Cheng G, Schuchman E, Mattson M (2000) Pivotal role for acidic sphingomyelinase in cerebral ischemia-induced ceramide and cytokine production, and neuronal apoptosis. J Mol Neurosci 15:85–97
Zelinski M, Murphy M, Lawson M, Jurisicova A, Pau K, Toscano N, Jacob D, Fanton J, Casper R, Dertinger S, Tilly J (2011) In vivo delivery of FTY720 prevents radiation-induced ovarian failure and infertility in adult female nonhuman primates. Fertil Steril 95(1440–1445):e1441–e1447
Zundel W, Giaccia A (1998) Inhibition of the anti-apoptotic PI(3)K/Akt/Bad pathway by stress. Genes Dev 12:1941–1946
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer-Verlag Wien
About this chapter
Cite this chapter
Hua, G., Kolesnick, R. (2013). Using ASMase Knockout Mice to Model Human Diseases. In: Gulbins, E., Petrache, I. (eds) Sphingolipids in Disease. Handbook of Experimental Pharmacology, vol 216. Springer, Vienna. https://doi.org/10.1007/978-3-7091-1511-4_2
Download citation
DOI: https://doi.org/10.1007/978-3-7091-1511-4_2
Published:
Publisher Name: Springer, Vienna
Print ISBN: 978-3-7091-1510-7
Online ISBN: 978-3-7091-1511-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)